Introduction
Features of myocardial remodeling include
cardiomyocyte hypertrophy, apoptosis, fibrosis and alterations in
cardiovascular function (1). The
renin-angiotensin (Ang) system (RAS) serves an important role in
cardiac remodeling. Ang II induces cardiac hypertrophy and fibrosis
in hypertension (2–4). Furthermore, a newly discovered member
of the RAS, (pro)renin receptor [(P)RR], is a novel drug target due
to its crucial involvement in cardiovascular injury (5,6).
Binding of (pro)renin/renin to (P)RR increases Ang I
formation from angiotensinogen and induces Ang II-independent
intracellular signaling (7). The
experimental evidence for the role of (P)RR was obtained from
studies of progressive nephropathy developed in
human-(P)RR-transgenic rats following the occurrence of
hypertension (8–10). Rats overexpressing the (P)RR gene
in vascular smooth muscle cells develop high blood pressure and a
high level of plasma aldosterone (9). Direct local intramyocardial
adenovirus-mediated gene delivery demonstrated that (P)RR triggers
the deterioration of cardiac function and participates in
extracellular matrix remodeling (11). However, AT1 receptor blockade does
not prevent the decrease in the ejection fraction, suggesting that
(P)RR-induced worsening of cardiac function is independent of Ang
II generation. Furthermore, rats with (P)RR overexpression develop
glomerulosclerosis with increased extracellular signal-regulated
kinase 1/2 (ERK1/2) phosphorylation (12). Also, human prorenin activates p38
mitogen activated protein kinase and phosphorylates heat shock
protein-27 in neonatal rat cardiomyocytes (12). These combined results suggest that
Ang II-independent effects of (P)RR are important in the
development of cardiovascular diseases.
These studies demonstrate that in rats with
hypertension, elevated (P)RR expression levels may correlate with
cardiovascular complications by a direct effect exerted by (P)RR
signaling mechanisms. However, the precise mechanisms of the (P)RR
signaling in the heart remain obscure.
Previous experiments have used an inhibitory peptide
termed the handle region peptide (HRP), which binds competitively
to the (P)RR with prorenin (13–15).
Ichihara et al (13)
reported that infusion of HRP decreases cardiac Ang II levels and
attenuates cardiac fibrosis in a diabetic rat model of nephropathy.
However, Feldt et al (14)
did not find that HRP prevents (pro)renin signaling in a
double-transgenic rat model. The discrepancy between these reports
may be due to the different animal models and different doses of
HRP used in experiments. Since prorenin is highly species-specific,
different HRPs exist for different rat models in which there are
different prorenin and renin levels (15). Therefore, it is necessary to
further investigate the effects of HRP on (P)RR and its signals in
various animal models. In the present study, HRP was used in an
aortic constricted rat model to investigate effects on cardiac
remodeling and the (P)RR signaling pathway.
Therefore, the overall goal of the present study was
to test the hypothesis that overexpression of cardiac (P)RR is
induced by aortic constriction, resulting in activation of the
phospholipase C (PLC)-β3-protein kinase C (PKC)-ERK1/2-Raf-1
proto-oncogene, serine/threonine kinase (Raf 1) signaling pathway,
thereby inducing cardiac remodeling and hypertension.
Materials and methods
Animals
Male Sprague-Dawley rats weighing 180–200 g (aged 6
weeks, Shandong Lukang Co., Ltd., Jining, China) were housed in a
temperature-controlled (24–26°C with 60% humidity) animal facility
(12-h light/dark cycle) with ad libitum access to rat chow
and tap water. All animal experiments were performed in accordance
with the NIH guidelines (Guide for the Care and Use of Laboratory
Animals) and were approved by the Animal Ethics Committee of
Taishan Medical University (Taian, China).
Experimental protocols
A total of 75 male Sprague-Dawley rats were
randomized to the abdominal aortic constriction and sham-operated
groups, as described previously (16). Briefly, animals were anesthetized
with sodium pentobarbitone (60 mg/kg, intraperitoneally).
Sham-operated animals underwent the same procedure without the
aortic ligation [control (C), n=15]. Aortic ligation surgery was
undertaken in the rest of the rats. Following recovery, they were
randomly allocated to the following groups: i) vehicle [aortic
ligation (AL), n=15]; ii) PLC-β3 inhibitor U73122 (Selleck
Chemicals Houston, TX, USA; U, n=15) 40 µg/kg/day for 4 weeks; iii)
(P)RR inhibitor HRP (Phoenix Pharmaceuticals Inc., Burlingame, CA,
USA; H, n=15) 4 µg/kg/day for 4 weeks; and iv) both U73122 and HRP
combined at the same doses (U+H, n=15) delivered by a subcutaneous
mini-pump.
Blood pressure was measured by the tail-cuff method
using Blood Pressure Meter (Softron BP-2012A; Beijing Incorporated,
Beijing, China) as described previously (16,17).
Following 4 weeks of treatment, the animals were
sacrificed by an overdose of sodium pentobarbitone (100 mg/kg,
intraperitoneally). Blood plasma was collected and hearts were
dissected and weighed. The left ventricle (LV) was dissected and
snap-frozen in isopentane/dry ice for immunohistochemical studies.
The remainder of the LV was preserved at −80°C for reverse
transcription-polymerase chain reaction (RT-PCR) and western blot
analysis.
Ang I and Ang II assays
Plasma samples were obtained by centrifugation of
blood (2,000 × g) at 4°C for 10 min. Plasma Ang I and Ang II
concentration (ng/ml) was quantitatively determined by a
radioimmunoassay using commercial kits (cat. no. 10059; Puer Weiye
Biotech Ltd., Beijing, China) based on methods described previously
(16–18).
Measurement of (P)RR, PLC-β3, PKC-α,
PKC-ε, ERK1/2 and Raf-1 mRNAs
Total RNA was extracted from heart tissue using
TRIzol® (Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). RT-PCR was carried out using PCR Reagent kits
with 5X M-MLV buffer and dNTPs (Takara Bio, Inc., Otsu, Japan).
Template cDNA was prepared using reverse transcriptase (Takara Bio,
Inc.). Detailed information on all primers used in the study,
including (P)RR, PLC-β3, PKC-α, PKC-ε, ERK1/2 and Raf-1 is
presented in Table I. The products
of PCR were size-fractionated on 1% agarose gels and detected by
ethidium bromide staining. Following quantification of band
intensities using a Bio-Rad Gel Documentation System (version 4.6;
Bio-Rad Laboratories, Inc., Hercules, CA, USA), the relative
steady-state level of mRNA was calculated following normalizing to
GAPDH (as internal reference gene) and presented as gene/GAPDH for
each sample in experimental groups. In each experimental run the
mRNA value for the control heart was given a value of 1 and the
amount of target gene mRNA in the experimental hearts was expressed
as a ratio of this value (16,17).
Then, the comparison of relative quantity of mRNA was statistically
conducted between groups.
 | Table I.Primers used in reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Primers used in reverse
transcription-quantitative polymerase chain reaction.
| Primers | Sequence (5′-3′) | Length (bp) |
|---|
| (P)RR | F:
CATAAGCATCTCGCCAAGG | 224 |
|
| R:
ACCAGGGATGTGTCGAATGA |
|
| PLC-β3 | F:
GGTCCTACCACTTCCTCCACTA | 317 |
|
| R:
GCCTCCTCTTCCTCCTTCAC |
|
| PKC-α | F:
GTTTACCCGGCCAACGACT | 134 |
|
| R:
GTGGGTTGCTTGAAGAAGCG |
|
| PKC-ε | F:
CAAGGTGTAGCGAGTGAGTGG | 314 |
|
| R:
CGGACTGTTGGTCTTCTGCT |
|
| ERK1 | F:
TATCAACACCACCTGCGACC | 129 |
|
| R:
ATGATCTCTGGGGCTCGGTA |
|
| ERK2 | F:
TTGTTCCCAAACGCTGACTC | 187 |
|
| R:
TAGGTAAGTCGTCCAGCTCC |
|
| Raf-1 | F:
AGCACCACCCTCTTTCACAA | 432 |
|
| R:
AGCATCACCTCACTGGCTTC |
|
| GAPDH | F:
CAGTGCCAGCCTCGTCTCAT | 595 |
|
| R:
AGGGGCCATCCACAGTCTTC |
|
Western blot analysis
Protein was extracted with Radioimmunoprecipitation
Assay buffer from heart tissue and subjected to western blot
analysis. Briefly, following protein concentration determination
via the bicinchoninic acid method, a total of 30 µg protein was
loaded per lane, separated in a 10% SDS-PAGE and transferred to
polyvinylidene membranes. The membranes were blocked at room
temperature for 3 h with 5% fat-free milk in TBS/0.5% Tween-20,
then incubated with primary antibodies (Abcam, Cambridge, USA)
against (P)RR (cat. no. ab40790), PLC-β3 (cat. no. ab14247), PKC-α
(cat. no. ab32376) and Raf-1 (cat. no. ab137435; all diluted to
1:1,200), and primary antibodies (Cell Signaling Technology, Inc.,
Danvers, MA, USA) against PKC-ε (cat. no. 2683) and ERK1/2 (cat.
no. 4695; both diluted to 1:1,000, at 4°C overnight respectively,
followed by horseradish peroxidase-conjugated secondary antibody
(cat. no. ZB-2301; Beijing Zhongshan Jinqiao Biotech Co., Ltd.,
Beijing, China; 1:5,000) for 2 h. Chemiluminescence was detected
with an enhanced chemiluminescence kit (Pierce; Thermo Fisher
Scientific, Inc.) and subsequent exposition to Hyperfilm (GE
Healthcare, Chicago, IL, USA). The signals of blots were quantified
by using a Bio-Rad gel documentation system (version 4.6) (Bio-Rad
Laboratories, Inc.). GAPDH (analyzed using an antibody from OriGene
Technologies, Inc., Beijing, China) served as the loading control.
The protein expression levels of (P)RR, PLC-β3, PKC-α, PKC-ε,
ERK1/2 and Raf-1 were calculated in relation to GAPDH. The value of
the control (sham-operated group) was expressed as 1 and then
values from other experimental groups were compared with this to
determine fold-change for each protein (16,17).
(P)RR immunohistochemistry
Sections of unfixed hearts were embedded in
PolyFreeze Tissue Freezing Medium (Polysciences, Warrington, PA,
USA) and snap-frozen (−35°C) in isopentane/dry ice. Cryostat
sections, 7 µm thick, were cut and fixed in cold 4%
paraformaldehyde for 30 min at 4°C. The sections were incubated
with the dilution of primary antibody against (P)RR (Abcam) at
dilution of 1:200 in PBS solution. Following washing with PBS, they
were incubated with peroxidase-conjugated anti-rabbit IgG (goat
polyclonal, 1:5,000; cat. no. ZF-0311; OriGene Technologies, Inc.,
Beijing, China) for 1 h at 4°C. The color was developed with 0.05%
3,3′-diaminobenzidine tetrahydrochloride (DAB) and 0.01%
H2O2. Myocyte (P)RR staining was assessed by
measuring the area of stained tissue within a given field. A total
of 20 fields were analyzed for each animal. The immunostaining area
of (P)RR was obtained in a given area and expressed as a percentage
of the entire image (16–18).
Statistical analysis
All data are expressed as the mean ± standard error
of the mean. Statistical significance of MAP, (P)RR, PLC-β3, PKC-α,
PKC-ε, ERK1/2 and Raf-1 and plasma Ang I, Ang II data was analyzed
by a Repeated Measures analysis of variance in SPSS (version 11.0;
SPSS, Inc., Chicago, IL, USA) followed by post-hoc multiple
comparison tests with the Least Significant Difference post hoc
test. P<0.05 was considered to indicate a statistically
significant differences.
Results
Inhibitors of (P)RR and PLC-β3
attenuate hypertension and cardiac hypertrophy induced by aortic
constriction in vivo
Mean arterial pressure (MAP) and heart rate changes
are presented in Table II. Aortic
constriction increased MAP (P<0.05), but HRP, U73122, or both
inhibitors combined, significantly decreased MAP compared with the
aortic ligation group (P<0.05). Compared with aortic restricted
(AL) rats, HRP and U73122 significantly decreased MAP (P<0.05).
There was no difference in heart rate between groups.
| Table II.Blood pressure, heart rate, Ang I and
Ang II concentration in groups of rats. |
Table II.
Blood pressure, heart rate, Ang I and
Ang II concentration in groups of rats.
| BP and HR | C | AL | U73122 | HRP | U+H |
|---|
| MAP (mmHg) | 100±3 | 155±5a | 128±5a,b | 131±6a,b | 126±6a,b |
| HR (beats
min−1) | 316±14 | 304±11 | 332±16 | 328±15 | 334±17 |
| Ang I and Ang
II |
| Ang I
(ng ml−1) | 21.8±1.2 |
32.5±2.8a |
16.8±1.0a,b |
20.5±0.9a,b |
10.2±0.7a,b |
| Ang II
(ng ml−1) | 5.8±0.3 |
10.2±0.8a |
5.2±0.5a,b |
4.5±0.2a,b |
4.3±0.5a,b |
There was no significant difference in body weight
among groups (Table III).
However, heart, left and right ventricle and atrial wall weights
were significantly increased in hypertensive rats from aortic
constriction (P<0.05). HRP, U73122 and the combination of the
two significantly reduced the whole heart, left and right
ventricle, and atrial weight (P<0.05), respectively. The ratios
of heart, left and right ventricle weight to body weight
demonstrated a significant difference in these animals (P<0.05;
Table III).
| Table III.Body and heart weights in groups of
rats. |
Table III.
Body and heart weights in groups of
rats.
|
| C | AL | U73122 | HRP | U+H |
|---|
| Body weight
(g) | 242±5 | 239±4 | 244±5 | 241±4 | 239±7 |
| Cardiac
parameters |
| HW
(g) | 0.921±0.062 |
0.969±0.035a |
0.823±0.029a,b |
0.845±0.044a,b |
0.812±0.024a,b |
| LV
(g) | 0.709±0.031 |
0.758±0.021a |
0.689±0.048a,b |
0.648±0.022a,b |
0.663±0.035a,b |
| RV
(g) | 0.165±0.008 |
0.175±0.009a |
0.156±0.005a,b |
0.159±0.002a,b |
0.146±0.002a,b |
| AW
(g) | 0.037±0.001 |
0.044±0.002a |
0.034±0.004a,b |
0.035±0.005b |
0.035±0.007a,b |
| HW/BW g/(100
g) | 0.379±0.007 |
0.408±0.005a |
0.336±0.01a,b |
0.350±0.006a,b |
0.338±0.008a,b |
| LV/BW g/(100
g) | 0.292±0.004 |
0.318±0.006a |
0.281±0.005a,b |
0.267±0.002a,b |
0.277±0.004a,b |
| RV/BW g/(100
g) | 0.068±0.006 |
0.073±0.004a |
0.063±0.003a,b |
0.065±0.004a,b |
0.061±0.004a,b |
| AW/BW g/(100
g) | 0.015±0.003 |
0.019±0.003a |
0.013±0.005a,b |
0.014±0.004b |
0.012±0.005a,b |
Aortic constriction significantly
affects levels of Ang I and Ang II in the circulation
Alterations of plasma Ang I and Ang II concentration
are presented in Table II. The
concentration of Ang I and Ang II significantly increased in AL
rats (P<0.05). Administration of HRP, U73122 and the combination
of the two significantly decreased plasma Ang I and Ang II levels
(P<0.05) compared with both control and AL rats,
respectively.
Increase in cardiac
PLC-b3-PKC-ERK1/2-Raf-1 signaling expression induced by aortic
constriction is reduced following inhibition of HRP and U73122
Cardiac (P)RR mRNA significantly increased in aortic
constricted rats (P<0.05). HRP treatment, U73122 treatment or
the two combined downregulated (P)RR mRNA in the heart (P<0.05;
Fig. 1A) respectively, compared
with AL animals. A similar alteration in cardiac (P)RR protein
level was observed (Fig. 1B).
Compared with controls, PLC-β3 mRNA and its protein levels
increased in AL rats, and the levels were significantly reduced
with treatment of HRP or U73122 or the two combined in rats,
compared with controls and AL animals (P<0.05; Fig. 1C and D).
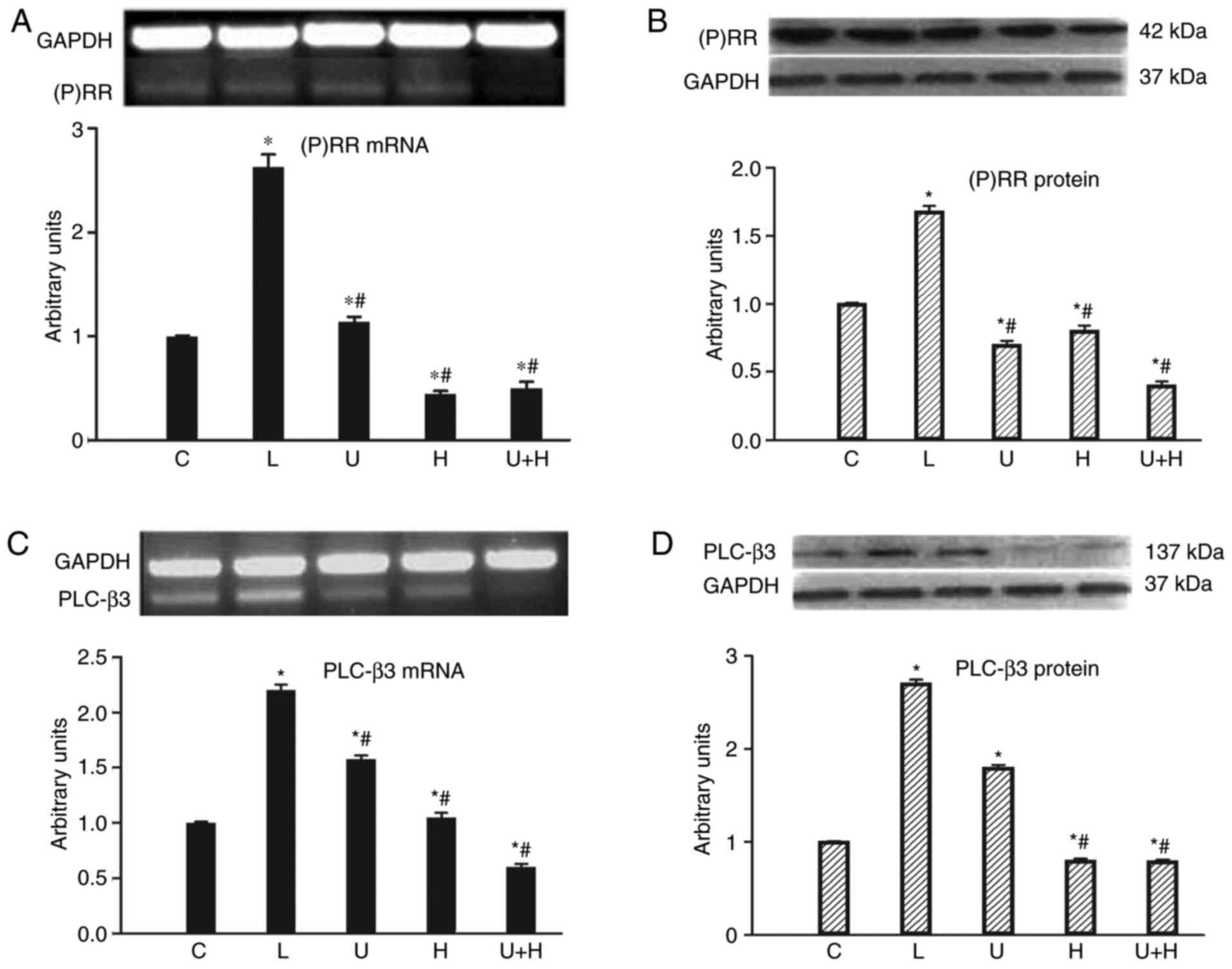 | Figure 1.Levels of (A) cardiac (P)RR mRNA (B)
and its protein, and (C) PLC-β3 mRNA and (D) its protein in the C
(n=15), AL, (n=13), U (n=13), HRP (H, n=14) or U73122 + HRP (U+H,
n=13)-treated rats. Data are expressed as the mean ± standard error
of the mean. *P<0.05 vs. control; #P<0.05 vs. AL.
AL, aortic ligation; (P)RR, (pro)renin receptor; U73122; H, handle
region peptide/HRP; U+H, U73122 + HRP; PLC, phospholipase C; C,
control. |
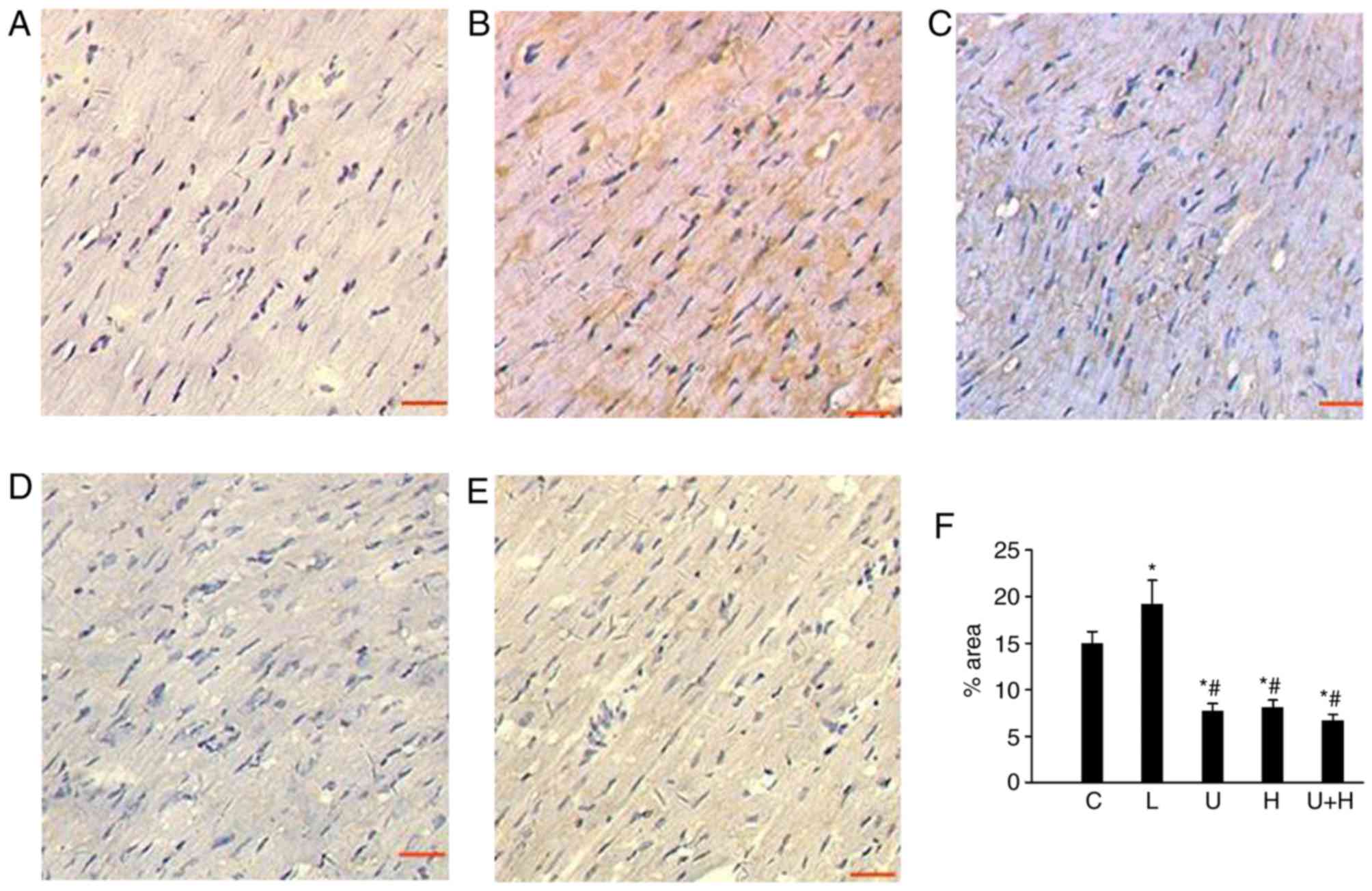
Immunohistochemistry results for (P)RR are presented
in Fig. 2. In the sham-operated
rats, (P)RR was expressed in cardiomyocytes at a basal level
(Fig. 2A and F). Cardiac (P)RR
immunostaining level was significantly upregulated in AL rats
(P<0.05; Fig. 2B and F).
U73122, HRP and the combined treatment of the two significantly
decreased cardiac (P)RR levels compared with AL and control
animals, respectively (P<0.05; Fig.
2C-F).
Cardiac PKC-α mRNA was significantly increased in AL
rats (P<0.05) and was significantly reduced by administration of
U73122, HRP and both reagents combined (P<0.05; Fig. 3A). Cardiac PKC-α protein levels
significantly increased in AL group (P<0.05) and were
significantly suppressed in U73122, HRP or combined groups
(P<0.05; Fig. 3B).
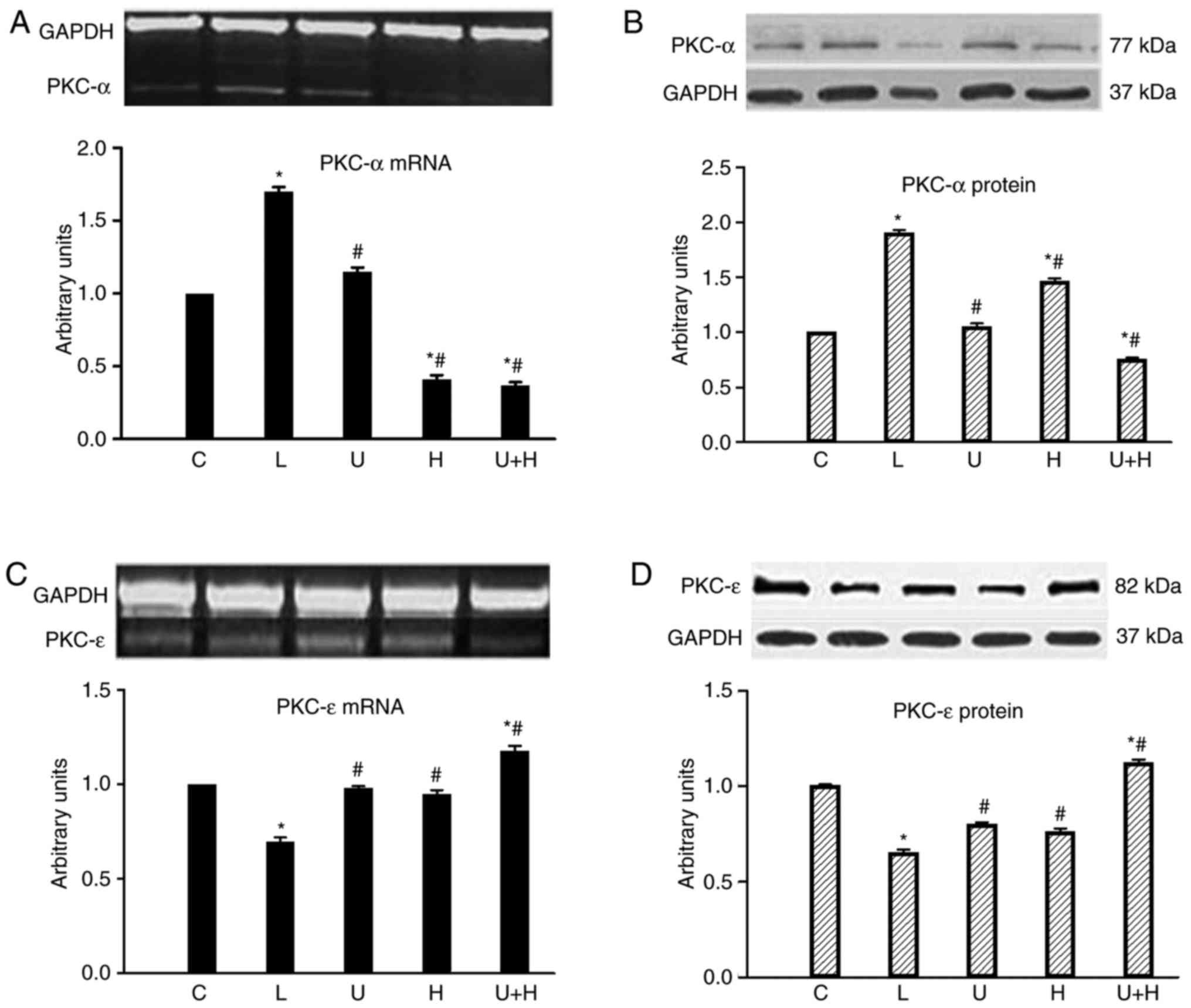 | Figure 3.Levels of (A) cardiac PKC-α mRNA and
(B) its protein, and (C) PKC-ε mRNA (D) and its protein in C
(n=14), AL (n=12), U (n=13), H (n=12) or U+H (n=13)-treated rats.
Data are expressed as the mean ± standard error of the mean.
*P<0.05 vs. C; #P<0.05 vs. AL. AL, aortic
ligation; (P)RR, (pro)renin receptor; U73122; H, handle region
peptide/HRP; U+H, U73122 + HRP; C, control; PKC, protein kinase
C. |
Cardiac PKC-ε mRNA level was significantly reduced
in AL rats (P<0.05), however it was significantly elevated by
treatment of U73122 (P<0.05) and both treatments combined
(P<0.05; Fig. 3C). The
alteration in cardiac PKC-ε protein expression level was similar to
the trend demonstrated in mRNA expression level (P<0.05;
Fig. 3D).
With aortic constriction, the level of ERK1/2 mRNA
in the heart significantly increased (P<0.05). Treatment with
U73122, HRP and the two reagents combined significantly
downregulated ERK1/2 mRNA (P<0.05; Fig. 4A). Furthermore, cardiac ERK1/2
protein levels were reduced by administration of both inhibitors
(P<0.05; Fig. 4B).
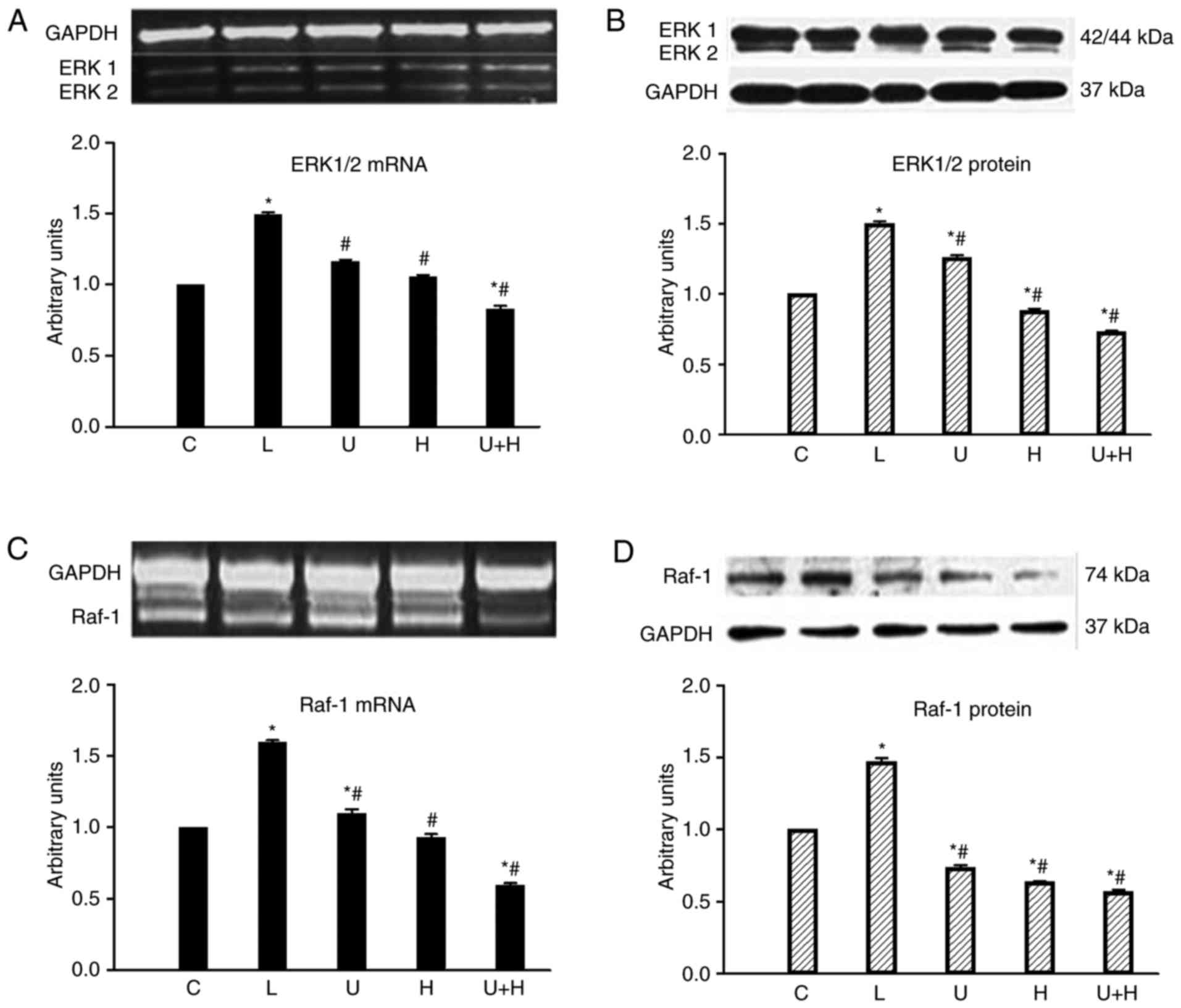 | Figure 4.Levels of (A) cardiac ERK1/2 mRNA (B)
and its protein, and (C) Raf-1 mRNA and (D) its protein in C
(n=14), AL (n=14), U (n=13), H (n=12) or U+H (n=13)-treated rats.
Data are expressed as the mean ± standard error of the mean.
*P<0.05 vs. control; #P<0.05 vs. AL. AL, aortic
ligation; (P)RR, (pro)renin receptor; U73122; H, handle region
peptide/HRP; U+H, U73122 + HRP; C, control; ERK1/2, extracellular
signal regulated kinase; Raf-1, Raf-1 proto-oncogene,
serine/threonine kinase. |
Aortic constriction increased Raf-1 mRNA levels
(P<0.05). U73122, HRP and both treatments combined decreased
Raf-1 mRNA (P<0.05), compared with AL rats (Fig. 4C). Cardiac Raf-1 protein levels
increased in AL rats and were then significantly downregulated in
U73122, HRP and combined treatment groups (P<0.05; Fig. 4D).
Discussion
The present study, to the best of the author's
knowledge, has described for the first time, an association between
aortic restriction by partial abdominal aortic ligation leading to
an increase in (P)RR and PLC-β3 expression and cardiac hypertrophy
(as evidenced by an increase in heart weight). Administration of
HRP and U73122 downregulated the expression of (P)RR and PLC-β3,
which indicated an association between the activities of cardiac
(P)RR and PLC-β3. The results obtained from the current
hypertensive animal model suggested a role for (P)RR and PLC-β3 in
the pathophysiology of heart diseases. The present study
demonstrated that aortic restriction upregulated the levels of
(P)RR in the heart through the enhancement of PLC-β3, PKC, ERK1/2
and Raf-1.
A number of studies have suggested involvement of
(P)RR in the development of cardiovascular diseases (19,20).
However, less is known about the mechanism by which the level of
(P)RR expression is regulated in the heart in hypertension.
Assessment of the mechanisms associated with aortic
restriction-induced (P)RR expression will help better understanding
of the role of the receptor in the heart in the pathophysiology of
hypertension complications and has the potential for development of
novel therapeutic tools for treatment of heart damage.
While the expression of (P)RR and PLC-β3 is
upregulated by aortic restriction, the results of the present study
also demonstrated that plasma Ang I and Ang II levels increased in
the circulation, which suggested that catalytic capacity in
angiotensinogen conversion leads to increased Ang II formation. The
(pro)renin receptor (P)RR has been considered to function by two
different mechanisms: i) Prorenin bound to this receptor generates
Ang I and then Ang II; and ii) prorenin/renin binding to (P)RR
leads to the activation of the signaling pathways followed by
upregulation of hypertrophic genes independent of Ang II (21–24).
The present study demonstrated that intracellular
PKC, ERK1/2 and Raf-1 signals enhanced (P)RR expression, as
inhibition of the receptor and PLC-β3 attenuated not only the
aortic restriction-induced expression of this receptor, but also
the expression levels of PKC-α, ERK1/2 and Raf-1 in the heart. The
level of cardiac PKC-α decreased following administration of
U73122, accompanied with a reduction of cardiac hypertrophy,
suggesting that PKC-α may act as a regulator of myocyte
hypertrophic growth. Furthermore, the present study demonstrated
that blockade of the receptor by HRP increased the PKC-ε level,
implicating that PKC-ε may serve an opposite role to PKC-α in the
regulation of the receptor expression in the signal pathway. These
results are consistent with previous reports of increased PKC-α
activity in a signaling network that programs developmental and
pathological cardiomyocyte hypertrophic growth (25–27).
The current data suggested that inhibition of PLC-β3
attenuated the aortic restriction-induced activation of PKC-α,
ERK1/2 and Raf-1, which suggested that PLC-β3 is an upstream
regulator of this intracellular signal. Notably, inhibition of both
(P)RR and PLC-β3 together produced a more profound effect on PKC-α,
ERK1/2 and Raf-1 and implied the interaction between these signals
in the pathway. PLC-β3 is considered to serve an important role in
the signal transduction mechanisms of cardiac hypertrophy (28–30).
The present study provided evidence for the involvement of PLC-β3
in the signal transduction mechanism in the transcriptional and
translational regulation of PKC, ERK1/2 and Raf-1 in the heart, as
gene and protein expression levels exhibited similar alteration
trends. The data indicated that the inhibitory response to U73122
and HRP was likely attributed to attenuation of specific PLC-β3
expression and subsequent activities on cardiac (P)RR. Although
inhibitors of PKC and ERK1/2 were not used in this study to measure
their specific responses, the inhibitory effects of U73122 and HRP
observed on the expression of PKC, ERK1/2 and Raf-1 indicated that
aortic restriction upregulated the expression of (P)RR through
mechanisms dependent on PLC-β3, PKC, ERK1/2 and Raf-1 signaling
pathways.
The present study confirmed the involvement of
intracellular PKC, ERK1/2 and Raf-1 signals in the regulation of
(P)RR expression. Furthermore, these intracellular factors may
influence expression of cardiac (P)RR by administration of this
receptor and PLC-β3 inhibitors. Since PKC and ERK1/2 have been
reported to be involved in myocyte hypertrophy (31,32),
the present findings demonstrated a similar effect for these
factors in cardiomyocytes in response to high blood pressure and
cardiac remodeling induced by aortic restriction.
Despite the interesting results of this study, there
are still possible limitations. First, the authors primarily
focused on the effects of two drugs (HRP and U73122) on the (P)RR
signaling pathway and cardiac remodeling. The authors did not test
the effects of other drugs on the pathway and the heart including
PKC inhibitors. However, the results of the present study
established that an association was present between (P)RR, PLC-β3
and various other signals. Secondly, fixed doses of the drugs were
administered during the treatment period. Dose escalation was not
conducted during the treatment period. In addition, this is the
first study of its kind in which HRP and U73122 were used to test
the effects of each drug and the combination of a (P)RR inhibitor
and a PLC-β3 inhibitor. Based on this study, further studies should
be conducted with more animal models.
In conclusion, the results of the present study
demonstrated that (P)RR and PLC-β3 genes together with PKC, ERK1/2
and Raf-1 are upregulated in the heart in hypertensive rats induced
by aortic restriction. The (P)RR and PLC-β3 were reduced in the
heart tissue on administration of their inhibitors. In addition,
PKC-α, ERK1/2 and Raf-1 were downregulated. It is likely that the
elevation of (P)RR expression induced by aortic restriction was due
to the PLC-β3 activation and subsequent signal transduction events
through PKC-ERK1/2-Raf-1 signaling. Ang II may initiate cardiac
hypertrophy and enhance PLC-β3 and ERK1/2 (33,34),
as well as increase the expression of Raf-1. Therefore, the results
suggested that (P)RR-PLC-β3-PKC-ERK1/2-Raf-1 signaling can be
considered as an important signaling pathway of (P)RR in the heart
and may therefore constitute novel therapeutic targets for the
prevention and treatment of cardiac hypertrophy.
Acknowledgements
Not applicable.
Funding
The present study was supported by Science and
Technology Department of Shandong Province (grant no.
2016GSF201207) and the National Natural Science Foundation of China
(grant no. 81270336).
Availability of data and materials
The analyzed datasets generated during the study are
available from the corresponding author on reasonable request.
Authors' contributions
YZ designed the present study, and wrote and revised
the manuscript. JW, CZ and JZ performed the experiments. CL, AZ and
AL analyzed the data. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
All animal experiments were performed in accordance
with the NIH guidelines (Guide for the Care and Use of Laboratory
Animals) and were approved by the Animal Ethics Committee of
Taishan Medical University (Taian, China).
Patient consent for publication
Not applicable.
Competing interest
The authors declare that they have no competing
interest.
References
|
1
|
De Mello WC: Novel aspects of angiotensin
II action in the heart. Implications to myocardial ischemia and
heart failure. Regul Pept. 166:9–14. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ainscough JF, Drinkhill MJ, Sedo A, Turner
NA, Brooke DA, Balmforth AJ and Ball SG: Angiotensin II type-1
receptor activation in the adult heart causes blood
pressure-independent hypertrophy and cardiac dysfunction.
Cardiovasc Res. 81:592–600. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Dang MQ, Zhao XC, Lai S, Wang X, Wang L,
Zhang YL, Liu Y, Yu XH, Liu Y, Li HH and Xia YL: Gene expression
profile in the early stage of angiotensin II-induced cardiac
remodeling: A time series microarray study in a mouse model. Cell
Physiol Biochem. 35:467–476. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yang J, Zhu HH, Chen GP, Ye Y, Zhao CZ,
Mou Y and Hu SJ: Inhibition of farnesyl pyrophosphate synthase
attenuates angiotensin II-induced cardiac hypertrophy and fibrosis
in vivo. Int J Biochem Cell Biol. 45:657–666. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chan SH and Chan JY: (Pro)renin receptor
as a therapeutic target for the treatment of hypertension?
Hypertension. 65:278–279. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Oshima Y, Morimoto S and Ichihara A: Roles
of the (pro)renin receptor in the kidney. World J Nephrol.
3:302–307. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gonzalez AA, Luffman C, Bourgeois CR, Vio
CP and Prieto MC: Angiotensin II-independent upregulation of
cyclooxygenase-2 by activation of the (Pro)renin receptor in rat
renal inner medullary cells. Hypertension. 61:443–449. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Burcklé CA, Jan Danser AH, Müller DN,
Garrelds IM, Gasc JM, Popova E, Plehm R, Peters J, Bader M and
Nguyen G: Elevated blood pressure and heart rate in human renin
receptor transgenic rats. Hypertension. 47:552–556. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Danser AH: (Pro)renin receptors: Are they
biologically relevant? Curr Opin Nephrol Hypertens. 18:74–78. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kaneshiro Y, Ichihara A, Sakoda M,
Takemitsu T, Nabi AH, Uddin MN, Nakagawa T, Nishiyama A, Suzuki F,
Inagami T and Itoh H: Slowly progressive, angiotensin
II-independent glomerulosclerosis in human (pro)renin
receptor-transgenic rats. J Am Soc Nephrol. 18:1789–1795. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Alcazar O, Cousins SW, Striker GE and
Marin-Castano ME: (Pro)renin receptor is expressed in human retinal
pigment epithelium and participates in extracellular matrix
remodeling. Exp Eye Res. 89:638–647. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sakoda M, Ichihara A, Kaneshiro Y,
Takemitsu T, Nakazato Y, Nabi AH, Nakagawa T, Suzuki F, Inagami T
and Itoh H: (Pro)renin receptor-mediated activation of
mitogen-activated protein kinases in human vascular smooth muscle
cells. Hypertens Res. 30:1139–1146. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ichihara A, Kaneshiro Y, Takemitsu T,
Sakoda M, Suzuki F, Nakagawa T, Nishiyama A, Inagami T and Hayashi
M: Nonproteolytic activation of prorenin contributesto development
of cardiac fibrosis in genetic hypertension. Hypertension.
47:894–900. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Feldt S, Maschke U, Dechend R, Luft FC and
Muller DN: The putative (pro)renin receptor blocker HRP fails to
prevent (pro)renin signaling. J Am Soc Nephrol. 19:743–748. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lu X, Danser AH and Meima ME: HRP and
prorenin: Focus on the (pro)renin receptor and vacuolar H+-ATPase.
Front Biosci (Schol Ed). 3:1205–1215. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang Y, Li B, Wang B, Zhang J, Wu J and
Morgan T: Alteration of cardiac ACE2/Mas expression and cardiac
remodelling in rats with aortic constriction. Chin J Physiol.
57:335–342. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang Y, Ma L, Wu J and Chen T:
Hydronephrosis alters cardiac ACE2 and Mas receptor expression in
mice. J Renin Angiotensin Aldosterone Syst. 16:267–274. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang Y, Wu J, Zhang Z, Wang B, Chen P and
Jing X: Effect of low sodium intake and β-blockade on renin
synthesis and secretion in mice with unilateral ureteral ligation.
Hypertens Res. 33:1258–1263. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Connelly KA, Advani A, Kim S, Advani SL,
Zhang M, White KE, Kim YM, Parker C, Thai K, Krum H, et al: The
cardiac (pro)renin receptor is primarily expressed in myocyte
transverse tubules and is increased in experimental diabetic
cardiomyopathy. J Hypertens. 29:1175–1184. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hirose T, Mori N, Totsune K, Morimoto R,
Maejima T, Kawamura T, Metoki H, Asayama K, Kikuya M, Ohkubo T, et
al: Gene expression of (pro)renin receptor is upregulated in hearts
and kidneys of rats with congestive heart failure. Peptides.
30:2316–2322. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nguyen G, Delarue F, Burcklé C, Bouzhir L,
Giller T and Sraer JD: Pivotal role of the renin/prorenin receptor
in angiotensin II production and cellular responses to renin. J
Clin Invest. 109:1417–1427. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Saris JJ, Hoen PA, Garrelds IM, Dekkers
DH, den Dunnen JT, Lamers JM and Jan Danser AH: Prorenin induces
intracellular signaling in cardiomyocytes independently of
angiotensin II. Hypertension. 48:564–571. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cousin C, Bracquart D, Contrepas A and
Nguyen G: Potential role of the (pro)renin receptor in
cardiovascular and kidney diseases. J Nephrol. 23:508–513.
2010.PubMed/NCBI
|
|
24
|
Nguyen G and Muller DN: The biology of the
(pro)renin receptor. J Am Soc Nephrol. 21:18–23. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Makary S, Voigt N, Maguy A, Wakili R,
Nishida K, Harada M, Dobrev D and Nattel S: Differential protein
kinase C isoform regulation and increased constitutive activity of
acetylcholine-regulated potassium channels in atrial remodeling.
Circ Res. 109:1031–1043. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Otani H, Yoshioka K, Nishikawa H, Inagaki
C and Nakamura T: Involvement of protein kinase C and RhoA in
protease-activated receptor 1-mediated F-actin reorganization and
cell growth in rat cardiomyocytes. J Pharmacol Sci. 115:135–143.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Palfi A, Bartha E, Copf L, Mark L, Gallyas
F Jr, Veres B, Kalman E, Pajor L, Toth K, Ohmacht R and Sumegi B:
Alcohol-free red wine inhibits isoproterenol-induced cardiac
remodeling in rats by the regulation of Akt1 and protein kinase C
alpha/beta II. J Nutr Biochem. 20:418–425. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Atef ME and Anand-Srivastava MB: Enhanced
expression of Gqα and PLC-β1 proteins contributes to vascular
smooth muscle cell hypertrophy in SHR: Role of endogenous
angiotensin II and endothelin-1. Am J Physiol Cell Physiol.
307:C97–C106. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang L, Malik S, Pang J, Wang H, Park KM,
Yule DI, Blaxall BC and Smrcka AV: Phospholipase Cε hydrolyzes
perinuclear phosphatidylinositol 4-phosphate to regulate cardiac
hypertrophy. Cell. 153:216–227. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Woodcock EA, Grubb DR and Iliades P:
Potential treatment of cardiac hypertrophy and heart failure by
inhibiting the sarcolemmal binding of phospholipase Cbeta1b. Curr
Drug Targets. 11:1032–1040. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Singal T, Dhalla NS and Tappia PS:
Regulation of c-Fos and c-Jun gene expression by phospholipase C
activity in adult cardiomyocytes. Mol Cell Biochem. 327:229–239.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Singal T, Dhalla NS and Tappia PS:
Reciprocal regulation of transcription factors and PLC isozyme gene
expression in adult cardiomyocytes. J Cell Mol Med. 14:1824–1835.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Schnabel P, Mies F, Nohr T, Geisler M and
Böhm M: Differential regulation of phospholipase C-beta isozymes in
cardiomyocyte hypertrophy. Biochem Biophys Res Commun. 275:1–6.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bai H, Wu LL, Xing DQ, Liu J and Zhao YL:
Angiotensin II upregulation of G alpha q/11, phospholipase C beta 3
and extracellular signal-regulated kinase 1/2 via angiotensin II
type 1 receptor. Chin Med J (Engl). 117:88–93. 2004.PubMed/NCBI
|