Introduction
Intracranial aneurysm (IA) is an abnormal expansion
in the intracranial arteries that weakens the arterial wall by
consistently pushing the vascular wall outwards, which leads to a
risk of aneurysm rupture (1,2). It
occurs in 2–3% of humans worldwide (3,4).
Although treatment and prevention options exist, the majority of
vascular remodeling cases lead to eventual rupture (5). At present, various molecular
mechanisms have been demonstrated to be involved in the
pathophysiology of IA, including cellular apoptosis (6,7).
Apoptosis, an energy-dependent form of cell death,
occurs in a number of physiological conditions, including tissue
remodeling, cell viability and cancer regression (8,9).
Studies have been conducted concerning the involvement of apoptosis
in IA growth and rupture (7,10).
Pentimalli et al (7)
revealed that aneurysm rupture is frequently associated with
elevated levels of apoptosis, which may serve an essential role in
weakening aneurysm walls, thereby resulting in dilation and
rupture. By using rabbits as animal models, Kadirvel et al
(11) reported that intrinsic
pathway apoptosis occurred in aneurysms induced by elastase.
However, few studies have been published concerning the apoptosis
events in IA mice, and the underlying molecular mechanisms are yet
to be established.
MicroRNAs (miRNAs/miRs) are a group of small (18–22
nucleotides) noncoding RNAs that negatively regulate gene
expression either via inhibiting translation or inducing the
degradation of target mRNAs (12),
which allows them to exhibit regulatory effects on various genes,
pathways and complex biological networks in cells. At present, a
wide range of miRNAs have been identified in mice, rat and human
cells, and studies have identified numerous roles for miRNAs in
normal cell homeostasis and disease (13–15).
Certain miRNAs function physiologically in the cardiovascular
system (16–18). For example, miR-9 has been reported
to contribute IA development by inhibiting proliferation and
weakening the contractility of smooth muscle cells (19), while miR-133 is reported to serve
as the key regulator in the phenotypic shift of vascular smooth
muscle cells (VSMCs) in vivo and in vitro (20). In addition, miR-145 has been
reported to be downregulated in atherosclerosis, vascular injury
and aneurysms (21,22). Recently, evidence has emerged that
demonstrates that certain specific miRNAs may serve a vital role in
the development of IA (23,24).
However, the role of miR-29a in the apoptosis process associated
with IA, and the underlying mechanisms, remain to be
elucidated.
The present study therefore used an experimental IA
model and demonstrated that miR-29a is able to regulate
mitochondrial apoptosis, which contributes to the process of
IA.
Materials and methods
Initiation of experimental IA
To induce cerebral aneurysms, male wild-type mice
(C57BL/6; 6–8 weeks; 30–32 g; n=20) were housed in a temperature
(25±1°C) and a humidity (55±5%) and maintained under 12 h light and
12 h dark cycle with free access to food and water. All mice were
purchased from Shanghai SLAC Laboratory Animals Co., Ltd.
(Shanghai, China). Under intraperitoneal Zoletile®
anesthesia (30 mg/kg) with xylazin (10 mg/kg), the left common
carotid artery and the posterior branches of the two renal arteries
were ligated to initiate cerebral aneurysms. Following the surgery,
1% normal saline was substituted for the drinking water to promote
the degree of hypertension. The mice were divided into a control
group with no surgery (n=10) and an aneurysm group (n=10) with
ligation on the left common carotid artery and the posterior
branches of the two renal arteries. The Institutional Animal Care
and Use Committee of the Affiliated Hospital of Hebei University of
Engineering (Hebei, China) approved all procedures prior to the
initiation of the study.
In order to evaluate the effect of miR-29a on IA,
the chemically modified agomir was used to increase miRNA
expression in vivo. miRNA agomir is a chemically modified,
cholesterylated, stable miRNA mimic, and its in vivo
delivery resulted in target silencing similar to the effects
induced by the overexpression of endogenous miRNA. Agomir-29a was
synthesized by Guangzhou RiboBio Co., Ltd. (Guangzhou, China). The
sequence of agomir-29a is 5′-ACUGAUUUCUUUUGGUGUUCAG-3′. The mice
received agomir-29a (100 µl) via tail vein injection (40 mg/kg body
weight; n=10) for 3 consecutive days. The mice in the no surgery
group (n=10) and the aneurysm group (n=10) with ligation were
treated as mentioned above. Expression of miR-29a was detected by
reverse transcription-quantitative polymerase chain reaction
(RT-qPCR; data not shown). At 200 days following the induction
procedure, all mice were euthanized with CO gas. Cerebral arteries
were dissected and stripped from brains under a surgical
microscope. Samples from the regions of aneurysmal dilation on left
posterior communicating artery in the aneurysm group and samples
from the circles of Willis in the control groups were obtained.
Cell cultures
Human brain VSMCs (HBVSMCs; cat. no. CSC-7824W) were
purchased from Creative Bioarray (Shirley, NY, USA). 293-T cells
were purchased from the Type Culture Collection of the Chinese
Academy of Sciences (Shanghai, China). All cells were cultured in
Dulbecco's modified Eagle medium supplemented with 10% fetal bovine
serum, which were purchased from Gibco (Thermo Fisher Scientific,
Inc., Waltham, MA, USA) and 1% penicillin (100 U/l) and
streptomycin (100 µg/ml). Cells were maintained at 37°C in a
humidified atmosphere with 5% CO2. Cells were seeded at
10×106 cells/dish in ten 15-cm dishes 1 day prior to
transfection. The miR-29a mimics, miR-29a inhibitor, and the
corresponding controls [mimics negative control (NC) or inhibitor
NC] were obtained from GeneCopoeia, Inc. (Rockville, MD, USA).
Transfection was performed using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol.
Cells were transfected with 50 nM miR-29a mimics,
miR-29a inhibitor or corresponding controls for 2 h, followed by
treatment with 200 µM H2O2 for 6 h, and cells
were harvested at different times (24, 36 and 48 h) for subsequent
experiments. Cells in the control group were treated with the same
medium without H2O2 (blank group).
Patient samples
Written informed consent was obtained from patients,
according to a protocol approved by the Ethics Committee of the
Affiliated Hospital of Hebei University of Engineering. In total, 2
ml peripheral blood samples were obtained from 24 pairs of patients
with IA and healthy volunteers from the Affiliated Hospital of
Hebei University of Engineering (recruited from March 2014 to April
2015). Twenty-four patients met the inclusion criteria and were
enrolled. Inclusion criteria (25)
were as follows; age ≤70 years (1)
and planned microsurgery or endovascular surgery for unruptured IA
(2). Exclusion criteria (25) were as follows; preoperative
intelligence quotient <80 (n=2) (1); initial modified Rankin scale ≥1 (n=1)
(2); and loss to follow-up (n=3)
(3). There were 28 males and 20
females, and the age range was 35–78 years.
RNA extraction
TissueLyser II (Qiagen, Inc., Valencia, CA, USA) was
used to homogenize the aneurysm and control samples obtained from
mice. Total RNA from tissue, cells and blood samples were isolated
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. Low
molecular weight RNA (<200 nucleotides) was isolated from the
total RNA by mirVana miRNA purification columns (Ambion; Thermo
Fisher Scientific, Inc.) for microarray analysis and RT-qPCR,
according to the manufacturer's protocol. RNA quality and quantity
were determined using a NanoDrop 2000 spectrophotometer (NanoDrop
Technologies; Thermo Fisher Scientific, Inc.).
Microarray and data analysis
Total RNA from samples from IA and sham mice
(3/group) was sent to LC Sciences, LLC (Houston, TX, USA) for miRNA
microarray analysis (miRCURY LNA™ miRNA Array kit; Exiqon A/S,
Vedbaek, Denmark). Data were analyzed by one-way analysis of
variance (ANOVA) using Tukey's multiple comparison test. Expression
normalization was performed using a cyclic LOWESS method (26).
RT-qPCR
RNA was reverse transcribed to cDNA from 100 ng
total RNA with a TIANScript II RT kit (Tiangen Biotech Co., Ltd.,
Beijing, China) at 37°C for 1 h. qPCR was performed with miScript
SYBRGreen PCR kit (Tiangen Biotech Co., Ltd.). Primer sequences
were as follows: U6 forward, 5′-CTCGCTTCGGCAGCACA-3′ and reverse,
5′-AACGCTTCACGAATTTGCGT-3′; miR-29a forward,
5′-TGCGCTAGCACCATCTGAAAT-3′ and reverse, 5′-CAGTGCAGGGTCCGAGGT-3′;
β-actin forward, 5′-GGGAAATCGTGCGTGACATTAAG-3′ and reverse,
5′-TGTGTTGGCGTACAGGTCTTTG-3′; caspase-3 forward,
5′-AGAGGAATGATTGGGGGTG-3′ and reverse, 5′-TTGCTAGGCAGTGGTAGCG-3′;
caspase-9 forward, 5′-CGGAATCACCAATCATTACAT-3′ and reverse,
5′-AGAAACGCCCACAACTGC-3′; caspase-8 forward,
5′-CAGCATTAGGGACAGGAATC-3′ and reverse, 5′-CAGTTATTCACAGTGGCCAT-3′;
myeloid cell leukemia 1 (Mcl-1) forward,
5′-TGAAATCGTTGTCTCGAGTGATG-3′ and reverse,
5′-TCACAATCGCCCCAGTTT-3′; and Cyt-c forward,
5′-CAGTGCCATACTGTGGAAAAGG-3′ and reverse,
5′-TGACCTGTCTTTCGTCCAAACA-3′. qPCR was performed on a StepOnePlus
Real-Time PCR System (Applied Biosystems; Thermo Fisher Scientific,
Inc.), with each reaction performed in triplicate. The PCR
amplification protocol was as follows: An initial 95°C for 5 min
and 40 cycles of 94°C for 15 sec; 55°C for 30 sec; and 70°C for 30
sec, followed by a final extension step of 72°C for 10 min. The
RT-qPCR assays were performed in triplicate and the alteration in
expression level was normalized to the expression of U6 using the
2−ΔΔCq method (27).
Bioinformatics TargetScan 7.0 (http://www.targetscan.org/) (28) and miRanda (http://www.microrna.org/) (29) target gene prediction software
selected Mcl-1 as a target gene of miR-29a.
Terminal deoxynucleotidyl
transferase-mediated bio-dUTP nick-end labelling (TUNEL) apoptosis
analysis of vascular tissues from sham and IA mice
For TUNEL analysis of mouse vascular tissues, the
Roche In Situ Cell Death Fluorescein Detection kit (Roche
Diagnostics, Basel, Switzerland) was used, according to the
manufacturer's protocol. Vascular tissues were fixed in 4%
paraformaldehyde for 3 h at 4°C, paraffin embedded and cut into 4
µm-thick sections. Tissue sections were subsequently de-waxed in
xylene, rehydrated in graded alcohols, and placed in
dH2O. The slides were incubated for 15 min at room
temperature with 20 µg/ml Proteinase K (Gibco BRL; Thermo Fisher
Scientific, Inc.). The slides were rinsed twice with PBS prior to
incubation in TUNEL reaction mixture for 60 min at 37°C. The
Nucleotide Mix was on ice and sufficient rTdT incubation buffer was
prepared for all experimental reactions, 50 µl rTdT incubation
buffer was added to each slide. The sections were covered with
plastic coverslips to ensure even distribution of the reagent. The
DNA fragments were labeled with the rTdT at 37°C for 1 h in the
dark. The sections were stained with 10 nM DAPI (Beyotime Institute
of Biotechnology, Shanghai, China) at 37°C for 3 min and observed
using an inverted fluorescence microscope (Olympus 1X51; Olympus
Corporation, Tokyo, Japan; magnification, ×100; number of fields,
n=5/tissue). TUNEL-positive cells were counted using ImageJ
software version 1.33–1.34 (http://rsbweb.nih.gov/ij/; National Institutes of
Health, Bethesda, MD, USA) for subsequent statistical analysis.
Luciferase reporter assay
miR-29a mimics, miR-29a inhibitors and negative
control (NC) miRs (mimics NC and inhibitor NC) were purchased from
Shanghai GenePharma Co., Ltd. (Shanghai, China). The sequences are
as follows: miR-29a mimics, 5′-TAGCACCATCTGAAATCGGTTA-3′; miR-29a
inhibitor, 5′-TAACCGATTTCAGATGGTGCTA-3′; mimics NC,
5′-AAATGTACTGCGCGTGGAGAC-3′; and inhibitor NC,
5′-UUCUCCGAACGUGUCACGUTT-3′. 293T cells (Type Culture Collection of
the Chinese Academy of Sciences, Shanghai, China) were seeded into
48-well plates (2×105 /well) and cotransfected with
miR-29a mimics/inhibitors/NC miRs (50 nM) and NFAT luciferase
reporter plasmids (200 ng; Promega Corporation, Madison, WI, USA)
containing a wild-type or mutant type of the Mcl-1 3′ untranslated
region (3′-UTR) using Lipofectamine® 2000 reagent
(Invitrogen; Thermo Fisher Scientific, Inc.). At 48 h following
transfection, luciferase activities were measured using the
Dual-Luciferase Reporter Assay System (Promega Corporation,
Madison, WI, USA). Firefly luciferase activity was normalized to
the activity of Renilla luciferase. Each transfection was
performed in triplicate.
Western blot analysis
Cells were lysed using radioimmunoprecipitation
buffer (cat. no. 211-40; AmyJet Scientific, Inc., Wuhan, China).
Total cell protein was extracted from cells or tissues. According
to the manufacturer's protocol, the protein concentration was
determined by a bicinchoninic acid protein assay kit (Beyotime
Institute of Biotechnology, Haimen, China). A total of 2 µg per
lane protein was electrophoresed with 10% SDS-PAGE. Subsequently,
proteins were transferred onto polyvinylidene difluoride membranes
(Bio-Rad Laboratories, Inc., Hercules, CA, USA) and blocked with 5%
skimmed milk at room temperature for 1 h. Following blocking with
milk, the membranes were incubated with specific primary antibodies
at 37°C for 2 h. β-actin was used as the internal reference. The
secondary antibody (1:10,000) was added to the membrane and
incubated for 1 h with agitation at room temperature. Rabbit
anti-cytochrome c (Cyt-c; 1:1,000; cat. no. 4280) and
anti-caspase-3 (caspase-3; cat. no. 9665; 1:1,500) were purchased
from Cell Signaling Technology, Inc. (Danvers, MA, USA). Rabbit
anti-Mcl-1 antibody (Mcl-1; 1:1,000; cat. no. sc-819) and
anti-β-actin (1:1,500; cat. no. sc-8432) were purchased from Santa
Cruz Biotechnology, Inc. (Dallas, TX, USA). The secondary
antibodies included horseradish peroxidase (HRP)-conjugated goat
anti-rabbit immunoglobulin G (1:2,000; cat. no. sc-2004) were
purchased from Santa Cruz Biotechnology, Inc. The signals were
visualized using Immobilon Western Chemiluminescent HRP Substrate
(EMD Millipore, Billerica, MA, USA) and autoradiography.
Densitometric levels of protein signals were quantified and
presented as their ratio to β-actin. Autoradiograms were quantified
by densitometry using Quantity One software version 4.4.0 (Bio-Rad
Laboratories, Inc.).
Cell viability determined using an MTT
assay
Cells (5×105) were seeded on 96-well
plates following trypsin digestion and cell counting, and five
wells in a row were used for each sample. Cells were incubated at
37°C with 5% CO2 for 5 days. An MTT stain was performed,
according to the manufacturer's protocol. Formazan was dissolved by
dimethyl sulfoxide. A microplate reader was used to measure the
optical density 450 value of solution.
Apoptosis assay
A total of 10×104 HBVSMCs transfected
with NC miRs, miR-29a mimics and miR-29a inhibitors, were harvested
by trypsinization at 48 h following transfection. An Annexin
V-Fluorescein Isothiocyanate kit (BD Biosciences, San Jose, CA,
USA) was used to detect apoptotic cells, according to the
manufacturer's protocol. The cells were analyzed with a flow
cytometer (FACScan; BD Biosciences) equipped with CellQuest
software (version 5.1; BD Biosciences). The relative ratio of early
apoptotic cells was compared with control transfection from each
experiment. Each sample was run in triplicate.
Statistical analysis
Data are presented as the mean ± standard deviation
from at least three separate experiments. All experiments were
repeated for at least three times with triplicated samples in each
experiment. One-way ANOVA using Tukey's multiple comparison test
was employed to evaluate the statistical differences among
different groups with SPSS version 19.0 software (IBM Corp.,
Armonk, NY, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
Genes and proteins involved in
mitochondrial apoptosis pathways are upregulated in IA mice
compared with sham controls
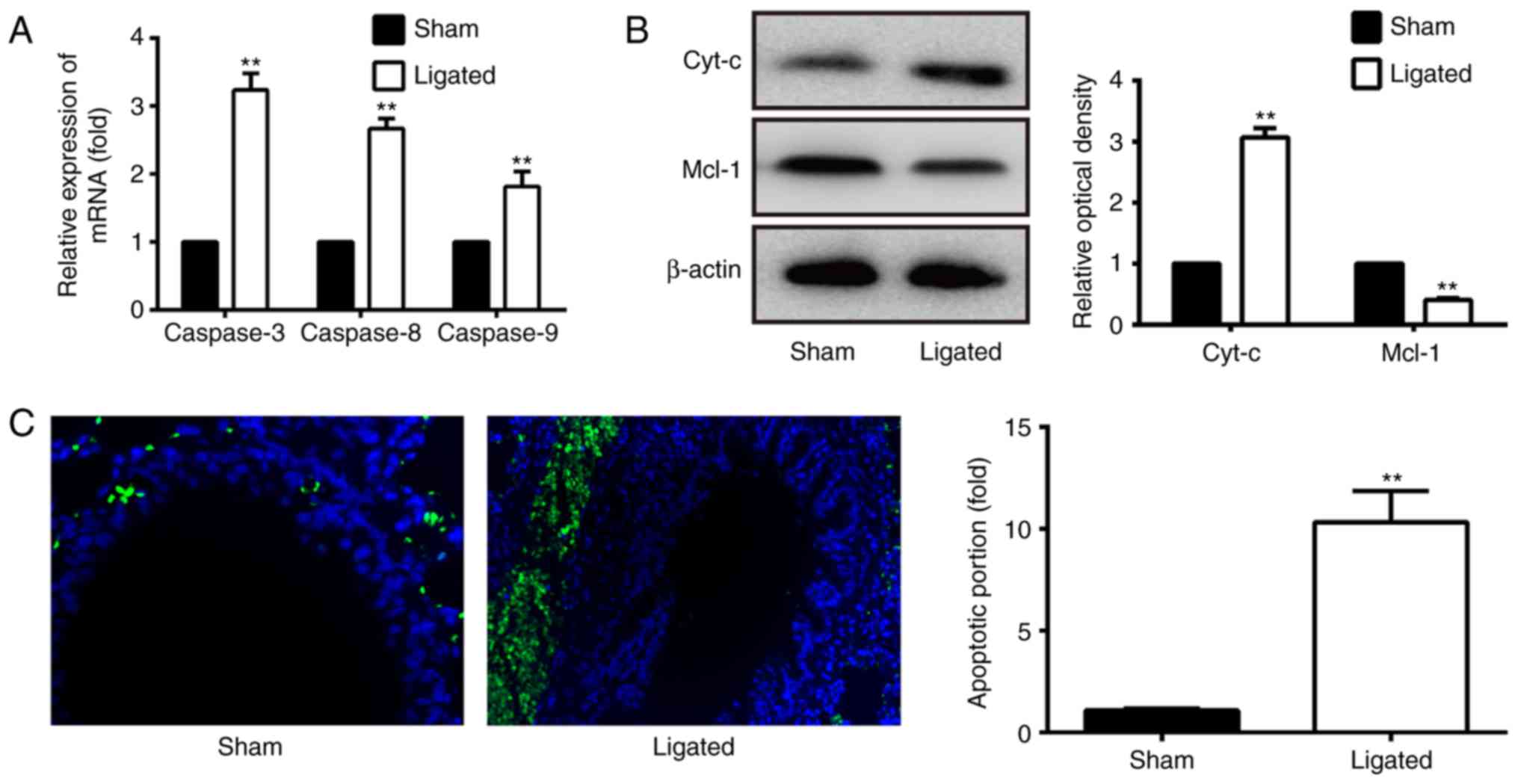
The activation of caspase-3, −8 and −9, which are
apoptotic caspases, may serve a vital role in the process of
mitochondrial apoptosis. In the present study, the mRNA expression
of caspases, including caspase-3, −8 and −9, was analyzed in sham
control and IA mice samples by RT-qPCR. The results demonstrated
that the expression levels of caspase-3, caspase-8 and caspase-9
were significantly upregulated in the IA group compared with sham
controls (P<0.01; Fig. 1A),
which indicate that increased apoptosis occurred in IA.
To further detect whether the mitochondrial pathway
is involved in the apoptosis observed in IA mice, the protein
expression of Cyt-c and Mcl-1 was detected by western blotting. The
results demonstrated that the protein expression of Cyt-c was
activated, while Mcl-1 expression was decreased, in the IA group
compared with the sham controls (Fig.
1B). Additionally, pathological analysis demonstrated that the
apoptosis of VSMCs, accompanied with decreased numbers of VSMCs,
occurred in the IA group (Fig.
1C). The apoptosis of vascular tissue between the two groups
was detected by a TUNEL assay (Fig.
1C). Quantification of TUNEL staining demonstrated that
apoptosis was significantly increased in the IA group compared with
the sham group (P<0.01). These results indicated that
mitochondrial apoptosis may participate in the process of IA.
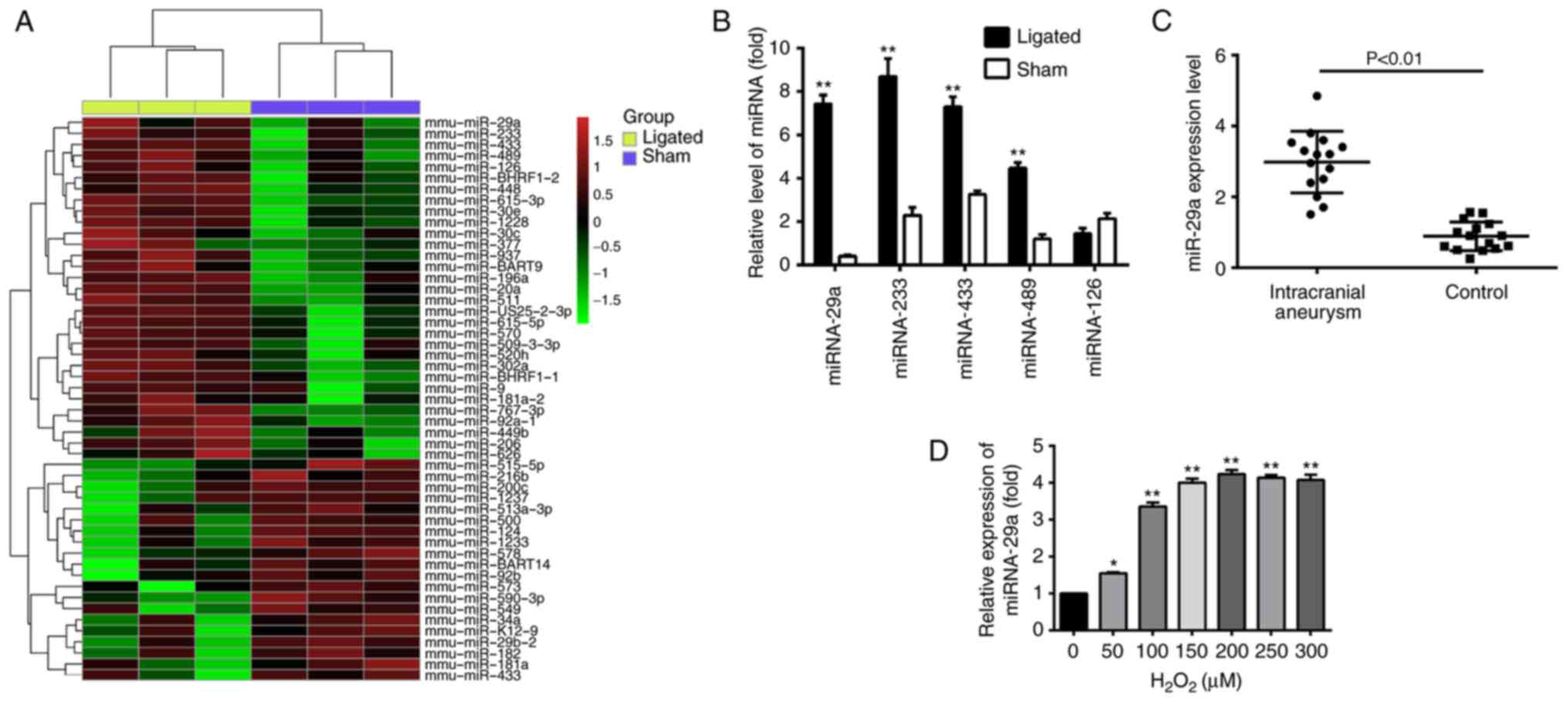
Expression profiles of miRNAs in IAs
and sham control arteries
miRNAs have been reported to regulate the
mitochondrial apoptosis pathway (30,31).
In the present study, to detect the miRNAs that may be involved in
the IA process, an miRNA array was used. Significantly different
expression levels of miRNAs were identified between the IA group
and the sham controls, with 31 miRNAs significantly upregulated and
20 miRNAs significantly downregulated (fold change value >2;
P<0.05) in the IA tissues compared with the control group
(Fig. 2A).
For validation of the microarray data, the most
dysregulated miRs, including miR-29a, miR-233, miR-433, miR-489 and
miR-126 were selected for RT-qPCR analysis. The relative expression
alterations of these miRNA analyzed by RT-qPCR were consistent with
the microarray analysis results, with the exception of miR-126,
which was not significantly altered between the sham control and IA
mice (P<0.05; Fig. 2B). In
order to determine whether miR-29a expression in IA mice models
exhibits a similar pattern to clinical patients, a clinical
experiment was performed to detect miR-29a expression in patients
with IA and healthy volunteers. The blood samples obtained from the
case group included 24 patients with IA, while the control group
included 24 healthy volunteers. miR-29a expression was detected and
compared between the two groups. The results of RT-qPCR
demonstrated that the expression of miR-29a in patients with IA was
significantly higher compared with healthy controls (P<0.01;
Fig. 2C).
miR-29a has been reported as a potential biomarker
in the development of IA (31). In
addition, it has been reported that upregulation of miR-29a may be
implicated in cell apoptosis by targeting Mcl-1 expression in rat
germ cell death (32). Previous
studies have demonstrated that miR-29a is able to induce cell
apoptosis by activating caspase proteins (33,34).
For example, miR-29a was reported to be responsible for the
activation of caspase-3-induced K562 cell apoptosis (35). Therefore, miR-29a was selected for
further investigation in the present study.
miR-29a downregulation inhibits the
apoptosis of HBVSMCs
An in vitro
H2O2-induced apoptosis model was established
to mimic IA (36). HBVSMCs were
treated with H2O2 (0–300 µM) and miR-29a
expression levels were significantly increased in a
concentration-dependent manner, peaking at 200 µM, compared with
the 0 µM group (Fig. 2D). Based on
these results, the concentration of 200 µM
H2O2 was selected for subsequent
experiments.
HBVSMC cells were transfected with miR-29a mimics or
inhibitor. Fig. 3A demonstrates
that the expression of miR-29a was significantly upregulated in the
miR-29a mimic treatment group compared with the mimics NC group
(P<0.01), while miR-29a expression was significantly decreased
in cells with miR-29a inhibitor treatment compared with the
inhibitor NC group (P<0.01). Following transfection with miR-29a
inhibitor for 24, 36 and 48 h, the cell viability of VSMCs treated
with H2O2 was significantly increased,
compared with the inhibitor NC treatment group (P<0.01; Fig. 3B). However, the
H2O2-induced reduction of HBVSMC viability
was significantly enhanced following transfection with miR-29a
mimics, compared with the mimics NC treatment group (P<0.01;
Fig. 3C). These results indicated
that the miR-29a inhibitor reversed
H2O2-induced cell viability inhibition, while
miR-29a mimics promoted H2O2-induced cell
viability inhibition.
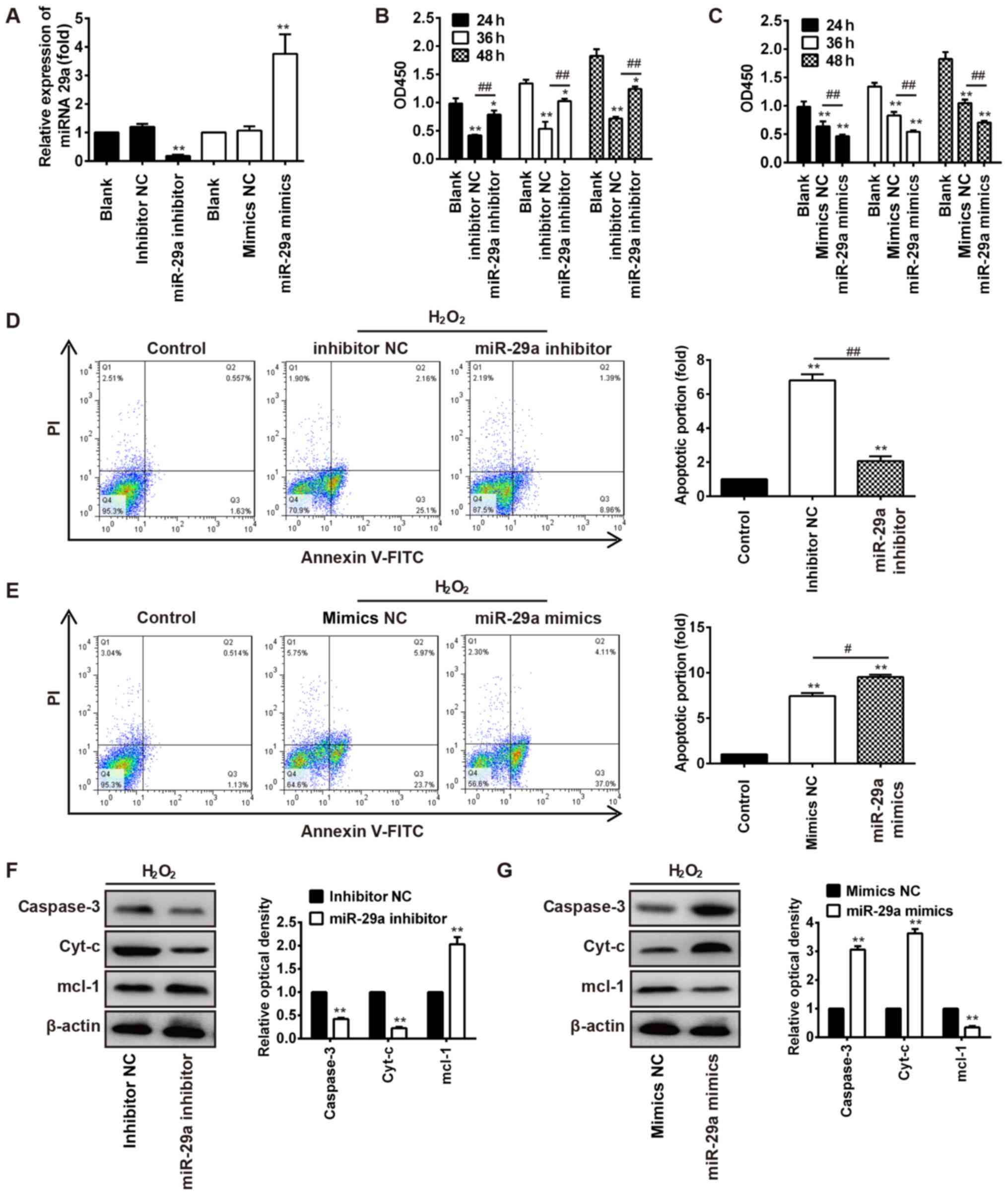 | Figure 3.miR-29a inhibition protects HBVSMCs
against H2O2-induced apoptosis. (A) miR-29a
expression was detected by reverse transcription-quantitative
polymerase chain reaction in HBVSMCs treated with miR-29a mimics or
miR-29a inhibitor. **P<0.01 vs. inhibitor NC or mimics NC group.
(B) Cell viability was analyzed using a Cell Counting kit-8 assay.
The OD 450 values were detected by using a SpectraMax microplate
reader. miR-29a inhibitor reversed
H2O2-induced cell viability inhibition in a
time-dependent manner in HBVSMCs. *P<0.05, **P<0.01 vs. Blank
group; ##P<0.01 vs. inhibitor NC group. (C)
Transfection of miR-29a mimics enhanced
H2O2-induced cell viability inhibition in
HBVSMCs. **P<0.01 vs. Blank group, ##P<0.01 vs.
mimics NC group. (D) Apoptotic cells were detected by Annexin
V-FITC and PI apoptosis detection kit, and were analyzed with a
flow cytometer. Treatment with miR-29a inhibitor protected HBVSMCs
from H2O2-induced apoptosis. Cells in the
lower left quadrant represent intact viable cells (Annexin negative
and PI negative), cells in the lower right quadrant represent early
apoptotic cells (Annexin positive and PI negative), cells in the
upper right quadrant represent late apoptotic cells (Annexin
positive and PI positive) and cells in the upper left quadrant
represent necrotic cells (Annexin negative and PI positive).
**P<0.01 vs. blank group; ##P<0.01, as indicated.
(E) miR-29a mimic treatment enhanced the
H2O2-induced apoptosis of HBVSMCs.
**P<0.01 vs. blank group; #P<0.05, as indicated.
(F) Western blotting results demonstrated that the protein levels
of caspase-3 and Cyt-c were significantly reduced following miR-29a
inhibitor transfection, while anti-apoptosis protein Mcl-1
expression was increased. **P<0.01 vs. inhibitor NC group. (G)
Western blotting results demonstrated that the protein levels of
the pro-apoptosis proteins, caspase-3 and Cyt-c, were significantly
increased following miR-29a mimics transfection, while
anti-apoptosis protein Mcl-1 expression was decreased. **P<0.01
vs. mimics NC group. miR, microRNA; HBVSMCs, human brain vascular
smooth muscle cells; NC, negative control; OD, optical density; PI,
propidium iodide; Cyt-c, cytochrome c; Mcl-1, myeloid cell leukemia
1; FITC, fluorescein isothiocyanate. |
Furthermore, treatment with 200 µM
H2O2 indicated that transfection of miR-29a
inhibitor protected against H2O2-induced
HBVSMC apoptosis, while miR-29a mimics worsened the
H2O2-induced HBVSMC apoptosis (P<0.05;
Fig. 3D and E). Western blot
analysis was performed to observe alterations in the protein
expression of apoptosis-associated proteins in
H2O2-induced HBVSMCs transfected with miR-29a
inhibitor or mimics. In H2O2-treated cells
with miR-29a inhibitor treatment, the expression levels of the
proapoptotic proteins caspase-3 and Cyt-c were decreased, while
Mcl-1 was increased, compared with the inhibitor NC group
(P<0.01; Fig. 3F). Conversely,
in cells treated with miR-29a mimics, the expression levels of the
proapoptotic proteins caspase-3 and Cyt-c were increased, while
Mcl-1 was decreased, compared with the mimics NC group (P<0.01;
Fig. 3G). These results indicated
that miR-29a may regulate cell apoptosis during IA.
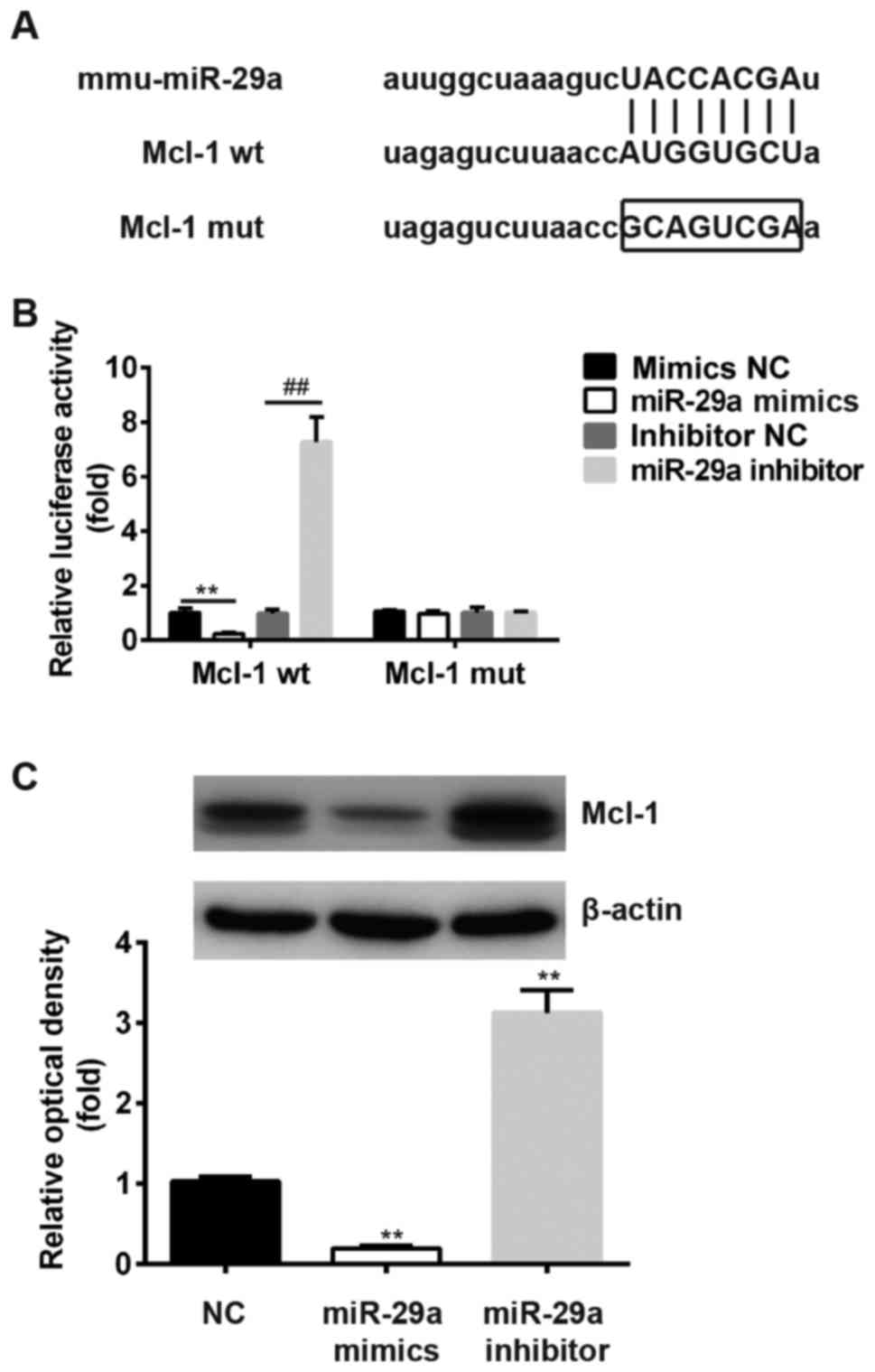
miR-29a directly targets Mcl-1
miR-29a has been reported to directly target Mcl-1
in intestinal epithelial cells (37). In the present study, bioinformatics
tools, TargetScan and miRanda, were used to identify the target
genes of miR-29a. Fig. 4A
demonstrates that Mcl-1 is a theoretical target gene of miR-29a.
The wild-type 3′-UTR of the Mcl-1 gene was cloned and inserted it
into the downstream region of a luciferase reporter gene.
Subsequently, miR-29a mimics/inhibitors were cotransfected with
different luciferase 3′-UTR constructs into 293T cells. The results
of luciferase assays demonstrated that miR-29a mimics significantly
weakened the relative luciferase activity of the luciferase
reporter containing the wild-type 3′-UTR of Mcl-1 mRNA compared
with mimics NC group, whereas the knockdown of miR-29a increased
luciferase activity (P<0.01; Fig.
4B). To determine whether Mcl-1 mRNA is directly targeted by
miR-29a, the predicted binding site of miR-29a in the 3′-UTR of
Mcl-1 was muted. By contrast, the luciferase activities of
luciferase reporters containing mutant 3′-UTRs of Mcl-1 luciferase
activity remained unaltered in miR-29a mimic/inhibitor-transfected
293T cells compared with the respective mimics/inhibitor NC groups
(Fig. 4B).
Results from the western blot analysis demonstrated
that the protein expression of Mcl-1 was markedly decreased
following miR-29a mimic treatment and increased following miR-29a
inhibitor treatment, compared with the NC group (Fig. 4C).
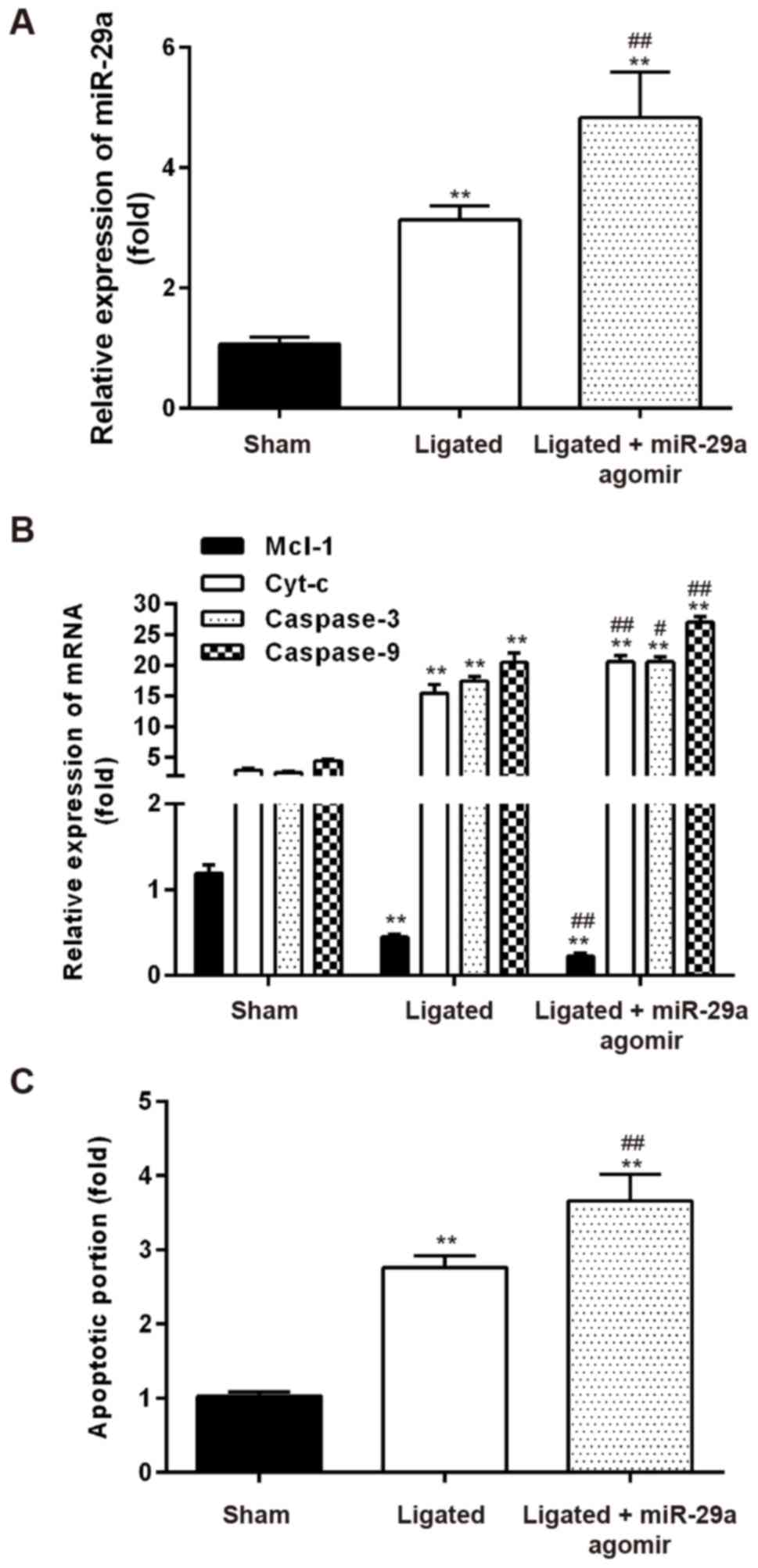
Overexpression of miR-29a promotes
apoptosis in vivo
As miR-29a overexpression in vitro enhanced
VHBSMC apoptosis (Fig. 3B) and
Mcl-1 was determined as a direct target of miR-29a (Fig. 4), the present study aimed to
determine whether miR-29a may regulate apoptosis by regulating
Mcl-1 expression in vivo. The miR-29a function in IA injury
was detected by administration of the agomir-miR-29a to mice. As
demonstrated in Fig. 5A, the
expression of miR-29a in IA ligated mice was significantly higher
compared with sham mice, while the level of miR-29a is further
significantly increased in agomir-miR-29a + IA ligated mice.
RT-qPCR results demonstrated that genes involved in mitochondrial
apoptosis pathways, including Cyt-c, caspase-3 and caspase-9, were
markedly increased, while Mcl-1 was decreased, in the IA ligated
mice compared with sham controls (Fig.
5B). In agomir-miR-29a + IA ligated mice, Cyt-c, caspase-3 and
caspase-9 were significantly increased, while the anti-apoptosis
gene Mcl-1 was decreased, compared with the IA ligated group
(Fig. 5B), indicating that miR-29a
overexpression promoted mitochondrial apoptosis in vivo.
TUNEL staining confirmed that apoptosis in tissue sections was
significantly increased in IA + miR-29a agomir mice, compared with
IA ligated mice (P<0.01; Fig.
5C).
Taken together, these results indicated that miR-29a
overexpression may stimulate mitochondrial apoptosis pathways,
subsequently contributing to the injury caused by IA.
Discussion
The development and growth of aneurysms involve the
complex events of arterial wall cells remodeling (38). Previous studies have demonstrated
that apoptosis is involved in the pathogenesis of aneurysms
(6,7). However, the molecular mechanisms
underlying the stimulation of apoptosis in the mouse IA model
remains unknown.
To investigate the apoptotic events of IA mice
model, the present study investigated the mRNA expression of
caspases associated with the mitochondrial apoptotic pathway,
including caspase-3, −8 and −9. The activation of caspases serves
an essential role during the process of apoptosis (39), which may be caused by an extrinsic
or intrinsic pathway, which lead to a terminal common pathway
(40). Caspase-8 activation is
involved in the extrinsic pathway and caspase-9 is involved in the
intrinsic pathway. In the mitochondrial pathway, the release of
Cyt-c constitutes an apoptotic complex, which subsequently results
in the activation of caspase-9. The activation of caspase-8 and −9
leads to the subsequent activation of caspase-3, which mediates
cell apoptosis (11). In the
present study, the expression levels of the proapoptotic genes,
caspase-3, −8 and −9, were markedly increased in IA mice samples,
indicating that the mitochondrial pathway may be involved in
apoptosis in the IA mouse model, as caspase activation serves an
important function in apoptosis.
It has been confirmed that miRNAs regulate various
biological events, including development, metabolism,
proliferation, differentiation and apoptosis (41). Studies have demonstrated the
contribution of miRNAs together with their target genes in
controlling IA injury in a number of models (18,19).
Therefore, to investigate the molecular mechanism of apoptosis in
IA injury, the present study focused on miRNA regulation. Results
from microarray analysis and RT-qPCR demonstrated that among the
upregulated miRNAs in IA mice models, miR-29a demonstrated the
highest upregulation level.
It has been demonstrated that miR-29a may regulate
the mitochondrial apoptosis pathway and activate caspases,
including caspase-3 and −8 (31).
Additionally, it has been reported that miR-29a expression was
significantly increased following ischemia-reperfusion (I/R) and
induced apoptosis, and transfection with anti-miR-29a reduced the
levels of apoptosis in cells and provided them with protection
against I/R injury (42). The
primary aim of the present study was to investigate whether miR-29
may be involved in IA-associated apoptosis, a key process in the
pathogenesis of IA. The present study identified that mitochondrial
apoptosis may occur in IA. To investigate the association between
miR-29a and caspases in IA, in vitro experiments were
performed, which demonstrated that miR-29a mimics induced the
activation of caspase-3, −8 and −9. These results indicated that
upregulation of miR-29a may promote the mitochondrial apoptosis
pathway in IA.
Apoptosis is a parameter of IA injury. VSMCs
constitute the majority of the blood vessel wall (43) and their growth or programmed cell
apoptosis serves an important role in the altered geometry of
vessels in vascular diseases, thus promoting the rupture of
aneurysms. Accordingly, VSMC apoptosis was utilized as an in
vitro model of IA (35).
Results from the present study demonstrated that miR-29a inhibition
attenuated the apoptosis of HBVSMCs; whereas, miR-29a
overexpression increased HBVSMC apoptosis.
Mcl-1 is reported to be associated with the
programming of differentiation and cell viability/death (44). In the present study, Mcl-1 was
identified as being directly targeted by miR-29a and involved in
IA-induced apoptosis, and an in vivo model was established
to determine whether apoptosis may be regulated by miR-29a in IA.
Results from the RT-qPCR and apoptosis detection analysis
demonstrated that the overexpression of miR-29a significantly
enhanced the mitochondrial apoptosis of IA injury, as may be
inferred from the increased mRNA expression levels of genes
involved in the mitochondrial pro-apoptosis pathways (caspase-3 and
−8 and Cyt-c), decreased anti-apoptotic gene Mcl-1 expression and
enhanced cell death by DNA fragmentation compared with the IA
ligated group.
It is therefore concluded that miR-29a may
contribute to the progression of IA by regulating the mitochondrial
apoptotic pathways. However, further studies are required for
verification of these results. Therefore, miR-29a may be a novel
target for the prevention and treatment of IA.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
WZ performed the experiments, contributed to data
analysis and wrote the paper. J-YS analyzed the data. HZ
conceptualized the study design, and contributed to data analysis
and experimental materials. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Experiments involving human and animal tissues were
approved by the Ethics Committee of The Affiliated Hospital of
Hebei University of Engineering. Written informed consent was
obtained from all human participants.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kacar E, Nas Fatih O, Erdogan C and
Hakyemez B: Intracranial aneurysm rupture during flow diverter
stent placement: Successful treatment with stent-in-stent
combination. Diagn Interv Imaging. 96:411–413. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Komotar RJ, Zacharia BE, Otten ML, Mocco J
and Lavine SD: Controversies in the endovascular management of
cerebral vasospasm after intracranial aneurysm rupture and future
directions for therapeutic approaches. Neurosurgery. 62:897–907.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lee D, Ahn SJ, Cho ES, Kim YB, Song SW,
Jung WS and Suh SH: High prevalence of intracranial aneurysms in
patients with aortic dissection or aneurysm: Feasibility of
extended aorta CT angiography with involvement of intracranial
arteries. J Neurointerv Surg. 9:1017–1021. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ronkainen A, Hernesniemi J, Ryynänen M,
Puranen M and Kuivaniemi H: A ten percent prevalence of
asymptomatic familial intracranial aneurysms: Preliminary report on
110 magnetic resonance angiography studies in members of 21 Finnish
familial intracranial aneurysm families. Neurosurgery. 35:208–213.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Frösen J, Piippo A, Paetau A, Kangasniemi
M, Niemelä M, Hernesniemi J and Jääskeläinen J: Remodeling of
saccular cerebral artery aneurysm wall is associated with rupture:
Histological analysis of 24 unruptured and 42 ruptured cases.
Stroke. 35:2287–2293. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Guo F, Li Z, Song L, Han T, Feng Q, Guo Y,
Xu J, He M and You C: Increased apoptosis and cysteinyl aspartate
specific protease-3 gene expression in human intracranial aneurysm.
J Clin Neurosci. 14:550–555. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pentimalli L, Modesti A, Vignati A,
Marchese E, Albanese A, Di Rocco F, Coletti A, Di Nardo P, Fantini
C, Tirpakova B and Maira G: Role of apoptosis in intracranial
aneurysm rupture. J Neurosurg. 101:1018–1025. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu CY, Su JC, Huang TT, Chu PY, Huang CT,
Wang WL, Lee CH, Lau KY, Tsai WC, Yang HP, et al: Sorafenib
analogue SC-60 induces apoptosis through the SHP-1/STAT3 pathway
and enhances docetaxel cytotoxicity in triple-negative breast
cancer cells. Mol Oncol. 11:266–279. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Apoptosis in cancer pathogenesis and
anti-cancer therapy. New perspectives and opportunities. Gregory
and Christopher D.: AntiCancer Res. 37:3712016.
|
|
10
|
Chen Z, Miao H, Feng H and Zhu G: Rupture
of an infectious intracranial aneurysm involving two parent
arteries after surgical treatment of infective endocarditis.
Neurosciences (Riyadh). 16:72–75. 2011.PubMed/NCBI
|
|
11
|
Kadirvel R, Ding YH, Dai D, Lewis DA and
Kallmes DF: Intrinsic pathway-mediated apoptosis in
elastase-induced aneurysms in rabbits. AJNR Am J Neuroradiol.
31:165–169. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen B, Li H, Zeng X, Yang P, Liu X, Zhao
X and Liang S: Roles of microRNA on cancer cell metabolism. J
Transl Med. 10:2282012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
An YR, Kim SJ, Oh MJ, Kim HM, Shim IS, Kim
PJ, Choi K and Hwang SY: Analysis of microRNA and gene expression
profiling in triazole fungicide-treated HepG2 cell line.
Toxicology. 303:94–98. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kirigin FF, Lindstedt K, Sellars M,
Ciofani M, Low SL, Jones L, Bell F, Pauli F, Bonneau R, Myers RM,
et al: Dynamic microRNA gene transcription and processing during T
cell development. J Immunol. 188:3257–3267. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sessa WC: MicroRNA regulation of
cardiovascular functions. Arterioscler Thromb Vasc Biol.
31:23692011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cui RR, Li SJ, Liu LJ, Yi L, Liang QH, Zhu
X, Liu GY, Liu Y, Wu SS, Liao XB, et al: MicroRNA-204 regulates
vascular smooth muscle cell calcification in vitro and in vivo.
Cardiovasc Res. 96:320–329. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cheng Y and Zhang C: MicroRNA-21 in
cardiovascular disease. J Cardiovasc Transl Res. 3:251–255. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Luo J, Jin H, Jiang Y, Ge H, Wang J and Li
Y: Aberrant expression of microRNA-9 contributes to development of
intracranial aneurysm by suppressing proliferation and reducing
contractility of smooth muscle cells. Med Sci Monit. 22:4247–4253.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Torella D, Iaconetti C, Catalucci D,
Ellison GM, Leone A, Waring CD, Bochicchio A, Vicinanza C, Aquila
I, Curcio A, et al: MicroRNA-133 controls vascular smooth muscle
cell phenotypic switch in vitro and vascular remodeling in vivo.
Circ Res. 109:880–893. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Engelhardt S and Leierseder S: Coinciding
functions for miR-145 in vascular smooth muscle and cardiac
fibroblasts. J Mol Cell Cardiol. 65:105–107. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rangrez AY, Massy ZA, Metzinger-Le Meuth V
and Metzinger L: miR-143 and miR-145: Molecular keys to switch the
phenotype of vascular smooth muscle cells. Circ Cardiovasc Genet.
4:197–205. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang GK, Zhu JQ, Zhang JT, Li Q, Li Y, He
J, Qin YW and Jing Q: Circulating microRNA: A novel potential
biomarker for early diagnosis of acute myocardial infarction in
humans. Eur Heart J. 31:659–666. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wu X, Zhang J, Huang Q, Yang P, Chen J and
Liu J: MicroRNA-92a regulates expression of kruppel-like factor2 in
rabbit model of intracranial aneurysm. Cell Mol Biol
(Noisy-le-grand). 61:44–48. 2015.PubMed/NCBI
|
|
25
|
Wardlaw JM and White PM: The detection and
management of unruptured intracranial aneurysms. Brain.
123:205–221. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bolstad BM, Irizarry RA, Astrand M and
Speed TP: A comparison of normalization methods for high density
oligonucleotide array data based on variance and bias.
Bioinformatics. 19:185–193. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Friedman RC, Farh KK, Burge CB and Bartel
DP: Most mammalian mRNAs are conserved targets of microRNAs. Genome
Res. 19:92–105. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Betel D, Wilson M, Gabow A, Marks DS and
Sander C: The microRNA.org resource: Targets and expression.
Nucleic Acids Res. 36:(Database Issue). D149–D153. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zeng CW, Zhang XJ, Lin KY, Ye H, Feng SY,
Zhang H and Chen YQ: Camptothecin induces apoptosis in cancer cells
via microRNA-125b-mediated mitochondrial pathways. Mol Pharmacol.
81:578–586. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang WH, Wang YH, Zheng LL, Li XW, Hao F
and Guo D: MicroRNA-29a: A potential biomarker in the development
of intracranial aneurysm. J Neurol Sci. 364:84–89. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Meunier L, Siddeek B, Vega A, Lakhdari N,
Inoubli L, Bellon RP, Lemaire G, Mauduit C and Benahmed M:
Perinatal programming of adult rat germ cell death after exposure
to xenoestrogens: Role of microRNA miR-29 family in the
down-regulation of DNA methyltransferases and Mcl-1. Endocrinology.
153:1936–1947. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Li MN, Li SS, Meng FS, Han P, Zhang WH and
Zhang LB: Mechanism and influence of down-regulation of miR-29a-3p
expression on apoptosis in mouse macrophage. J South China Agricult
Univ. 39:64–69. 2018.
|
|
34
|
Khamisipour G, Mansourabadi E, Naeimi B,
Moazzeni A, Tahmasebi R, Hasanpour M, Mohammadi MM, Mansourabadi Z
and Shamsian S: Knockdown of microRNA-29a regulates the expression
of apoptosis-related genes in MCF-7 breast carcinoma cells. Mol
Clin Oncol. 8:362–369. 2018.PubMed/NCBI
|
|
35
|
Wang X, Zhong H, Wang L, Dong Y, Jia A, Mo
Q and Zhang C: MiR-29 induces K562 cell apoptosis by
down-regulating FoxM1. Med Sci Monit. 21:3115–3120. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wu L, Li X, Li Y, Wang L, Tang Y and Xue
M: Proliferative inhibition of danxiongfang and its active
ingredients on rat vascular smooth muscle cell and protective
effect on the VSMC damage induced by hydrogen peroxide. J
Ethnopharmacol. 126:197–206. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lv B, Liu Z, Wang S, Liu F, Yang X, Hou J,
Hou Z and Chen B: MiR-29a promotes intestinal epithelial apoptosis
in ulcerative colitis by down-regulating Mcl-1. Int J Clin Exp
Pathol. 7:8542–8552. 2014.PubMed/NCBI
|
|
38
|
Laine MT, Björck M, Beiles CB, Szeberin Z,
Thomson I, Altreuther M, Debus ES, Mani K, Menyhei G and Venermo M:
Few internal iliac artery aneurysms rupture under 4 cm. J Vasc
Surg. 65:76–81. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Piao L, Canguo Z, Wenjie L, Xiaoli C,
Wenli S and Li L: Lipopolysaccharides-stimulated macrophage
products enhance Withaferin A-induced apoptosis via activation of
caspases and inhibition of NF-κB pathway in human cancer cells. Mol
Immunol. 81:92–101. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chang JP, Chen MC, Liu WH, Lin YS, Huang
YK, Pan KL, Ho WC, Fang CY, Chen CJ and Chen HC: Mitochondrial
apoptotic pathway activation in the atria of heart failure patients
due to mitral and tricuspid regurgitation. Exp Mol Pathol.
99:65–73. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zandberga E, Kozirovskis V, Ābols A,
Andrējeva D, Purkalne G and Linē A: Cell-free microRNAs as
diagnostic, prognostic, and predictive biomarkers for lung cancer.
Genes Chromosomes Cancer. 52:356–369. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wang L, Niu X, Hu J, Xing H, Sun M, Wang
J, Jian Q and Yang H: After myocardial ischemia-reperfusion,
miR-29a, and Let7 could affect apoptosis through regulating IGF-1.
Biomed Res Int. 2015:2454122015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Xu F, Ahmed AS, Kang X, Hu G, Liu F, Zhang
W and Zhou J: MicroRNA-15b/16 attenuates vascular neointima
formation by promoting the contractile phenotype of vascular smooth
muscle through targeting YAP. Arterioscler Thromb Vasc Biol.
35:2145–2152. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yang T, Buchan HL, Townsend KJ and Craig
RW: MCL-1, a member of the BLC-2 family, is induced rapidly in
response to signals for cell differentiation or death, but not to
signals for cell proliferation. J Cell Physiol. 166:523–536. 1996.
View Article : Google Scholar : PubMed/NCBI
|