Introduction
γδ T cells, a small population of unconventional or
innate T cells, are characterized by the expression of γδ receptors
(1). A variety of diseases such as
tuberculosis (2), leprosy
(3), typhoid fever (4), brucellosis (5), tularemia (6), ehrlichiosis (7), malaria (8) and toxoplasmosis (9) can stimulate the expansion of γδ T
cells. γδ T cell receptors (TCRs) have various ligand-binding
sites, allowing γδ T cells to recognize a broad range of pathogenic
agents by common molecular patterns (10). They can kill target cells directly
because of their cytotoxic activity or indirectly by influencing
the activity of other immune cells (11). γδ T cells also exhibit
antigen-presenting ability; in particular, blood Vγ9δ2 T cells are
capable of responding to microbes, tumors as well as cluster of
differentiation (CD)4+ and CD8+ T cells
(12). Owing to these features
shared by innate and adaptive immune cells, γδ T cells are defined
as unconventional T cells (13).
Vγ9δ2 T cells are a major subset in the peripheral blood (14), and can typically be activated by
phosphomonoester antigen and expand in the blood of infected
individuals (15). In previous
studies, researchers have identified various characteristics of
different γδ T cell subsets, particularly those of Vγ9δ2 T cells,
which include immune defense capacity against tumors (16) and antiviral defense ability
(17,18). In infectious diseases, Vγ9δ2 T
cells are capable of recognizing phosphoantigens produced via the
methylerythritol phosphate biochemical pathway of various bacteria,
parasites and fungi, after which these cells become activated
(13). Therefore, Vγ9δ2 T cells
have been considered a promising candidate for immunotherapeutic
drug development and represent a novel therapeutic tool (19). Consequently, a lot of research has
focused on exploiting the potential of Vγ9δ2 T cells, by first
trying to effectively expand this subset of cells. However, the
initial proportion of γδ T cells in human peripheral blood
mononuclear cells (PBMCs) is only 3–5% (20), making their expansion in
vitro difficult, particularly during long-term culture.
There is still no widely accepted method for the
specific expansion of γδ T cells. The different culture methods for
γδ T cell expansion reported globally by different laboratories
since 2000 are summarized in Table
I. To yield abundant γδ T cells with high vitality, most
researchers preferred to expand γδ T cells from PBMCs instead of
purifying them prior to in vitro culture. This is reasonable
because cell-cell contact is necessary (21) for the effective expansion of γδ T
cells and less donor peripheral blood is required. It is well known
that antigenic stimulant and cytokines are essential for γδ T cells
(20). Multiple common antigens
have been used to stimulate expansion of γδ T cells, including
isopentenyl pyrophosphate (IPP) (22), (E)-4-hydroxy-3-methyl-but-2-enyl
pyrophosphate (HMB-PP) (23),
zoledronate (Zometa) (24) and
bromohydrin pyrophosphate (BrHPP) (25). IPP is a natural antigen for Vγ9δ2 T
cells, which can directly stimulate these cells in the absence of
accessory antigen-presenting cells (22). Therefore, IPP has been chosen to
expand γδ T cells in the present study and typical doses (Table I) were used. The other antigens can
also effectively stimulate γδ T cells through various ways; for
example, BrHPP is a synthetic analog of IPP and functions like IPP
(26). Conversely, Zometa leads to
the accumulation of IPP (1) and
HMB-PP acts as an intermediate metabolite of microbial isoprenoid
biosynthesis (1). These two agents
are regarded as indirect or synthetic simulants of γδ T cells. In
addition, interleukin 2 (IL-2) is an essential cytokine for
maintaining T cell expansion and has been widely used in culture of
γδ T cells (27). Considering the
non-specific function of IL-2 in terms of T cell expansion, a
step-wise increase in IL-2 concentration was tested for culture.
Low dose IL-2 (100 U/ml) was used during the early stages of
culture (0–5 days) until γδ T cells reached the logarithmic phase,
allowing the other T cell subsets to die. Subsequently, the dose of
IL-2 was increased to 1,000 U/ml to offer a better expansion
environment for γδ T cells in the logarithmic phase. As a control,
the effects of 1,000 U/ml IL-2 from start to finish were also
detected. To understand the effects of various treatments on cell
status, Vγ9δ2 T cell growth in different culture conditions was
examined.
 | Table I.Culture methods for γδ T cell
expansion since 2000. |
Table I.
Culture methods for γδ T cell
expansion since 2000.
| Author, year | Cell source | Stimulant | Cytokines
(concentration) | Relative increase
in the number of cells (times) | Duration of culture
(days) | Refs. |
|---|
| Duault et
al, 2016 | PBMC | BrHPP | IL-2 (400 U/ml),
IL-33 (100, 500 or 1,000 ng/ml) | 10-12 | 10 | (40) |
| Klimpel et
al, 2003 | PBMC | None | IL-2 (100 U/ml),
IL-15 (1,000 U/ml) | 10-12 | 8 | (41) |
| Kondo et al,
2011 | PBMC | Zoledronate
(Zometa) | IL-2 (1,000
U/ml) | 12-16 | 14 | (24,42) |
| Sato et al,
2009 |
|
|
|
|
|
|
| Barcy et al,
2008 | PBMC | None | IL-2 (20 U/ml) IL-7
(100 U/ml) | 6-8 | 14 | (43) |
| Rincon-Orozco et
al, 2005 | PBMC | IPP | IL-2 (100
U/ml) | – | 14 | (44) |
| Cabillic et
al, 2010 | PBMC | BrHPP/Zoledronate
(Zometa) | IL-2 (400
U/ml) | 12-14 | 14 | (45) |
| Casetti et
al, 2009 | PBMC | IPP | IL-2 (6.5 U/ml),
IL-15 (10 ng/ml), TNF-α (1.7 ng/ml) | 6-8 | 10 | (46) |
| Devilder et
al, 2009 | PBMC |
Phytohemagglutinin | None | 6-8 | – | (47) |
| Tsai et al
2015 | Magnetically
isolated γδ T cells | None | None | – | – | (48) |
| McGill et
al, 2016 | Magnetically
isolated γδ T cells | None | IL-2 (10 U/ml) | 8-10 | – | (49) |
Lentiviral transduction is a highly efficient method
for genetic modification by integrating exogenous genes into host
cells (28). Lentiviruses can
infect non-dividing cells and are regarded as a powerful tool for
basic research (28). HIV-1
pseudotypes with a protein coat of vesicular stomatitis virus
glycoprotein, enables the lentivirus to be transduced into the
majority of mammalian cells (29),
regardless of cell cycle stage. However, artefacts caused by open
reading frame (ORF) disruption or gene activation, can be
introduced into the host cells, in knockdown studies or
over-expression (28).
Furthermore, depending on cell types the modulation efficiency of
gene expression differs, particularly between primary cells and
cell lines (28). Therefore,
experiments should be carefully designed and a proper titration of
the viral vector is necessary. Previously, given the low cell
viability and the limitation of culturing time, transducing
lentiviruses into primary T cells has been an issue for a number of
researchers, especially in cell subsets that account for a small
percentage of cells in PBMCs such as γδ T cells (28). To solve this problem, certain
researchers have chosen to use CH-296 (30), a recombinant human fibronectin
widely used in retroviral transduction, instead of lentiviruses. In
order to achieve high transduction efficiency, the present study
aimed to find the optimum transducing time and multiplicity of
infection (MOI). Considering that the lentivirus generally needs
5–7 days to express completely, to guarantee that γδ T cells could
withstand the damage caused by lentiviral transduction and remain
active, cell growth was examined to find the appropriate time when
the quantity and proportion of Vγ9δ2 T cells in PBMCs reached the
highest level.
In the present study, optimal culture methods for
Vγ9δ2 T cells were investigated in order to satisfy various
experimental purposes. One protocol was confirmed to be suitable
for genetic modification of Vγ9δ2 T cells by lentiviral
transduction. The results provided effective and convenient methods
to expand Vγ9δ2 T cells that fulfill various purposes for
scientific and application studies.
Materials and methods
Donor samples
A total of 5 healthy volunteer donors without a
history of autoimmune or other diseases were recruited from Nanfang
Hospital between January 2016 and April 2018; their age ranged from
20–28 years old, and 3 of them were female and 2 male. The study
was approved by the Ethics Committee of the Southern Medical
University (Guangzhou, China). Prior to sample collection, written
informed consent was obtained from all subjects. PBMCs were
isolated from whole blood collected in K2 EDTA vacuum blood
collection tubes (367525; BD Biosciences, Franklin Lakes, NJ, USA)
using Ficoll-Hypaque (Axis-Shield Diagnostics Ltd., Dundee, UK) by
density gradient centrifugation according to the manufacturer's
protocols.
Flow cytometry
All cell numbers were accurately counted using flow
cytometry according to the manufacturer's protocols. To analyze the
proportion of Vγ9δ2 T cells, PBMCs were collected and stained at
4°C for 20 min in the dark with the following antibodies:
Phycoerythrin (PE)-conjugated anti-γδ TCR antibody (12-9959-42;
eBioscience; Thermo Fisher Scientific Inc., MA, USA) and
allophycocyanin (APC)-conjugated anti-CD8 antibody (17-0087-42;
eBioscience; Thermo Fisher Scientific Inc.). Briefly, the cells
were first collected and counted. Then 1X PBS was used to wash the
cells twice following which, they were incubated with 1,000-fold
diluted antibody in 5% FBS PBS at 4°C for 20 min in the dark. The
cells were then washed twice with 5% FBS PBS and resuspended in 1%
paraformaldehyde (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany),
then immediately analyzed using flow cytometry (Attune NxT; Thermo
Fisher Scientific Inc.) and FlowJo version 7.0 (FlowJo LLC,
Ashland, OR, USA). As Vγ9δ2 T cells are
CD4−CD8− T cells (27), Vγ9δ2 T cells were defined as γδ
T+CD8− cells.
Cell lines and γδ T cells culture
293 cells (cat. no. CRL-11268) were purchased from
the American Type Culture Collection (Manassas, VA, USA) and used
for lentiviral packaging. The 293 cells were cultured in Dulbecco's
modified Eagle's medium (Corning Incorporated, Corning, NY, USA)
containing 10% fetal bovine serum (FBS; Corning Incorporated), 1%
CTS™ GlutaMAX™-I (Gibco; Thermo Fisher Scientific Inc.) and 1%
Minimum Essential Medium Non-Essential Amino Acids Solution (Gibco;
Thermo Fisher Scientific Inc.) at 37°C with 5% CO2.
PBMCs were stimulated with different doses of IPP
(Sigma-Aldrich; Merck KGaA) and recombinant human IL-2 (PeproTech,
Inc., Rocky Hill, NJ, USA) in RPMI 1640 medium (Corning
Incorporated) supplemented with 10% FBS at 37°C with 5%
CO2. Briefly, PBMCs were isolated from whole blood of
healthy donors and the proportion of Vγ9δ2 T cells was detected
using flow cytometry. Subsequently, the cells were divided into
four groups and exposed to different culture conditions, as
described in Table II. During the
16 day culture period, the proportion of Vγ9δ2 T cells was
monitored using flow cytometry and cell growth was observed under a
fluorescence microscope (Nikon Corporation, Tokyo, Japan). γδ T
cells were sorted from PBMCs using antiγδ TCR-labeled MACS magnetic
beads (Miltenyi Biotec GmbH, Bergisch Gladbach, Germany).
| Table II.Expanding γδ T cells using different
combinations of IPP and IL-2. |
Table II.
Expanding γδ T cells using different
combinations of IPP and IL-2.
|
|
| IL-2 (U/ml) |
|---|
|
|
|
|
|---|
| Group | IPP (µg/ml) | Day 0–5 | Day 6–16 |
|---|
| a | 2 | 1,000 | 1,000 |
| b | 2 |
100 | 1,000 |
| c | 5 | 1,000 | 1,000 |
| d | 5 |
100 | 1,000 |
Cell Counting Kit-8 (CCK-8) assay
Cell viability of Vγ9δ2 T cells in different culture
conditions was assessed using the CCK-8 assay. Cells were seeded in
96-well plates (Shanghai ExCell Biology, Inc., Shanghai, China) at
a density of 2.5×105 cells/100 µl/well and 10 µl/well
CCK-8 solution (Dojindo Molecular Technologies, Inc., Kumamoto,
Japan) was added. Cells were incubated at 37°C with 5%
CO2 for 4 h, then the supernatants were transferred to a
new plate and the absorbance was measured at 450 and 630 nm using a
Varioskan® Flash microplate reader (Thermo Fisher
Scientific, Inc.). Cell viability was calculated as the optical
density (OD) value at 450 nm divided by the OD value at the
reference wavelength 630 nm.
Lentiviral packaging and
transduction
The 293 cells were inoculated in T75 culture flasks
(Nalge Nunc International, Penfield, NY, USA) at a density of
2×106 cells and allowed to reach 70–80% confluence the
day prior to infection. The lentiviral plasmid
pHAGE-fullEF1a-MCS-IZsGreen 6 µg, and packaging plasmids psPAX2 4.5
µg and pMD2.G 2.4 µg (all Invitrogen; Thermo Fisher Scientific
Inc.) were transfected into 293 cells using X-tremeGENE™ HP DNA
Transfection Reagent (Roche Applied Science, Mannheim, Germany) for
16 h at 37°C in 5% CO2, according to the manufacturer's
protocol. Following 48–72 h, supernatants containing lentiviral
particles were harvested and filtered through a 0.45-µm filter (EMD
Millipore, Billerica, MA, USA) to remove cell debris. The
supernatants were concentrated by ultracentrifugation at 50,000 × g
at 4°C for 90 min, and the lentiviral particle pellet was
resuspended in 100% FBS and stored at −80°C. The viral titers of
concentrated lentiviral particles were measured by infecting 293
cells seeded at a density of 1×105 cells/well in a
12-well plate (Nalge Nunc International) with viral serial
dilutions. Three days later, the green fluorescent protein (GFP)
expression was detected using flow cytometry and the viral titer
was calculated using the following equation: viral titer (Tu/µl) =
(% GFP+ cells × number of cells transduced)/virus
volume.
For lentiviral transduction, PBMCs stimulated with
IPP and IL-2 were seeded at 1×106 cells/ml in 6-well
plates (Nalge Nunc International), and the concentrated lentivirus
was added at MOI=50 at 37°C and 5% CO2. The transduction
was repeated 2–3 times a day. After 5–7 days culture, the cells
were collected, and the expression of GFP and γδ TCR was detected
using flow cytometry, as described above.
ELISA
Following lentiviral transduction, the cells were
seeded into the 96-well plates (Nalge Nunc International, Penfield,
NY, USA) for 18 h and the cell culture supernatant of lentivirus
infected or uninfected Vγ9δ2 T cells was collected to detect
secreted interferon gamma (IFN-γ) levels of using an ELISA kit
(EH008-96; Shanghai ExCell Biology, Inc., Shanghai, China),
according to the manufacturer's protocol.
Statistical analysis
All data are presented as the means ± standard
deviation and are the results of 3 independent experiments. The
IFN-γ secretion levels amongst different groups were analyzed using
one-way analysis of variance followed by least significant
difference post hoc test. All statistical analyses were performed
using the SPSS version 17.0 (SPSS, Inc., Chicago, IL, USA).
P-values were two-sided and P<0.05 was considered to indicate a
statistically significant difference.
Results
Optimum culture conditions for γδ T
cells
The primary proportion of Vγ9δ2 T cells in PBMCs
(3.14%) was consistent with that observed in a previous study
(26) using flow cytometry
(Fig. 1A). Subsequently, different
doses of IPP and IL-2 at separate stages (Table II) were tested in order to
determine a suitable protocol for culturing Vγ9δ2 T cells.
Following 3–6 days of culture, clonal clusters were observed
(Fig. 1B), suggesting the start of
the logarithmic growth phase. To quantify the viability of Vγ9δ2
cells, CCK-8 assays were performed and the results indicated no
significant difference in cell viability amongst the four different
groups (Fig. 1C). Furthermore, to
determine whether different combinations of IPP and IL-2 could
influence the culture conditions of γδ T cells during the early
stages of culture and encourage entry into the logarithmic growth
phase, the Vγ9δ2 T cell proportion was determined using flow
cytometry (Fig. 1D and E) and the
total number of cells in culture were counted (Fig. 1F). The results from day 4 revealed
that groups a and c, which were exposed to high doses of IL-2
(1,000 U/ml) from the beginning of culture, possessed significantly
higher total cell numbers and a higher proportion of Vγ9δ2 T cells
compared to groups b and d. This in turn also resulted in a higher
Vγ9δ2 T cell count (Fig. 1G).
However, during the early stages of culture (0–5 days) there was no
difference between high and low doses of IPP in terms of the
proportion of Vγ9δ2 T cells (Fig.
1E). On day 16, the time period generally used for ex
vivo T cell long-term culture (31,32),
there was no difference in the proportion of Vγ9δ2 T cells amongst
the four groups (Fig. 1E).
However, group b, where cells were cultured in 2 µg/ml IPP plus 100
U/ml IL-2 at the early stage and 1,000 U/ml IL-2 at the later
stage, demonstrated the highest total cell count at 16 days
(Fig. 1F) and the highest number
of Vγ9δ2 T cells (Fig. 1G;
Table III). These results
indicated that the group b protocol could maintain long-term
culture of Vγ9δ2 T cells, which is necessary for genetic
modification of T cells through lentiviral transduction because
enough time is allowed for expression of exogenous genes. For this
reason, the group b culture protocol was chosen to expand the Vγ9δ2
T cells for genetic modification.
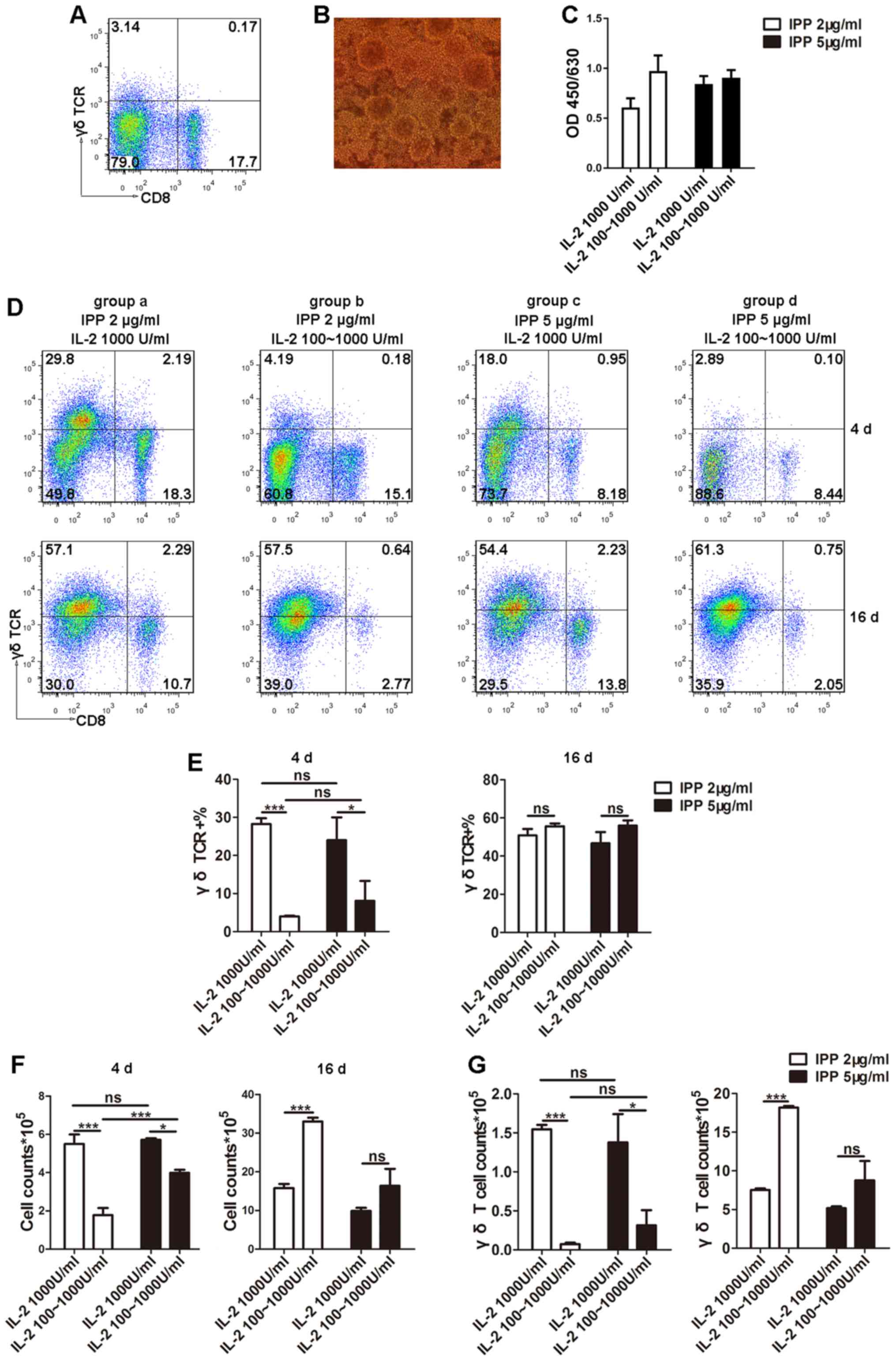 | Figure 1.Optimization of culture conditions
for γδ T cells. (A) Proportion of Vγ9δ2 T cells in PBMCs from
healthy donors was determined using flow cytometry. (B) Following
3–6 days of culture, the clonal clusters of γδ T cells were
observed using microscopy (magnification, ×20). (C) Cell viability
was detected by Cell Counting Kit-8. (D and E) PBMCs were
stimulated with different doses of IPP and IL-2, and the
proportions of Vγ9δ2 T cells were detected using flow cytometry
during the logarithmic growth phase (3–6 days) and the plateau
phase (16 days). (F) Cell counts were determined to reflect the
efficiency of γδ T cell expansion in different culture conditions.
(G) From the total cell count and proportion of Vγ9δ2 T cells, the
Vγ9δ2 T cell counts were calculated. Data are expressed as the mean
± standard deviation. *P<0.05, ***P<0.001. The experiments
were repeated three times. CD, cluster of differentiation; IL-2,
interleukin 2; IPP, isopentenyl pyrophosphate; ns, not significant;
PBMC, peripheral blood mononuclear cell, TCR, T cell receptor. |
| Table III.Summary of Vγ9δ2 T cells harvested
under four different culture conditions and recommended application
for the culture methods. |
Table III.
Summary of Vγ9δ2 T cells harvested
under four different culture conditions and recommended application
for the culture methods.
| Group | a | b | c | d |
|---|
| Initial seeding
density (×105 cells) | 2.00 | 2.00 | 2.00 | 2.00 |
| Day 4 |
| Total
cell count (×105 cells) |
5.50±0.50 |
1.78±0.38 |
5.73±0.08 |
3.99±0.16 |
| Vγ9δ2 T
cell (%) | 28.25±1.55 |
4.02±0.17 |
24±6.00 |
8.10±5.21 |
| Vγ9δ2 T
cell count (×105 cells) |
1.55±0.06 |
0.07±0.02 |
1.38±0.33 |
0.31±0.19 |
| Day 16 |
| Total
cell count (×105 cells) |
15.8±1.10 |
33.00±1.00 |
9.90±0.80 | 16.40±4.40 |
| Vγ9δ2 T
cell (%) |
47.75±6.76 |
55.15±4.39 |
52.50±18.09 | 53.25±4.40 |
| Vγ9δ2 T
cell count (×105 cells) |
7.52±0.40 |
18.18±0.40 |
5.18±1.21 |
8.77±0.75 |
| Application | Short-term
culture | Long-term
culture | Unsuitable | Unsuitable |
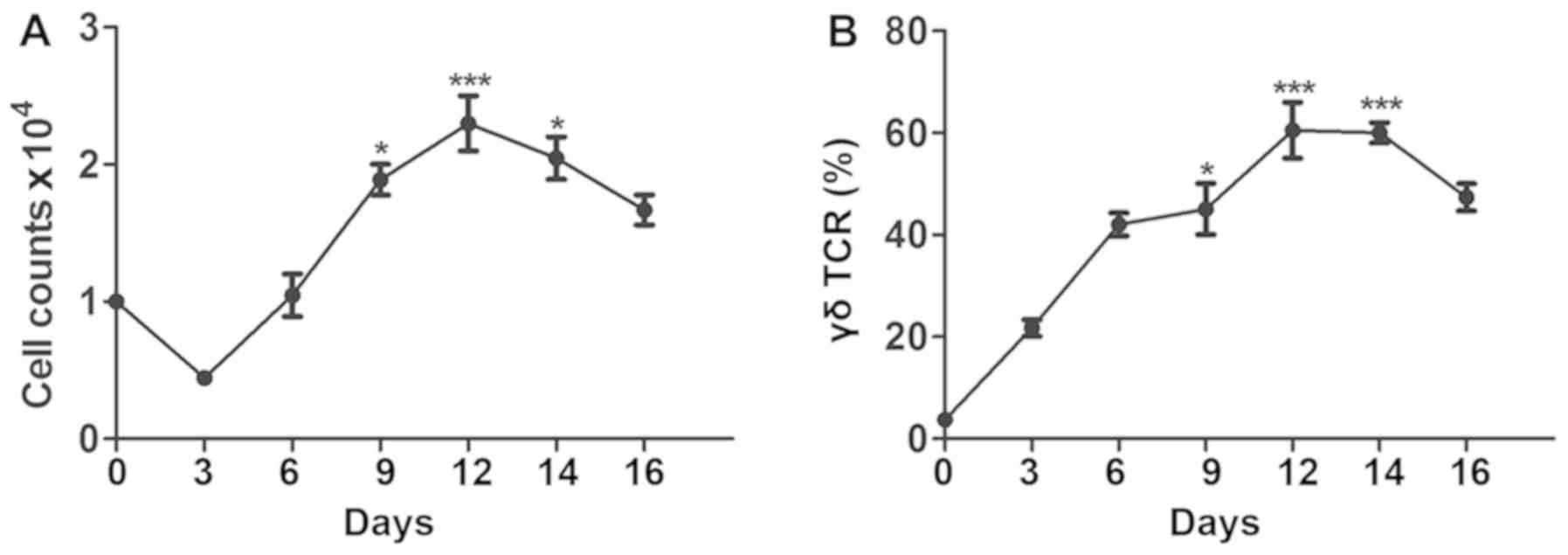
Expansion of γδ T cells under optimum
culture conditions for lentiviral infection
After determining the appropriate conditions for
culturing Vγ9δ2 T cells, cell expansion was monitored in order to
find the optimal time point for lentiviral transduction. Since a
minimum of 5–7 days is required for expression of an exogenous gene
using a recombinant lentivirus, long-term culture of Vγ9δ2 T cells
is necessary. The culture condition with 2 µg/ml of IPP plus 100
U/ml IL-2 at the early stage and 1,000 U/ml IL-2 at the later stage
(group b) was believed to be the most suitable for genetic
modification. Therefore, the relative cell number and proportion of
Vγ9δ2 T cells under this condition were determined every 3 days. As
presented in Fig. 2A, during the
first 3 days of culture, the number of PBMCs decreased greatly and
only 50% of cells survived during this period. In line with this, a
lot of cell debris was observed in the culture under the
microscope. However, the proportion of Vγ9δ2 T cells increased
gradually (Fig. 2B). On day 6, the
cell count returned to the initial level, and the average
proportion of Vγ9δ2 T cells reached ~40%. Clonal clusters were also
observed under the microscope (data not shown). As the cells
entered the logarithmic growth phase (6–12 days), both the cell
count and the proportion of Vγ9δ2 T cells rapidly increased. During
the mid-logarithmic growth phase (8–14 days), the culture contained
the maximum cell count and highest proportion of Vγ9δ2 T cells,
indicating the optimum time to perform subsequent experiments.
Finally, during the later stages of culture (10–20 days), the
plateau phase, the cell count and proportion of Vγ9δ2 T cells
started to decrease.
Lentiviral transduction of γδ T
cells
Lentiviral transduction is an efficient method for
modulating gene expression that has been extensively used in life
science research (28). However,
the transduction protocol should be carefully designed to improve
transduction efficiency, particularly for primary cells. According
to the previous experiment, 8–10 days was selected as the optimum
culture time for γδ T cells prior to treatment. Therefore,
lentivirus was added to γδ T cells at this time point at an MOI=50,
which has been demonstrated to be most suitable for primary T cells
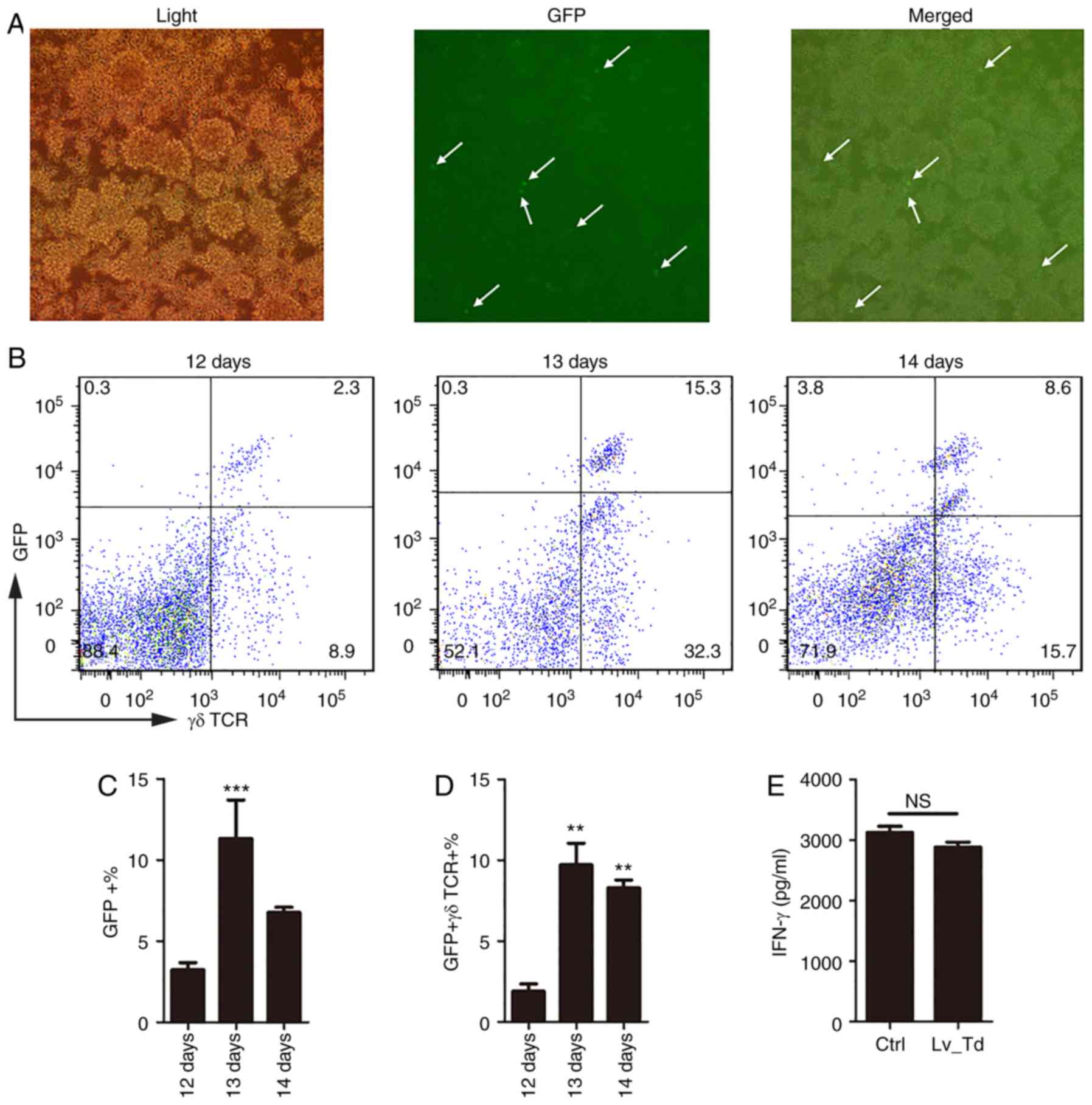
(33). The reporter GFP carried by
the recombinant lentivirus was observed under a fluorescence
microscope (Fig. 3A) and
quantified using flow cytometry (Fig.
3B), 5–7 days following lentiviral transduction. The proportion
of cells expressing both GFP and γδ TCR was daily detected between
days 12–14 of culture. The results confirmed that 5 days following
infection (the 13th day of the entire culturing period), there was
a significantly higher expression of GFP (P<0.001) and γδ T cell
expansion was influenced by the lentivirus but still maintained
high proportion (Fig. 3B). To
further investigate whether these γδ T cells were suitable for
experimental purposes, γδ T cells were magnetically isolated and
IFN-γ secretion was measured to confirm whether γδ T cells were
able to retain their function following lentiviral transduction
(Fig. 3E). According to the ELISA
results (Fig. 3E), cells
transfected with the lentivirus still secreted high levels of IFN-γ
5 days following lentiviral transduction, and there was no
significant difference between the uninfected control group and the
lentiviral transduction group. These results indicated that
lentiviral transduction did not affect the IFN-γ secretion-ability
of γδ T cells. Therefore, lentiviral infection did not appear to
cause damage to cells, and the harvested cells may be used to
perform subsequent functional experiments.
Discussion
Using different doses of IPP and IL-2, a set of
methods for Vγ9δ2 T cell culture were established, for meeting
different experimental requirements. A total of four different
combinations of IPP and IL-2 tested in the present study could
effectively stimulate the expansion of Vγ9δ2 T cells. Notably,
following 16 days of culture, cell counts for the four different
groups increased 3–10 fold and the proportion of Vγ9δ2 T cells was
>50%. To quickly stimulate the expansion of Vγ9δ2 T cells or
increase the rate of Vγ9δ2 T cells entering the logarithmic growth
phase, a high dose of IL-2 (1,000 U/ml) was necessary, whereas 2
µg/ml IPP could be used for reasons of economy. The most suitable
culture method for experiments requiring long-term culture of Vγ9δ2
T cells consisted of adding 2 µg/ml IPP plus 100 U/ml IL-2 at the
early stage, and increasing to 1,000 U/ml IL-2 at the later stage.
Under this culture condition, the largest number of Vγ9δ2 T cells
could be harvested. This culture condition was optimal for
long-term culture of Vγ9δ2 T cells, in order to satisfy the time
restraints of lentiviral transduction. According to the growth
pattern of Vγ9δ2 T cells under this culture condition, 8–10 days
was considered the most appropriate timing to perform lentiviral
transduction. Following 5 days of lentiviral transduction, the
proportion of double-positive GFP and γδ TCR cells reached the
highest level, and the harvested cells were still capable of
secreting IFN-γ.
Effective expansion of Vγ9δ2 T cells is necessary in
order to perform research on this subset of cells. IPP is regarded
as a direct stimulant of Vγ9δ2 T cells and has been used in
previous studies for the expansion of Vγ9δ2 T cells (Table I). In the present study, IPP was
used to stimulate γδ T cells in PBMCs at a high (5 µg/ml) and low
(2 µg/ml) dose, in combination with different concentrations of
IL-2. Since Vγ9δ2 T cells are CD4−CD8− T
cells (27), cells were labeled
with CD8 fluorescent antibody to distinguish the Vγ9δ2 T cells from
other subsets of γδ T cells. By comparing the four different
culture conditions, it was observed that high doses of IL-2 at the
early stage of γδ T cell culture led to an acceleration of γδ T
cell expansion and increased the rate at which cells entered the
logarithmic growth phase. The different doses of IPP did not result
in any difference both in cell count and proportion of Vγ9δ2 T
cells, but the combination of the high dose of IPP with low dose of
IL-2 at the early stage was most effective at stimulating cell
expansion. Studies on the basic function of Vγ9δ2 T cells,
including cytotoxicity, cytokine and chemokine secretion, antigen
presentation, and immunomodulatory activity (34–36)
generally require quick and effective expansion of Vγ9δ2 T cells,
suggesting that initial harvesting of sufficient Vγ9δ2 T cells is
necessary. Therefore, as presented in Table III, the culture method in group a
was regarded as the most suitable for experiments focusing on Vγ9δ2
T cell functional assays, as Vγ9δ2 T cells could expand 2–3 times
during the 6–8-day culture period. The doses of IL-2 and IPP did
not influence the final culture results in terms of Vγ9δ2 T cell
proportion and T cell activity, indicating that all four methods
could stimulate effective expansion of Vγ9δ2 T cells. However, in
group b where 2 µg/ml IPP combined with 100 U/ml IL-2 was used at
the early stage and 1,000 U/ml IL-2 at the later stage, more Vγ9δ2
T cells were harvested compared with the other groups. This may be
because a low dose of IPP exerts less damage on the cells compared
with a high IPP dose (37), while
a low dose of IL-2 at the early stage causes other cell subsets to
die, leading to less cell competition with γδ T cells. Studies
investigating other aspects of Vγ9δ2 T cells, including Vγ9δ2 T
cell growth and genetic modification, generally require a large
number of cells or need longer incubation times. Therefore, for
these experiments, the group b protocol is suitable, as the largest
number of Vγ9δ2 T cells was harvested and long-term culture of
cells was possible (Table III).
Overall the results indicated two effective culture methods for
Vγ9δ2 T cells expansion. For short-term expansion of Vγ9δ2 T cells
(0–6 days), the most suitable culture method was adding 2 µg/ml IPP
and 1,000 U/ml IL-2; while for long-term culture of Vγ9δ2 T cells
to obtain a large number of cells, the most suitable culturing
method was adding 2 µg/ml IPP and 100 U/ml IL-2 at the early stage
and increasing the dose of IL-2 to 1,000 U/ml when the cells
reached logarithmic growth phase (and clonal clusters could be
observed).
Lentiviral transduction is the main method for
genetic modification of primary cells and can be used in adoptive
antigen-specific T cell therapy (38). A previous study demonstrated the
anti-tumor effects of γδ T cells in various cancer types (39), leading to increasing interest in
understanding the potential application of Vγ9δ2 T cells in
adoptive T cell therapy, particularly in metastatic melanoma.
Transferring adoptive T cells to cancer patients with tumor
infiltrating lymphocytes expanded ex vivo may develop
anti-tumor immune responses (36).
γδ T cells with tumor-infiltrating ability are considered to be one
of the most effective lymphocyte subsets because of their ability
to kill tumor cells in various types of cancers, including
leukemia, neuroblastoma and carcinomas (39). For these reasons, genetic
modification of γδ T cells has become a promising research
tool.
Unlike cell lines, primary cells, particularly γδ T
cells, which account for a small percentage of PBMCs, are difficult
to use for lentiviral transduction because of their sensitivity to
cell death and limited proliferative capacity (28). Therefore, the experimental design
should be carefully considered to improve transduction efficiency.
In the present study, based on the different γδ T cell culture
methods, an optimized lentiviral transduction protocol for γδ T
cells was developed by investigating the optimum time for
transduction. Since lentiviral transduction generally requires 5–7
days for expressing an exogenous gene, long-term culture of Vγ9δ2 T
cells was necessary. Therefore, the group b culture method using 2
µg/ml IPP plus 100 U/ml IL-2, at the early stage and 1,000 U/ml
IL-2 at the later stage was selected. Furthermore, by monitoring γδ
T cells, it was identified that the mid-logarithmic growth phase
(8–10 days) was when there was the greatest proportion of γδ T
cells, indicating the most suitable time point to perform
subsequent experiments. MOI=50 is suitable for T cell lentiviral
transduction (35) because of the
high transduction rate and relatively low toxicity. In the present
study, the protocol for lentiviral transduction of γδ T cells
resulted in a relatively high transduction ratio and low damage of
cell function at the same time. On day 5 post-lentiviral
transduction, the transduction rate reached the highest level, the
proportion of Vγ9δ2 T cells remained high and the IFN-γ secretory
function was unchanged.
In conclusion, Vγ9δ2 T cells exhibit distinct
proliferation characteristics during ex vivo culture and studies
using Vγ9δ2 T cells have to take this into account when designing
experimental procedures. In the present study, two culture methods
for expansion of Vγ9δ2 T cells were developed to satisfy various
experimental purposes, using different combinations of IPP and IL-2
doses. However, these methods are only tailored for culturing these
cells in the laboratory. If a larger scale culture of Vγ9δ2 T cells
is required to meet the needs of clinical applications or
bioengineering, further optimization of the methods is required.
According to the long-term culture method and the growth of Vγ9δ2 T
cells, lentiviral transduction was optimized for these cells. In
the future, the genetic modification efficiency and functional
effects of the modified cells may be explored, to better understand
the application of these cells for biological therapy. The present
study provided a set of optimized protocols for Vγ9δ2 T cell
culture to fulfill various research purposes; therefore, it will
greatly promote basic research and clinic application of γδ T
cells.
Acknowledgements
Not applicable.
Funding
The present study was supported by National Natural
Science Foundation of China (grant nos. 81772150 and 81571951),
Guangdong Natural Science Foundation (grant no. 2016A030311001),
Science and Technology Project of Guangdong Province (grant no.
2017A020212007) and Science and Technology Project of Guangzhou
(grant no. 201707010215).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LM and RNW conceived and designed the experiments.
WTH, JHY and RNW performed the experiments. RNW, QW, CYZ and WJX
analyzed the data. LM provided reagents, materials and analysis
tools. LM and RNW drafted the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
the Southern Medical University (Guangzhou, China). Prior to sample
collection, written informed consent was obtained from all
subjects.
Patient consent for publication
All healthy volunteers provided consent for
publication.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Pang DJ, Neves JF, Sumaria N and
Pennington DJ: Understanding the complexity of γδ T-cell subsets in
mouse and human. Immunology. 136:283–290. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Balbi B, Moller DR, Kirby M, Holroyd KJ
and Crystal RG: Increased numbers of T lymphocytes with gamma
delta-positive antigen receptors in a subgroup of individuals with
pulmonary sarcoidosis. J Clin Invest. 85:1353–1361. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Modlin RL, Pirmez C, Hofman FM, Torigian
V, Uyemura K, Rea TH, Bloom BR and Brenner MB: Lymphocytes bearing
antigen-specific gamma delta T-cell receptors accumulate in human
infectious disease lesions. Nature. 339:544–548. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hara T, Mizuno Y, Takaki K, Takada H,
Akeda H, Aoki T, Nagata M, Ueda K, Matsuzaki G and Yoshikai Y:
Predominant activation and expansion of V gamma 9-bearing gamma
delta T cells in vivo as well as in vitro in Salmonella infection.
J Clin Invest. 90:204–210. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bertotto A, Gerli R, Spinozzi F, Muscat C,
Scalise F, Castellucci G, Sposito M, Candio F and Vaccaro R:
Lymphocytes bearing the gamma delta T cell receptor in acute
Brucella melitensis infection. Eur J Immunol. 23:1177–1180. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Poquet Y, Kroca M, Halary F, Stenmark S,
Peyrat MA, Bonneville M, Fournié JJ and Sjöstedt A: Expansion of
Vgamma9 Vdelta2 T cells is triggered by Francisella
tularensis-derived phosphoantigens in tularemia but not after
tularemia vaccination. Infect Immun. 66:2107–2114. 1998.PubMed/NCBI
|
|
7
|
Caldwell CW, Everett ED, McDonald G, Yesus
YW and Roland WE: Lymphocytosis of gamma/delta T cells in human
ehrlichiosis. Am J Clin Pathol. 103:761–766. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Perera MK, Carter R, Goonewardene R and
Mendis KN: Transient increase in circulating gamma/delta T cells
during Plasmodium vivax malarial paroxysms. J Exp Med. 179:311–315.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Scalise F, Gerli R, Castellucci G,
Spinozzi F, Fabietti GM, Crupi S, Sensi L, Britta R, Vaccaro R and
Bertotto A: Lymphocytes bearing the gamma delta T-cell receptor in
acute toxoplasmosis. Immunology. 76:668–670. 1992.PubMed/NCBI
|
|
10
|
Tanaka Y, Sano S, Nieves E, De Libero G,
Rosa D, Modlin RL, Brenner MB, Bloom BR and Morita CT: Nonpeptide
ligands for human gamma delta T cells. Proc Natl Acad Sci USA.
91:8175–8179. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Viey E, Fromont G, Escudier B, Morel Y, Da
Rocha S, Chouaib S and Caignard A: Phosphostim-activated gamma
delta T cells kill autologous metastatic renal cell carcinoma. J
Immunol. 174:1338–1347. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Brandes M, Willimann K and Moser B:
Professional antigen-presentation function by human gammadelta T
Cells. Science. 309:264–268. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lawand M, Déchanet-Merville J and
Dieu-Nosjean MC: Key features of gamma-delta T-cell subsets in
human diseases and their immunotherapeutic implications. Front
Immunol. 8:7612017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Beetz S, Marischen L, Kabelitz D and Wesch
D: Human gamma delta T cells: candidates for the development of
immunotherapeutic strategies. Immunol Res. 37:97–111. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Morita CT, Lee HK, Wang H, Li H, Mariuzza
RA and Tanaka Y: Structural features of nonpeptide prenyl
pyrophosphates that determine their antigenicity for human gamma
delta T cells. J Immunol. 167:36–41. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kabelitz D, Wesch D, Pitters E and Zöller
M: Potential of human gammadelta T lymphocytes for immunotherapy of
cancer. Int J Cancer. 112:727–732. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Poccia F, Agrati C, Martini F, Capobianchi
MR, Wallace M and Malkovsky M: Antiviral reactivities of gammadelta
T cells. Microbes Infect. 7:518–528. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Poccia F, Battistini L, Cipriani B,
Mancino G, Martini F, Gougeon ML and Colizzi V:
Phosphoantigen-reactive Vgamma9Vdelta2 T lymphocytes suppress in
vitro human immunodeficiency virus type 1 replication by
cell-released antiviral factors including CC chemokines. J Infect
Dis. 180:858–861. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hiasa A, Nishikawa H, Hirayama M, Kitano
S, Okamoto S, Chono H, Yu SS, Mineno J, Tanaka Y, Minato N, et al:
Rapid alphabeta TCR-mediated responses in gammadelta T cells
transduced with cancer-specific TCR genes. Gene Ther. 16:620–628.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Puan KJ, Jin C, Wang H, Sarikonda G, Raker
AM, Lee HK, Samuelson MI, Märker-Hermann E, Pasa-Tolic L, Nieves E,
et al: Preferential recognition of a microbial metabolite by human
Vgamma2Vdelta2 T cells. Int Immunol. 19:657–673. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Witkin AJ and Hsu J: Induction of gamma
delta T-lymphocyte effector functions by bisphosphonate zoledronic
acid in cancer patients in vivo. Retina. 102:2310–2311. 2013.
|
|
22
|
Poquet Y, Constant P, Halary F, Peyrat MA,
Gilleron M, Davodeau F, Bonneville M and Fournié JJ: A novel
nucleotide-containing antigen for human blood gamma delta T
lymphocytes. Eur J Immunol. 26:2344–2349. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Eberl M, Engel R, Beck E and Jomaa H:
Differentiation of human gamma-delta T cells towards distinct
memory phenotypes. Cell Immunol. 218:1–6. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kondo M, Izumi T, Fujieda N, Kondo A,
Morishita T, Matsushita H and Kakimi K: Expansion of human
peripheral blood γδ T cells using zoledronate. J Vis Exp. 2:6–11.
2011.
|
|
25
|
Espinosa E, Belmant C, Pont F, Luciani B,
Poupot R, Romagné F, Brailly H, Bonneville M and Fournié JJ:
Chemical synthesis and biological activity of bromohydrin
pyrophosphate, a potent stimulator of human gamma delta T cells. J
Biol Chem. 276:18337–18344. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gertner-Dardenne J, Bonnafous C, Bezombes
C, Capietto AH, Scaglione V, Ingoure S, Cendron D, Gross E, Lepage
JF, Quillet-Mary A, et al: Bromohydrin pyrophosphate enhances
antibody-dependent cell-mediated cytotoxicity induced by
therapeutic antibodies. Blood. 113:4875–4884. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hintz M, Reichenberg A, Altincicek B, Bahr
U, Gschwind RM, Kollas AK, Beck E, Wiesner J, Eberl M and Jomaa H:
Identification of (E)-4-hydroxy-3-methyl-but-2-enyl pyrophosphate
as a major activator for human gammadelta T cells in Escherichia
coli. FEBS Lett. 509:317–322. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shearer RF and Saunders DN: Experimental
design for stable genetic manipulation in mammalian cell lines:
Lentivirus and alternatives. Genes Cells. 20:1–10. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Naldini L, Blömer U, Gallay P, Ory D,
Mulligan R, Gage FH, Verma IM and Trono D: In vivo gene delivery
and stable transduction of nondividing cells by a lentiviral
vector. Science. 272:263–267. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Heemskerk MHM, Hagedoorn RS, van der Hoorn
MAWG, van der Veken LT, Hoogeboom M, Kester MG, Willemze R and
Falkenburg JH: Efficiency of T-cell receptor expression in
dual-specific T cells is controlled by the intrinsic qualities of
the TCR chains within the TCR-CD3 complex. Blood. 109:235–243.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Carlens S, Gilljam M, Chambers BJ, Aschan
J, Guven H, Ljunggren HG, Christensson B and Dilber MS: A new
method for in vitro expansion of cytotoxic human CD3-CD56+ natural
killer cells. Hum Immunol. 62:1092–1098. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
da Silva RF, Petta CA, Derchain SF, Alici
E and Guimarães F: Up-regulation of DNAM-1 and NKp30, associated
with improvement of NK cells activation after long-term culture of
mononuclear cells from patients with ovarian neoplasia. Hum
Immunol. 75:777–784. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhou CY, Wen Q, Chen XJ, Wang RN, He WT,
Zhang SM, Du XL and Ma L: Human CD8(+) T cells transduced with an
additional receptor bispecific for both Mycobacterium tuberculosis
and HIV-1 recognize both epitopes. J Cell Mol Med. 20:1984–1998.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Dudley ME, Wunderlich JR, Robbins PF, Yang
JC, Hwu P, Schwartzentruber DJ, Topalian SL, Sherry R, Restifo NP,
Hubicki AM, et al: Cancer regression and autoimmunity in patients
after clonal repopulation with antitumor lymphocytes. Science.
298:850–854. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gentles AJ, Newman AM, Liu CL, Bratman SV,
Feng W, Kim D, Nair VS, Xu Y, Khuong A, Hoang CD, et al: The
prognostic landscape of genes and infiltrating immune cells across
human cancers. Nat Med. 21:938–945. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Dudley ME, Wunderlich JR, Yang JC, Sherry
RM, Topalian SL, Restifo NP, Royal RE, Kammula U, White DE,
Mavroukakis SA, et al: Adoptive cell transfer therapy following
non-myeloablative but lymphodepleting chemotherapy for the
treatment of patients with refractory metastatic melanoma. J Clin
Oncol. 23:2346–2357. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Eberl M, Hintz M, Reichenberg A, Kollas
AK, Wiesner J and Jomaa H: Microbial isoprenoid biosynthesis and
human gammadelta T cell activation. FEBS Lett. 544:4–10. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
van der Veken LT, Hagedoorn RS, van Loenen
MM, Willemze R, Falkenburg JHF and Heemskerk MHM: Alphabeta T-cell
receptor engineered gammadelta T cells mediate effective
antileukemic reactivity. Cancer Res. 66:3331–3337. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Silva-Santos B, Serre K and Norell H: γδ T
cells in cancer. Nat Rev Immunol. 15:683–691. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Duault C, Franchini DM, Familliades J,
Cayrol C, Roga S, Girard JP, Fournié JJ and Poupot M: TCRVγ9 γδ T
Cell Response to IL-33: A CD4 T Cell-Dependent Mechanism. J
Immunol. 196:493–502. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Klimpel GR, Matthias MA and Vinetz JM:
Leptospira interrogans activation of human peripheral blood
mononuclear cells: Preferential expansion of TCR gamma delta+ T
cells vs TCR alpha beta+ T cells. J Immunol. 171:1447–1455. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Sato K, Kondo M, Sakuta K, Hosoi A, Noji
S, Sugiura M, Yoshida Y and Kakimi K: Impact of culture medium on
the expansion of T cells for immunotherapy. Cytotherapy.
11:936–946. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Barcy S, De Rosa SC, Vieira J, Diem K,
Ikoma M, Casper C and Corey L: Gamma delta+ T cells involvement in
viral immune control of chronic human herpesvirus 8 infection. J
Immunol. 180:3417–3425. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Rincon-Orozco B, Kunzmann V, Wrobel P,
Kabelitz D, Steinle A and Herrmann T: Activation of V gamma 9V
delta 2 T cells by NKG2D. J Immunol. 175:2144–2151. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Cabillic F, Toutirais O, Lavoué V, de La
Pintière CT, Daniel P, Rioux-Leclerc N, Turlin B, Mönkkönen H,
Mönkkönen J, Boudjema K, et al: Aminobisphosphonate-pretreated
dendritic cells trigger successful Vgamma9Vdelta2 T cell
amplification for immunotherapy in advanced cancer patients. Cancer
Immunol Immunother. 59:1611–1619. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Casetti R, Agrati C, Wallace M, Sacchi A,
Martini F, Martino A, Rinaldi A and Malkovsky M: Cutting edge:
TGF-beta1 and IL-15 Induce FOXP3+ gammadelta regulatory T cells in
the presence of antigen stimulation. J Immunol. 183:3574–3577.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Devilder MC, Allain S, Dousset C,
Bonneville M and Scotet E: Early triggering of exclusive IFN-gamma
responses of human Vgamma9Vdelta2 T cells by TLR-activated myeloid
and plasmacytoid dendritic cells. J Immunol. 183:3625–3633. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Tsai CY, Liong KH, Gunalan MG, Li N, Lim
DS, Fisher DA, MacAry PA, Leo YS, Wong SC, Puan KJ, et al: Type I
IFNs and IL-18 regulate the antiviral response of primary human γδ
T cells against dendritic cells infected with Dengue virus. J
Immunol. 194:3890–3900. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
McGill JL, Rusk RA, Guerra-Maupome M,
Briggs RE and Sacco RE: Bovine gamma delta T Cells Contribute to
exacerbated IL-17 production in response to co-infection with
Bovine RSV and Mannheimia haemolytica. PLoS One. 11:e01510832016.
View Article : Google Scholar : PubMed/NCBI
|