Introduction
During aging, a number of physiological functions
are altered and the cessation of cell division is accompanied by
specific changes in cell function, morphology and gene expression.
These changes may contribute to age-associated diseases, including
hypertension, chronic coronary disease and diabetes (1). Vascular endothelial cells (VEC) are
highly specialized and active cells exhibiting antithrombotic and
anti-inflammatory properties. These cells are critically involved
in the maintenance of vascular homeostasis by regulating vascular
tone and integrity, as well as remodeling processes (2). Vascular cell senescence, which
accompanies aging, promotes endothelial cell dysfunction (3) and is associated with increased
vascular risk (4). Pathological
states, including those observed in oxidative stress conditions,
invoke irreversible growth arrest in vitro within a few
days, a term referred to as stress-induced premature senescence
(SIPS) (5,6). Previous evidence suggests that
premature senescence of endothelial cells may lead to endothelial
dysfunction and atherogenesis (3).
Hydrogen sulfide (H2S) has attracted
considerable interest as an endogenous gaseous mediator and
potential pharmacological/therapeutic tool. It is endogenously
generated from cysteine, in reactions catalyzed by cystathionine
β-synthase (CBS) or cystathionine γ-lyase (CSE) (7). Studies in various species, including
humans have demonstrated that H2S is involved in diverse
physiological and pathophysiological processes, including
regulation of blood pressure (8),
inflammation (9) and metabolic
disorders (10). The beneficial
effects of H2S may be mediated through its antioxidant
effects. H2S is a potent inhibitor of
O2- formation and gp91phox expression induced
by TNF-α in pulmonary artery endothelial cells (PAECs) (11). Furthermore, H2S protects
endothelial cells against oxidized low-density lipoprotein (LDL)
and hydrogen peroxide (H2O2)-mediated cell
cytotoxicity (12). H2S
also appears to be a potent scavenger of oxygen-derived free
radicals (13), which may
contribute to the protective role of NaHS against the toxicity of
H2O2in vitro and in
vivo(14). Findings of a
previous study from our laboratory indicated that the endogenous
CSE/H2S system is downregulated in adipose tissues
during aging (15). Another study
reported that thermotolerance and lifespan of Caenorhabditis
elegans (C. elegans) was increased when exposed to
H2S, which was mediated by SIR-2.1 activity (16).
SIR-2.1 is a C. elegans ortholog to sirtuin 1
(SIRT1), which is commonly known as nicotinamide adenine
dinucleotide (NAD+)-dependent class III histone
deacetylase. This enzyme has been shown to modulate lifespan in
yeast, worms, flies and mice. A mammalian SIRT1 homolog, silent
information regulator-2 (Sir2), is highly conserved in organisms
ranging from archaea to humans and it has been shown to regulate
cell cycle, senescence, apoptosis and metabolism by interacting
with a number of molecules, including p53 (17) and Foxo1 (18). A previous study demonstrated that
SIRT1 inhibition induces premature senescence-like growth arrest in
human cancer cells (19). The aim
of the present study was to investigate the effects of
H2S, using the donor NaHS, on the inhibition of
H2O2-induced senescence in human umbilical
vein endothelial cells (HUVECs) and the role of SIRT1 in this
process.
Materials and methods
Materials
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide
(MTT), dimethyl sulfoxide (DMSO), collagenase II, sodium
hydrosulfide (NaHS) and DL-propargylglycine (PPG) were purchased
from Sigma Aldrich (Zwijndrecht, Netherlands). Endothelial cell
growth supplement (ECGS) was purchased from ScienCell Research
Laboratories, Inc. (Carlsbad, CA, USA). p21 and SIRT1 antibodies
and the Senescence β-Galactosidase Staining Kit were purchased from
the Beyotime Institute of Biotechnology (Shanghai, China). Luminol
reagent and polyvinylidene fluoride (PVDF) membranes were purchased
from Millipore (Billerica, MA, USA).
Cell cultures
HUVECs were isolated from newborn umbilical cords
and cultured in M199 (Gibco, Grand Island, NY, USA) supplemented
with 20% fetal bovine serum (FBS) and 2% EGCS at 37°C under 5%
CO2 in a humidified atmosphere. Cells were used during
passages two or three.
Immunohistochemistry
Immunohistochemical staining of the HUVECs for
factor VIII-like antigen (fVIII-AGN) was performed on confluent
cultures grown in 35-mm dishes. The media was aspirated and the
cells were washed with phosphate-buffered saline (PBS) prior to
fixing in bovine serum albumin (BSA) for 15 min at room
temperature. The cells were washed again with PBS and incubated
with a 1:40 (v/v) dilution of horseradish peroxidase
(HRP)-conjugated rabbit anti-human fVIII:AGN (Bioss Inc., Woburn,
MA, USA) for 45 min. The dishes were washed in triplicate with PBS,
rinsed briefly with distilled water and mounted with buffered
glycerol on glass cover slips. HRP-bound primary antibody was
detected and observed using 3,3′-diaminobenzidine (DAB). Smooth
muscle cells served as negative controls.
Senescence-associated β-galactosidase
(SA-β-gal) staining
SA-β-gal activity was measured using a senescence
cell staining kit. HUVECs were pretreated with various
concentrations of NaHS, 5 mM NAM or a combination of NaHS and NAM
for 48 h. The cells were then placed in media supplemented with 25
μM H2O2 for 1 h. The media were then replaced
with normal medium and incubated for an additional 72 h. The cells
were washed twice with PBS and the HUVECs were fixed and stained
for SA-β-gal activity using the Senescence β-Galactosidase Staining
Kit. The cells were then incubated at 37°C for 16 h and
SA-β-gal-positive cells were observed using microscopy, which
included counting >400 cells in three independent fields. The
percentage of SA-β-gal-positive cells was determined by counting
the number of green cells within a sample (20).
Cell cycle assay
To determine the effect of
H2O2 on cell cycle progression, HUVECs were
grown for 1 h with or without 25 μmol/l H2O2.
Cells were collected using trypsinization and centrifugation for 5
min at 300 × g and were fixed with 70% ethanol at 4°C overnight.
Cells were centrifuged to remove alcohol, stained with 50 mmol/l
propidium iodide and washed twice with cold PBS. HUVECs were
subjected to flow cytometric analyses with FACSCalibur and
CellQuest software (BD Biosciences, Franklin Lakes, NJ, USA). Cell
cycles were analyzed and the proportion of cells in the
G0/G1, S and G2/M phases was
recorded.
Immunoblot analyses
Protein extracts were prepared using the mammalian
cell extraction kit following the manufacturer’s instructions.
Protein concentrations were determined using a BCA Protein Assay
kit (Pierce, Biotechnology, Inc., Rockford, IL, USA). Extracted
proteins were treated with 5% sodium dodecyl sulfate-polyacrylamide
gel electrophoresis (SDS-PAGE) sample buffer, then heated at 100°C
for 10 min and separated using electrophoresis on a 10%
SDS-polyacrylamide gel. Equal amounts of protein were separated
using SDS-PAGE and then transferred to PVDF membranes. The
membranes were incubated in a blocking buffer containing BSA (1%)
and Tween-20 (0.1%, v/v) in Tris-buffered saline with Tween-20
(TBST) at room temperature for 2 h, and inoculated overnight at 4°C
with the primary antibodies, anti-human β-actin (1:500) and
anti-human SIRT1 (1:300). The membranes were then inoculated with
goat anti-rabbit (1:3,000) and goat anti-mouse (1:1,500)
HRP-conjugated secondary antibodies at room temperature for 2 h.
Each membrane was developed using enhanced chemiluminescence
detection and quantified by densitometry.
Enzymatic activity assay
A SIRT1 enzyme activity assay was performed to
determine the effect of H2S on activity using a
commercially available kit (Genmed, Plymouth, MN, USA). After
preparing cell lysates, the SIRT1 activity assay was performed in a
96-well plate according to the manufacturer’s instructions. The
reaction product emitted fluorescence, which was detected using an
excitation wavelength of 350 nm and an emission wavelength of 405
nm.
Cell proliferation assay
Proliferation of HUVECs was determined using an MTT
assay (21). Forty-eight hours
after cell seeding, the media were removed and 210 μl fresh culture
media and 50 μl MTT solution (5 mg/ml in PBS) were added to each
well, followed by incubation for 2 h at 37°C in a 5% CO2
atmosphere. The cells were cultured for 4 h at 37°C in a 5%
CO2 atmosphere and the optical density of the solution
was evaluated using a microplate spectrophotometer at 595 nm.
Cell scratch assay
To determine the functional consequences of
senescence induced by H2O2, the in
vitro scratch injury model was used. Cells were seeded in a
96-well plate and treated 24 h after seeding. Twenty-four hours
after treatment, a thin-line, devoid of cells, was made by
scratching the culture plate bottom with a 10 μl pipette tip.
Following scratching, the wells were washed with PBS and fresh
media were added. Two images were captured per well; the width of
the scratch was measured at four points per image with Image-Pro
Plus and the means were calculated.
Statistical analyses
Results are expressed as mean ± standard error of
the mean (SEM) or mean ± standard deviation (SD). Differences
between groups were evaluated using analysis of variance, Dunnett’s
test or the least significant difference t-test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Establishing a senescence model in
HUVECs
To investigate the effect of H2S on HUVEC
senescence, we utilized an established senescence model, which
involved incubating the cells with 25 μmol/l
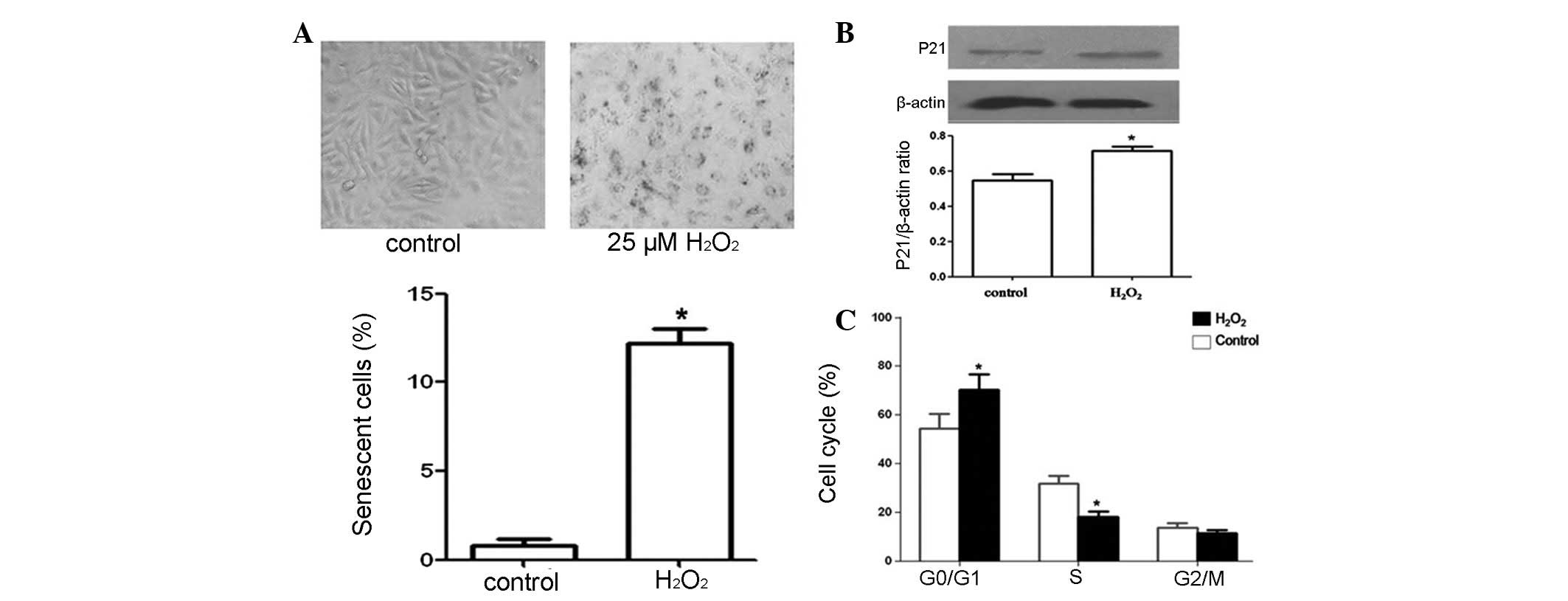
H2O2 for 1 h (22). Using light microscopy, we confirmed
the presence of senescent HUVECs, which exhibited increased cell
size and cytoplasmic granularity (Fig.
1A). The number of SA-β-gal-positive cells (Fig. 1A) and the proportion of HUVECs in
the G0/G1 phase (Fig. 1C) were increased, indicating the
presence of senescent cells. In order to confirm our results, we
investigated p21 levels, which were increased in senescence
(23). Immunoblot analyses
indicated that p21 levels were increased in HUVECs treated with
H2O2 (Fig.
1B). Collectively, these results indicate that the low
concentration of H2O2 used in our study
induced cell senescence.
H2S protects against HUVEC
senescence
SA-β-gal is a well-accepted biochemical marker of
cell senescence (24). Examination
of SA-β-gal activity in HUVECs treated with
H2O2 (25 μM) revealed a significant increase
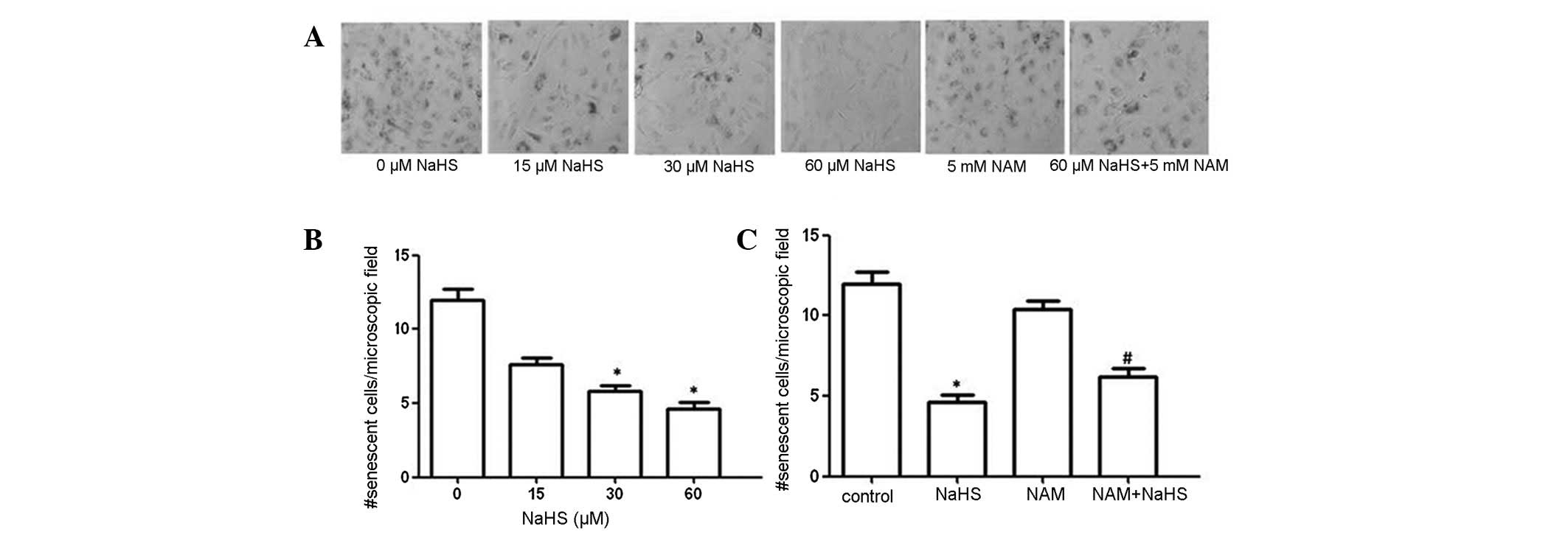
in SA-β-gal-positive cells, which reached 11.2±1.06% (Fig. 1A). However, increases in
SA-β-gal-positive cells were significantly attenuated in the NaHS
(60 μM) group (Fig. 2A and B).
Progression through the cell cycle is a critical
cellular process and cell cycle arrest during the G1
phase is a characteristic exhibited by senescent cells. Our results
demonstrated that treatment with 25 μM H2O2
arrested HUVECs in the G0/G1 phase as the
proportion of cells in the G0/G1 phase was
~70.2% compared to 54.4% in the control group. NaHS (60 μM)
pretreatment eliminated the effects of H2O2
and reduced the proportion of cells in the
G0/G1 phase to 58.1% (Table I). These results indicate that
H2S protects against HUVEC senescence.
 | Table IEffect of H2S on cell
cycle arrest of HUVECs induced by 25 μM
H2O2. |
Table I
Effect of H2S on cell
cycle arrest of HUVECs induced by 25 μM
H2O2.
| Group |
G0/G1 (%) | S (%) | G2/M
(%) |
|---|
| Control | 54.41±5.83 | 31.83±3.26 | 13.76±1.85 |
|
H2O2 | 70.24±6.31a | 18.21±2.11 | 11.55±1.23 |
| NaHS | 58.16±5.34 | 29.08±2.52 | 12.91±1.36 |
|
H2O2+NaHS | 53.72±5.12b | 32.14±3.43 | 14.14±1.69 |
H2S enhances the activation of
SIRT1
To determine whether H2S regulates HUVEC
senescence through a SIRT1-mediated pathway, we examined the
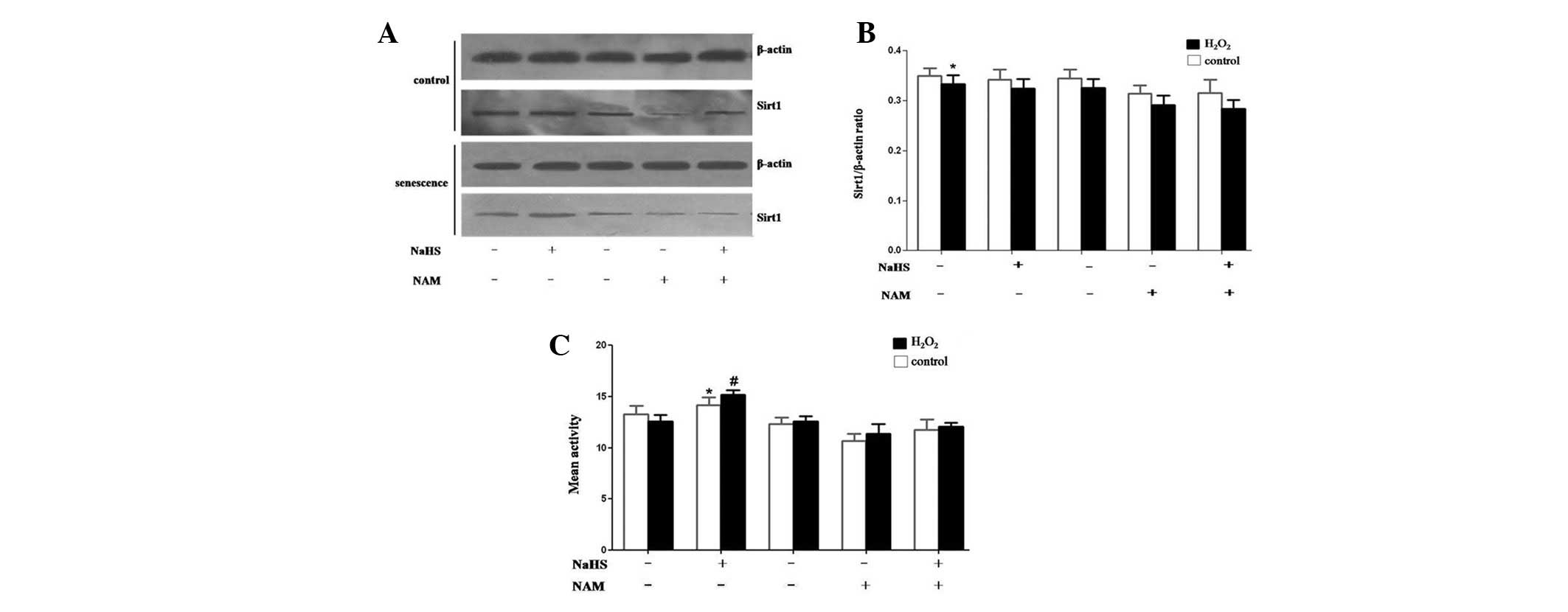
expression and activity of SIRT1. Immunoblot analyses indicated
that SIRT1 levels were decreased in the H2O2
(25 μM) treatment group compared to the control, and NaHS (60 μM)
treatment did not rescue SIRT1 expression (Fig. 3A and B). In contrast to its effect
on protein expression, NaHS enhanced SIRT1 deacetylase activity
in vitro (Fig. 3C),
indicating a direct effect on SIRT1-mediated pathways. These
results suggest that NaHS blocks senescence, cell differentiation
and stress-induced apoptosis, and promotes cell growth by
increasing SIRT1 deacetylase activity.
Inhibition of SIRT1 by NAM attenuates the
anti-senescent effects of H2S
To elucidate the role of SIRT1, HUVECs were
pretreated with NaHS and/or NAM, a selective SIRT1 inhibitor, for
48 h prior to treatment with H2O2 (25 μM) for
1 h. NAM attenuated the decrease in SA-β-gal-positive cells
inferred by NaHS alone (Fig. 2A and
C).
H2S prevents
H2O2-mediated dysfunction in HUVECs
Since cell cycle arrest is a common hallmark of
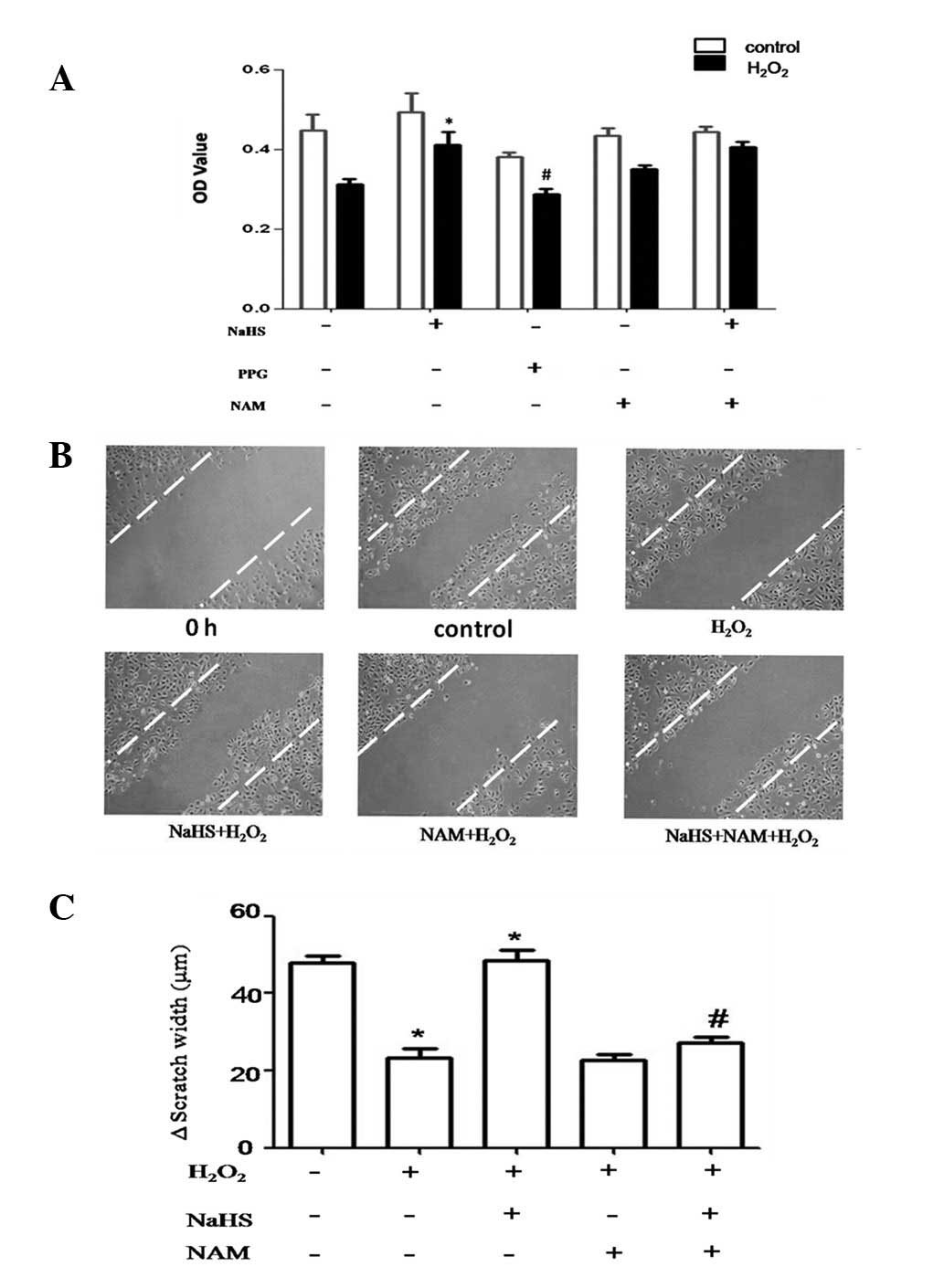
cellular senescence, we examined cell proliferation using the MTT
assay. Our study indicated that NaHS (60 μM) improved
H2O2-induced decreases in HUVEC
proliferation. This reduction in proliferation was similar, but not
significant, in cells pretreated with NAM and NaHS. Treatment with
NaHS alone, however, was effective at rescuing the
anti-proliferative effects of H2O2 (Fig. 4A). To further examine whether
H2S attenuates senescence-induced endothelial cell
dysfunction, we monitored cell migration using a scratch assay
(25). Cell migration was
significantly reduced by H2O2 and
pretreatment with NaHS eliminates this decrease. However,
pretreatment with NAM and NaHS was associated with significantly
decreased cell migration, which is similar to that observed in
cells treated with H2O2 (Fig. 4C). We conclude that H2S
prevents the reduction in cell migration associated with
senescence, an effect that is prevented by NAM. These results
indicate that H2S protects HUVECs against senescence
through SIRT1.
Discussion
A number of studies have suggested that aging is an
independent risk factor for the development of cardiovascular
diseases. Previous studies further demonstrated that cellular
senescence is involved in various pathological conditions, which
are not limited to the cardiovascular system (26). Cellular senescence is a process by
which cells irreversibly exit the cell cycle and cease to divide in
response to a variety of stresses, including those observed during
oxidative states (27). In this
study, we established an H2O2-induced
senescent model in vitro using HUVECs to investigate the
protective role of H2S in cell senescence.
Results of the present study demonstrated that a
dose of 25 μM H2O2 increased the number of
SA-β-gal-positive cells, which was eliminated following treatment
with the H2S donor, NaHS. These results demonstrated the
importance of H2S in preventing HUVEC senescence. The
results obtained during cell cycle analyses were consistent with
this observation. Coincidently, previous studies have shown that
H2S increases the lifespan in C. elegans(16). Our results suggest that
H2S may be responsible for retarding the aging process.
Furthermore, we explored the mechanism of H2S against
HUVEC senescence.
The results of our study indicated that SIRT1
expression was unchanged after HUVECs were pretreated with 60 μM
NaHS for 48 h, while SIRT1 enzyme activity was enhanced, indicating
that SIRT1 is a key sensor system for regulating endothelial cell
survival, proliferation and senescence. A recent study suggested
that SIRT1 overexpression in a mouse model led to a significant
improvement in animal health during aging (28). Another study demonstrated that the
protective effects of SIRT1 may be due to the regulation of
acetylation/deacetylation of key proteins (29). One study indicated that SIRT1
protein levels decrease rapidly with each increase in cell passage
and this leads to premature senescence (30). SIRT1 protein stability may play a
role in the progressive loss of SIRT1 associated with aging;
however, the mechanisms remain elusive. One study indicated that
post-translational modification by sumoylation affects the activity
of SIRT1 (31). Another study
demonstrated that ~10–25% of liver proteins, including actin,
tubulin and glyceraldehyde-3-phosphate dehydrogenase (GAPDH),
undergo sulfhydration under physiological conditions. Sulfhydration
appears to be a physiological post-translational modification for
proteins (32). We consider that
H2S may activate SIRT1 via sulfhydration of the SIRT1
protein. It is possible that SIRT1 is indirectly activated via
other H2S-induced physiological alterations. The
hypothesis that H2S shifts redox homeostasis, thereby
increasing available NAD+ (or the NAD+/NADH
ratio) and resulting in increased SIRT1 activity is
controversial.
HUVEC senescence and the consequent reduction of
their proliferative and migration ability may contribute to
miopragia associated with advanced age. In the present study, we
observed that HUVECs treated with 25 μM H2O2
have a reduced ability to migrate, while NaHS treatment prevented
this process. However, the mechanisms by which H2S
improves cell proliferation remain unclear. It is accepted that
increased cellular senescence is associated with decreased cell
proliferation in vivo since senescent cells are not able to
divide (33). Therefore, it may be
concluded that NaHS (60 μmol/l) improves HUVEC proliferation by
delaying cell senescence.
Exogenous H2S has notable effects on
mammalian physiology that improve survival in changing
environmental conditions (17).
Despite the molecular mechanisms involved in the modulation of
SIRT1 activity, H2S affects SIRT1 activity and
attenuates senescence, thus establishing that H2S
exhibits novel endothelial protective effects. FoxO1, is a
downstream target of SIRT1, which has been shown to modulate
G1-S and G2-M phase transition by
coordinating the expression of multiple important cell cycle
regulators (34). In future
studies, we aim to evaluate the effect of H2S on FoxO1
via SIRT1 expression and activity.
A previous study indicated that
H2S-releasing diclofenac derivatives, a novel class of
non-steroidal anti-inflammatory drugs (NSAIDs), may be of clinical
value in the treatment of osteolytic bone disease (35). In addition, preclinical studies
have indicated that the novel hydrogen sulfide-modulating agent,
S-propargyl-L-cysteine (SPRC) is a potent cardioprotective
candidate (36). Our study
provides a novel therapeutic role for H2S, which
protects against HUVEC senescence.
Acknowledgements
This study was supported by grants from the National
Natural Science Foundation of China (81170277, ZS Jiang), the
Special Research Fund for the Doctoral Program of Higher Education
of China Project (20124324110003, ZS Jiang) and the Aid Program for
Science and Technology Innovative Research Team in Higher
Educational Institutions of Hunan Province.
Abbreviations:
|
H2S
|
hydrogen sulfide
|
|
SIPS
|
stress-induced premature
senescence
|
|
CSE
|
cystathionine γ-lyase
|
References
|
1
|
Sniderman AD and Furberg CD: Age as a
modifiable risk factor for cardiovascular disease. Lancet.
371:1547–1549. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Behrendt D and Ganz P: Endothelial
function. From vascular biology to clinical applications. Am J
Cardiol. 90:40L–48L. 2002.PubMed/NCBI
|
|
3
|
Minamino T, Miyauchi H, Yoshida T, et al:
Endothelial cell senescence in human atherosclerosis: role of
telomere in endothelial dysfunction. Circulation. 105:1541–1544.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Serrano AL and Andrés V: Telomeres and
cardiovascular disease: does size matter? Circ Res. 94:575–584.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gorbunova V, Seluanov A and Pereira-Smith
OM: Expression of human telomerase (hTERT) does not prevent
stress-induced senescence in normal human fibroblasts but protects
the cells from stress-induced apoptosis and necrosis. J Biol Chem.
277:38540–38549. 2002. View Article : Google Scholar
|
|
6
|
Frippiat C, Chen QM, Zdanov S, Magalhaes
JP, Remacle J and Toussaint O: Subcytotoxic H2O2 stress triggers a
release of transforming growth factor-beta 1, which induces
biomarkers of cellular senescence of human diploid fibroblasts. J
Biol Chem. 276:2531–2537. 2001. View Article : Google Scholar
|
|
7
|
Rimondi E, di Iasio MG, Gonelli A,
Celeghini C, Secchiero P and Zauli G: Hydrogen sulfide
down-regulates the expression and release of osteoprotegerin (OPG)
by vascular endothelial cells. Invest New Drugs. 30:1731–1735.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lavu M, Bhushan S and Lefer DJ: Hydrogen
sulfide-mediated cardioprotection: mechanisms and therapeutic
potential. Clin Sci (Lond). 120:219–229. 2011.PubMed/NCBI
|
|
9
|
Predmore BL and Lefer DJ: Hydrogen
sulfide-mediated myocardial pre- and post-conditioning. Expert Rev
Clin Pharmacol. 4:83–96. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Whiteman M, Gooding KM, Whatmore JL, et
al: Adiposity is a major determinant of plasma levels of the novel
vasodilator hydrogen sulphide. Diabetologia. 53:1722–1726. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Muzaffar S, Jeremy JY, Sparatore A, Del
Soldato P, Angelini GD and Shukla N: H2S-donating
sildenafil (ACS6) inhibits superoxide formation and gp91phox
expression in arterial endothelial cells: role of protein kinases A
and G. Br J Pharmacol. 155:984–994. 2008.
|
|
12
|
Jeney V, Komódi E, Nagy E, et al:
Suppression of hemin-mediated oxidation of low-density lipoprotein
and subsequent endothelial reactions by hydrogen sulfide (H(2)S).
Free Radic Biol Med. 46:616–623. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Geng B, Chang L, Pan C, Qi Y, Zhao J, Pang
Y, Du J and Tang C: Endogenous hydrogen sulfide regulation of
myocardial injury induced by isoproterenol. Biochem Biophys Res
Commun. 318:756–763. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Whiteman M, Cheung NS, Zhu YZ, et al:
Hydrogen sulphide: a novel inhibitor of hypochlorous acid-mediated
oxidative damage in the brain? Biochem Biophys Res Commun.
326:794–798. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Feng X, Chen Y, Zhao J, Tang C, Jiang Z
and Geng B: Hydrogen sulfide from adipose tissue is a novel insulin
resistance regulator. Biochem Biophys Res Commun. 380:153–159.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Miller DL and Roth MB: Hydrogen sulfide
increases thermotolerance and lifespan in Caenorhabditis
elegans. Proc Natl Acad Sci USA. 104:20618–20622. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Luo J, Nikolaev AY, Imai S, Chen D, Su F,
Shiloh A, Guarente L and Gu W: Negative control of p53 by Sir2alpha
promotes cell survival under stress. Cell. 107:137–148. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Brunet A, Sweeney LB, Sturgill JF, et al:
Stress-dependent regulation of FOXO transcription factors by the
SIRT1 deacetylase. Science. 303:2011–2015. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ota H, Tokunaga E, Chang K, et al: Sirt1
inhibitor, Sirtinol, induces senescence-like growth arrest with
attenuated Ras-MAPK signaling in human cancer cells. Oncogene.
25:176–185. 2006.PubMed/NCBI
|
|
20
|
Dimri GP, Lee X, Basile G, Acosta M, et
al: A biomarker that identifies senescent human cells in culture
and in aging skin in vivo. Proc Natl Acad Sci USA. 92:9363–9367.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Köse GT, Korkusuz F, Ozkul A, Soysal Y,
Ozdemir T, Yildiz C and Hasirci V: Tissue engineered cartilage on
collagen and PHBV matrices. Biomaterials. 26:5187–5197.
2005.PubMed/NCBI
|
|
22
|
Oeseburg H, Iusuf D, van der Harst P, van
Gilst WH, Henning RH and Roks AJ: Bradykinin protects against
oxidative stress-induced endothelial cell senescence. Hypertension.
53:417–422. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Freedman DA and Folkman J: CDK2
translational down-regulation during endothelial senescence. Exp
Cell Res. 307:118–130. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Okatani Y, Wakatsuki A and Reiter RJ:
Protective effect of melatonin against homocysteine-induced
vasoconstriction of human umbilical artery. Biochem Biophys Res
Commun. 277:470–475. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liang CC, Park AY and Guan JL: In vitro
scratch assay: a convenient and inexpensive method for analysis of
cell migration in vitro. Nat Protoc. 2:329–333. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lakatta EG and Levy D: Arterial and
cardiac aging: major shareholders in cardiovascular disease
enterprises: Part I: aging arteries: a ‘set up’ for vascular
disease. Circulation. 107:139–146. 2003.
|
|
27
|
Ben-Porath I and Weinberg RA: The signals
and pathways activating cellular senescence. Int J Biochem Cell
Biol. 37:961–976. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Herranz D, Muñoz-Martin M, Cañamero M, et
al: Sirt1 improves healthy ageing and protects from metabolic
syndrome-associated cancer. Nat Commun. 1:32010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zu Y, Liu L, Lee MY, Xu C, et al: SIRT1
promotes proliferation and prevents senescence through targeting
LKB1 in primary porcine aortic endothelial cells. Circ Res.
106:1384–1393. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sasaki T, Maier B, Bartke A and Scrable H:
Progressive loss of SIRT1 with cell cycle withdrawal. Aging Cell.
5:413–422. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yang Y, Fu W, Chen J, Olashaw N, Zhang X,
Nicosia SV, Bhalla K and Bai W: SIRT1 sumoylation regulates its
deacetylase activity and cellular response to genotoxic stress. Nat
Cell Biol. 9:1253–1262. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mustafa AK, Gadalla MM, Sen N, et al:
H2S signals through protein S-sulfhydration. Sci Signal.
2:ra722009.
|
|
33
|
Smith HS: Hydrogen sulfide’s involvement
in modulating nociception. Pain Physician. 12:901–910. 2009.
|
|
34
|
Ho KK, Myatt SS and Lam EW: Many forks in
the path: cycling with FoxO. Oncogene. 27:2300–2311. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Frantzias J, Logan JG, Mollat P, Sparatore
A, Del Soldato P, Ralston SH and Idris AI: Hydrogen
sulphide-releasing diclofenac derivatives inhibit breast
cancer-induced osteoclastogenesis in vitro and prevent osteolysis
ex vivo. Br J Pharmacol. 165:1914–1925. 2012. View Article : Google Scholar
|
|
36
|
Zheng Y, Xu J, Ma G, Zhang J, Zhu Q, Liu
H, Zhang P, Zhu Y and Cai W: Bioavailability and pharmacokinetics
of S-propargyl-L-cysteine, a novel cardioprotective agent, after
single and multiple doses in Beagle dogs. Xenobiotica. 42:304–309.
2012. View Article : Google Scholar : PubMed/NCBI
|