Introduction
End-stage renal disease (ESRD) is generally
diagnosed when the glomerular filtration rate (GFR) is <15
ml/min/1.73 m2, which can be similar to that in uremia
(1,2). ESRD patients have two treatment
options: Long-term dialysis or kidney transplantation. Owing to
problems arising from donors, matching type and race, numerous
patients have to undergo dialysis treatments, including
hemodialysis (HD) or peritoneal dialysis (PD), as an alternative to
renal replacement therapy (3).
Compared with HD, PD has been associated with notably better or at
least similar survival of patients, lower cost and is superior to
retaining residual renal function (4,5);
however, few patients are reluctant to utilize PD for extended
periods due to the progression of peritoneal fibrosis. Thus,
applying PD for >10 years is exceedingly rare (6). The occurrence of peritoneal fibrosis
is mainly due to the composition of PD solution (high-glucose
content, low pH, elevated osmolality, increased concentration of
lactate and the degradation products of glucose), uremic toxins,
refractory or recurrent infectious peritonitis, and chronic
inflammation (7–9). Therefore, understanding the origin of
peritoneal fibrosis is critical to maintaining the integrity of the
peritoneal membrane and to prolong PD treatment in such patients.
At present, no studies have investigated this; thus, the underlying
mechanism is unclear.
Uric acid (UA) is the metabolic end product of
nucleic acid purine (including nucleic acids in food), which is
mainly eliminated via renal excretion. The levels of UA are an
indispensable marker in the detection of rare inborn errors of
metabolism of the degradation system of purine nucleotides
(10). Hyperuricemia has been
frequently observed in patients with chronic kidney disease (CKD)
due to decreases in GFR. In numerous, serum UA has been reported as
a predictor of the progression of renal disease, as >90% of
hyperuricemia is due to the impaired renal excretion of UA
(11,12). Thus, whether high levels of blood
UA have an effect on peritoneal fibrosis, as well as reducing the
levels of UA alleviates peritoneal fibrosis, requires further
investigation. On this basis, the present study employed peritoneal
mesothelial cells to determine the role of serum UA.
Materials and methods
Reagents
Fetal bovine serum (FBS; 10%), Dulbecco's Modified
Eagle's medium (DMEM)/F12 medium, 1% (v/v) penicillin and
streptomycin were obtained from Invitrogen (Thermo Fisher
Scientific, Inc., Waltham, MA, USA). UA, Triton X-100, MTT,
dimethyl sulfoxide (DMSO), DAPI, Hoechst 33342 were purchased from
Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). Anti-phosphorylated
(p)-P38, anti-total (t)-P38, anti-mothers against decapentaplegic
homolog 3 (Smad3), anti-p-Smad3, anti-E-cadherin, anti-transforming
growth factor (TGF)-β1, anti-α-smooth muscle actin (SMA),
anti-vimentin and anti-β-actin were purchased from Santa Cruz
Biotechnology, Inc. (Dallas, TX, USA). An enhanced
chemiluminescence kit was also purchased (Pierce; Thermo Fisher
Scientific, Inc.).
Cell culture
The cell line employed in the present study was the
human peritoneal mesothelial cell line, HMrSV5, which was obtained
from the Cell Culture Centre (Chinese Academy of Medical Sciences,
Beijing, China). HMrSV5 cells were cultured in DMEM/F12 medium
containing 10% FBS at 37°C in a humidified environment with 5%
CO2. The cells were dissociated using 0.2% trypsin plus
0.02% EDTA solution at 37°C and sub-cultured once in 4 days. The
cells were then rinsed with PBS, the medium was exchanged and the
cells were cultured further.
MTT assay
Cell viability was determined by an MTT assay.
Briefly, HMrSV5 cells in the logarithmic growth phase were cultured
at a density of 5×103 cells/well in 96-well plates. The
cells were exposed to various concentrations (0, 200, 400, 600,
800, 1,000, 1,200 and 1,500 µmol/l) of UA for 24, 48 and 72 h. For
analysis, 20 µl of MTT substrate was added to each well and the
plates were incubated for an additional 4 h at 37°C with 5%
CO2. The medium was removed, and the cells were
solubilized in 150 µl DMSO to dissolve the colored formazan
crystals for 15 min. The absorbance of each well was recorded on a
microplate spectrophotometer at 490 nm using a microplate reader
(ASYS Expert Plus, ASYS Hitech GmbH, Eugendorf, Austria). Relative
cellular growth was determined using the ratio of average
absorbance in treatment cells vs. the average absorbance in control
cells. The cell viability was calculated as the ratio of optical
densities. UA concentrations of 200 and 400 µmol/l were used in the
experiments below.
Terminal
deoxynucleotidyl-transferase-mediated dUTP nick end labelling
(TUNEL) assay
TUNEL assays were performed using an
ApopTag® kit (Merck KGaA) according to the
manufacturer's protocols. HMrSV5 cells were fixed with 4%
paraformaldehyde solution for 30 min and then blocked using PBS
buffer containing 2% H2O2 for 5 min at room
temperature. Following washing with PBS, cells were treated with
0.1% Triton X-100 on ice for 2 min to increase the permeability of
the cell membrane. Subsequently, the cells were incubated with 50
µl TUNEL reagent in a humidified chamber for 1 h at 37°C. To detect
the nuclei, the cells were incubated with DAPI for 2 min at room
temperature in the dark and mounted in neutral resin. An
epifluorescent microscope (FV300, Olympus Corporation, Tokyo,
Japan) was used to detect apoptotic cells (magnification, ×40). A
total of 3 randomly selected number of fields were examined.
Immunofluorescence staining
HMrSV5 cells were seeded in 6-well plates. After the
indicated treatment, the cells were fixed with 4% paraformaldehyde
for 15 min at 0°C, permeabilized with 0.2% Triton X-100 in PBS for
5 min, and blocked with 50 mg/ml bovine serum albumin
(Sigma-Aldrich) in PBS at room temperature for 30 min. Following
incubation with primary antibodies against vimentin (1:200;
sc-5565; Santa Cruz Biotechnology, Inc.) and E-cadherin (1:200;
sc-7870; Santa Cruz Biotechnology, Inc.) overnight at 4°C, the
cells were washed three times with PBS. The
tetramethylrhodamine-conjugated goat anti-mouse IgG antibody
(1:100; sc-57606 Santa Cruz Biotechnology, Inc.) was applied for
0.5 h at room temperature. The cell nuclei were counterstained with
Hoechst 33342 for 15 min at room temperature and the images were
captured using a fluorescence microscope (magnification, ×10;
excitation wavelength, 430 nm; FV300, Olympus Corporation).
Western blot analysis
HMrSV5 cells were lysed with
radioimmunoprecipitation assay lysis buffer (Beyotime Institute of
Biotechnology, Haimen, China) on ice for 30 min and centrifuged at
15,000 × g at 4°C for 30 min. The supernatant was collected, and
protein concentrations were determined using a bicinchoninic acid
assay kit (Pierce; Thermo Fisher Scientific, Inc.). Equal protein
lysate samples (10 µg) were separated via 12% SDS-PAGE and then
transferred to nitrocellulose membranes. The membranes were blocked
at room temperature for 2 h in TBS with 0.1% Tween-20 (TBS-T)
containing 5% skim milk, followed by incubation with primary
antibodies at 4°C overnight. The primary antibodies used in the
present study were: Anti-p-P38 (1:400; sc-7973), anti-P38 (1:400;
sc-7972), anti-Smad3 (1:400; sc-101154), anti-p-Smad3 (1:400;
sc-517575), anti-E-cadherin (1:400; sc-7870), anti-TGF-β1 (1:400;
sc-146), anti-α-SMA (1:400; sc-324317), anti-vimentin (1:400;
sc-5565) and anti-β-actin (1:400; sc-47778). The membrane was then
incubated with horseradish peroxidase-conjugated secondary
antibodies (sc-2039 and sc-2094; Santa Cruz Biotechnology, Inc.)
for 2 h at room temperature. Blots were developed using an enhanced
chemiluminescence kit (Pierce; Thermo Fisher Scientific, Inc.).
Relative expression was quantified using Quantity One Software
(version 643274, rev. A; Bio-Rad Laboratories, Inc., Hercules, CA,
USA).
Reverse transcription
semi-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was extracted from HMrSV5 cells using
TRIzol® RNA isolation (Gibco; Thermo Fisher Scientific,
Inc.) and purified with DNase I (Invitrogen; Thermo Fisher
Scientific, Inc.). RNA (1 µg) was reverse transcribed into the
first strand of cDNA using a SuperScript Reverse Transcriptase
system (Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocols. qPCR reactions were performed using a
SuperScript™ First-Strand Synthesis kit (Invitrogen; Thermo Fisher
Scientific, Inc.) and a Gene Amp PCR system 9700 (PerkinElmer,
Inc., Waltham, MA, USA). qPCR was performed with the following
thermocycling conditions: First at 95°C for 3 min, 30 cycles at
95°C for 30 sec, with different annealing temperatures (55°C for
TGF-β1; 60°C for GAPDH) for 30 sec, at 72°C for 30 sec, and
extension at 72°C for 10 min. The amplified products were separated
by electrophoresis on a 2% agarose gel and visualized by ethidium
bromide staining for 30 min at room temperature. Each product was
visualized after separation and GAPDH was used to normalize the
mRNA expression levels. Image density was quantified with a
FluoroImager SI (GE Healthcare, Chicago, IL, USA). The sequences of
TGF-β1 and GAPDH genes were obtained from the GenBank database
(https://www.ncbi.nlm.nih.gov/genbank/), and specific
primers were designed over an exon-exon junction with Primer
Premier 5.0 (Premier Biosoft International, Palo Alto, CA,
USA).
The primers used for amplification of TGF-β1 were as
follows: Forward, 5′-AACATGATCGTGCGCTCTGCAAGTGCAGC-3′ and reverse,
5′-AAGGAATAGTGCAGACAGGCAGGA-3′; GAPDH, forward,
5′-CCTTCATTGACCTCAACTAC-3′ and reverse,
5′-CCAAAGTTGTCATGGATG-3′.
Statistical analysis
Statistical analysis was performed using SPSS 19.0
(IBM Corp., Armonk, NY, USA). All values were expressed as the mean
± standard deviation, which were obtained from at least three
independent experiments for each condition. Statistical
significance was determined by performing one-way analyses of
variance followed by Tukey's multiple comparison test. P<0.05
was considered to indicate a statistically significant difference,
and P<0.01 was considered to indicate a highly statistically
significant difference.
Results
Effects of UA on the cell viability of
peritoneal mesothelial cells
In order to examine the cytotoxic effects of UA on
peritoneal mesothelial cells, HMrSV5 cells were incubated with UA
at various concentrations (0–1,500 µmol/l) for 24–72 h, and the
cell viability was determined using an MTT assay. As presented in
Fig. 1A, UA-treated HMrSV5 cells
(≤400 µmol/l) did not exhibit significant reductions in cell
viability compared with the control (P>0.05); however, above
this concentration, the cell viability significantly decreased
(P<0.01). In particular, UA of 200 or 400 µmol/l did not notably
alter the cell viability at different durations as demonstrated in
Fig. 1B. To further analyze
UA-induced apoptosis, a TUNEL experiment was conducted. The results
revealed that ≤400 µmol/l UA is almost non-toxic to cells, whereas
>400 µmol/l of UA was observed to increase the cytotoxicity of
the cells (Fig. 1C). Similar
results were acquired via analysis under a light microscope
(Fig. 1D). Following treatment
with 0–400 µmol/l UA, apoptosis was not notably induced. In the
follow-up epithelial-mesenchymal transition (EMT) experiments, UA
concentrations of 200 and 400 µmol/l were used.
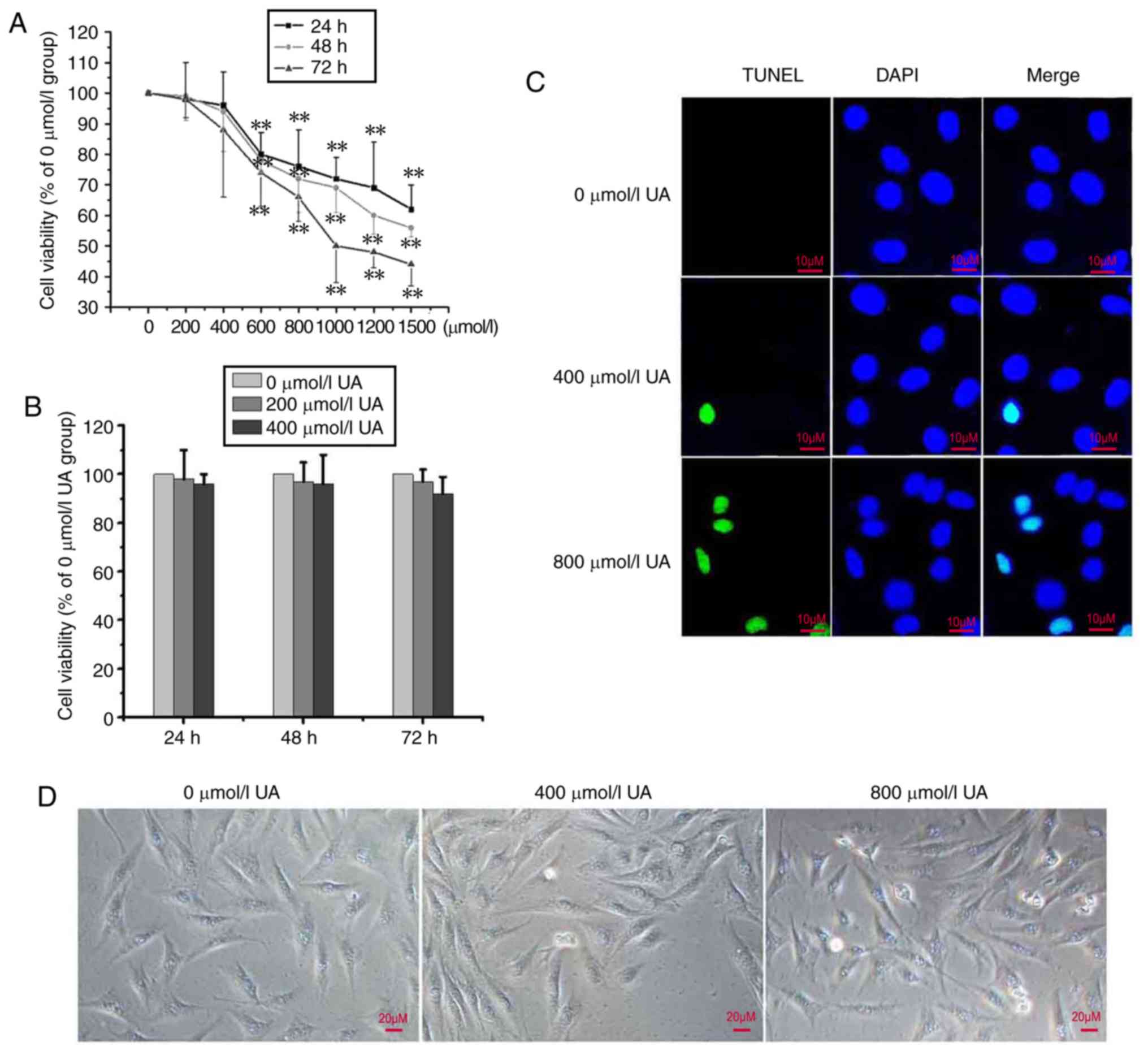 | Figure 1.UA inhibits cell viability in
peritoneal mesothelial cells. (A) HMrSV5 cells were treated with
0–1, 500 µmol/l of UA for 24, 48 and 72 h, and the cell viability
was detected via an MTT assay (n=3). Values are presented as the
mean ± standard deviation; **P<0.01 vs. 0 µmol/l group. (B)
Cells were treated with 0, 200 and 400 µmol/l of UA for 24, 48 and
72 h, and cell viability was detected using an MTT assay. (C) Cells
were treated with 0, 400 and 800 µmol/l of UA for 48 h, and the
TUNEL-stained cells were detected. Scale bar=10 µm. (D) Cells were
treated as aforementioned, and the cells were observed under a
light microscope. Scale bar, 20 µm. Results are presented as the
mean ± standard deviation. TUNEL, terminal
deoxynucleotidyl-transferase-mediated dUTP nick end labelling; UA,
uric acid. |
Effects of UA on EMT of peritoneal
mesothelial cells
A previous study demonstrated that EMT contributes
to the development of peritoneal fibrosis (13). In order to understand whether UA is
involved in the regulation of peritoneal fibrosis, the effects of
UA on the EMT of cultured human peritoneal mesothelial cells were
examined. As demonstrated in Fig.
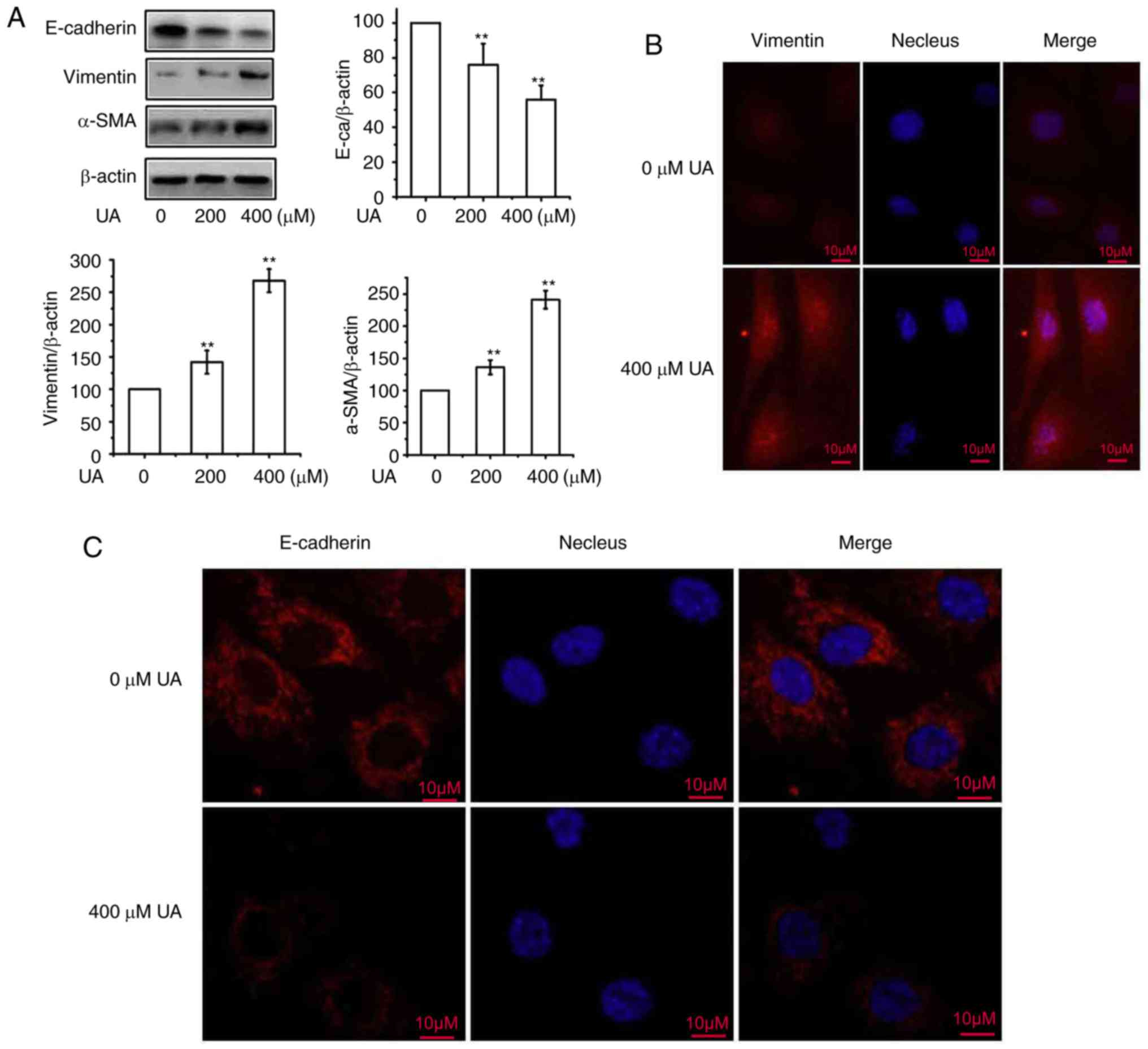
2A, compared with the control, treatment with UA significantly
upregulated the expression of α-SMA and vimentin, and significantly
downregulated the expression of E-cadherin in a dose-dependent
manner, following treatment with 0, 200 and 400 µmol/l UA for 48 h.
The effects of UA-induced EMT of HMrSV5 cells were subsequently
assessed via immunofluorescence. As presented in Fig. 2B and C, it was observed that
treatment with UA led to the increased the expression of vimentin
and decreased E-cadherin expression, indicating the characteristics
of a mesenchymal phenotype. These results confirm that UA acts
positively to regulate EMT in peritoneal mesothelial cells.
Effects of UA on TGF-β1/Smad3 pathway
of peritoneal mesothelial cells
It is well known that activation of TGF-β1/Smad3
signaling is critical for the development of EMT (14). Thus, whether UA may regulate EMT
via the activation of the signaling pathway in HMrSV5 cells was
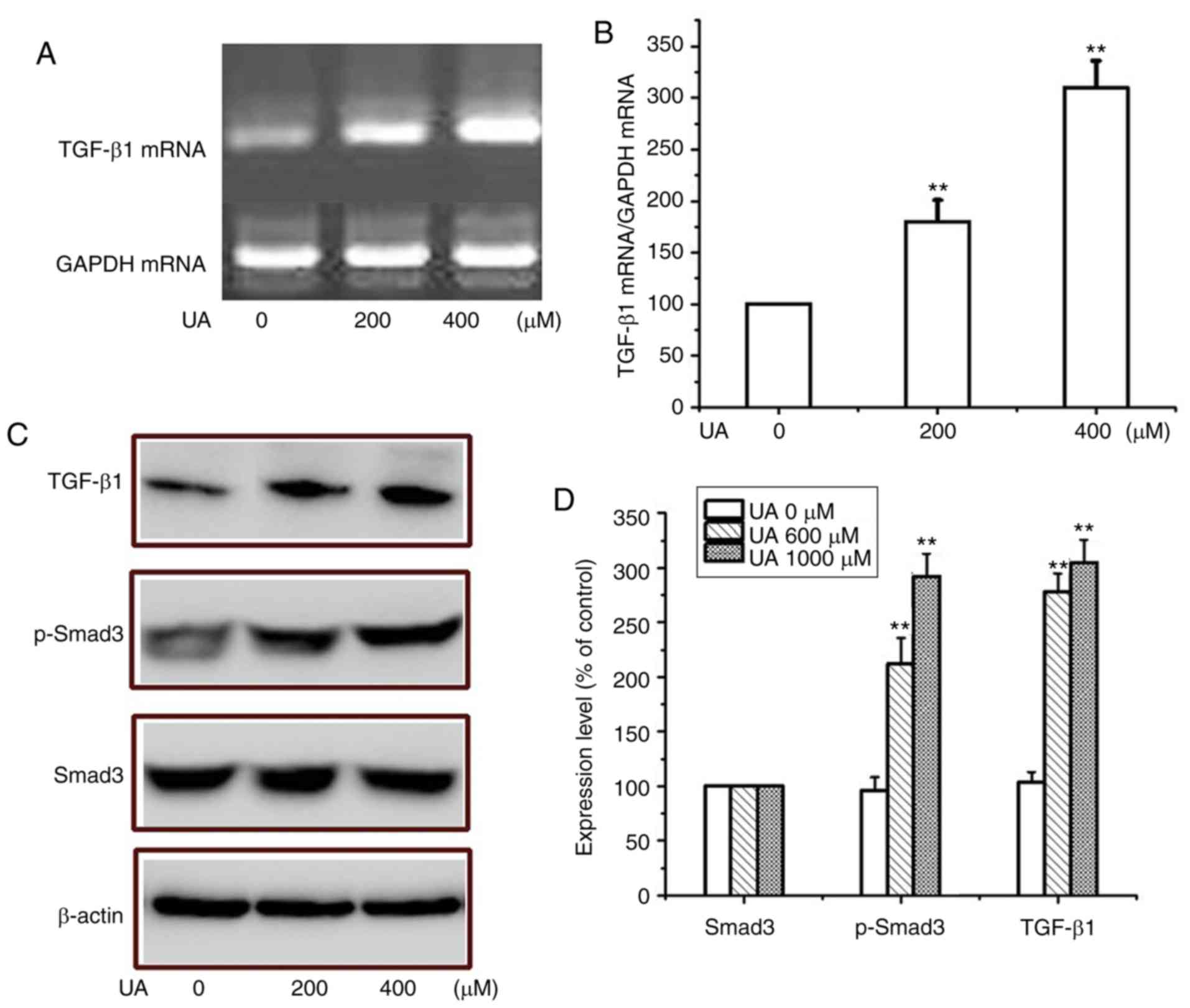
examined. To demonstrate whether UA serves a role in regulating the
activation of the TGF-β1/Smad3 pathway during EMT, the expression
levels of TGF-β1 and Smad3 were determined. As demonstrated in
Fig. 3A and B, following UA
treatment, the expression of TGF-β1 was significantly increased
compared with the control group, which may be associated with
upregulation of α-SMA expression and downregulation of E-cadherin.
Similarly, as presented in Fig. 3C and
D, UA treatment promoted the phosphorylation of Smad3 in a
dose-dependent manner; however, the levels of total Smad3 did not
change significantly. Therefore, UA may serve a critical role in
mediating the activation of TGF-β1/Smad3 in peritoneal mesothelial
cells undergoing EMT.
Effects of UA on mitogen-activated
protein kinase (MAPK)/P38 pathway of peritoneal mesothelial
cells
P38 MAPKs serve important roles in the invasion and
fibrosis of tumor cells. It has been suggested that P38 MAPK
signaling regulates EMT (15).
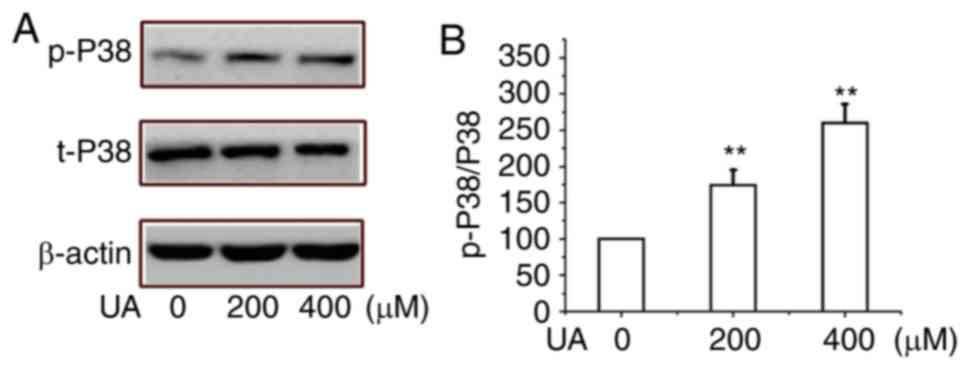
Thus, the effects of UA-induced EMT were determined by
investigating the important pathway of MAPK-p38 proteins in HMrSV5
cells. As demonstrated in Fig. 4A and
B, the exposure of peritoneal mesothelial cells to UA promoted
the expression of p-P38, which indicated that UA treatment
activated the pathway in a dose-dependent manner; however, the
expression of t-P38 did not change significantly. These findings
suggest that UA treatment markedly induced EMT and regulated
MAPK-P38 signaling pathway in HMrSV5 cells.
Discussion
Recent studies have demonstrated that EMT of
peritoneal epithelial cells may be one of the mechanisms underlying
peritoneal fibrosis, in which epithelial cells lose their
characteristic cell-cell junctions, polarized cell-surface
molecules and apical-basolateral polarity, while acquiring
properties typical of mesenchymal cells and increased migratory,
invasive, and fibrogenic potentials (16,17).
UA has been reported to serve a pathogenic role in CKD; however,
its effect on EMT or peritoneal fibrosis has not yet been
elucidated. Therefore, in the present study, the effects of UA on
the EMT of peritoneal mesothelial cells were investigated. It was
demonstrated that UA induced downregulation of E-cadherin, and
upregulation of α-SMA and vimentin in human peritoneal mesothelial
cells in vitro. Overall, it was proposed that UA promotes
the EMT process of peritoneal mesothelial cells. Previous studies
have reported that oxidative stress serves an important role in the
development of fibrosis under various pathological conditions,
while few studies suggest that UA may induce oxidative stress, and
be a mediator of EMT (18,19). UA may indirectly promote the
development of EMT by inducing oxidative stress. Conversely, UA
induces the release of cellular inflammatory mediators, including
monocyte chemoattractant protein-1, cyclooxygenase-2 and tumor
necrosis factor-α (20). Notably,
it is well established that these pro-inflammatory cytokines are
strongly associated with the occurrence of EMT (21,22).
Furthermore, in the present study, signaling
cascades involved in the effects of UA-induced EMT were
investigated. Emerging evidence has demonstrated that TGF-β1/Smad3
signaling serves a pivotal role in EMT (23). The results also indicated that
TGF-β1 is crucial for the initiation of EMT, and the TGF-β1/Smad3
pathway is regulated by UA, which can directly promote EMT.
Numerous studies have previously demonstrated that the activity of
the MAPK pathway is required for TGF-β1-induced EMT in mammary
epithelial cells (24,25). The present study reported that
exposure of peritoneal mesothelial cells to UA promoted the
expression of p-P38, which suggested that UA-induced EMT is closely
associated with the activation of P38.
In conclusion, the present study investigated
whether UA promotes TGF-β1-induced EMT, as well as the potential
pathways involved in this crucial transition of peritoneal
mesothelial cells. The findings demonstrate a potential association
between UA and the progression of EMT in peritoneal mesothelial
cells. Future investigation is required to further understand the
role of UA in peritoneal fibrosis during long-term peritoneal
dialysis. The potential for alleviating peritoneal fibrosis by
lowering the serum UA levels is promising, and thus should be
evaluated in well-designed randomized controlled studies in the
future.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
C-YD was the guarantor of integrity for the entire
study and responsible for the study concepts, the definition of
intellectual content, the clinical studies, and statistical
analysis. YL was responsible for the study concepts, the study
design, literature research, statistical analysis and manuscript
preparation, editing and review. JH was responsible for aquisition,
analysis and interpretation of data, the definition of intellectual
content and manuscript preparation, editing and review. KW
performed experimental studies and data analysis. C-YZ performed
data acquisition. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Grams ME, Li L, Greene TH, Tin A, Sang Y,
Kao WH, Lipkowitz MS, Wright JT, Chang AR, Astor BC and Appel LJ:
Estimating time to ESRD using kidney failure risk equations:
Results from the African American study of kidney disease and
hypertension (AASK). Am J Kidney Dis. 65:394–402. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jun M, Turin TC, Woodward M, Perkovic V,
Lambers Heerspink HJ, Manns BJ, Tonelli M and Hemmelgarn BR:
Assessing the validity of surrogate outcomes for ESRD: A
meta-analysis. J Am Soc Nephrol. 26:2289–2302. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Purnell TS, Luo X, Cooper LA, Massie AB,
Kucirka LM, Henderson ML, Gordon EJ, Crews DC, Boulware LE and
Segev DL: Association of race and ethnicity with live donor kidney
transplantation in the United States from 1995 to 2014. JAMA.
319:49–61. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Teixeira JP, Combs SA and Teitelbaum I:
Peritoneal dialysis: Update on patient survival. Clin Nephrol.
83:1–10. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Weinhandl ED, Foley RN, Gilbertson DT,
Arneson TJ, Snyder JJ and Collins AJ: Propensity-matched mortality
comparison of incident hemodialysis and peritoneal dialysis
patients. J Am Soc Nephrol. 21:499–506. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Morinelli TA, Luttrell LM, Strungs EG and
Ullian ME: Angiotensin II receptors and peritoneal dialysis-induced
peritoneal fibrosis. Int J Biochem Cell Biol. 77:240–250. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fernández-Perpén A, Pérez-Lozano ML, Bajo
MA, Albar-Vizcaino P, Sandoval Correa P, del Peso G, Castro MJ,
Aguilera A, Ossorio M, Peter ME, et al: Influence of
bicarbonate/low-GDP peritoneal dialysis fluid (BicaVera) on in
vitro and ex vivo epithelial-to-mesenchymal transition of
mesothelial cells. Perit Dial Int. 32:292–304. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ueno T, Nakashima A, Doi S, Kawamoto T,
Honda K, Yokoyama Y, Doi T, Higashi Y, Yorioka N, Kato Y, et al:
Mesenchymal stem cells ameliorate experimental peritoneal fibrosis
by suppressing inflammation and inhibiting TGF-β1 signaling. Kidney
Int. 84:297–307. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Huh JY, Seo EY, Lee HB and Ha H:
Glucose-based peritoneal dialysis solution suppresses adiponectin
synthesis through oxidative stress in an experimental model of
peritoneal dialysis. Perit Dial Int. 32:20–28. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jasinge E, Kularatnam GAM, Dilanthi HW,
Vidanapathirana DM, Jayasena KLSPKM, Chandrasiri NDPD, Indika NLR,
Ratnayake PD, Gunasekara VN, Fairbanks LD and Stiburkova B: Uric
acid, an important screening tool to detect inborn errors of
metabolism: A case series. BMC Res Notes. 10:4542017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Anker SD, Doehner W, Rauchhaus M, Sharma
R, Francis D, Knosalla C, Davos CH, Cicoira M, Shamim W, Kemp M, et
al: Uric acid and survival in chronic heart failure: Validation and
application in metabolic, functional, and hemodynamic staging.
Circulation. 107:1991–1997. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Richette P and Bardin T: Gout. Lancet.
375:318–328. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wakabayashi K, Hamada C, Kanda R, Nakano
T, Io H, Horikoshi S and Tomino Y: Adipose-derived mesenchymal stem
cells transplantation facilitate experimental peritoneal fibrosis
repair by suppressing epithelial-mesenchymal transition. J Nephrol.
27:507–514. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Brodeur AC, Roberts-Pilgrim AM, Thompson
KL, Franklin CL and Phillips CL: Transforming growth
factor-β1/Smad3-independent epithelial-mesenchymal transition in
type I collagen glomerulopathy. Int J Nephrol Renovasc Dis.
10:251–259. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hipp S, Berg D, Ergin B, Schuster T,
Hapfelmeier A, Walch A, Avril S, Schmalfeldt B, Höfler H and Becker
KF: Interaction of Snail and p38 mitogen-activated protein kinase
results in shorter overall survival of ovarian cancer patients.
Virchows Arch. 457:705–713. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ouanouki A, Lamy S and Annabi B:
Periostin, a signal transduction intermediate in TGF-β-induced EMT
in U-87MG human glioblastoma cells, and its inhibition by
anthocyanidins. Oncotarget. 9:22023–22037. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tennakoon AH, Izawa T, Kuwamura M and
Yamate J: Pathogenesis of type 2 epithelial to mesenchymal
transition (EMT) in renal and hepatic fibrosis. J Clin Med.
5:E42015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu RM and Gaston Pravia KA: Oxidative
stress and glutathione in TGF-beta-mediated fibrogenesis. Free
Radic Biol Med. 48:1–15. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zharikov S, Krotova K, Hu H, Baylis C,
Johnson RJ, Block ER and Patel J: Uric acid decreases NO production
and increases arginase activity in cultured pulmonary artery
endothelial cells. Am J Physiol Cell Physiol. 295:C1183–C1190.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kang DH, Nakagawa T, Feng L, Watanabe S,
Han L, Mazzali M, Truong L, Harris R and Johnson RJ: A role for
uric acid in the progression of renal disease. J Am Soc Nephrol.
13:2888–2897. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Strippoli R, Moreno-Vicente R, Battistelli
C, Cicchini C, Noce V, Amicone L, Marchetti A, Del Pozo MA and
Tripodi M: Molecular mechanisms underlying peritoneal EMT and
fibrosis. Stem Cells Int. 2016:35436782016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li S, Lu J, Chen Y, Xiong N, Li L, Zhang
J, Yang H, Wu C, Zeng H and Liu Y: MCP-1-induced ERK/GSK-3β/Snail
signaling facilitates the epithelial-mesenchymal transition and
promotes the migration of MCF-7 human breast carcinoma cells. Cell
Mol Immunol. 14:621–630. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Margetts PJ, Bonniaud P, Liu L, Hoff CM,
Holmes CJ, West-Mays JA and Kelly MM: Transient overexpression of
TGF-{beta}1 induces epithelial mesenchymal transition in the rodent
peritoneum. J Am Soc Nephrol. 16:425–436. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Mulder KM: Role of Ras and Mapks in
TGFbeta signaling. Cytokine Growth Factor Rev. 11:23–35. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Witte D, Otterbein H, Förster M, Giehl K,
Zeiser R, Lehnert H and Ungefroren H: Negative regulation of
TGF-β1-induced MKK6-p38 and MEK-ERK signalling and
epithelial-mesenchymal transition by Rac1b. Sci Rep. 7:173132017.
View Article : Google Scholar : PubMed/NCBI
|