Introduction
Bronchial asthma is a chronic airway inflammatory
disease. As of 2008, ~300 million people worldwide have been
diagnosed with asthma, with China accounting for ~10% (~30 million)
of all cases (1). As a potential
life-threating condition, asthma has become a serious health
problem worldwide. Recently established treatment protocols have
significantly improved asthma control rates; however, symptomatic
treatment represents the primary available therapeutic option, and
inhaled corticosteroids and bronchodilators are the most common
medications used to treat asthma symptoms (2). Currently, the absence of effective
etiological treatments represents a limitation in improving the
asthma control rate.
Allergic asthma is a common type of asthma, and
children are twice as likely to develop this disease compared with
adults (3). Immune-mediated
inflammation is the principal factor involved in allergic asthma
pathogenesis, and a variety of inflammatory mediators and cytokines
are involved in this process, including interferon-γ (IFN-γ),
interleukin-4 (IL-4) and immunoglobulin E (IgE) (4). Dendritic cells (DCs) are involved in
the initiation and maintenance of the inflammatory chain reaction
underlying allergic asthma (5,6).
Previous studies have shown that DC surface molecules, including
major histocompatibility complex class II (MHC-II) molecules and
costimulatory proteins (7,8), and DC-secreted cytokines (9) can regulate the differentiation of
naïve T cells into type 1 or type 2 T-helper (Th2) cells, or
regulatory T cells (Tregs). DCs are associated with airway
epithelial cells, mast cells and eosinophils, three types of cells
that are involved in asthmatic pathology (10–12).
Asthma is caused by the activation and recruitment of inflammatory
cells and secretion of pro-inflammatory factors following exposure
to an antigen (13), suggesting
that asthma could be treated by suppressing this process.
CC chemokine receptor 7 (CCR7) is primarily
expressed on the surface of DCs, T-lymphocytes and B-lymphocytes
(14) and it has been shown to
promote the internalization of antigens by DCs, and to regulate
cell survival, migration, and to induce DC maturation (15,16).
Immune tolerance is the state of unresponsiveness of the immune
system to a particular antigen (17). A previous study has shown that the
CCR7-dependent migration of DCs from the lungs to draining lymph
nodes is involved in the transport of inhaled silver particles, and
this process is essential to induce peripheral tolerance of T cells
(18). However, this previous
study was primarily focused on the process of immune tolerance
associated with antigen presentation. The mechanism underlying the
role of CCR7-expressing DCs in the regulation of immune tolerance
in the airways during allergic asthma remains unclear and requires
further investigation. Therefore, the present study investigated
the effects of CCR7 knockdown and overexpression on DC-mediated
immune tolerance in the lungs of rats with allergic asthma.
Materials and methods
Culture of bone marrow-derived
immature DCs (imDCs)
The present study was approved by The Ethics
Committees of The People's Hospital of Shanxi Province. A total of
33 specific pathogen-free (SPF) male Wistar rats (age, 6–8 weeks;
weight, 180–200 g) were obtained from The Laboratory Animal Center
of Hangzhou Hibio Technology Co., Ltd. The animals were
acclimatized to laboratory conditions (23°C, 12-h light/dark cycle,
50% humidity and ad libitum access to food and water) for 2
weeks prior to experimentation. Animals were sacrificed by
intravenous injection of pentobarbital sodium (150 mg/kg) and
animal death was confirmed by lack of reflexes, heartbeat and
breathing. Specific tissues were collected for experimentation.
Bone marrow was collected from the femurs and tibiae of Wistar rats
as previously described (19). The
intact femurs and tibiae were kept in 70% ethanol for 30 min, and
then rinsed with fresh RPMI medium (Gibco; Thermo Fisher
Scientific, Inc.). The extremities of the femurs and tibiae were
removed with scissors, and the bone marrow was flushed using 5–10
ml medium with a 19-gauge syringe. A lysis buffer containing
ammonium chloride (KHCO3 1.0 g/l; NH4CL 8.3
g/l; EDTA-Na2 0.037 g/l) was used to lyse red blood
cells and the samples were washed twice to isolate the bone marrow
cells. Bone marrow cells were resuspended in RPMI medium
supplemented with recombinant rat IL-4 (10 ng/ml; PeproTech, Inc.)
and recombinant rat granulocyte monocyte colony-stimulating factor
(GM-CSF; 10 ng/ml; PeproTech, Inc.), and cultured at a density of
1×106 cells/ml in six-well plates with 3 ml culture
medium/well at 37°C in a 5% CO2 humidified incubator.
Fresh culture medium and cytokines were added at day 3. Cell
aggregates attached to the dish surface were observed between day 3
and 4. The culture medium was removed and replaced by fresh medium
with GM-CSF at day 5 and 6.
The phenotype of the cultured dendritic cells was
examined by investigating the protein expression levels of α E2
integrin (OX62) and MHC-II, as assessed by flow cytometry (Fig. S1), conducted as follows: One well
of a 6-well plate was centrifuged at 300 × g for 5 min at 37°C, and
the cell pellet was collected and resuspended in 5X volume of
washing solution (0.01 M PBS). The cells were rinsed twice and
resuspended in 200 µl of staining buffer (cat. no. 420201;
BioLegend, Inc.) at a concentration of 1×106 cells/ml.
The cells were equally divided into two 1.5-ml centrifuge tubes;
one tube was an experimental group, whereas the other was a blank
group. Phycoerythrin (PE)-conjugated anti-MHC class II (1:50; cat.
no. 12-0920-82; Invitrogen; Thermo Fisher Scientific, Inc.) and
PE-conjugated anti-Ox62 (1:50; cat. no. 12-1030-82; Invitrogen;
Thermo Fisher Scientific, Inc.) antibodies were added separately
and incubated for 30 min at room temperature in the dark; the blank
group for each assay was incubated without primary antibody. MHC-II
and OX62 were directly detected using an Accuri C6 flow cytometer
(BD Biosciences); the blank group was used to determine the
negative region. The test was repeated three times.
Transfection of imDCs with short
hairpin RNA (shRNA) targeting CCR7
The sequences of the shRNA targeting CCR7 and the
control sequences were as follows: shRNA-CCR7,
5′-TGGATCTTTGGTGCCTACCTGTGTA-3′; control shRNA,
5′-TTCTCCGAACGTGTCACGTAA-3′. shRNA-CCR7 and control shRNA was
inserted into a pHBAd-U6-GFP backbone, which was packaged into an
adenovirus. The 293A cells were transfected with the shCCR7
plasmid, and the adenovirus was harvested. The plasmid, shRNA and
adenovirus were all supplied by Hangzhou Hibio Technology Co., Ltd.
As control, an empty vector was used, and the infection efficiency
was assessed by western blot analysis (Fig. S2). Then, 1×106 imDCs
were suspended in 1 ml adenoviral supernatant (MOI=50) with 1% FBS
(Invitrogen; Thermo Fisher Scientific, Inc.), 10 ng/ml GM-CSF and
10 ng/ml IL-4 at 37°C for 72 h. Subsequently, cells were
centrifuged at 300 × g at 37°C for 2 h. After infection, imDCs were
washed twice in PBS and incubated with RPMI-1640 medium containing
10% fetal calf serum (FCS; Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10 ng/ml GM-CSF and 10 ng/ml IL-4 at 37°C with 5%
CO2. The cells were harvested for injection 48 h after
infection.
Transfection of imDC with adenovirus
overexpressing CCR7
NotI and PstI restriction sites were
added to the ends of the CCR7 coding sequence, which was cloned
into a pDC316-MCMV-ZsGreen backbone, which was packaged into an
adenovirus. The recombinant adenoviral vector containing the rat
CCR7 gene was prepared as previously described by Hibio
Technology Co., Ltd. (20), and
the centrifugal enhancement method was used to increase the
infection efficiency (21). Empty
plasmid vector was used as negative control. Viral packaging was
verified by the detection of green fluorescent protein via
fluorescence microscopy (data not shown). Subsequently,
1×106 imDCs were suspended with 1% FBS, 10 ng/ml GM-CSF
and 10 ng/ml IL-4, adding the adenovirus at MOI=50, and cells were
subsequently centrifuged at 300 × g at 37°C for 2 h. After
infection, the imDCs were washed twice in PBS and incubated with
RPMI-1640 medium containing 10% FCS supplemented with 10 ng/ml
GM-CSF and 10 ng/ml IL-4 at 37°C with 5% CO2. The cells
were harvested for injection 48 h after transfection. The infection
efficiency of CCR7 was determined by western blot analysis
(Fig. S3).
Establishment of animal models
Healthy SPF male Wistar rats were used to establish
an animal model of allergic asthma, according to a modified
previously described protocol (22). Rats were immunized by
intraperitoneal injection of 100 µg ovalbumin (OVA; Sigma-Aldrich;
Merck KGaA) adsorbed into 400 µg aluminium hydroxide in 0.2 ml
sterile saline (0.9%) on day 1 and 8. After 2 weeks, rats were
exposed to aerosol spray containing 1% OVA in 0.9% sterile saline
for 30 min every day, 6 days every week, for a total of 8 weeks.
The experimental rats were divided into three groups (n=10 in each
group) as follows: i) Control (Con) group, rats injected with
2×105 cultured wild-type imDCs; ii) CCR7 overexpression
(Over) group, rats injected with 2×105 cultured imDCs
infected with adenoviral particles overexpressing CCR7; and iii)
CCR7 interference (Sh) group, containing rats injected with
2×105 cultured imDCs infected with adenoviral particles
encoding a shRNA targeting CCR7. imDCs were injected via the tail
vein at day 1, 10 and 18.
Immunohistochemistry of CCR7 and
OX62
For immunohistochemical staining, lung tissues were
fixed at room temperature for 5 days using 4% formaldehyde
solution, embedded in paraffin and sectioned (4-µm), dewaxed in
100% xylene (twice in total, 20 min each time), rehydrated in a
graded alcohol series (100% for 5 min, 95% for 5 min, 80% for 5
min), and incubated with 0.3% H2O2 in
methanol to block the endogenous peroxidase activity for 15 min at
37°C. Mounted sections were boiled in 10 mM citrate solution (pH
6.0) for 20 min for antigen retrieval, and incubated overnight at
4°C with the following primary antibodies: Anti-rat CCR7 (1:50;
cat. no. bs-1305R; Bioss, Inc.) and anti-rat OX62 (1:50; cat. no.
sc-53085; Santa Cruz Biotechnology, Inc.). Horseradish
peroxidase-conjugated secondary antibody (1:25; cat. no. PV-6001;
Beijing Zhongshan Golen Bridge Biotechnology Co., Ltd.; OriGene
Technologies, Inc.) was incubated with the sections at 37°C for 30
min. A two-step technique (SuperPicture Third Generation IHC
Detection kit; Invitrogen; Thermo Fisher Scientific, Inc.) was used
to visualize the stained samples, and 0.1 mg/ml
3,3′-diaminobenzidine diluted in a 0.02% H2O2
solution (Vector Laboratories, Inc.) was used as chromogen for 2
min at room temperature. The tissue sections were counterstained
with hematoxylin (1 g/ml, 2 min) and eosin (5 g/l, 30 sec) at room
temperature, and observed using a light microscope (magnifications,
×100 and ×400; Olympus Corporation).
Western blot assay
Lung tissues were homogenized in RIPA buffer
(Pierce; Thermo Fisher Scientific, Inc.) and the protein
concentration was determined using the bicinchoninic acid protein
assay (Pierce; Thermo Fisher Scientific, Inc.). Total protein was
separated by SDS-PAGE on 10% gels. In total, 30 µg protein was
loaded in each lane. Proteins were then transferred to PVDF
membranes (EMD Millipore). Membranes were blocked for 2 h at room
temperature in 5% non-fat dry milk and incubated with a primary
antibody anti-rat CCR7 (1:1,000; cat. no. ab32527; Abcam) or
anti-OX62 (1:1,000; cat. no. bs-1274R; Bioss, Inc.) at 4°C
overnight. Membranes were then incubated with the appropriate
horseradish peroxidase-labeled anti-mouse [1:1,000; cat. no.
GAM007; MultiSciences (Lianke) Biotech Co., Ltd.] or anti-rabbit
IgG [1:1,000; cat. no. GAR0072; MultiSciences (Lianke) Biotech Co.,
Ltd.] secondary antibodies at room temperature for 50 min.
Anti-β-actin (1:1,000; cat. no. sc-47778; Santa Cruz Biotechnology
Inc.) or GAPDH (1:1,000; cat. no. sc-32233; Santa Cruz
Biotechnology Inc.) was used as the loading control. Bands were
visualized by autoradiography (ChemiDoc XR+System; Bio-Rad
Laboratories, Inc.) and quantified by densitometry (ImageJ V1.46;
National Institutes of Health). The results were normalized to
GAPDH or to β-actin. All experiments were performed in
triplicate.
Bronchoalveolar lavage fluid (BALF)
collection and cell counting
After the animal model was successfully established
(8 weeks), the animals were anesthetized and the lungs were
exposed. The right principal bronchus was occluded using a hemostat
clamp. A total of 4 ml saline was used for the bronchoalveolar
lavage in the left lung, and 3–4 ml of fluid was collected after
flushing twice. The alveolar lavage fluid was centrifuged at 300 ×
g for 10 min at room temperature, and the supernatant was used for
ELISA. Next, 0.5 ml saline was added to 0.5 ml BALF containing
sediment and mixed. Then, the samples were smeared and the cells
were counted using the Wright-Giemsa staining technique [Wright's
dyeing powder (1 g), Jimsa dyeing powder (0.5 g), 20 min
incubation, room temperature] and observed with a CX21 light
microscope (Olympus Corporation). Cells were counted under a
microscope using a hemocytometer and a counting place (25×16 grid
cells); the number of cells in each corner of the plate was
calculated.
ELISA
After the animal model was successfully established,
blood (4 ml) was extracted from the vena cava, anticoagulated with
heparin and centrifuged (500 × g for 10 min at room temperature),
and serum was collected. ELISA kits were used for measuring IFN-γ
(cat. no. EK0374), IL-4 (cat. no. EK0406), IL-12 (cat. no.
A01152-2), IL-10 (cat. no. EK0418), TGF-β (cat. no. EK0514; all
Boster Biological Technology) and IgE (cat. no. E-EL-R0517c;
Elabscience Biotechnology, Inc.) concentrations in serum and BALF
supernatant, according to the manufacturer's protocol.
Statistical analysis
All assays were repeated three times. The data are
presented as the mean ± SD. Statistical analysis was performed
using SPSS 11.5 software (SPSS, Inc.). One-way ANOVA was performed
to compare multiple groups, followed by Student-Newman-Keuls-Q
post-hoc test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Expression of CCR7 in lung tissues
following CCR7-overexpressing DC injection in vivo
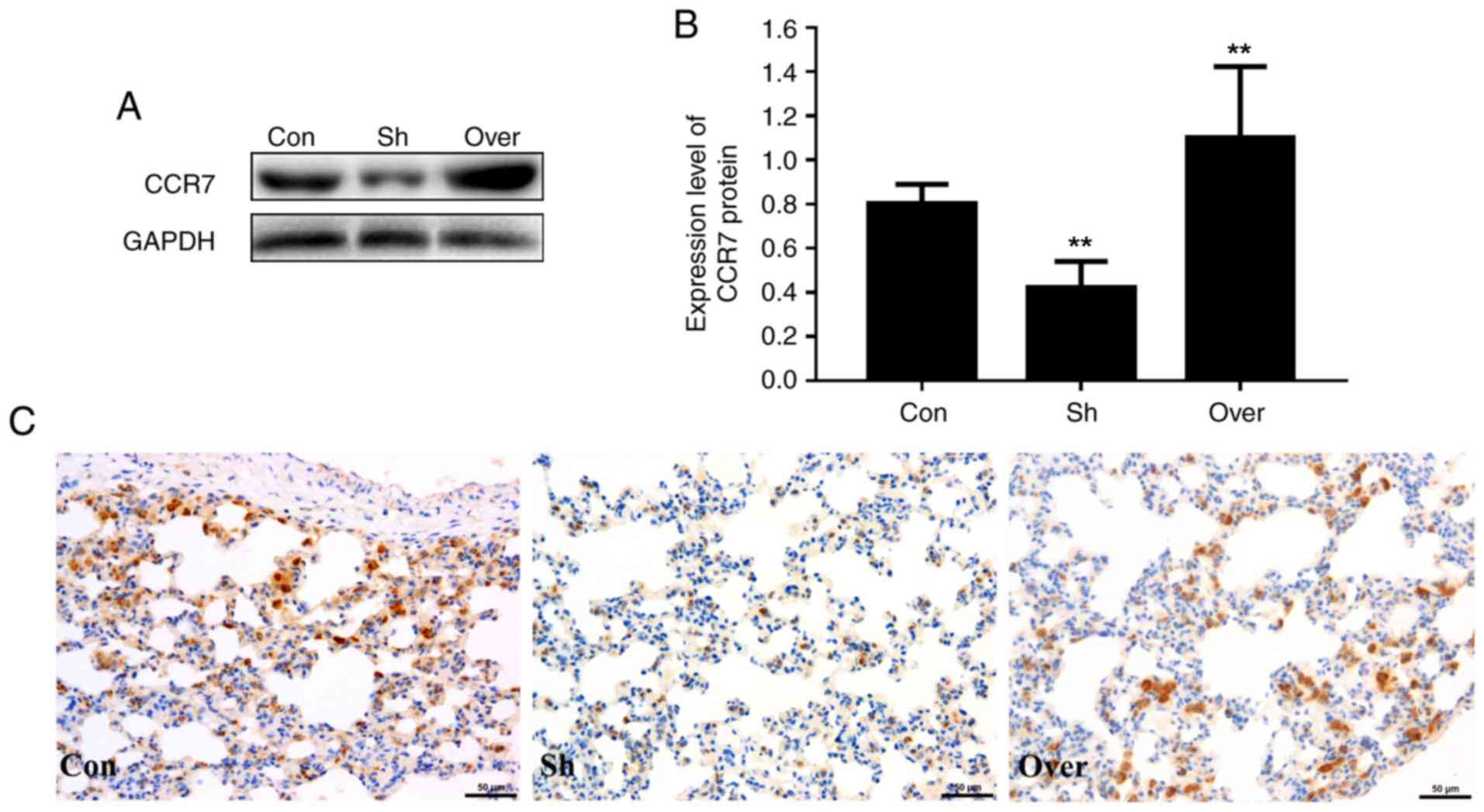
Immunohistochemical analysis suggested that the
protein expression level of CCR7 was increased in the airways of
rats in the CCR7-overexpressing group (Fig. 1). In addition, the cell membranes
were stained yellow and brown, indicating the localization of CCR7
in the peribronchial stroma (Fig.
1C). Compared with the control group, the protein expression of
CCR7 was significantly higher in the airways of rats in the
overexpression adenovirus-infected (Over) group and lower in the
shRNA adenovirus-infected (Sh) group (P<0.01; Fig. 1B). The protein expression levels of
CCR7 in various groups detected by western blotting was consistent
with the immunohistochemical results (Fig. 1A).
CCR7-overexpressing DCs promote
inflammatory cell infiltration in the lungs of rats with allergic
asthma
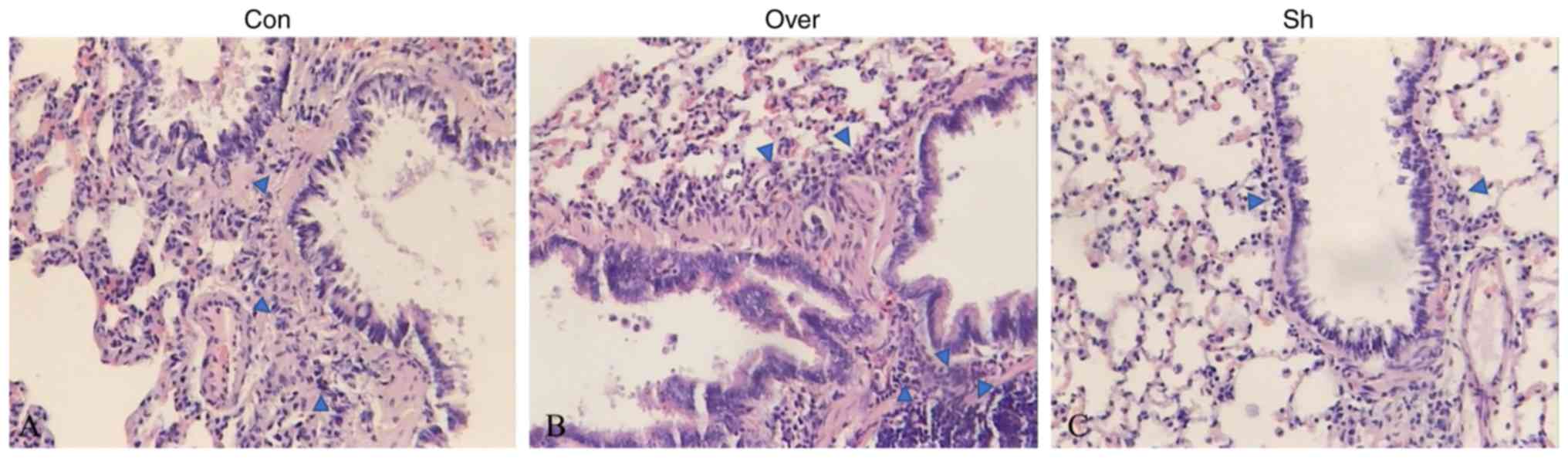
HE staining suggested the presence of numerous
inflammatory cells in the lung tissues in the control group
(Fig. 2). Additionally, the number
of infiltrated inflammatory cells, including lymphocytes,
eosinophils and neutrophils (as determined by cell morphology
following H&E staining), was increased in the airway and lung
tissues in the Over group compared with the Con group, and the
airway wall was markedly thicker in the CCR7 overexpression group
(Fig. S4). In the Sh group, the
bronchiole structure was normal and the number of inflammatory
cells infiltrating the lung tissue was reduced.
Expression of DC-specific antigens in
lung tissues is positively associated with the expression level of
CCR7
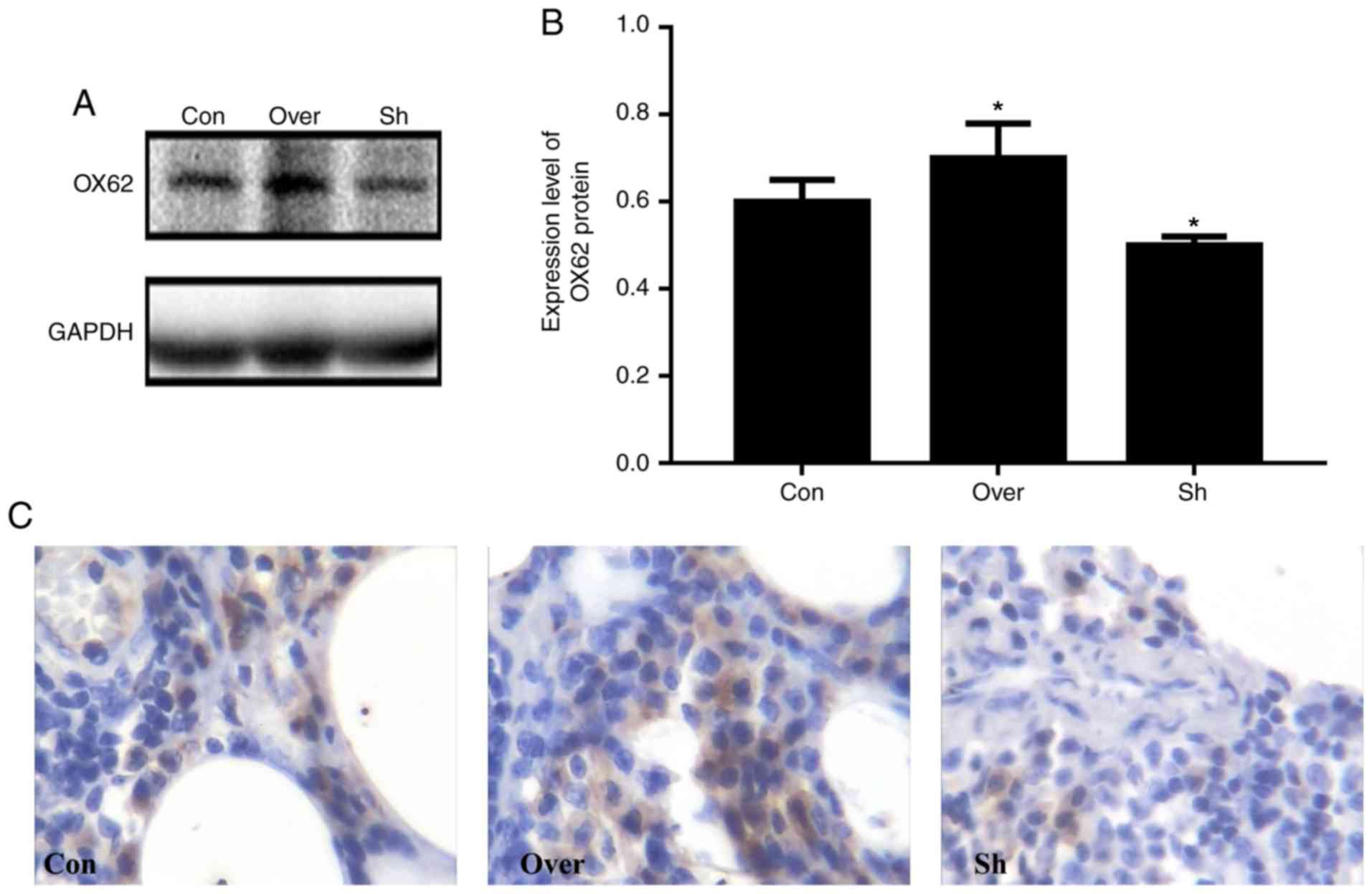
OX62 was used a marker for DC detection in the
airways as it is specifically expressed by DCs (23). The CCR7 Over group exhibited higher
expression levels of OX62 compared with the Con and Sh groups
(Fig. 3). In addition, the protein
expression level of OX62 was lower in the CCR7 interference group
compared with the control group (P<0.05).
Positive correlation between the
expression levels of CCR7 and the presence of leukocytes,
neutrophils and lymphocytes in BALF
The numbers of leukocytes (P<0.01), neutrophils
(P<0.05) and lymphocytes (P<0.01) were higher in the CCR7
Over group compared with the Con group and the Sh group (Fig. 4). Conversely, the expression of
eosinophils was not affected by CCR7 overexpression. The present
data suggested that CCR7 expression could affect the number and
types of immune cells recruited to the airways and infiltrating the
lung tissues.
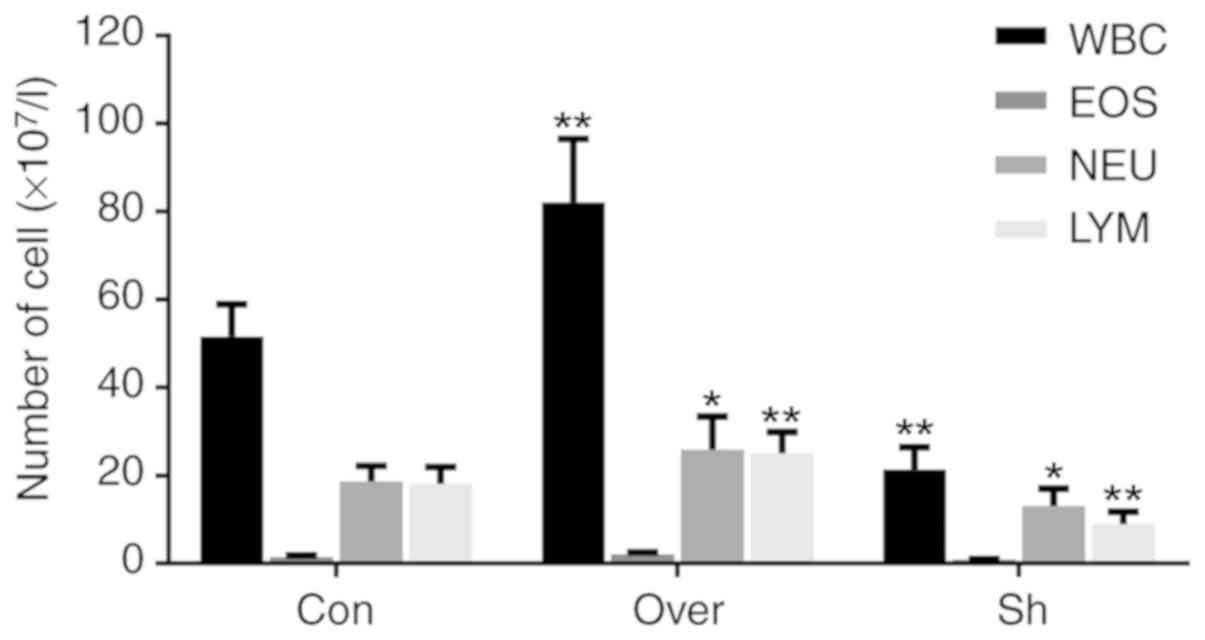 | Figure 4.Cell count and classification of
immune cells in bronchoalveolar lavage fluid. Numbers of WBC, NEU,
EOS and LYM in each group. *P<0.05 vs. Con; **P<0.01 vs. Con.
CCR7, CC chemokine receptor 7; Con, control group; Over, CCR7
overexpression group; Sh, CCR7 knockdown group; WBC, leukocytes;
EOS, eosinophil; NEU, neutrophils; LYM, lymphocytes. |
Expression levels of cytokines and IgE
in the BALF and serum are associated with the expression level of
CCR7
Compared with the control group, the protein
expression levels of IL-12, IL-4, IFN-γ and IgE, in both the BALF
supernatant (Fig. 5) and serum
(Fig. 6), were increased in the
CCR7 Over group compared with the Con and the Sh groups
(P<0.01). Compared with the control group, the protein
expression levels of IL-10 and TGF-β in the BALF supernatant
(Fig. 5) and serum (Fig. 6) were significantly lower in the
CCR7 Over group, and higher in the Sh group (P<0.01).
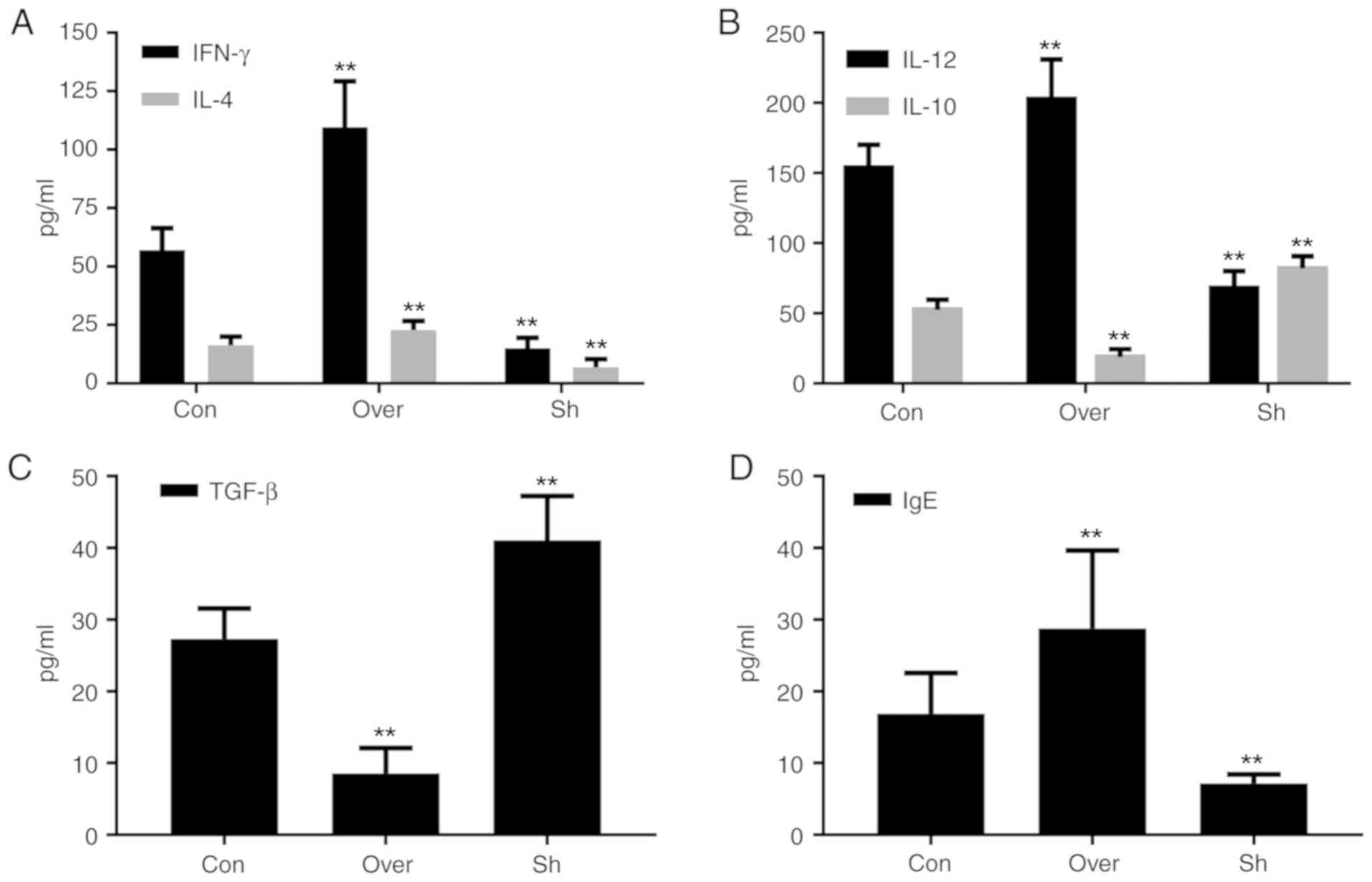 | Figure 5.Protein expression levels of (A)
IFN-γ and IL-4, (B) IL-12 and IL-10, (C) TGF-β and (D) IgE in
bronchoalveolar lavage fluid. Protein expression levels of various
cytokines were assessed by ELISA. **P<0.01 vs. Con. CCR7, CC
chemokine receptor 7; IFN-γ, interferon-γ; IL, interleukin; Con,
control group; Over, CCR7 overexpression group; Sh, CCR7 knockdown
group; TGF-β, transforming growth factor-β; IgE, immunoglobulin
E. |
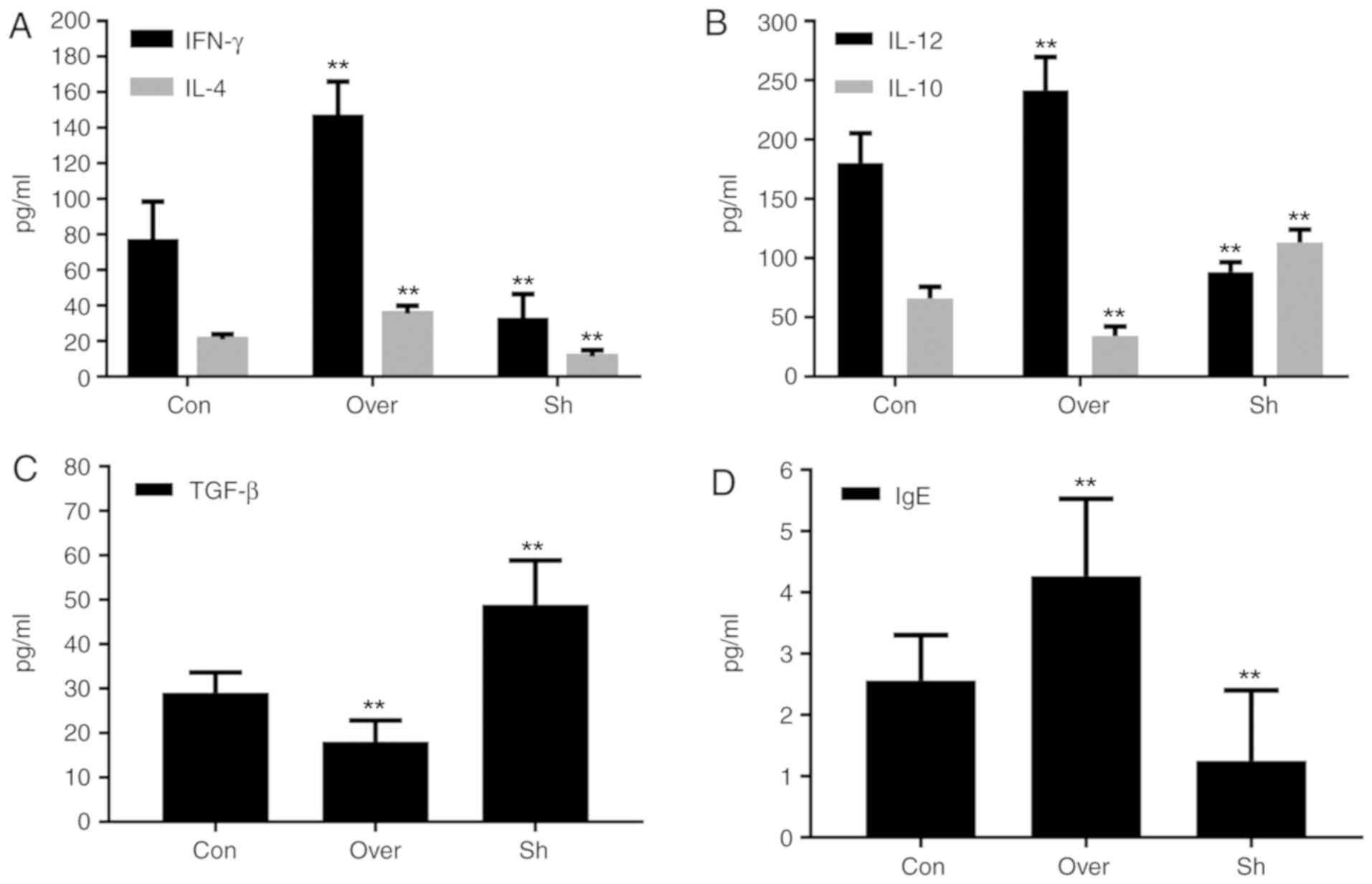 | Figure 6.Protein expressions of (A) IFN-γ and
IL-4, (B) IL-12 and IL-10, (C) TGF-β and (D) IgE in serum. Protein
expression levels of various cytokines were assessed by ELISA.
**P<0.01 vs. control group. CCR7, CC chemokine receptor 7;
IFN-γ, interferon-γ; IL, interleukin; Con, control group; Over,
CCR7 overexpression group; Sh, CCR7 knockdown group; TGF-β,
transforming growth factor-β; IgE, immunoglobulin E. |
Discussion
One of the most important functions of the immune
system is to prevent pathogen-associated damage, which is limited
through effective recognition of the exogenous antigens and
initiation of the immune response (24). The immune system allows the body to
maintain its internal environment via the immune tolerance
mechanism (25). Both immune
response and immune tolerance mechanisms serve important roles in
allergy-induced asthma (26).
Therefore, in the present study, the effects of CCR7 overexpression
and knockdown in DCs on the mechanisms of immune tolerance were
investigated in an animal model of allergic asthma.
In the present study, DC markers were examined, and
the expression levels of OX62 were decreased following
shRNA-mediated CCR7 knockdown in DCs. The present results suggested
that CCR7 may serve an important role in the regulation of the
immune response in DC-induced asthma. Previous studies have
demonstrated that imDCs can internalize, process and deliver
antigens, and it is also associated with the induction of immune
tolerance (9); however, mature DCs
exhibit immune stimulating abilities (9). Interestingly, CCR7 expression is
limited to mature DCs (27). imDCs
have been used as a model to study the association between changes
in the expression level of cytokines and the immune inflammatory
response in allergic asthma (28).
In the present study, HE and Wright-Giemsa staining of lung tissues
and BALF showed an increased number of infiltrating inflammatory
cell in the CCR7 overexpression group. The present findings
suggested that CCR7 may promote immune inflammation. Conversely,
there was no significant association between eosinophils and CCR7
expression; the increase and aggregation of eosinophils may be
related to other factors, such as IL-5 (29). However, a previous study showed
that CCR7-deficient animals fail to induce tolerance to inhaled
environmental innocuous antigens (17). The reasons underlying the
differences between the present and this previous study may be
multifactorial. Notably, the animal model is different; in the
present study, continuous inhalation of OVA was used to induce
bronchial asthma in rats, whereas the previous study established an
immune tolerance model via the inhalation of increasing doses of
the antigens. Moreover, the present study used a rat allergy model,
whereas the previous study was performed in mice. In addition, the
CCR7-deficient animals in the previous study may exhibit additional
defects, such as impaired T cell recirculation, that may influence
the immune system in these animals.
CCR7 possesses a variety of functions and properties
in DCs. A previous study has shown that knockdown of CCR7 increases
the expression levels of CD80, CD86, IFN-γ, IL-12/23 and IL-1α in
DC (30). CCR7 silencing also
increases the resistance to infection by increasing the number of
neutrophils in the lung airways (31). In the present study, various
cytokines, including IgE, were found to be involved in
allergy-induced asthma and their expression level decreased
following knockdown of CCR7. The present results suggested that a
decrease in the expression level of CCR7 may suppress the immune
response in patients with allergy-induced asthma. Accumulating
evidence demonstrated that the mucosa in asthmatic airways contains
a large number of activated Th cells, as well as higher levels of
IL-4 and IL-5. In vitro studies have shown that IL-4 can
induce B lymphocytes to synthesize IgE, promoting airway
hyper-responsiveness (32–34), although IFN-γ inhibits this effect
(35). IL-12 is a major cytokine
that regulates immune balance by promoting the expression of
certain cytokines, such as IFN-γ, and inhibiting the secretion of
other cytokines, such as IL-4 and IL-5 (36). Therefore, IFN-γ, IL-4, IL-12 and
IgE serve important roles in the pathogenesis of allergy-induced
asthma. Overall, decreased CCR7 expression not only reduced
inflammatory cell infiltration, but also decreased the expression
levels of various inflammatory factors.
IL-10 and TGF-β are cytokines involved in the
process of immune tolerance (37,38).
IL-10 exhibits a wide range of functions, including the inhibition
of Th2 cytokine and IgE production and it is involved in decreasing
mast cell and eosinophil function (39). In addition, IL-10 increases the
secretion of IgG4 and regulates the IgG4/IgE ratio (40). TGF-β is a pro-inflammatory cytokine
that regulates lymphocyte homeostasis, suppresses Th2 cell
activation and promotes Treg cell differentiation (41). In the present study, CCR7 knockdown
increased the protein expression levels of IL-10 and TGF-β in
allergy-induced asthma, suggesting that CCR7 may serve an important
role in immune tolerance in allergy-induced asthma. The induction
of T cell antigen-specific immune tolerance may represent a novel
strategy for the treatment of various immune inflammatory diseases,
including allergy-induced asthma.
The immune tolerance to allergens in asthmatic
patients is due to the activation and proliferation of Th cells
(26). Therefore, mechanisms
underlying immune tolerance defects may be important for the
pathogenesis of allergy-induced asthma. Knockdown of CCR7 in DCs
caused the cells to remain in an immature state, promoting immune
tolerance. This effect may further reduce the activity of DCs,
leading to decreases in the expression levels of cytokines involved
in the immune response in allergy-induced asthma, and increases in
the expression levels of cytokines involved in immune
tolerance.
CCR7 has important roles in DC-mediated immune
inflammation and immune tolerance in patients with allergy-induced
asthma (17). Notably, the present
study presents certain limitations, since the expression level of
CCR7 in DCs was investigated only in the lungs. In the future, it
may be useful to examine the role of CCR7-expressing DCs in other
tissues prone to allergic inflammation. Moreover, further studies
are required to examine the CCR7-dependent chemotaxis and
CCR7-mediated signal transduction pathways, which may provide
insights into novel therapeutic approaches for the treatment of
patients with allergic asthma.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by The Shanxi
Scholarship Council of China (grant no. 2014-Focus 8).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YL designed and performed the majority of the study
and data analysis, and drafted the manuscript. YD contributed to
the conception and design of the study. AZ provided pathological
assistance, and was involved in the data analysis and
interpretation. RJ, XN and XX contributed to interpretation of the
data and analyses. All of the authors have read and approved the
manuscript.
Ethics approval and consent to
participate
The present study was approved by The Ethics
Committees of The People's Hospital of Shanxi Province.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bateman ED, Hurd SS, Barnes PJ, Bousquet
J, Drazen JM, FitzGerald JM, Gibson P, Ohta K, O'Byrne P, Pedersen
SE, et al: Global strategy for asthma management and prevention:
GINA executive summary. Eur Respir J. 31:143–178. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tesse R, Borrelli G, Mongelli G,
Mastrorilli V and Cardinale F: Treating pediatric asthma according
guidelines. Front Pediatr. 6:2342018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Schatz M and Rosenwasser L: The allergic
asthma phenotype. J Allergy Clin Immunol Pract. 2:645–648. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lambrecht BN and Hammad H: The immunology
of asthma. Nat Immunol. 16:45–56. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chairakaki AD, Saridaki MI, Pyrillou K,
Mouratis MA, Koltsida O, Walton RP, Bartlett NW, Stavropoulos A,
Boon L, Rovina N, et al: Plasmacytoid dendritic cells drive acute
asthma exacerbations. J Allergy Clin Immunol. 142:542–556.e12.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lambrecht BN and Hammad H: The role of
dendritic and epithelial cells as master regulators of allergic
airway inflammation. Lancet. 376:835–843. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lombardi V, Singh AK and Akbari O: The
role of costimulatory molecules in allergic disease and asthma. Int
Arch Allergy Immunol. 151:179–189. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kuchroo VK, Das MP, Brown JA, Ranger AM,
Zamvil SS, Sobel RA, Weiner HL, Nabavi N and Glimcher LH: B7-1 and
B7-2 costimulatory molecules activate differentially the Th1/Th2
developmental pathways: Application to autoimmune disease therapy.
Cell. 80:707–718. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kadowaki N: Dendritic cells: A conductor
of T cell differentiation. Allergol Int. 56:193–199. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Webb LM and Tait Wojno ED: The role of
rare innate immune cells in Type 2 immune activation against
parasitic helminths. Parasitology. 144:1288–1301. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lambrecht BN, Persson EK and Hammad H:
Myeloid cells in Asthma. Microbiol Spectr. 5:MCHD–0053-2016. 2017.
View Article : Google Scholar
|
|
12
|
Thomas SY, Whitehead GS, Takaku M, Ward
JM, Xu X, Nakano K, Lyons-Cohen MR, Nakano H, Gowdy KM, Wade PA and
Cook DN: MyD88-dependent dendritic and epithelial cell crosstalk
orchestrates immune responses to allergens. Mucosal Immunol.
11:796–810. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mims JW: Asthma: Definitions and
pathophysiology. Int Forum Allergy Rhinol. 1 (Suppl 5):S2–S6. 2015.
View Article : Google Scholar
|
|
14
|
Hauser MA and Legler DF: Common and biased
signaling pathways of the chemokine receptor CCR7 elicited by its
ligands CCL19 and CCL21 in leukocytes. J Leukoc Biol. 99:869–882.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Förster R, Davalos-Misslitz AC and Rot A:
CCR7 and its ligands: Balancing immunity and tolerance. Nat Rev
Immunol. 8:362–371. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wei G, Jie Y, Haibo L, Chaoneng W, Dong H,
Jianbing Z, Junjie G, Leilei M, Hongtao S, Yunzeng Z and Junbo G:
Dendritic cells derived exosomes migration to spleen and induction
of inflammation are regulated by CCR7. Sci Rep. 7:429962017.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Akdis CA and Akdis M: Mechanisms of
allergen-specific immunotherapy and immune tolerance to allergens.
World Allergy Organ J. 8:172015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hintzen G, Ohl L, del Rio ML,
Rodriguez-Barbosa JI, Pabst O, Kocks JR, Krege J, Hardtke S and
Förster R: Induction of tolerance to innocuous inhaled antigen
relies on a CCR7-dependent dendritic cell-mediated antigen
transport to the bronchial lymph node. J Immunol. 177:7346–7354.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tiurbe G, Matuschek A, Kämmerer U,
Schneider M, Thiede A, Ulrichs K and Otto C: Inhibitory effects of
rat bone marrow-derived dendritic cells on naïve and
alloantigen-specific CD4+ T cells: A comparison between
dendritic cells generated with GM-CSF plus IL-4 and dendritic cells
generated with GM-CSF plus IL-10. BMC Res Notes. 2:122009.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xin HM, Peng YZ, Yuan ZQ and Guo H: In
vitro maturation and migration of immature dendritic cells after
chemokine receptor 7 transfection. Can J Microbiol. 55:859–866.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Vaysse L and Arveiler B: Transfection
using synthetic peptides: Comparison of three DNA-compacting
peptides and effect of centrifugation. Biochim Biophys Acta.
1474:244–250. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Martin JG and Tamaoka M: Rat models of
asthma and chronic obstructive lung disease. Pulm Pharmacol Ther.
19:377–385. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Brenan M and Puklavec M: The MRC OX62
antigen: A useful marker in the purification of rat veiled
cellswith the biochemical properties of an integrin. J Exp Med.
175:1457–1465. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Delgado M and Gonzalez-Rey E: Role of
cortistatin in the stressed immune system. Front Horm Res.
48:110–120. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lübbers J, Rodríguez E and van Kooyk Y:
Modulation of immune tolerance via siglec-sialic acid interactions.
Front Immunol. 9:28072018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Głobińska A, Boonpiyathad T, Satitsuksanoa
P, Kleuskens M, van de Veen W, Sokolowska M and Akdis M: Mechanisms
of allergen-specific immunotherapy: Diverse mechanisms of immune
tolerance to allergens. Ann Allergy Asthma Immunol. 121:306–312.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Dieu MC, Vanbervliet B, Vicari A, Bridon
JM, Oldham E, Aït-Yahia S, Brière F, Zlotnik A, Lebecque S and Caux
C: Selective recruitment of immature and mature dendritic cells by
distinct chemokines expressed in different anatomic sites. J Exp
Med. 188:373–386. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Saeidi M, Masoud A, Shakiba Y, Hadjati J,
Mohyeddin Bonab M, Nicknam MH, Latifpour M and Nikbin B:
Immunomodulatory effects of human umbilical cord Wharton's
jelly-derived mesenchymal stem cells on differentiation, maturation
and endocytosis of monocyte-derived dendritic cells. Iran J Allergy
Asthma Immunol. 12:37–49. 2013.PubMed/NCBI
|
|
29
|
Furuta GT, Atkins FD, Lee NA and Lee JJ:
Changing roles of eosinophils in health and disease. Ann Allergy
Asthma Immunol. 113:3–8. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Qian J, Xu X, Ding J, Yin R, Sun Y, Xue C,
Wang J, Ding C, Yu S, Liu X, et al: Newcastle disease virus-like
particles induce DC maturation through TLR4/NF-κB pathway and
facilitate DC migration by CCR7-CCL19/CCL21 axis. Vet Microbiol.
203:158–166. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kling JC, Mack M and Körner H: The absence
of CCR7 results in dysregulated monocyte migration and
immunosuppression facilitating chronic cutaneous leishmaniasis.
PLoS One. 8:e790982013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Robinson DS, Hamid Q, Ying S, Tsicopoulos
A, Barkans J, Bentley AM, Corrigan C, Durham SR and Kay AB:
Predominant TH2-like bronchoalveolar T-lymphocyte population in
atopic asthma. N Engl J Med. 326:298–304. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Oh CK, Geba GP and Molfino N:
Investigational therapeutics targeting the IL-4/IL-13/STAT-6
pathway for the treatment of asthma. Eur Respir Rev. 19:46–54.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Schoenborn JR and Wilson CB: Regulation of
interferon-gamma during innate and adaptive immune responses. Adv
Immunol. 96:41–101. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gouveia ACC, Braga FG, Mota M, Silva FMC,
Brugiolo ASS, Oliveira EE, Ayupe MC, Teixeira HC and Ferreira AP:
Enhanced expression of PD-L1 and IFN-γ on dendritic cells is
associated with BCG-induced Th2 inhibition. Cytokine. 99:163–172.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zheng H, Ban Y, Wei F and Ma X: Regulation
of interleukin-12 production in antigen-presenting cells. Adv Exp
Med Biol. 941:117–138. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Carrier Y, Yuan J, Kuchroo VK and Weiner
HL: Th3 cells in peripheral tolerance. I. Induction of
Foxp3-positive regulatory T cells by Th3 cells derived from
TGF-beta T cell-transgenic mice. J Immunol. 178:179–185. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ng TH, Britton GJ, Hill EV, Verhagen J,
Burton BR and Wraith DC: Regulation of adaptive immunity; the role
of interleukin-10. Front Immunol. 4:1292013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jutel M, Akdis M, Budak F,
Aebischer-Casaulta C, Wrzyszcz M, Blaser K and Akdis CA: IL-10 and
TGF-beta cooperate in the regulatory T cell response to mucosal
allergens in normal immunity and specific immunotherapy. Eur J
Immunol. 33:1205–1214. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Soyer OU, Akdis M and Akdis CA: Mechanisms
of subcutaneous allergen immunotherapy. Immunol Allergy Clin North
Am. 31:175–190, vii-viii. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Martín-Orozco E, Norte-Muñoz M and
Martínez-García J: Regulatory T cells in allergy and Asthma. Front
Pediatr. 5:1172017. View Article : Google Scholar : PubMed/NCBI
|