Introduction
Colorectal cancer (CRC) is the third-leading cause
of cancer mortality in most countries, and there were an estimated
134,490 new cases and 49,190 deaths in the United States in 2015
(1,2). Although the mortality rate from CRC
has declined by 3% over the past decade, 30–40% of CRC patients
still develop metastases and 50% die of CRC recurrence (3). CRC, which is associated with poor
prognosis, is usually diagnosed at a late stage (4–7).
Critical cancer hallmarks include the ability of tumor cells to
invade and migrate to distant tissues and to exhibit enhanced cell
proliferation (8); therefore,
elucidation of these cancer-promoting mechanisms may lead to drug
discoveries for novel therapies and subsequent improvement of
survival rates of CRC patients.
Filamins (FLNs), a family of actin-binding proteins,
comprise FLNa, FLNb and FLNc. FLNa, also known as filamin-A, was
identified in macrophages in 1975 and was originally characterized
as a protein that could cross-link actin filaments and form rigid
gels (9–11). Its contribution to the formation of
the actin cytoskeleton serves a vital role in responses to
extracellular stimulation. The function of FLNa has been revealed
in studies that have collectively identified over 90
filamin-binding proteins that are involved in cell migration, cell
adhesion, phosphorylation, cell signaling, receptor activation and
other important cellular functions (12).
The cross-linking structure of FLNa accompanies
extracellular signals to the cellular cytoskeleton. FLNa forms a
homodimer and interacts with cortical actin to form a dynamic
three-dimensional structure (13).
By interacting with transmembrane receptor complexes, adaptor
molecules and second messengers, FLNa regulates signaling events
involved in cell motility. The molecular function of FLNa in cell
chemotaxis remains debatable and seems to vary according to the
levels of FLNa and its interacting partners (14). Abnormal expression of FLNa is
associated with a wide spectrum of human disorders (15–17)
that are caused by abnormal interactions between FLNa and its
corresponding partners (18,19).
Zhu et al (20) considered
that FLNa regulated the activation of epidermal growth factor
receptor (EGFR) in human melanoma cells.
Epidermal growth factor receptor (EGFR) is a
tyrosine kinase receptor, also known as ErbB1. EGFR is the
prototype of the ErbB family and is expressed in nearly all
epithelial tissues (21,22). EGFR is a major regulator of cell
proliferation, metabolism, survival and motility (23); its inappropriate activation has an
important role in several types of cancer (24). Ligands that activate EGFR lead to
receptor tyrosine kinase autophosphorylation and activate several
important signaling molecules, such as those within
RAS/RAF/mitogen-activated protein kinase kinase (MEK)/ERK and
PI3K/Akt pathways (25–28); EGFR subsequently transmits signals
through these pathways (29).
The present study measured the proliferative and
migratory abilities of the SW480 cell line after experimentally
silencing FLNa expression. Then, the effects of FLNa silencing on
the activation of EGFR and related signaling molecules were
explored. Finally, the relationship between FLNa expression and the
clinical pathology of CRC patients was determined using
immunohistochemical analysis.
Materials and methods
Cell culture
The CRC cell line, SW480, was cultured in RPMI 1640
medium containing 10% fetal bovine serum (FBS) in a 5%
CO2 incubator at 37°C. SW480 cells were obtained from
Columbia University as a gift; RPMI 1640 medium was purchased from
Gibco (Thermo Fisher Scientific, Inc.); FBS was purchased from
Hyclone (GE Healthcare Life Sciences).
Transfection of short hairpin (sh)RNA
plasmids
CRC SW480 cells were seeded at a density of
4×105/ml, cultured for 24 h, transfected with pSIF1-FLNa
shRNA (SW480/KD group;
5′-GGGCTGACAACAGTGTGGTGCCTTCCTGTCAGAGCACCACACTGTTGTCAGCCCTTTTT-3′;
from our laboratory) or control shRNA (SW480/Ctrl group;
5′-TTGTCCGAACGTGCGAGGAGGCTTCCTGTCAGACCTCCTCGCACGTTCGGACAATTTTT-3;
from our laboratory) containing pSIF1 plasmids (2 µg; from our
laboratory) was used for transfection using
Lipofectamine® 2000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.), and selected with puromycin (2 µg/ml) for 4
weeks. Protein expression was examined by western blot analysis as
detailed below.
MTS assay
SW480/KD and SW480/Ctrl cells were placed in
separate 96-well plates (5×103/well). After starvation
with serum-free 1640 medium (SFM) for 2 h, the cells were treated
with various concentrations (0, 4, 20 and 100 nM) of EGF (Miltenyi
Biotec GmbH); after 48 h incubation, cell viability was determined
using MTS tetrazolium substrate (Cell Titer 96 Aqueous One Solution
Cell Proliferation Assay; Promega Corporation). Subsequently, a
spectrophotometer (Thermo Fisher Scientific, Inc.) was used to
determine the optical density at 490 nm as previously described
(30).
Wound healing assay
SW480/KD and SW480/Ctrl cells were starved for 4 h
with SFM and were then scratched with 10 µl pipette tips and washed
with PBS to remove the floating cells. Subsequently, the two groups
were untreated or treated with 20 nM EGF for 24 h. An inverted
microscope (magnification ×10) was used to assess the width of the
scratch; images of the same field were captured every 8 h (31).
Transwell assay
After 4 h of starvation with SFM, SW480/KD and
SW480/Ctrl cells were seeded in the top chamber (0.4 µm
polycarbonate-membrane inserts) with a concentration of
106 cells per 200 µl in SFM. The bottom chamber was
filled with 600 µl of RPMI 1640/10% FBS medium. Cells were allowed
to migrate for 24 h (untreated or treated with 20 nM EGF).
Non-migratory cells on the upper surface were then removed using a
wet cotton swab. The migrated cells on the surface of the lower
membrane were stained by H&E and counted under a light
microscope (magnification ×200; Olympus Corporation) in 5 random
fields to obtain the average number (32).
Western blotting
SW480 cells were treated with or without EGF (20 nM)
for 5, 10 and 30 min and then lysed with cold lysis buffer (Amyjet
Scientific, Inc,). The protein concentration was quantified using a
bicinchoninic acid assay kit. Proteins (20 µg/lane) were separated
via 10% SDS-PAGE and were transferred onto polyvinylidene
difluoride membranes. Subsequently, PVDF membranes were blocked
with 3% bovine serum albumin (Thermo Fisher Scientific, Inc.) at
room temperature for 2 h and then were incubated with primary
antibodies in 4°C for 12 h, followed by incubation with secondary
antibodies at 25°C for 1.5 h and visualization with enhanced
chemiluminescence reagent (Beyotime Institute of Biotechnology).
ImageJ software (v1.52; National Institutes of Health) was used to
quantity proteins. Primary antibodies included the following:
Rabbit anti-FLNa antibody (1:1,000; cat. no. MAB1680; EMD
Millipore), rabbit anti-EGFR antibody (1:1,000; cat. no.
18986-1-AP; ProteinTech Group, Inc.), rabbit anti-phosphorylated
(p-)EGFR antibody (Tyr1068; 1:2,000; cat. no. 3777; Cell Signaling
Technology, Inc.), rabbit anti-Akt antibody (1:1,000; cat. no.
10176-2-AP; ProteinTech Group, Inc.), rabbit anti-p-Akt antibody
(Ser473; 1:1,000; cat. no. 66444-1-Ig; ProteinTech Group, Inc.),
rabbit anti-ERK1/2 antibody (1:4,000; cat. no. 16443-1-AP;
ProteinTech Group, Inc.), rabbit anti-p-ERK1/2 antibody
(Thr202/Tyr204; 1:3,000; cat. no. 4370, Cell Signaling Technology.)
and mouse anti-β-actin antibody (1:5,000; cat. no. 60008-1-Ig;
ProteinTech Group, Inc.). Horseradish peroxidase (HRP)-conjugated
anti-mouse IgG (1:5,000; cat. no. 7076) and anti-rabbit IgG
(1:5,000; cat. no. 7074; both Cell Signaling Technology) secondary
antibodies were used.
Xenograft experiments in vivo
A total of 20 BALB/c male nude mice (age, 6–8 weeks;
weight, 15.3–18.2 g) were purchased from the Experimental Animal
Center of Hebei Medical University. Mice were housed at 20–26°C and
40–60% humidity under a 12:12 h light/dark cycle, with access to
food and water ad libitum. The present study was approved by
the Laboratory Animal Ethical Committee of the Fourth Hospital of
Hebei Medical University. Xenograft experiments were performed as
previously described (33). Cells
(2×106) suspended in Hank's balanced salt solution
(HyClone; GE Healthcare Life Sciences) were injected in the right
thigh. After 2 weeks, this experiment was ended. The isolated solid
tumors were measured for size, weight and volume.
Patients and specimens
A total of 82 patients with CRC (51.1±15.9 years
old; range, 42–74 years), who were admitted to the general surgery
department at the 980th Hospital of the PLA Joint Logistics Support
Force between November 2015 and May 2016, were included in the
present study. All subjects provided written informed consent.
Colorectal cancer tissues were obtained from the 980th Hospital of
the PLA Joint Logistics Support Force (Bethune International Peace
Hospital) with the approval of the Hospital Ethics Committee and
were used according to ethical procedures.
Immunohistochemical (IHC)
staining
Immunohistochemical staining was conducted to
determine the protein expression of FLNa and Ki-67 by using of a
diaminobenzidine kit (OriGene Technologies, Inc.) according to the
manufacturer's instructions. Tissues were fixed with 4%
paraformaldehyde at room temperature for 48 h, embedded in paraffin
and sectioned (5 µm). Briefly, after microwave-antigen retrieval,
endogenous peroxidase activity was inhibited by incubation with 3%
H2O2. Sections were blocked with 5% goat
serum (Absin Bioscience, Inc.) at room temperature for 40 min, then
incubated overnight at 4°C with a polyclonal rabbit anti-Ki-67
antibody (1:100; cat. no. 27309-1-AP; ProteinTech Group, Inc.) or
anti-FLNa antibody (1:100; cat. no. 67133-1-Ig; ProteinTech Group,
Inc.). After washing with PBS, the sections were incubated with a
secondary antibody at 37°C for 30 min [HRP-conjugated goat
anti-mouse IgG (1:1,000; cat. no. SA00001-1) and goat anti-rabbit
IgG (1:1,000; cat. no. SA00001-2), both ProteinTech Group, Inc.].
Sections were counterstained with hematoxylin and eosin at room
temperature for 4 min to visualize nuclei. Five different fields
were randomly selected per sample. Images were acquired using a
Leica microscope (magnification ×100; Leica DM6000B; Leica
Microsystems GmbH) and were digitized with LAS version 4.4 (Leica
Microsystems GmbH). The thresholds for dichotomizing FLNa IHC were
set to the medians for each measurement.
Statistical analysis
All data are reported as mean ± standard deviation.
All statistics and graphs were obtained using GraphPad Prism 5
(GraphPad Software, Inc.); SPSS 19.0 software (IBM Corp.) was used
for Kaplan-Meier plots and ANOVA. Cell viability, cell number, band
density and gene expression were analyzed by ANOVA followed by
Bonferroni post hoc test when making comparisons in datasets
containing multiple groups. Spearman's test was used for the
correlation analysis of FLNa and Ki67. The maximal log-rank method
was used for optimal cut-point determination and adjusted P-values.
Cut-points within the central 80% of ordered FLNa protein
expression were considered. P<0.05 was considered to indicate a
statistically significant difference.
Results
FLNa knockdown accelerates EGF-induced
cell proliferation and migration in SW480 cells
To elucidate the role of FLNa in colon carcinoma
cells, EGF-treated SW480 cells were transfected with shFLNa to
knockdown FLNa expression. The efficacy of the knockdown was
confirmed via western blotting. FLNa expression was significantly
downregulated in the SW480/KD group (0.48±0.01) compared with that
of the SW480/Ctrl group (1.13±0.03; P<0.01; Fig. 1A). A cell viability assay
demonstrated that FLNa knockdown promoted EGF-induced cell
proliferation in a dose-dependent manner in the SW480/KD group
(P<0.01; Fig. 1B). Cell scratch
migration assays demonstrated that FLNa knockdown significantly
accelerated cell migration rates, and this effect was independent
of EGF treatment (Fig. 1C and D).
A Transwell assay also indicated that the number of migrated cells
in the SW480/KD group was greater compared with the SW480/Ctrl
group (76±3 vs. 21±5; P<0.05) and the migration ability was even
further increased in the SW480/KD group compared with those in the
EGF-treatment group (Fig. 1E and
F).
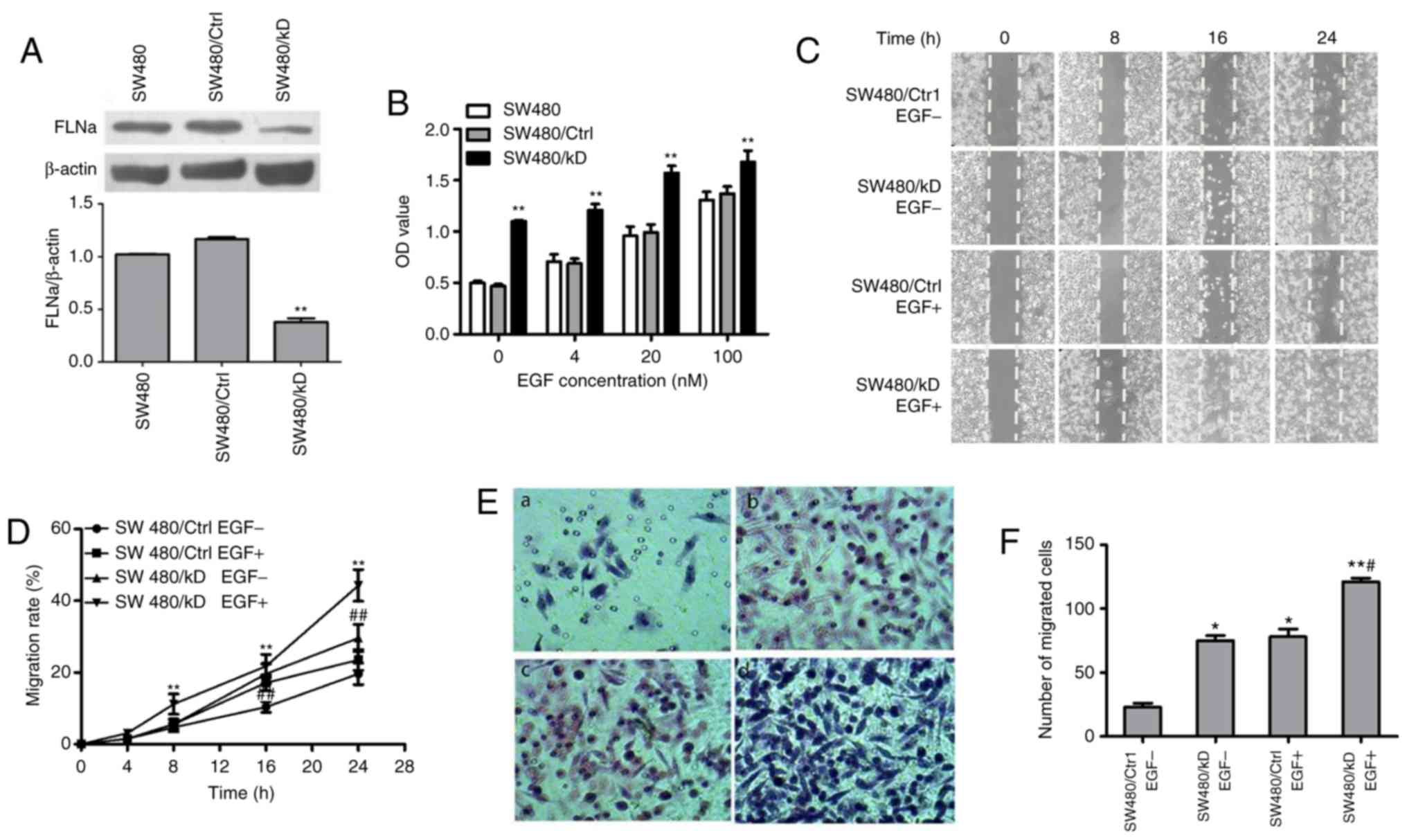 | Figure 1.FLNa knockdown promotes proliferation
and migration of SW480 cells. (A) Western blot analysis confirmed
the shRNA-mediated silencing of FLNa expression in SW480 cells
transfected with shRNA FLNa for 4 weeks compared with cells
transfected with control. **P<0.01 vs. SW480/Ctrl, n=3. (B)
SW480 cells were transfected with shRNA FLNa or control shRNA, then
treated with EGF (0, 4, 20 and 100 nM) for 24 h. Cell viability was
determined using the MTS assay at 0, 5, 10 and 30 min. Data are
presented as the mean ± standard deviation. **P<0.01 and vs.
control, n=3. (C) Representative photomicrographs of a scratch
assay performed with EGF-treated SW480 cells transfected with shRNA
FLNa or control after scratching at 0, 8, 16 and 24 h.
Magnification ×10. (D) Quantification of SW480 cells migrating into
the scratch gap. The data are illustrated as the mean ± standard
error of mean from three independent experiments. **P<0.01 vs.
SW480/Ctrl EGF-treatment or ##P<0.01 vs. SW480/KD
EGF-treatment. (E) Boyden-chamber assay demonstrating that
EGF-treated SW480 cells transfected with shRNA FLNa or control for
24 h traversed the filter to the other side and were stained by
hematoxylin and eosin. Magnification ×200. (F) Quantification of
cells migrated to the lower side of the membrane. *P<0.05 or
**P<0.01 vs. SW480/Ctrl EGF-; #P<0.05 vs. SW480/KD
EGF-. FLNa, Filamin A; sh, short hairpin; EGF, epidermal growth
factor; KD, knockdown; Ctrl, control. |
Effects of FLNa on EGFR
phosphorylation in CRC cells
Because the activation of EGFR, Akt and ERK
signaling is required for cell proliferation and migration
(34), the effect of
shRNA-mediated FLNa knockdown on EGF-stimulated SW480 cells was
evaluated via western blotting. The protein expression levels of
EGFR, Akt and ERK, and their activated phosphorylated forms, were
detected in SW480/KD and SW480/Ctrl cells treated with EGF for 0,
5, 10 and 30 min. FLNa expression in the SW480/KD group was
significantly decreased at every tested time point compared with
the SW480/Ctrl group (Fig. 2A and
B). Furthermore, FLNa knockdown significantly increased the
p-EGFR/EGFR (Fig. 2C), p-Akt/Akt
(Fig. 2D) and p-ERK/ERK (Fig. 2E) protein ratios compared with the
control (P<0.01), indicating that FLNa knockdown activated the
EGFR, Akt and ERK signaling pathways. These data suggested that
FLNa regulated cell proliferation and migration via EGFR, Akt and
ERK signaling in colon carcinoma cells.
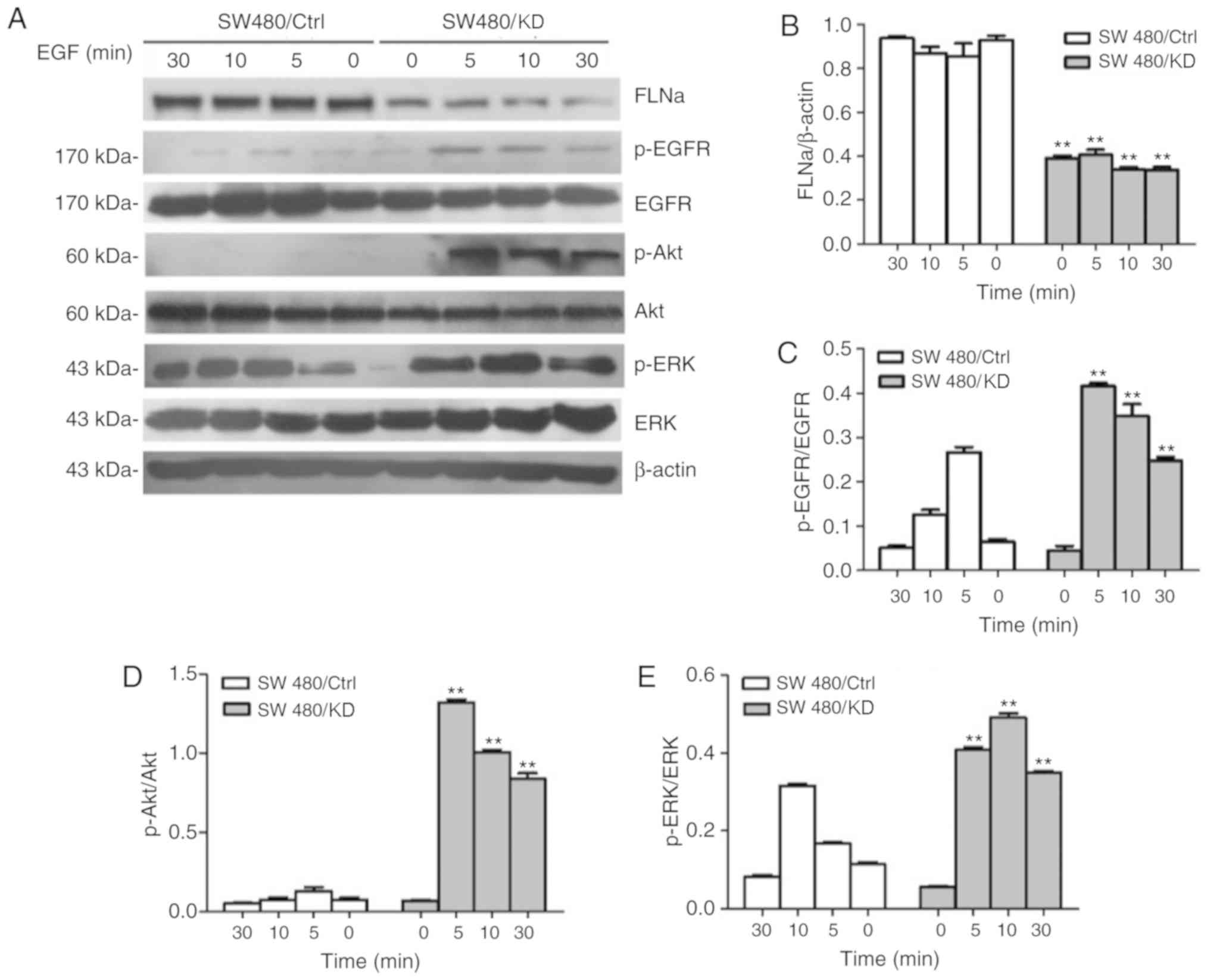 | Figure 2.FLNa knockdown promotes EGF-induced
EGFR/Akt/ERK phosphorylation in SW480 cells. SW480 cells were
transfected with shRNA FLNa or control shRNA for 24 h. Then cells
were treated with or without EGF (20 nM) for 0, 5, 10 and 30 min.
(A) Total proteins were extracted and subjected to western blotting
with antibodies against FLNa, EGFR, p-EGFR, p-Akt, Akt, p-ERK and
ERK; β-actin is shown here as a loading control. (B) Band
intensities for the ratio of FLN/β-actin were measured. (C) Band
intensities for the ratio of p-EGFR/EGFR, (D) p-Akt/Akt and (E)
p-ERK/ERK were measured and normalized to β-actin. **P<0.01 vs.
SW480/Ctrl. FLNa, Filamin A; EGF, epidermal growth factor; sh,
short hairpin; EGFR, epidermal growth factor receptor; p-,
phosphorylated. |
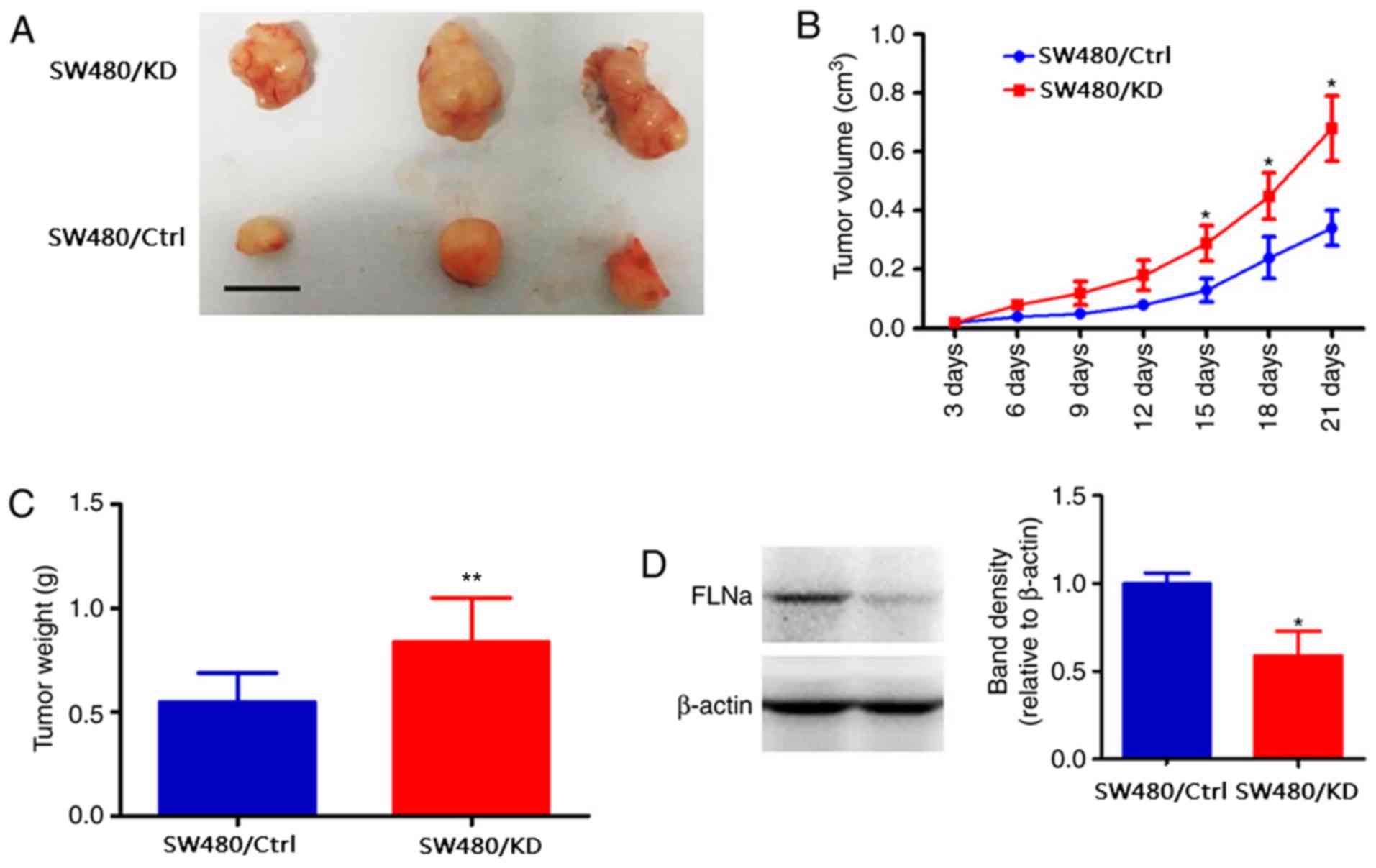
FLNa silencing promotes SW480 cell
growth in vivo
To further confirm the oncogenic efficiency of FLNa
knockdown, SW480/KD or SW480/Ctrl were subcutaneously inoculated
into the left flank (SW480/KD) and right flank (SW480/Ctrl in nude
mice, respectively; n=8). After 14 days, the tumors were larger in
the SW480/KD group compared with the SW480/Ctrl group (0.613±0.114
vs. 0.378±0.068 cm3, P<0.05; Fig. 3A and B). In addition, the tumor
weight at the end of the experiment was increased in the SW480/KD
group (0.512±0.031 g) compared with the SW480/Ctrl group
(0.371±0.044 g; P<0.05; Fig.
3C). Western blotting confirmed that the FLNa levels were
significantly decreased in the SW480/KD tumor tissues compared with
those in the control (Fig. 3D).
These results further indicated that FLNa knockdown accelerated CRC
tumor growth.
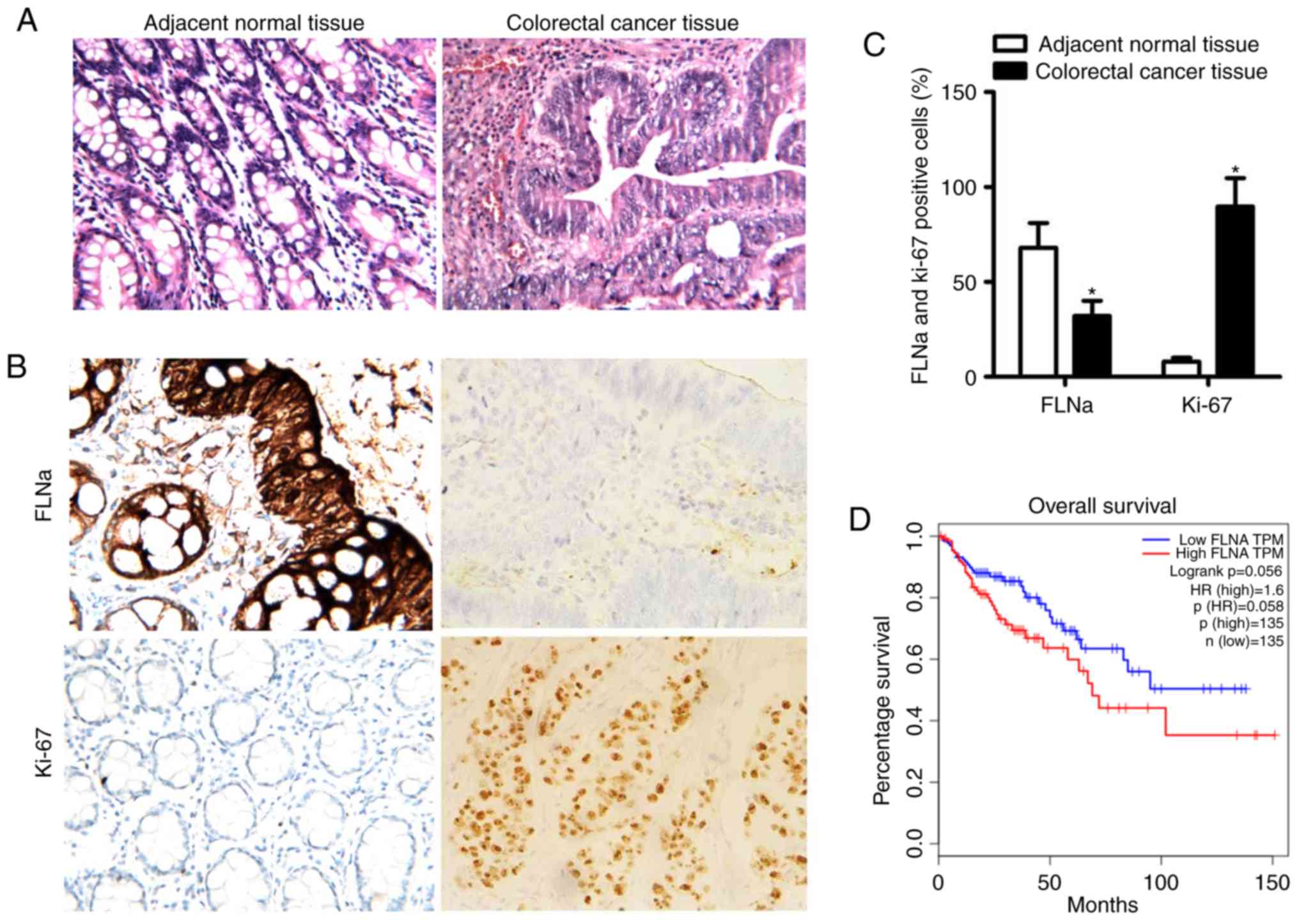
Expression of FLNa in CRC tissues from
patients and correlation with Ki-67 expression
The characteristics of the 82 CRC patients (median,
51.1±15.7 years; range, 42–74 years old) enrolled in the present
study are listed in Table I. The
expression levels of FLNa exhibited a significant association with
TNM stage and lymph node metastasis in CRC, but not with gender,
age, diameter of tumor, or degree of differentiation (Table I). Representative photographs of
hematoxylin and eosin and immunohistochemical staining for FLNa and
Ki-67 in adjacent normal tissues and CRC tissues (Fig. 4A and B) revealed significantly
higher expression of Ki-67 and lower expression of FLNa in CRC
tissues compared with adjacent normal tissues (P<0.05; Fig. 4C). FLNa expression had a
significant negative correlation with Ki-67 expression (Table II), which may explain how FLNa
knockdown promoted tumor growth. By contrast, low expression of
FLNa did not influence the overall survival time for the patients
with CRC, as evidenced by Kaplan-Meier survival analysis (Fig. 4D).
 | Table I.Relationship between FLNa and
clinical-pathological features. |
Table I.
Relationship between FLNa and
clinical-pathological features.
|
|
| FLNa expression
(%) |
|
|---|
|
|
|
|
|
|---|
| Characteristic | N (%) | Low (n=40) | High (n=42) | P-value |
|---|
| Age |
|
|
| 0.3928 |
|
≤60 | 38 (46.3) | 20 (50.0) | 18 (42.8) |
|
|
>60 | 44 (53.7) | 20 (50.0) | 24 (57.1) |
|
| Sex |
|
|
| 0.9913 |
|
Female | 43 (52.4) | 21 (52.5) | 22 (52.4) |
|
|
Male | 39 (47.6) | 19 (47.5) | 20 (47.6) |
|
| Diameter of tumor,
cm |
|
|
| 0.6414 |
| ≤4 | 42 (51.2) | 20 (50.0) | 22 (52.4) |
|
|
>4 | 40 (48.8) | 20 (50.0) | 20 (47.6) |
|
| Degree of
differentiation |
|
|
| 0.9827 |
|
Low | 37 (45.1) | 18 (45.0) | 19 (45.2) |
|
|
Moderate or high | 45 (54.9) | 22 (55.0) | 23 (54.8) |
|
| TNM stage |
|
|
| 0.0008 |
|
I–II | 36 (43.9) | 10 (25.0) | 26 (61.9) |
|
|
III–IV | 46 (56.1) | 30 (75.0) | 16 (38.1) |
|
| Lymph node
metastasis |
|
|
| 0.0007 |
|
Yes | 48 (58.5) | 31 (77.5) | 17 (40.5) |
|
| No | 34 (41.5) | 9 (22.5) | 25 (59.5) |
|
| Table II.Relationship between the expression
of FLNa and Ki-67. |
Table II.
Relationship between the expression
of FLNa and Ki-67.
|
| Ki-67 expression
(%) |
|
|
|---|
|
|
|
|
|
|---|
| FLNa
expression | Negative
(n=38) | Positive
(n=44) | rs | P-value |
|---|
| Negative
(n=40) | 12 (31.6) | 28 (63.6) | −0.32 | 0.004 |
| Positive
(n=42) | 26 (68.4) | 16 (36.4) |
|
|
Discussion
As a large cytoplasmic protein, FLNa mainly
crosslinks actin filaments, membrane receptors and signaling
intermediates (35). The specific
role of FLNa in cancer metastasis has remained elusive. Through
MTS, wound healing and Transwell assays in the present study, it
was identified that FLNa knockdown increased the proliferative and
migratory abilities of CRC cells in vitro. In addition, FLNa
knockdown was demonstrated to promote tumor growth in vivo.
These findings are in accord with an earlier report on breast
cancer cells that overexpressed ErbB2, in which knockdown of FLNa
promoted division and metastasis of tumor cells (36).
CRC cells invade and metastasize to distant sites
and these phenomena are accompanied by aberrant activation of cell
signaling (37). A number of
studies have reported filamin structures and filamin-binding
proteins that are involved in cell signaling and other important
cellular functions. For example, FLNa is involved in the
organization of the actin network (10,38),
acts as a ‘molecular switch’ to convert mechanical stimuli to
chemical signals (39). These
multiple interactions suggest that FLNa is a key component of a
variable signaling-scaffold complex (10). In addition, EGFR is an
actin-binding protein and has become an indispensable molecular
target for cancer therapy. EGFR is activated by certain ligands
(such as EGF) and provokes various cellular signaling events
resulting in increased cell proliferation, invasion and metastasis
(29). EGFR carries out these
functions through activating multiple signaling cascades, including
RAS/RAF/MEK/ERK and PI3K/Akt pathways (40). The present study demonstrated that
the phosphorylation levels of EGFR, ERK and Akt were significantly
increased in the FLNa-knockdown group compared with the control,
whereas no difference was observed in the total EGFR, ERK and Akt
expression levels between these two groups. The present results
indicated that lower expression of FLNa promoted the proliferative
and migratory ability of CRC cells by activating EGFR and its
downstream signaling proteins. Under certain circumstances, this
aberrantly activated pathway may lead to abnormal cell growth and
invasion, which could subsequently result in drug resistance or
metastasis to distant sites (41).
An important limitation of the present study was that only one cell
line was used; therefore, future studies are required to confirm
the findings of the present study by using additional cell
lines.
Accumulating evidence suggests that FLNa
participates in the development of multiple tumors. Xu et al
(42) demonstrated that FLNa
regulates focal-adhesion disassembly and inhibits breast cancer
cell migration and invasion. A previous study showed that there is
a positive relationship between FLNa and VEGF in patients with lung
cancer (43). However, a separate
study emphasized that FLNa interacting with other proteins inhibits
CRC progression (44), indicating
that FLNa may represent a novel cancer-suppressor gene for treating
colorectal adenocarcinoma. Thus, the current literature suggests a
dual role for FLNa and complex underlying mechanisms in various
types of cancer. The present study found that FLNa knockdown
correlated with TNM stage and lymph node metastasis in patients
with CRC, which is in accordance with a previous study on gastric
cancer (45). A possible mechanism
for the downregulation of the FLNa gene in CRC cells may be
mutagenesis. Notably, FLNa expression was demonstrated to be
negatively correlated with the expression of Ki-67, which is a
reliable index of cancer progression. The present data suggested
that FLNa may have protective roles as a negative regulator in CRC
SW480 cells by promoting proliferation via the activation of many
signaling pathways. Hence, FLNa may represent a novel prognostic
marker and therapeutic target for treating CRC.
Acknowledgements
Not applicable.
Funding
This work was supported by the National Natural
Science Foundation of China (grant no. 81071846 to TNZ), the
Natural Science Foundation of Hebei Province of China (grant no.
H2013505059 to TNZ), the Department of Science and Technology of
Hebei Province of China (grant no. 12396107D to RJZ, and grant nos.
14397707D, 09966114D and 092461102D to TNZ) and the Wu Jieping
Medical Foundation (grant nos. 320.6750.12604, 320.6750.14063 and
320.6799.15005 to TNZ).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
KW performed the experiments and data analysis. KW
and TZ participated in the design and coordination of experimental
work. KW and RZ were involved in data acquisition. KW and TZ
drafted the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Laboratory
Animal Ethical Committee of the Fourth Hospital of Hebei Medical
University (Shijiazhuang, China). Experimental procedures were
implemented in accordance with the guidelines and regulations of
the Hospital Ethics Committee and were performed according to
ethical procedures of the 980th Hospital of the PLA Joint Logistics
Support Force (Bethune International Peace Hospital).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ferlay J, Steliarova-Foucher E, Lortet
Tieulent J, Rosso S, Coebergh JW, Comber H, Forman D and Bray F:
Cancer incidence and mortality patterns in Europe: Estimates for 40
countries in 2012. Eur J Cancer. 49:1374–1403. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Naishadham D and Jemal A:
Cancer statistics, 2016. CA Cancer J Clin. 66:7–30. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sinclair P, Singh A, Riaz AA and Amin A:
An unsolved conundrum: The ideal follow up strategy after curative
surgery for colorectal cancer. Gastrointest Endosc. 75:1072–1079.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Vogelstein B, Fearon ER, Hamilton SR, Kern
SE, Preisinger AC, Leppert M, Nakamura Y, White R, Smits AM and Bos
JL: Genetic alterations during colorectal-tumor development. N Engl
J Med. 319:525–532. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fan N, Kang R, Ge X, Li M, Liu Y, Chen HM
and Gao CF: Identification of alpha-2-HS-glycoprotein precursor and
tubulin beta chain as serology diagnosis biomarker of colorectal
cancer. Diagn Pathol. 12:532014. View Article : Google Scholar
|
|
6
|
Thomson DM, Krupey J, Freedman SO and Gold
P: The radioimmunoassay of circulating carcinoma bryonic antigen of
the human digestive system. Proc Natl Acad USA. 64:161–167. 1969.
View Article : Google Scholar
|
|
7
|
Duffy MJ: Role of tumor markers in
patients with solid cancers: A critical review. Eur J Intern Med.
18:175–184. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hartwig JH and Stossel TP: Isolation and
properties of actin, myosin and a new actin-binding protein in
rabbit alveolar macrophages. J Biol Chem. 250:5696–5705.
1975.PubMed/NCBI
|
|
10
|
Stossel TP, Condeelis J, Cooley L, Hartwig
JH, Noegel A, Schleicher M and Shapiro SS: Filamins as integrators
of cell mechanics and signalling. Nat Rev Mol Cell Biol. 2:138–145.
2001. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gomer RH and Lazarides E: Switching of
filamin polypeptides during myogenesis in vitro. J Cell Biol.
96:321–329. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Savoy RM and Ghosh PM: The dual role of
filamin a in cancer: Can't live with (too much of) it, can't live
without it. Endocr Relat Cancer. 20:R341–R356. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Djinovic-Carugo K and Carugo O: Structural
portrait of filamin interaction mechanisms. Cur Protein Pept Sci.
11:639–650. 2010. View Article : Google Scholar
|
|
14
|
Xu YJ, Bismar TA, Su J, Xu B, Kristiansen
G, Varga Z, Teng L, Ingber DE, Mammoto A, Kumar R and Alaoui-Jamali
MA: Filamin a regulates focal adhesion disassembly and suppresses
breast cancer cell migration and invasion. J Exp Med.
207:2421–2437. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Feng Y, Chen MH, Moskowitz IP, Mendonza
AM, Vidali L, Nakamura F, Kwiatkowski DJ and Walsh CA: Filamin a
(FLNA) is required for cell-cell contact in vascular development
and cardiac morphogenesis. Proc Natl Acad Sci USA. 103:19836–19841.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang Y, Kreisberg JI, Bedolla RG,
Mikhailova M, deVere White RW and Ghosh PM: A 90 kDa fragment of
filamin A promotes casodex-induced growth inhibition in
Casodex-resistant and rogen receptor positive C4-2 prostate cancer
cells. Oncogene. 26:6061–6070. 2006. View Article : Google Scholar
|
|
17
|
Kyndt F, Gueffet JP, Probst V, Jaafar P,
Legendre A, Le Bouffant F, Toquet C, Roy E, McGregor L, Lynch SA,
et al: Mutations in the gene encoding filamin a as a cause for
familial cardiac valvular dystrophy. Circulation. 115:40–49. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nakamura F, Heikkinen O, Pentikainen OT,
Osborn TM, Kasza KE, Weitz DA, Kupiainen O, Permi P, Kilpeläinen I,
Ylänne J, et al: Molecular basis of filamin A-FilGAP interaction
and its impairment in congenital disorders associated with filamin
a mutations. PLoS One. 4:e49282009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhou AX, Hartwig JH and Akyurek LM:
Filamins in cell signaling transcription and organ development.
Trends Cell Biol. 20:113–123. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhu TN, He HJ, Kole S, D'Souza T, Agarwal
R, Morin PJ and Bernier M: Filamin A-mediated downregulation of the
exchange factor Ras-GRF1 correlates with decreased matrix
metalloproteinase-9 expression in human melanoma cells. J Biol
Chem. 282:14816–14826. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yarden Y and Sliwkowski MX: Untangling the
ErbB signalling network. Nat Rev Mol Cell Biology. 2:127–137. 2001.
View Article : Google Scholar
|
|
22
|
Roskoski R: The ErbB/HER family of
protein-tyrosine kinases and cancer. Pharmacol Res. 79:34–74. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yewale C, Baradia D, Vhora I, Patil S and
Misra A: Epidermal growth factor receptor targeting in cancer: A
review of trends and strategies. Biomaterials. 34:8690–8707. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
David II, Gallo RM and Settleman J:
Mutational activation of ErbB family receptor tyrosine kinases:
Insights into mechanisms of signal transduction and tumorigenesis.
Bioessays. 29:558–565. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lo HW: EGFR targeted therapy in malignant
Glioma: Novel aspects and mechanisms of drug resistance. Curr Mol
Pharmacol. 3:37–52. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Navolanic PM, Steelman LS and McCubrey JA:
EGFR family signaling and its association with breast cancer
development and resistance to chemotherapy. Int J Oncol.
22:237–252. 2003.PubMed/NCBI
|
|
27
|
Park OK, Schaefer TS and Nathans D: In
vitro activation of Stat3 by epidermal growth factor receptor
kinase. Proc Natl Acad Sci USA. 93:13704–13708. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bowman T, Garcia R, Turkson J and Jove R:
STATs in oncogenesis. Oncogene. 19:2474–2488. 2013. View Article : Google Scholar
|
|
29
|
Mclendon RE, Turner K, Perkinson K and
Rich J: Second messenger systems in human gliomas. Arch Pathol Lab
Med. 131:1585–1590. 2007.PubMed/NCBI
|
|
30
|
Wang CZ, Xie JT, Zhang B, Ni M, Fishbein
A, Aung HH, Mehendale SR, Du W, He TC and Yuan CS: Chemopreventive
effects of Panax notoginseng and its major constituents on SW480
human colorectal cancer cells. Int J Oncol. 31:1149–1156.
2007.PubMed/NCBI
|
|
31
|
Kaleağasıoğlu F and Berger MR:
Differential effects of erufosine on proliferation, wound healing
and apoptosis in colorectal cancer cell lines. Oncol Rep.
31:1407–1416. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
LI K, Zhou ZY, JI PP and Luo HS: Knockdown
of β-catenin by siRNA influences proliferation, apoptosis and
invasion of the colon cancer cell line SW480. Oncol Lett.
11:3896–3900. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Nakano H, Miyazawa T, Kinoshita K, Yamada
Y and Yoshida T: Functional screening identifies a microRNA,
miR-491 that induces apoptosis by targeting Bcl-X(L) in colorectal
cancer cells. Int J Cancer. 127:1072–1080. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cardoso AP, Pinto ML, Pinto AT, Oliveira
MI, Pinto MT, Gonçalves R, Relvas JB, Figueiredo C, Seruca R,
Mantovani A, et al: Macrophages stimulate gastric and colorectal
cancer invasion through EGFR Y(1086), c-Src, Erk1/2 and akt
phosphorylation and smallGTPase activity. Oncogene. 33:2123–2133.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Begonja AJ, Hoffmeister KM, Hartwig JH and
Falet H: FlnA-null megakaryocytes prematurely release large and
fragile platelets that circulate poorly. Blood. 118:2285–2295.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Popowicz GM, Schleicher M, Noegel AA and
Holak TA: Filamins: Promiscuous organizers of the cytoskeleton.
Trends Biochem Sci. 31:411–419. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Martin GS: Cell signaling and cancer.
Cancer Cell. 4:167–174. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang K, Ash JF and Singer SJ: Filamin, a
new high-molecular- weight protein found in smooth muscle and
nonmuscle cells. Proc Natl Acad Sci USA. 72:4483–4486. 1977.
View Article : Google Scholar
|
|
39
|
Ehrlicher AJ, Nakamura F, Hartwig JH,
Weitz DA and Stossel TP: Mechanical strain in actin networks
regulates FilGAP and integrin binding to filamin a. Nature.
478:260–263. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yi WY, Hong W, Kang HJ, Kim HJ, Zhao W,
Wang A, Seong YS and Bae I: Inhibition of the PI3K/Akt pathway
potentiates cytotoxicity of EGFR kinase inhibitors in
triple-negative breast cancer cells. J Cell Mol Med. 17:648–656.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Abrams SL, Steelman LS, Shelton JG, Wong
EW, Chappell WH, Bäsecke J, Stivala F, Donia M, Nicoletti F, Libra
M, et al: The Raf/MEK/ERK pathway can govern drug resistance,
apoptosis and sensitivity to targeted therapy. Cell Cycle.
9:1781–1791. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Xu Y, Bismar TA, Su J, Xu B, Kristiansen
G, Varga Z, Teng L, Ingber DE, Mammoto A, Kumar R and Alaoui-Jamali
MA: Filamin a regulates focal adhesion disassembly and suppresses
breast cancer cell migration and invasion. J Exp Med.
207:2421–2437. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Uramoto H, Akyurek LM and Hanagirl T: A
positive relationship between Filamin and VEGF in patiens with lung
cancer. Anticancer Res. 30:3939–3944. 2010.PubMed/NCBI
|
|
44
|
Park YL, Park SY, Lee SH, Kim RB, Kim JK,
Rew SY, Myung DS, Cho SB, Lee WS, Kim HS and Joo YE: Filamin a
interacting protein 1-like expression inhibits progression in
colorectal cancer. Oncotarget. 7:72229–72241. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Sun GG, Sheng SH, Jing SW and Hu WN: An
antiproliferative gene FLNA regulates migration and invasion of
gastric carcinoma cell in vitro and its clinical significance.
Tumor Biol. 3:2641–2648. 2014. View Article : Google Scholar
|