Introduction
Ischemic stroke is the most common type of
cerebrovascular disease, accounting for 85% of all cerebrovascular
diseases. It is ranked third among life-threatening diseases and is
the most disabling disease worldwide (1). It severely affects human health and,
within 3 months following a stroke, ~15–30% of survivors suffer
from permanent disability, including paralysis, memory disorders,
thought disorders, linguistic problems and akinesia (2). Of these individuals, ~20% require
care as they cannot perform self-care. In western countries,
>70% of the population >65 years old experience a stroke
(3), and the number of patients
continues to increase (3).
Although the pathology and physiology of ischemic stroke involve
different mechanisms, increasing evidence has indicated that
ischemic damage and inflammation lead to disease progression
(4). Cerebral ischemia induces
pathological pathways in an ischemia cascade reaction and causes
irreversible damage of ischemic core neurons (5).
At present, substantial evidence has shown that the
essential mechanism of cerebral ischemic injury involves oxidative
stress and inflammatory reaction (6). The use of psychotherapeutic drugs for
anti-inflammatory treatment has provided scope clinically,
including for the treatment of tumors (7), and it has shown preliminary treatment
effects. Consequently, theoretical and clinical investigations
aimed at enhancing the inflammatory mechanism following cerebral
ischemia is likely to generate novel opportunities for the
treatment of cerebral infarction (6,8).
Toll-like receptor 4 (TLR4) is a transmembrane
receptor protein composed of an extracellular region, transmembrane
region and intracellular region (9). Through recognition of high-mobility
group protein 1 (HMGB1), heat shock protein, fibrous protein,
necrocytosis components and other molecules released at sites of
histocyte damage, TLR4 further activates the signal transduction
pathway in the cell to enhance the synthesis and release of
cytokines, promoting the maturation and differentiation of immune
cells and regulating the immune response (9). In peripheral regions, TLR4 is
predominantly distributed on lymphocytes, macrophages, dendritic
cells and other innate immunocytes, and is involved in immune
responses to bacteria and other exogenous pathogens. In the central
nervous system, TLR4 distributes extensively on astrocytes,
microglial cells, vascular endothelium and smooth muscle
cytomembranes, and is involved in the pathogenic process of
Parkinson's disease, Alzheimer's disease and other
neurodegenerative diseases (10).
The activated TLR4 can activate nuclear factor-κB (NF-κB) in a
resting state, which translocates to the cell nucleus to initiate
the gene expression of TNF-(4 interleukin (IL)-1, cyclooxygenase-2
(COX-2) and other adhesion molecules through combining with myeloid
differentiation antigen 88 (MyD88), leading to inflammatory
reactions (11).
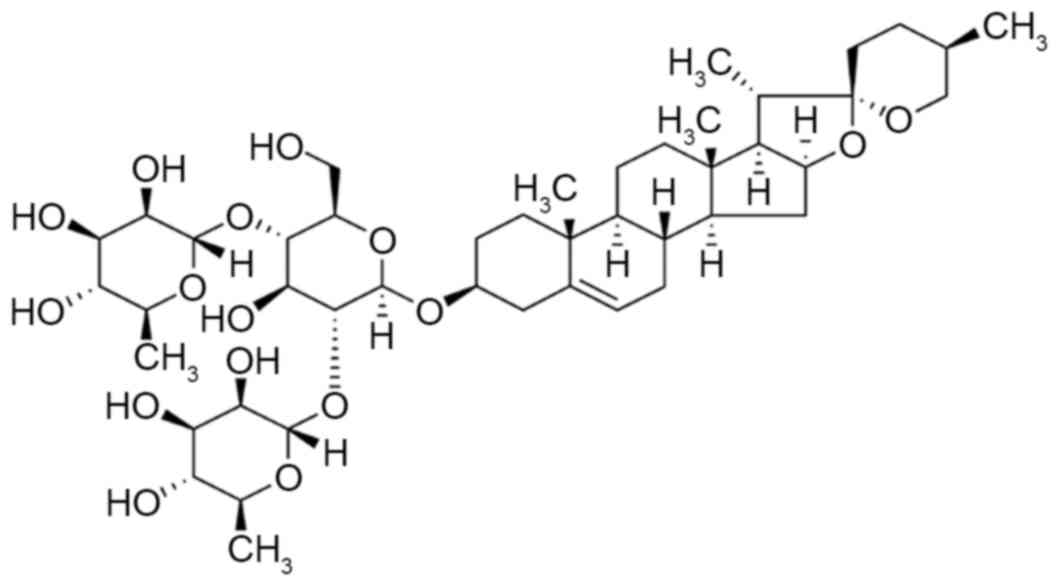
Diosgenin, generally known as saponin, is a natural
and synthetic steroid sapogenin belonging to a screw sterane
alcohol glucoside (Fig. 1)
(12). The relative molecular mass
is 414.63, and is present extensively in leguminosae and
dioscoreaceae plants. Diosgenin is found in the seeds of fenugreek
Trigonella foenum-graecum L. (13). It can also be extracted from the
tuber of Dioscorea zingiberensis C. H. Wright, D.
nipponica Makino, D. panthaica Prain et Burkill and
D. nipponica Makino ssp. Rosthornii (Prain et Burkill) C. T.
Ting through methods, including hydrolyzation, fermentation and
extraction (14). Diosgenin, as an
essential raw material for the synthesis of steroid hormone drugs
and steroidal contraceptives, is used to produce pregnenolone,
progesterone and cortisone (15).
Previous in-depth investigations of the pharmacologic actions of
diosgenin have been performed. Diosgenin has antitumor effects in
addition to blood lipid-regulating, anti-platelet aggregating and
bilifaction-promoting effects (14). It is an essential drug for the
treatment of cardiovascular disease, encephalitis, dermatosis and
tumors (15,16). Therefore, the aim of the present
study was to investigate the neuroprotective effect of dioscin on
inhibiting the effects of ischemic stroke and its possible
mechanisms.
Materials and methods
Animals
Adult male Sprague-Dawley rats (8–10 weeks old,
200–230 g) were purchased from the Hunan Experimental Animal Centre
of Hunan University of Chinese Medicine (Hunan, China) and fed a
commercially available liquid diet. The rats were housed in
separate cages in a room with controlled temperature (22–24°C), 12
h light:dark cycle (light between 8:00 and 20:00) and humidity
(50–55%). The protocols used in the present study followed the
National Institutes of Health guidelines for the Care and Use of
Laboratory Animals (National Institutes of Health, Bethesda, MD,
USA), were approved by the institutional animal ethics committee of
Hunan University of Chinese Medicine of Medicine. Following 1 week
of acclimatization, the rats were randomly divided into three
groups (n=8 per group) as follows: Sham group, stroke model group,
and dioscin treatment group. In the dioscin treatment group, the
rats were intragastrically treated with 80 mg/kg/day of dioscin for
4 weeks.
Middle cerebral artery occlusion
(MCAO)
All rats were anesthetized with 2.5% isoflorane
(Forane; Abbot Japan, Tokyo, Japan) and were maintained at 37±1°C
during the experiment. The right common carotid artery was exposed
using a midline neck incision, and 4-cm poly-L-lysine-coated nylon
thread (3–0) was inserted into the internal carotid artery through
the right common carotid artery and gently advanced until
resistance was detected in the blood flow trace. MCAO was
maintained for 2 h, following which the thread was gently removed
to restore blood flow. The wounds were sterilized and sutured, and
the rats were allowed to recover from anesthesia.
Neurological assessment
The modified neurological severity score was used to
assess neurologic severity scores. The neurologic function was
graded with scores between 0 and 5 (0, no neurologic deficit; 5,
maximal deficit). A score of 1 indicated that the rats were unable
perform the test or the tested reflex, and a higher score indicated
more severe injury.
Following anesthesia with 2.5% isoflorane, the rats
were sacrificed and their brains were removed, cleaned and
solidified using pre-cooled normal saline (4°C) for 5 min. The
prepared slices (5 µM) were stained with 2%
2,3,5-triphenyltetrazolium chloride (Sigma; Merck Millipore;
Darmstadt, Germany) and fixed in 10% buffered formalin solution.
Images were captured using light microscopy (Nikon Eclipse
TE2000-U; Nikon, Tokyo, Japan).
Determination of biological
indicators
Following anesthesia with 2.5% isoflorane, the rats
were sacrificed, following which their brains were removed and
cleaned, and the hippocampus was separated. Total cellular protein
was lysed from the hippocampal tissue using
radioimmunoprecipitation lysis buffer (Beyotime Institute of
Biotechnology, Jiangsu, China) and the protein content was
determined using a BCA protein assay kit (Beyotime Institute of
Biotechnology). The proteins (2–5 inate of Biotechnologyionthe
activities of IL-1 activities of Biotechusing ELISA kits. The color
intensity was measured at 450 nm. The proteins (10 µot were
incubated with the caspase-3 and caspase-9 activity kits
(Ac-DEVD-pNA and Ac-LEHD-pNA) for 2 h at 37°C. The color intensity
was measured at 405 nm.
Western blot analysis
Total cellular protein was lysed from the
hippocampal tissue using radioimmunoprecipitation lysis buffer
(Beyotime Institute of Biotechnology) and the protein content was
determined using a BCA protein assay kit (Beyotime Institute of
Biotechnology). The proteins (50 µg) were subjected to 8–12%
SDS-PAGE and then transferred onto a nitrocellulose membrane (EMD
Millipore, Billerica, MA, USA). The membrane was blocked by
incubation with 5% (w/v) nonfat milk in Tris-buffered saline with
Tween-20 (Sigma-Aldrich; Merck Millipore). The membrane was then
incubated with anit-IRAK1 (cat. no. sc-7883, 1:500, Santa Cruz
Biotechnology, Inc., Dallas, TX, USA), anti-TRAF6 (cat. no.
sc-7221, 1:500, Santa Cruz Biotechnology, Inc.), anti-HMGB-1 (cat.
no. sc-135809, 1:500, Santa Cruz Biotechnology, Inc.), anti-TLR4
(cat. no. sc-293072, 1:500, Santa Cruz Biotechnology, Inc.),
anti-MyD88 (cat. no. sc-11356, 1:500, Santa Cruz Biotechnology,
Inc.), anti-NF-κB (cat. no. sc-7178, 1:500, Santa Cruz
Biotechnology, Inc.) and anti-GAPDH (cat. no. AG019, 1:2,000,
Beyotime Institute of Biotechnology) antibodies overnight at 4°C.
The blots were then incubated with anti-rabbit horseradish
peroxidase-conjugated antibodies (cat. no. A0208, 1:5,000, Beyotime
Institute of Biotechnology) for 1 h at room temperature. The
protein blank was detected using enhanced chemiluminescence
(Beyotime Institute of Biotechnology) method and images were
captured using the Image Lab (version 3.0; Bio-Rad Laboratories,
Inc., Hercules, CA, USA).
Statistical analysis
All data are expressed as the mean ± standard
deviation and analysed using SPSS software (version 19.0; IBM
Corp., Armonk, NY, USA). One-way analysis of variance was performed
to compare the differences between two groups, followed by Duncan's
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Effect of dioscin on infarct volume
and neurological scores in rats with ischemic stroke
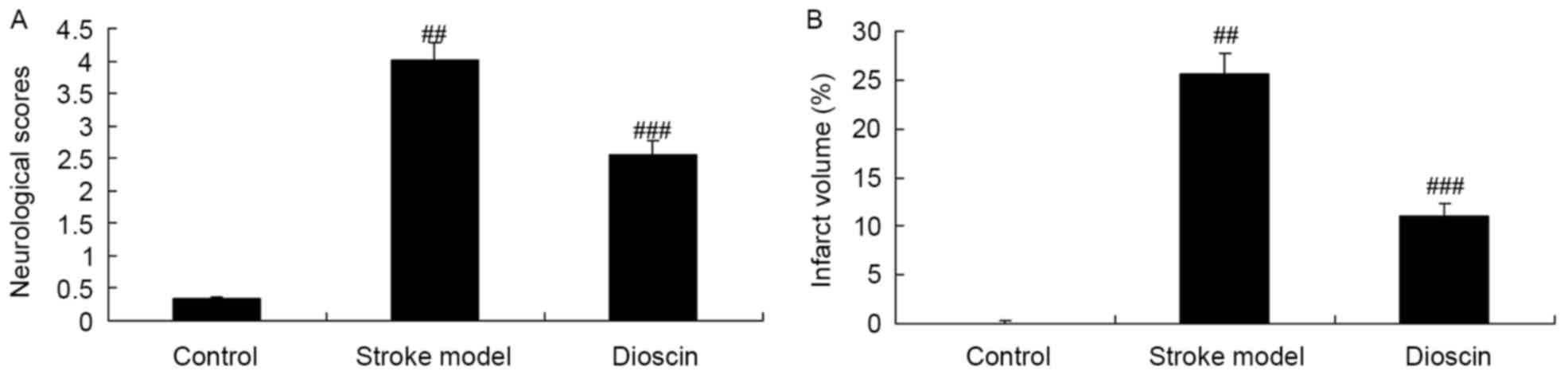
In order to confirm the effect of dioscin on
ischemic stroke, the infarct volume and neurological scores were
determined. As shown in Fig. 2A,
there were significant increases in infarct volume and neurological
scores in the ischemic stroke group, compared with the sham control
group. By contrast, treatment with dioscin significantly reduced
the ischemic stroke-induced infarct volume and neurological scores
in the ischemic stroke model (Fig.
2B).
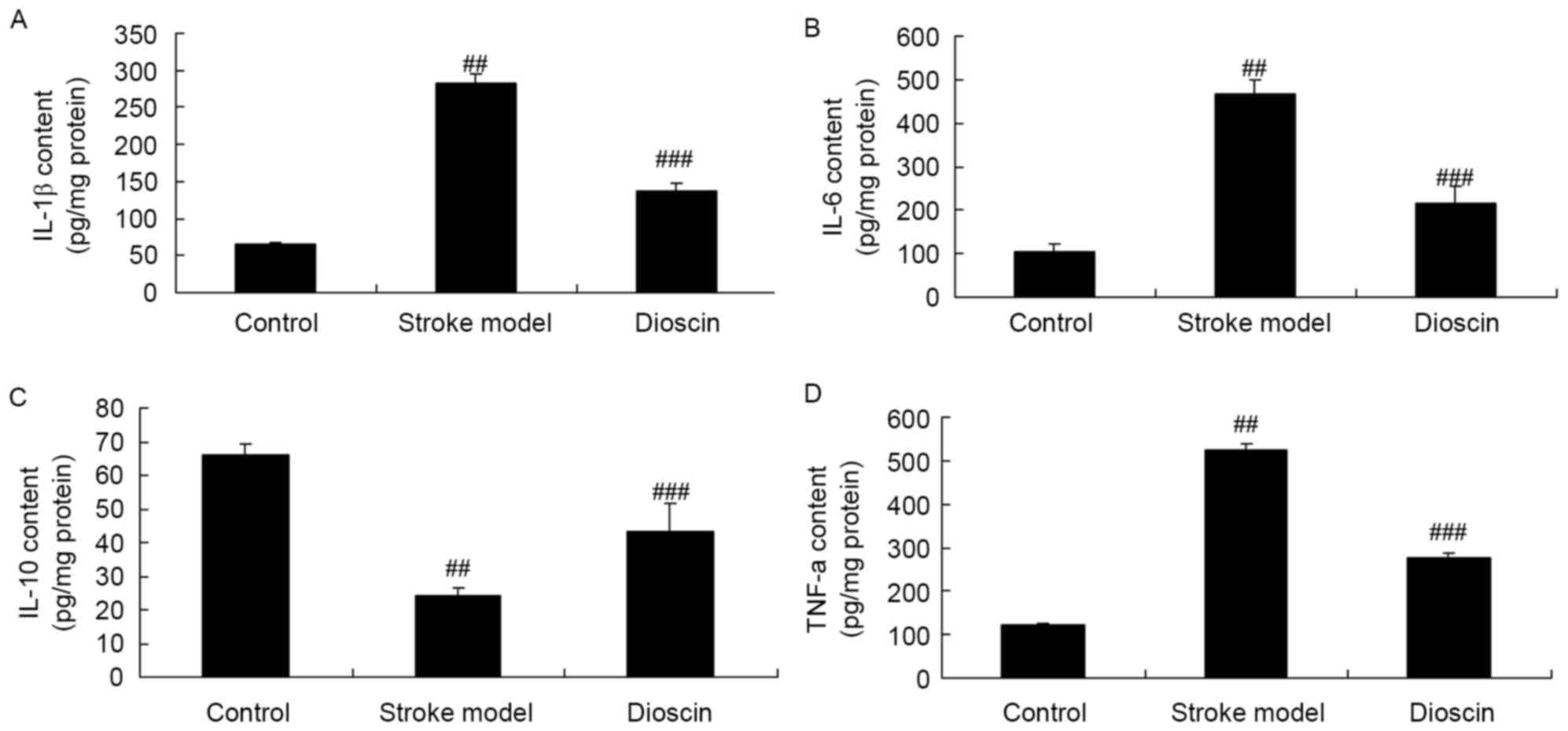
Effect of dioscin on inflammatory
responses in the rat ischemic stroke model
To further confirm the inhibitory effect of dioscin
in the ischemic stroke rats, the activities of IL-1e activities
ofts, e respowere measured using ELISA kits. As shown in Fig. 3A-D, the results indicated that
there were increases in the activities of IL-1β, IL-6 and TNF-α and
a decreased in the activity of IL-10 in the rats of the ischemic
stroke model, compared with those in the control group. In
addition, dioscin treatment significantly inhibited the activities
of IL-1activities of igniin the rat ischemic stroke model (Fig. 3).
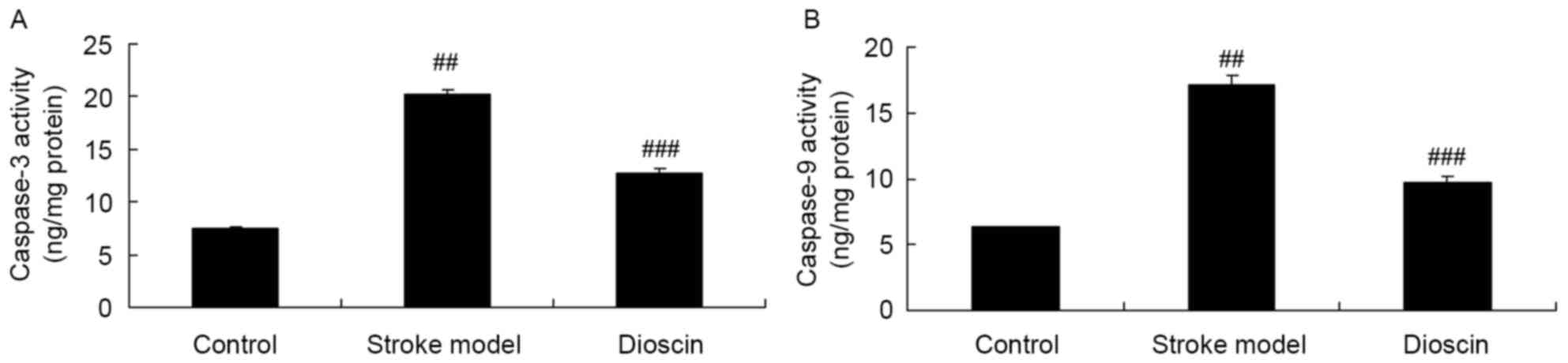
Effect of dioscin on activities of
caspase-3/9 in the rat ischemic stroke model
To further confirm the involvement of caspase
activation in the effect of dioscin on apoptosis in rats with
ischemic stroke, caspase-3 and caspase-9 kits were used to measure
these indices. The activities of caspase-3 and caspase-9 in the
rats of the ischemic stroke model were increased, compared with
those in the sham control group (Fig.
4A and B). Treatment with dioscin significantly inhibited the
activities of caspase-3 and caspase-9 activity in the ischemic
stroke model (Fig. 4A and B).
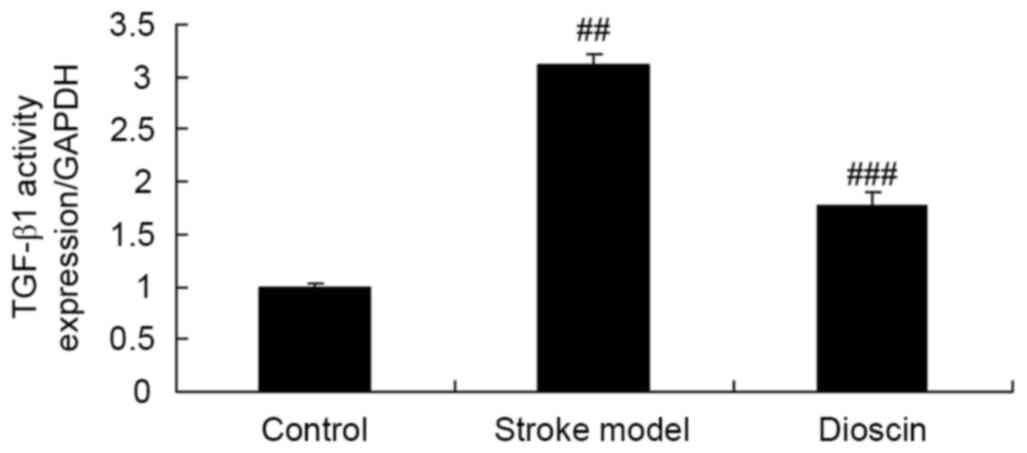
Effect of dioscin on the activity of
TGF-β1 in the rat ischemic stroke model
To determine whether the effect of dioscin affects
the activity of TGF-acin the rats in the ischemic stroke model, the
activities of TGF-c swere measured using ELISA kits. Compared with
that in the sham control group, the activity of TGF-actin the
ischemic stroke group was enhanced (Fig. 5). Treatment with dioscin
significantly suppressed the activity of TGF-β1 in the ischemic
stroke model (Fig. 5).
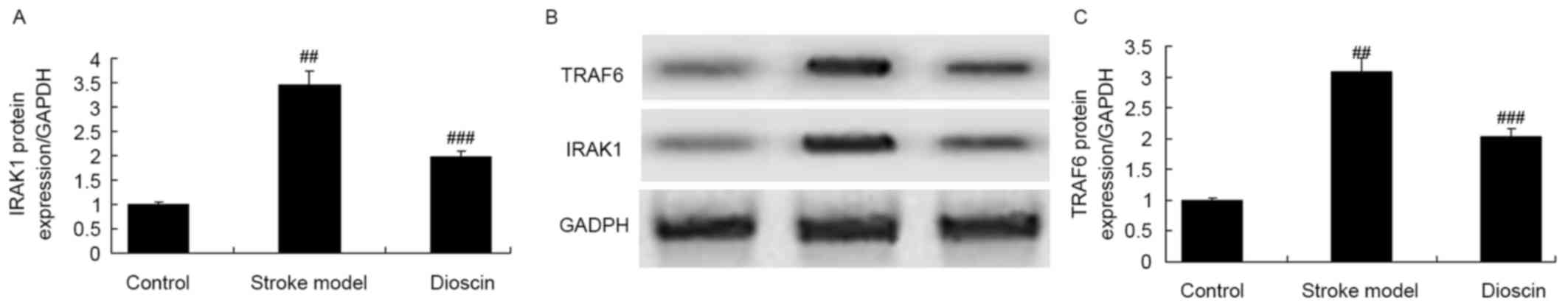
Effect of dioscin on the protein
expression levels of IRAK1 and TRAF6 in the rat ischemic stroke
model
The present study examined the protein expression
levels of IRAK1 and TRAF6 in the rats of the ischemic stroke model
treated with dioscin. As shown in Fig.
6A-C, the protein expression levels of IRAK1 and TRAF6 in the
ischemic stroke group were higher, compared with those of the sham
control group. Dioscin markedly reduced the protein expression
levels of IRAK1 and TRAF6 in the ischemic stroke model (Fig. 6A-C).
Effect of dioscin on the protein
expression levels of HMGB-1 in the rat ischemic stroke model
The present study assessed whether dioscin affected
the protein expression levels of HMGB-1 in the rats ischemic stroke
model. As shown in Fig. 7A and B,
the protein expression of HMGB-1 in the ischemic stroke group was
increased, compared with that in the sham group. The induced
protein expression of HMGB-1 in the ischemic stroke group was
significantly suppressed by dioscin (Fig. 7A and B).
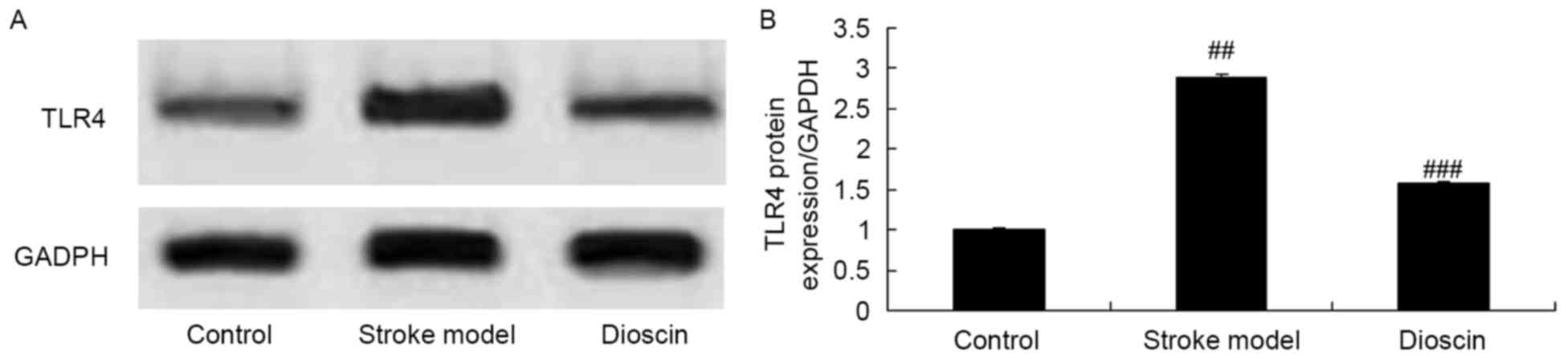
Effect of dioscin on the protein
expression levels of TLR4 in the rat ischemic stroke model
In order to examine the effects of the
administration of dioscin on stroke, western blot analysis was
performed to measure the protein expression levels of TLR4 in the
ischemic stroke group. As shown in Fig. 8A and B, the protein expression
level of TLR4 in the ischemic stroke group was markedly increased,
compared with that on the sham control group. However, treatment
with dioscin significantly suppressed the protein expression of
TLR4 in the ischemic stroke group (Fig. 8A and B).
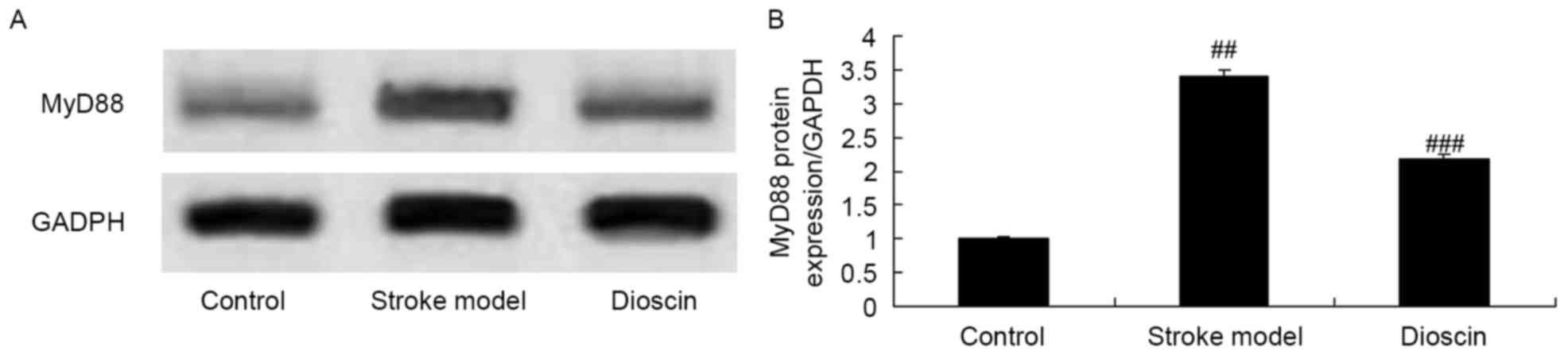
Effect of dioscin on the protein
expression of MyD88 in the rats ischemic stroke model
The present study also examined the protein
expression levels of MyD88 in rats with ischemic stroke treated
with dioscin using western blotting analysis. As shown in Fig. 9A and B, the protein expression of
MyD88 in ischemic stroke group was higher, compared with that in
the sham control group. Dioscin treatment significantly suppressed
the protein expression of MyD88 in the ischemic stroke group
(Fig. 9A and B).
Effect of dioscin on the protein
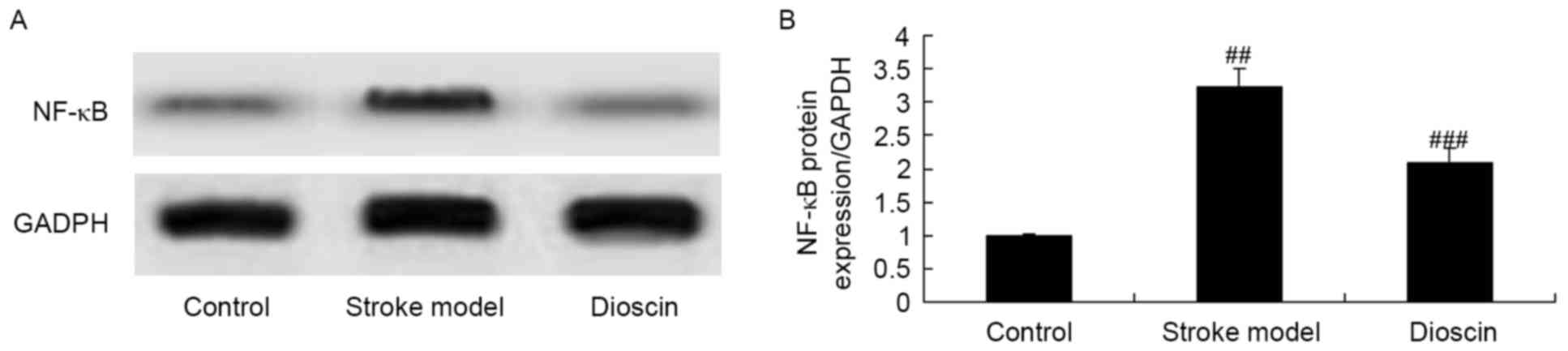
expression of NF-κB in the rat ischemic stroke model
To investigate the possible role of NF-κB
invpathways in the effects of dioscin on ischemic stroke, the
protein expression levels of NF-κB were examined using western blot
analysis. The results of the western blot analysis showed that the
protein expression of NF-κB was significantly increased in the
ischemic stroke group, compared with that in the sham control
group. By contrast, dioscin treatment significantly suppressed the
protein expression of NF-κB in the ischemic stroke group (Fig. 10A and B).
Discussion
Acute cerebral infarction is also known as apoplexy
or cerebral apoplexy. It is one of the most complicated nervous
system diseases with the highest level of damage. It has been
ranked as the third leading cause of mortality and the leading
contributor to disability (17).
Following thrombosis, sudden interruption or cerebral blood flow or
emboli, patients suffer from symptoms, including paralysis,
language damage and eyesight loss (18). In addition, <15% of cases are
caused by hemorrhagic stroke or cardiac arrest. According to
statistical data of the United States in 2014, the average rate of
stroke occurrence was once every 40 sec, with associated mortality
every 4 min (4). The estimated
mortality rate has reached 41.6% and, with an increasingly aging
population the rates are likely to increase correspondingly. The
data obtained in the present study suggested that treatment with
dioscin significantly reduced ischemic stroke-induced infarct
volume and neurological scores in the ischemic stroke model.
The pathological and physiological mechanisms of
cerebral ischemia/reperfusion injury include energy failure,
excitability, amino acid toxicity, oxidative stress damage, nitric
oxide, inflammatory reaction, cell apoptosis, mitochondria stress
and autophagy. On reviewing the time-scale of cerebral
ischemia/reperfusion injury, inflammatory reactions become apparent
up to 2 h following ischemia, and further aggravate cerebral tissue
damage (19). As a result of
hematoencephalic barrier effects, the inflammatory reactions
following ischemia are divided into central sleep apnea and
peripheral (20). Initially, at
the early stage of cerebral ischemia/reperfusion injury, free
radicals, cell toxicity and other substances released by the
‘infarction core’ activate the microglia inherent inflammatory
cells in the central nervous system, and induce microglial cells to
release TNF-roglia inherent inflammatory cells in the central
nervous sfollowing ischemia (21).
Tao et al confirmed that dioscin ameliorates inflammation
through TLR4 signaling via the inhibition of HMGB-1 in an in
vitro cerebral ischemia/reperfusion injury model (22).
These results showed that Dioscin treatment
significantly inhibited the activities of IL-1β, IL-6, TNF-α,
caspase-3 and caspase-9 in the ischemic stroke rats, which showed
that the dioscin-induced inhibition of ischemic stroke was
dependent on the inflammation and anti-apoptotic effects.
TLRs are one of the most important pattern
recognition receptor families, and have key effects on the
induction of inflammatory reactions and generation of the
inflammatory medium process (23).
All TLRs are enriched in the extracellular region of leucine-rich
repeat sequences, and is necessary for recognizing
pathogen-associated molecular patterns (PAMPs) (24). In addition, the intracellular Tol
I-interleukin-1 receptor structural domain is required for
initiating intracellular signal transduction. The aforementioned
structure can recognize conserved molecular structures and
microorganism products, including the endogenous molecular patterns
of PAMPs and tissue damage (25).
A previous study have shown that the expression of TLR4 is
upregulated in an ischemic stroke model (25), expressed predominantly on
microglial cells and neurons. In addition, in a mouse model with
TLR4 defect, nerve function loss and brain damage were less marked,
compared with those in wild-type mice (26). TLR activation leads to the
translocation of NF-κB. It is a key regulatory factor mediating the
expression of inflammatory mediators and the inflammatory reaction.
Inhibiting its activity reduces brain damage (27). The results of the present study
showed that dioscin significantly suppressed the activity of
TGF-β1, reduced the protein expression levels of IRAK1 and TRAF6,
and suppressed the protein expression levels of HMGB-1 and TLR4 in
the rats of the ischemic stroke model. Liu et al showed that
dioscin alleviates alcoholic liver fibrosis through the
TLR4/MyD88/NF-κB signaling pathway (13). Therefore, the effect of dioscin on
ischemic stroke may be associated with the TLR4 pathway.
The TLR4/NF-kB signal transduction pathway activates
downstream inflammatory cytokines, including TNF-a, IL-6, IL-10,
C-X-C motif chemokine ligand-10, interferon-F and chemokines, and
upregulates the expression of cell adhesion molecules. In addition,
the expression of matrix metalloproteinase (MMP)-9 in ischemic
brain tissue increases with pro-inflammatory effects (11,28).
The cytokines generated following ischemia can induce the
generation of MMP, which causes an increase in vasopermeability and
blood-brain barrier (BBB) damage. This further promotes
microvascular basement membrane protein hydrolysis and vasogenic
cerebral edema generation, and leads to increased cerebral
ischemia, BBB destruction, encephaledema, neuronal cell death, and
further aggravation of brain damage (10). Qi et al found that dioscin
alleviates LPS-induced inflammatory kidney injury via the
TLR4/MyD88 signaling pathway (12). The results of the present study
showed that dioscin treatment significantly also suppressed the
protein expression levels of MyD88 and NF-88 in the rats of the
ischemic stroke model, and downregulated the TLR4/MyD88/NF-4/
pathway.
In conclusion, based on the observations of the
present study, dioscin significantly reduced ischemic
stroke-induced infarct volume and neurological scores, and
inhibited ischemic stroke-induced inflammation and expression
levels of TGF-ession levels ofced inflammain the ischemic stroke
model. These findings provide novel insights into the mechanisms of
dioscin as a potent anti-inflammatory agent, which may be used to
treat ischemic stroke via the TLR4/MyD88/NF-reat ischemi Therefore,
dioscin may have anti-inflammatory, anti-apoptotic and other
effects, which require confirmation in the future, in addition to
clinical application to provide further data to support the
findings obtained in the present study.
References
|
1
|
Lin YN, Hu CJ, Chi JY, Lin LF, Yen TH, Lin
YK and Liou TH: Effects of repetitive transcranial magnetic
stimulation of the unaffected hemisphere leg motor area in patients
with subacute stroke and substantial leg impairment: A pilot study.
J Rehabil Med. 47:305–310. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Verdecchia P, Reboldi G, Angeli F,
Trimarco B, Mancia G, Pogue J, Gao P, Sleight P, Teo K and Yusuf S:
Systolic and diastolic blood pressure changes in relation with
myocardial infarction and stroke in patients with coronary artery
disease. Hypertension. 65:108–114. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jusufovic M, Sandset EC, Bath PM, Karlson
BW and Berge E: Scandinavian Candesartan Acute Stroke Trial Study
Group: Effects of blood pressure lowering in patients with acute
ischemic stroke and carotid artery stenosis. Int J Stroke.
10:354–359. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Muchada M, Rubiera M, Rodriguez-Luna D,
Pagola J, Flores A, Kallas J, Sanjuan E, Meler P, Alvarez-Sabin J,
Ribo M and Molina CA: Baseline National Institutes of Health stroke
scale-adjusted time window for intravenous tissue-type plasminogen
activator in acute ischemic stroke. Stroke. 45:1059–1063. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lindman BR, Zajarias A, Madrazo JA, Shah
J, Gage BF, Novak E, Johnson SN, Chakinala MM, Hohn TA, Saghir M
and Mann DL: Effects of phosphodiesterase type 5 inhibition on
systemic and pulmonary hemodynamics and ventricular function in
patients with severe symptomatic aortic stenosis. Circulation.
125:2353–2362. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xin Q, Cheng B, Pan Y, Liu H, Yang C, Chen
J and Bai B: Neuroprotective effects of apelin-13 on experimental
ischemic stroke through suppression of inflammation. Peptides.
63:55–62. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Boehme AK, McClure LA, Zhang Y, Luna JM,
Del Brutto OH, Benavente OR and Elkind MS: Inflammatory markers and
outcomes after lacunar stroke: Levels of inflammatory markers in
treatment of stroke study. Stroke. 47:659–667. 2016.PubMed/NCBI
|
|
8
|
Folyovich A, Biró E, Orbán C, Bajnok A,
Varga V, Béres-Molnár AK, Vásárhelyi B and Toldi G: Relevance of
novel inflammatory markers in stroke-induced immunosuppression. BMC
Neurol. 14:412014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang P, Guo ZF, Xu YM, Li YS and Song JG:
N-Butylphthalide (NBP) ameliorated cerebral ischemia
reperfusion-induced brain injury via HGF-regulated TLR4/NF-κB
signaling pathway. Biomed Pharmacother. 83:658–666. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang B, Choi JJ, Eum SY, Daunert S and
Toborek M: TLR4 signaling is involved in brain vascular toxicity of
PCB153 bound to nanoparticles. PLoS One. 8:e631592013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ye L, Yang Y, Zhang X, Cai P, Li R, Chen
D, Wei X, Zhang X, Xu H, Xiao J, et al: The role of bFGF in the
excessive activation of astrocytes is related to the inhibition of
TLR4/NFκB signals. Int J Mol Sci. 17:pii: E37. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Qi M, Yin L, Xu L, Tao X, Qi Y, Han X,
Wang C, Xu Y, Sun H, Liu K and Peng J: Dioscin alleviates
lipopolysaccharide-induced inflammatory kidney injury via the
microRNA let-7i/TLR4/MyD88 signaling pathway. Pharmacol Res.
111:509–522. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu M, Xu Y, Han X, Yin L, Xu L, Qi Y,
Zhao Y, Liu K and Peng J: Dioscin alleviates alcoholic liver
fibrosis by attenuating hepatic stellate cell activation via the
TLR4/MyD88/NF-κB signaling pathway. Sci Rep. 5:180382015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Guo Y, Xing E, Song H, Feng G, Liang X, An
G, Zhao X and Wang M: Therapeutic effect of dioscin on
collagen-induced arthritis through reduction of Th1/Th2. Int
Immunopharmacol. 39:79–83. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lee HJ, Park JS, Yoon YP, Shin YJ, Lee SK,
Kim YS, Hong JH, Son KH and Lee CJ: Dioscin and methylprotodioscin
isolated from the root of Asparagus cochinchinensis suppressed the
gene expression and production of airway MUC5AC mucin induced by
phorbol ester and growth factor. Phytomedicine. 22:568–572. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhao X, Xu L, Zheng L, Yin L, Qi Y, Han X,
Xu Y and Peng J: Potent effects of dioscin against gastric cancer
in vitro and in vivo. Phytomedicine. 23:274–282. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cramer SC and Hill MD: REGENESIS-LED
Investigators: Human choriogonadotropin and epoetin alfa in acute
ischemic stroke patients (REGENESIS-LED trial). Int J Stroke.
9:321–327. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Punt M, van Alphen B, van de Port IG, van
Dieën JH, Michael K, Outermans J and Wittink H: Clinimetric
properties of a novel feedback device for assessing gait parameters
in stroke survivors. J Neuroeng Rehabil. 11:302014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang LC, Wu CL, Cheng YY and Tsai KJ:
Deletion of nuclear localizing signal attenuates proinflammatory
activity of prothymosin-alpha and enhances its neuroprotective
effect on transient ischemic stroke. Mol Neurobiol. 54:582–593.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu W, Wang X, Zheng Y, Shang G, Huang J,
Tao J and Chen L: Electroacupuncture inhibits inflammatory injury
by targeting the miR-9-mediated NF-κB signaling pathway following
ischemic stroke. Mol Med Rep. 13:1618–1626. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liesz A and Kleinschnitz C: Editorial:
Mechanisms of neuroinflammation and inflammatory neurodegeneration
in acute brain injury. Front Cell Neurosci. 9:3002015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tao X, Sun X, Yin L, Han X, Xu L, Qi Y, Xu
Y, Li H, Lin Y, Liu K and Peng J: Dioscin ameliorates cerebral
ischemia/reperfusion injury through the downregulation of TLR4
signaling via HMGB-1 inhibition. Free Radic Biol Med. 84:103–115.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Vartanian KB, Stevens SL, Marsh BJ,
Williams-Karnesky R, Lessov NS and Stenzel-Poore MP: LPS
preconditioning redirects TLR signaling following stroke: TRIF-IRF3
plays a seminal role in mediating tolerance to ischemic injury. J
Neuroinflammation. 8:1402011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Derkow K, Krüger C, Dembny P and Lehnardt
S: Microglia induce neurotoxic IL-17+ γδ T cells dependent on TLR2,
TLR4, and TLR9 activation. PLoS One. 10:e01358982015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Belinga VF, Wu GJ, Yan FL and Limbenga EA:
Splenectomy following MCAO inhibits the TLR4-NF-κB signaling
pathway and protects the brain from neurodegeneration in rats. J
Neuroimmunol. 293:105–113. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sun M, Deng B, Zhao X, Gao C, Yang L, Zhao
H, Yu D, Zhang F, Xu L, Chen L and Sun X: Isoflurane
preconditioning provides neuroprotection against stroke by
regulating the expression of the TLR4 signalling pathway to
alleviate microglial activation. Sci Rep. 5:114452015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang J, Wu Y, Weng Z, Zhou T, Feng T and
Lin Y: Glycyrrhizin protects brain against ischemia-reperfusion
injury in mice through HMGB1-TLR4-IL-17A signaling pathway. Brain
Res. 1582:176–186. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang J, Fu B, Zhang X, Chen L, Zhang L,
Zhao X, Bai X, Zhu C, Cui L and Wang L: Neuroprotective effect of
bicyclol in rat ischemic stroke: Down-regulates TLR4, TLR9, TRAF6,
NF-κB, MMP-9 and up-regulates claudin-5 expression. Brain Res.
1528:80–88. 2013. View Article : Google Scholar : PubMed/NCBI
|