Metabolic adaptations are closely associated with
alterations in cellular behavior. In the past 20 years, there has
been a growing interest in cancer metabolism, particularly on
glucose metabolism (1). Cancer cells
are able to reprogram their energy metabolism to meet the increased
biogenetic demands required for their rapid and uncontrolled growth
(2). Cells from normal tissues
mainly generate adenosine 5′-triphosphate (ATP) through the
mitochondrial oxidative phosphorylation. In these cells, glucose is
transformed to pyruvate through glycolysis, and most pyruvate
enters mitochondrial oxidative metabolism for efficient energy
generation (3). However, most cancer
cells consume glucose through glycolysis, even in the presence of
sufficient oxygen; this phenomenon is called the Warburg effect,
which leads to the production of pyruvate and lactate as final
metabolites (4). This enhanced
aerobic glycolysis allows cancer cells to better proliferate by
generating sufficient amounts of ATP and other biomolecules,
including nucleotides, amino acids and fatty acids (5).
The pentose phosphate pathway (PPP), also known as
the phosphogluconate pathway or the hexose monophosphate shunt, is
a metabolic pathway parallel to glycolysis, and represents the
first committed step of glucose metabolism (6). The PPP serves a pivotal role in
supporting cancer cell survival and growth by generating pentose
phosphate for nucleic acid synthesis and providing
nicotinamide-adenine dinucleotide phosphate (NADPH), which is
needed for fatty acid synthesis and cell survival under stress
conditions (7). Previous studies
indicate that PPP flux can be directly or indirectly modulated in
cancer cells, in order to improve cell survival and proliferation
(2,7). Therefore, the regulatory network of PPP
flux represents an important metabolic adaptation in a number of
environmental contexts in human malignancies, including cancer.
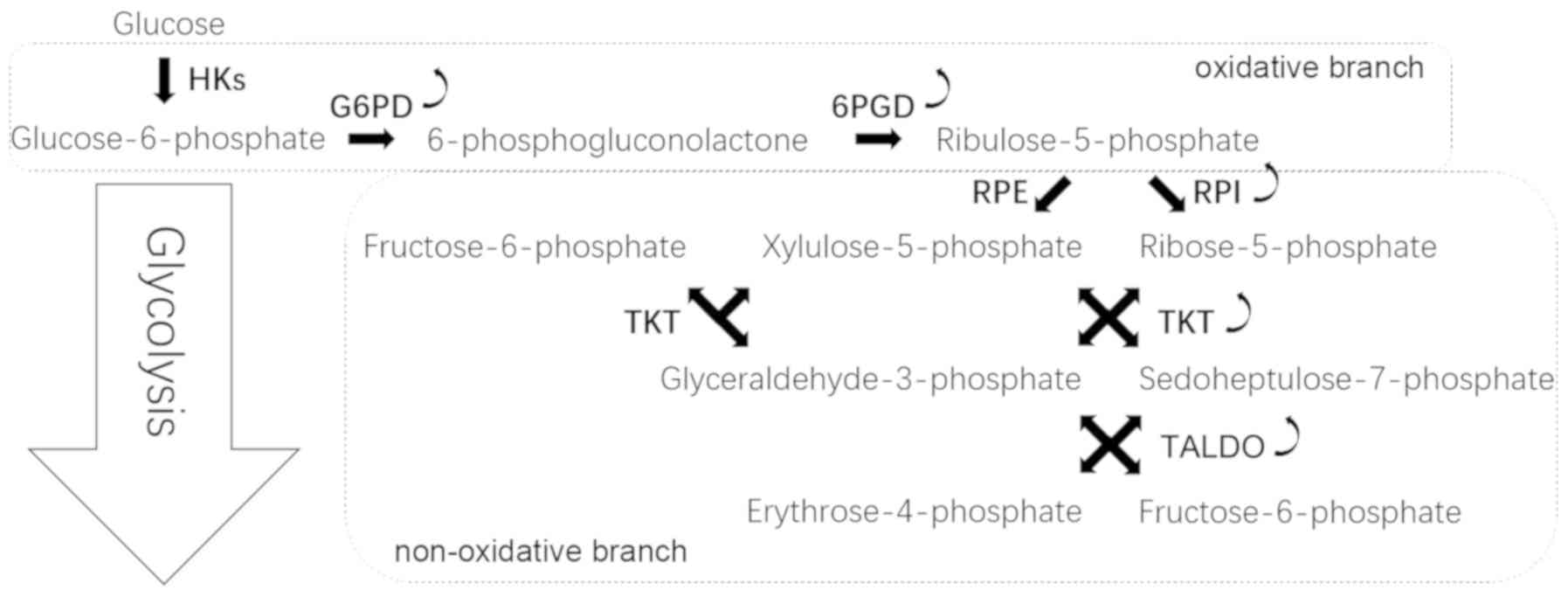
The PPP occurs in the cytosol and comprises two
irreversible oxidative reactions followed by a series of reversible
interconversions (Fig. 1). The PPP
is thus divided into two biochemical branches: An oxidative and a
non-oxidative branch. The oxidative branch converts glucose
6-phosphate (G6P) into ribulose-5-phosphate (Ru5P), CO2
and NADPH (8). NADPH is vital to
maintain the reduction-oxidation (redox) balance under stress
conditions and allows cells to proliferate rapidly (9). The non-oxidative branch yields the
glycolytic intermediates fructose 6-phosphate (F6P), glyceraldehyde
3-phosphate (G3P) and sedoheptulose sugars, resulting in the
production of sugar phosphate precursors for amino acid synthesis
and ribose-5-phosphate (R5P), which is essential for nucleic acid
synthesis (10).
The PPP is primarily regulated during the G6PD
reaction. G6PD catalyzes the irreversible oxidation of G6P into
6-phosphogluconolactone in a rate-limiting step; the first molecule
of NADPH is generated during this reaction (11). G6PD acts as a ‘gatekeeper’ of this
pathway and is therefore the rate-limiting enzyme in the PPP.
Subsequently, G6PD activity not only determines the flux
partitioning between glycolysis and PPP, but also reflects the
oxidative PPP flux (12). G6PD is
overexpressed in cancer cells, and Ju et al (13) demonstrated that the elevated
expression of G6PD is predictive of poor survival of patients with
cancer, indicating that G6PD may serve a vital role in
tumorigenesis. There are two cellular isomers of G6PD, a dimer and
a tetramer; the dimer stability has been demonstrated to have an
important role in vivo (14).
High pH and ionic strength are beneficial for the dimer synthesis,
whereas low pH generates a shift toward the tetramer synthesis
(15).
The tumor suppressor p53 binds to G6PD and inhibits
the formation of the active dimer and suppresses NADPH production,
glucose consumption and biosynthesis, which results in inhibition
of the PPP (16). Polo-like kinase 1
(Plk1) is a key regulator of cell mitosis and enhances PPP flux and
macromolecule biosynthesis through the direct phosphorylation of
G6PD to promote the formation of G6PD active dimer. This is an
essential feature of Plk1 as a promoter of cancer cell cycle
progression and growth (17). In
addition, glycosylation activates G6PD activity, and modification
of G6PD with an O-linked β-N-acetylglucosamine sugar increases the
glucose flux to the PPP (18).
Mammalian target of rapamycin complex 1 upregulates the
transcriptional and the post-transcriptional expression of G6PD to
activate PPP (19). p21-activated
kinase 4 increases G6PD activity by enhancing Mdm2-mediated p53
ubiquitination and degradation (20). Furthermore, suppression of G6PD
lowers glutathione levels, decreases NADPH production, reduces the
capacity to scavenge reactive oxygen species (ROS) and enhances the
oxaliplatin-induced apoptosis through ROS-mediated damage in
vitro (13). These results
indicate that G6PD may be a potential prognostic biomarker and
represent a promising target in cancer therapy.
The 6-phosphogluconolactone hydrolase irreversibly
hydrolyzes 6-phosphogluconolactone into 6-phosphogluconate (6PG).
6PG is then oxidatively decarboxylated by 6PGD, leading to the
synthesis of Ru5P, CO2 and a second molecule of NADPH.
Upregulation of 6PGD activity has been identified in various types
of cancer, including breast, acute myeloid leukemia (AML), ovarian
and lung cancers (21–23).
The enzyme 6PGD is commonly activated in human
cancer cells after lysine acetylation, which promotes
NADP+ binding to 6PGD and the formation of active dimers
of 6PGD (24). In this pathway,
activated 6PGD enhances the oxidative phase of PPP, and nucleotide
or RNA biosynthesis. This reaction serves a role in maintaining
intracellular Ru5P at a physiological level that is sufficient to
fulfill the metabolic requirements of rapidly growing cancer cells
(25). In addition,
3-phosphoglycerate (3-PG) directly binds to the active site of 6PGD
and competes with its substrate, 6PG, to inhibit 6PGD. Furthermore,
the glycolytic enzyme phosphoglycerate mutase 1 (PGAM1) controls
intracellular levels of 3-PG (26).
A recent study reported that attenuation of PGAM1 results in
abnormal accumulation of 3-PG, which inhibits 6PGD and subsequently
suppresses the oxidative PPP and anabolic biosynthesis. Malic
enzyme forms a physiological hetero-oligomer with 6PGD, which
increases 6PGD activity (27).
The enzyme RPI converts Ru5P into R5P, and the
enzyme RPE converts Ru5P into xylulose-5-phosphate (Xu5P). It has
been demonstrated that ribose-5-phosphate isomerase A (RPIA)
regulates cancer growth and tumorigenesis (28). In addition, RPIA is significantly
overexpressed in colorectal cancer and hepatocellular carcinoma
(HCC) (29,30). RPIA also activates β-catenin by
entering the nucleus to form a complex with adenomatous polyposis
coli and β-catenin, thus modulating cell proliferation and
oncogenicity (29).
TKT and TALDO are two enzymes that convert R5P and
Xu5P, and the gluconeogenetic intermediates G3P and F6P. TKT and
TALDO are responsible for complex interconversion reactions within
the non-oxidative PPP (10). TKT
converts excess R5P into G3P and F6P through a number of reactions,
G3P is metabolized alongside further steps of glycolysis, and F6P
is converted into G6P that re-enters the oxidative PPP to generate
additional NADPH (31). Elevated TKT
expression levels were reported in lung cancer cells, breast cancer
cells and prostate cancer cells (21,22).
TKT expression is closely regulated by the nuclear
factor, erythroid 2-like 2 (NRF2)/Kelch-like ECH-associated protein
1/BTB and CNC homolog 1 oxidative stress sensor pathway in various
types of cancer (32). For example,
exposure to ultraviolet A increases cancer proliferation by
upregulating intracellular concentrations of TKT in melanoma
(33). In addition, fructose
stimulates TKT activity and is preferentially used over glucose to
generate nucleic acids via the non-oxidative PPP (34). Higher vertebrates obtain
transketolase-like 1 (TKTL1) by genome duplication and exon
skipping (35,36). TKTL1 upregulation is a general
phenomenon in epithelial malignancies, ocular adnexal tumors,
malignant pleural effusion and other types of cancer (37–39).
TKTL1 is therefore considered a novel tumor marker and a potential
good target in cancer treatment (40).
TALDO catalyzes the reversible transfer of a
three-carbon unit between various sugar phosphates (from ketose to
aldose sugar phosphates) (10). A
previous study has revealed that TALDO is significantly
overexpressed in gastric adenocarcinoma (41). Furthermore, its expression is
associated with metastatic behavior in HCC (42). In addition, a combination of arginine
and ascorbic acid decreases intracellular NADPH levels by reducing
TALDO activity in the PPP (43).
Numerous regulatory pathways for tumor cells exist
within the PPP, and most reactions in glycolysis are crucial to
maintain tumor cell function. Since PPP and glycolysis are
metabolically linked for sharing the common intermediate G6P, the
increased glycolysis during reperfusion concomitantly led to
decreased PPP rate (44). The
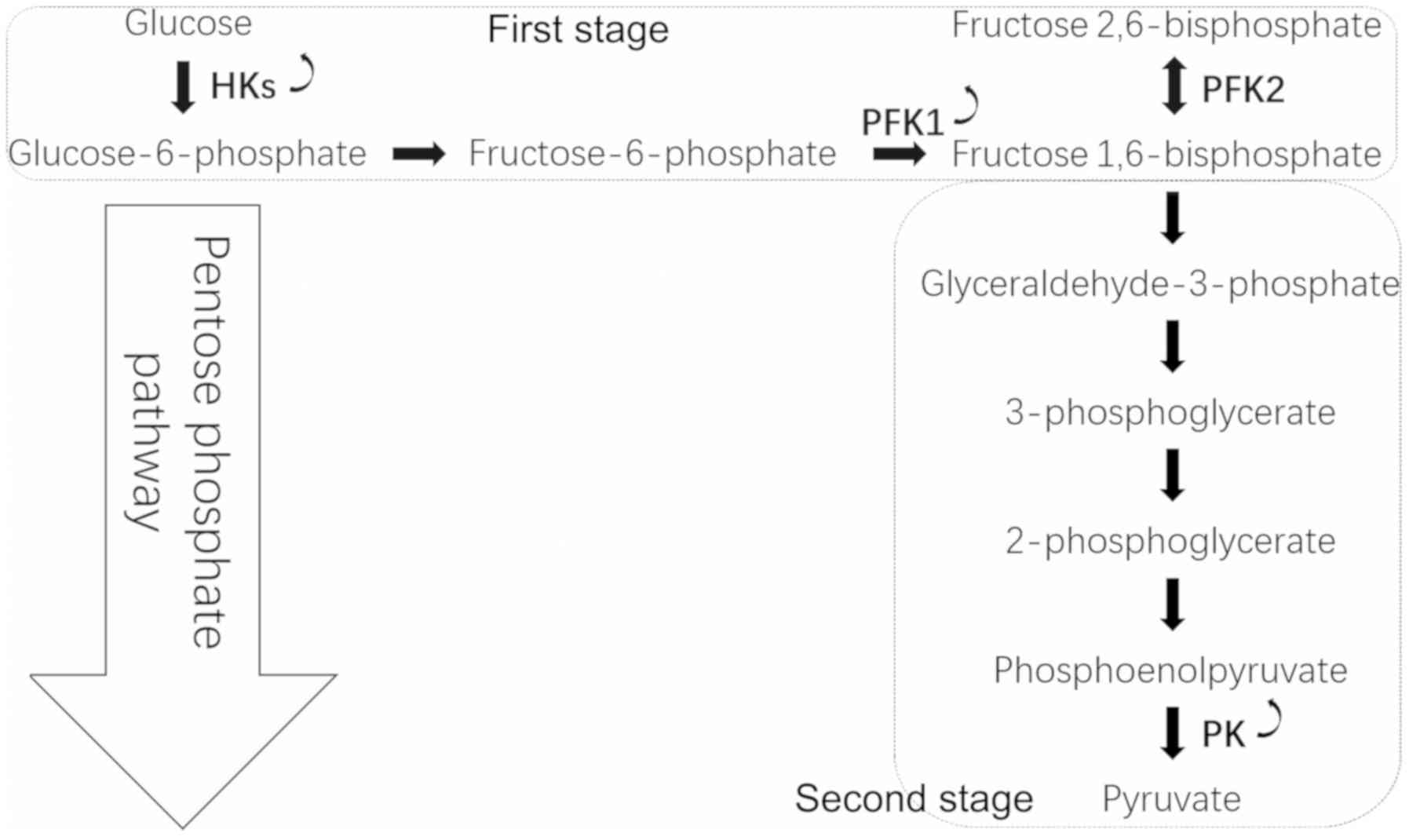
conversion of glucose to pyruvate occurs in two stages (Fig. 2). In the first stage, phosphorylated
forms of pyruvate intermediates are synthesized, leading to ATP
synthesis. Hexokinase (HK) phosphorylates glucose into G6P, and
phosphofructokinase-1 (PFK1) catalyzes the conversion of F6P to
fructose 1,6-bisphosphate. In the subsequent stage, ATP is
generated by substrate-level phosphorylation and metabolism of
glucose. The final step of glycolysis is catalyzed by the pyruvate
kinase (PK) enzyme that leads to the synthesis of pyruvate and ATP.
In cancer cells, the glycolytic reaction generates a ‘bottleneck’
effect by increasing the upstream part of the glycolytic flux up to
PK and decreasing the glycolytic flux from PK downward (45).
HK catalyzes glucose phosphorylation, which is one
of the regulatory reactions of glycolysis. To maintain the Warburg
effect, cancer cells upregulate HK. Four isoforms of HK exist
(HK1-HK4). HK2, which may be in a soluble form in the cytoplasm or
bound to the mitochondrial outer membrane, has a glucose affinity
100-fold higher than HK1, HK3 and HK4 (46). In addition, the expression of HK1 may
be sufficient for normal cell metabolism. However, the accelerated
anabolic metabolism in cancer cells demands a robust HK activity.
Therefore, the induction of HK2 expression is required. Overall,
HK2 is elevated in cancer cells, promotes glycolysis and inhibits
mitochondrial-mediated apoptosis (47).
The induction of HK2 expression by oncogenic Ras is
crucial for accelerated ribonucleotide synthesis (48). Bcl-2-associated athanogene (BAG)-3, a
member of the BAG cochaperone family that comprises six BAGs
(BAG1-BAG6), increases HK2 expression by interacting with HK2 mRNA
(49). Hypoxia-inducible factor
(HIF)-1α induces the expression of the glycolytic enzyme HK2. The
sustained expression of the oncogene forms of the human
papillomavirus E6 and E7 is vital to maintain HK2 expression levels
by upregulating the pro-oncogene MYC and downregulating microRNA
(miR)-143-3p (50). In AML, an
internal tandem duplication mutation in the Fms-like tyrosine
kinase 3 gene upregulates the level of mitochondrial HK2, causing a
significant increase in aerobic glycolysis; therefore, leukemic
cells become highly dependent on glycolysis, which increases their
sensitivity to the pharmacological inhibition of glycolytic
activity (51). In addition, the
histone-lysine N-methyltransferase NSD2 is recruited to and
methylates HK2 promoters (52).
NSD2-driven tamoxifen-resistant cancers exhibit an enhanced PPP
activity, elevated NADPH production and reduced ROS levels. For
example, treatment of ovarian cancer xenografted mice with the HK2
inhibitor 3-bromopyruvate attenuates tumor growth and confers a
survival advantage (53).
PFK1 irreversibly phosphorylates F6P into
fructose-1,6-bisphosphate. This reaction is a crucial and a
rate-limiting step in glycolysis. It has been demonstrated that
PFK1 activity is increased in cancer cell lines, and expression of
PFK1 is upregulated in breast and liver cancers (54,55). In
addition, PFK1 is regulated by ATP and F6P substrates (56).
In response to hypoxia, O-GlcNAcylation suppresses
PFK1 activity and redirects glucose towards the PPP, which provides
an advantage for cancer cell growth (57). A Krüppel-associated box-type
zinc-finger protein named p53 inhibitor of TIGAR activation (PITA)
is a selective regulator of p53, and PITA transgenic mice exhibit
increased PFK1 activity and elevated glycolytic rate (58). The PFK1 platelet isoform (PFKP), the
predominant PFK1 isoform, is overexpressed in human glioblastoma
cells and promotes aerobic glycolysis and brain cancer cell
proliferation (59). In addition,
the loss of phosphatase and tensin homolog (PTEN) and activation of
epidermal growth factor receptor (EGFR)-dependent phosphoinositide
3-kinase cause AKT activation, which in turn increases PFKP
stability (59). In leukemic cells,
the cyclin D3-cyclin dependent kinase 6 (CDK6) phosphorylates PFKP
and suppresses its activity (60),
thus shifting the glucose-derived carbon into the PPP. Through this
mechanism, cyclin D3-CDK6 enhances NADPH production to neutralize
ROS. Snail1, which is a key transcriptional repressor of
epithelial-mesenchymal transition (EMT), represses PFKP, leading to
the glucose flux switch to PPP and the generation of NADPH
(61). In addition, heme
oxygenase-1/carbon monoxide reduces methylation of
6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase 3 (PFKFB3) in
cancer cells, thus redirecting glucose from the glycolysis pathway
to the PPP, ensuring cancer cell resistance against oxidative
stress (62). The dynamic regulation
of PFKP enhances the survival of cancer cells undergoing metabolic
stress and therefore increases their ability to metastasize in
vivo.
PK converts phosphoenolpyruvate into pyruvate during
the third irreversible reaction of glycolysis; thus, PK serves an
important role in the control of metabolism in cancer cells. The
ratio between the active and inactive forms of PK in cancer cells
determines whether glucose is used for OXPHOS or for PPP to support
cell growth (63). Low pyruvate
kinase activity increases glucose influx into PPP for biosynthesis
while high pyruvate kinase activity increases OXPHOS and decreases
glucose influx into PPP (64). PK
possesses two isoforms generated by alternative splicing of PK
named M1 and M2, of which expressions are location and time
dependent: The pyruvate kinase M2 isoform (PKM2) is preferentially
expressed in cancer cells, where complex regulation of its activity
is essential for the control of cellular metabolism (65).
Upon glucose starvation, cellular levels of
succinylaminoimidazole-carboxamide riboside, an intermediate of the
de novo purine nucleotide synthesis pathway, are increased.
This leads to the stimulation of PKM2 activity in cancer cells,
which alters cellular energy level, glucose uptake and lactate
generation (66). Following EGFR
activation, PKM2 binds and phosphorylates histone H3 at T11.
PKM2-dependent histone H3 modification contributes to EGF-induced
cyclin D1 and c-MYC expression, tumor cell proliferation, cell
cycle progression and brain tumorigenesis (67). In human lung cancer cells, the marked
increase in intracellular ROS leads to the inhibition of the
glycolytic enzyme PKM2 by oxidation of Cys358, which requires the
transfer of glucose flux into the PPP, stimulating redox potential
and ROS detoxification (68). In
addition, PKM2 gene transcription is activated by HIF-1 by direct
interaction with the HIF-1α subunit (69). Serine binds to and activates human
PKM2, and the PKM2 activity in cells after depletion of serine is
reduced. This reduction in PKM2 activity switches the cells into a
fuel-saving mode in which more pyruvate is transferred to
mitochondria to support cell proliferation (70).
Elevated expression of G6PD is associated with HCC
metastases and poor prognosis of patients with HCC, and G6PD
knockdown inhibits the proliferation, migration and invasion of HCC
cell lines in vitro (74). In
addition, G6PD promotes HCC cell migration and invasion by
activating the signal transduction and activator of transcription 3
(STAT3) pathway to induce EMT (74).
The transcription factor NRF2 is required for G6PD induction, and
miR-1 is involved in its activation (75). BAG directly interacts with G6PD to
suppress the PPP flux, DNA synthesis and HCC cell growth (76). Furthermore, PTEN binds to G6PD to
prevent formation of the active G6PD dimer, which subsequently
inhibits the PPP. However, the AKT coactivator T cel1
leukemia/lymphoma protein IA promotes G6PD activity and increases
G6PD pre-mRNA splicing and protein expression (77). Inhibitor of differentiation and DNA
binding-1 (ID1), regulates c-MYC through Wnt/β-catenin pathway
activation to promote G6PD promoter transcription and activate the
PPP (78), which confers to HCC
cells an oxaliplatin chemoresistance (79). In addition, ID1 activates the PPP to
increase NADPH production and reduce intracellular ROS levels, thus
promoting chemotherapy resistance in HCC.
Numerous key enzymes from the glycolysis pathway are
involved in the carcinogenesis of HCC. The major distinction
between HCC cells and normal hepatocytes is the difference in
enzymes that catalyze the first step of glucose metabolism. In
normal hepatocytes, this step is catalyzed by glucokinase, whereas
this enzyme is lacking in HCC cells and is replaced by HK2
(80). The long non-coding RNA
taurine upregulated gene 1 (TUG1) controls cell migration and
glycolysis by regulating the p21/miR-455-3p axis, which affects HK2
stability during translation but not transcription (81). In addition, miR-125a overexpression
significantly decreases HK2 protein level in HCC cells, which
indicates that miR-125a directly targets HK2 (82). In addition, overexpression of STAT3
upregulates HK2 mRNA and HK2 protein expression (83). Furthermore, hypomethylation in the
HK2 promoter CpG island (CGI) N-shore region increases HK2
expression, and hypermethylation in the HK2-CGI suppresses HK2
expression by inhibiting the interaction between a hypoxia response
element and HIF-1α (84).
G6PD is closely associated with molecular subtypes
of breast cancer, and its upregulation is a negative prognostic
factor in breast cancer (85,86). It
has been demonstrated that G6PD silencing increases the glycolytic
flux, reduces lipid synthesis and increases glutamine uptake in
breast cancer cells, whereas TKT silencing reduces glycolysis flux
(31). Overexpression of NSD2 in
breast cancer induces cancer resistance to tamoxifen by
upregulating G6PD and HK2 expression, which enhances PPP flux
(52). In addition, G6PD expression
and activity are continuously unregulated in breast cancer cells,
and it has been reported that G6PD inhibition leads to an increase
in 5′-AMP-activated protein kinase (AMPK) signaling, a decrease in
lipid biosynthesis and the inhibition of breast cancer cell growth
and survival (21). Furthermore, TKT
expression is associated with tumor size in the 4T1/BALB/c
syngeneic model, and high TKT levels are associated with poor
survival (87).
The YAP/TEAD/p65 axis upregulates HK2 transcription,
which promotes breast cancer cell migration. This axis may
therefore represent a potential therapeutic target for treatment of
metastatic breast cancer (88). It
has been reported that inhibition of hexokinase using
2-deoxyglucose induces chloroquine-resistance in breast cancer
(89). In addition, metformin
stimulates the glycolytic flux caused by starvation by interfering
with HK2 activity (90).
Furthermore, the AMPK-dependent phosphorylation of PFKFB3
substitutes oxidative respiration by glycolysis, which causes
inhibition of cell death and of antitumor efficiency of the
microtubule toxin in breast cancer cells (91). Sonic hedgehog phosphorylates PFKFB3
to promote glycolysis and proliferation of breast cancer cells,
which is mediated by smoothened and p38/MK2 (92). In addition, the
6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase 4 (PFKFB4) in
breast cancer cells can phosphorylate the oncogenic steroid
receptor coactivator-3, which rapidly increases its transcriptional
activity and promotes the glucose flux switch towards purine
synthesis (93). Furthermore, PKM2
is phosphorylated at tyrosine 105 and forms oncogenic dimers in
breast cancer cells, whereas PKM2 is largely unphosphorylated and
forms non-tumorigenic tetramers in non-transformed MCF10A cells
(94). Moreover, the intragenic DNA
methylation-mediated binding of the protein brother of regulator of
imprinted sites on the replacement exon of PK is associated with
cancer-specific splicing that promotes the Warburg effect and thus
breast cancer progression (95).
The metabolic processes in cancer cells differ from
those in normal cells. In malignancies, cancer cell proliferation
is stimulated. Elevated PPP activity in cancer cells may
distinguish cancer cells from normal cells, and the enzymes
involved in PPP may therefore represent novel targets for diagnosis
and treatment of various types of cancer. The present review
demonstrated that cancer cells have acquired numerous mechanisms
that circumvent PPP and glycolysis regulation. However, further
investigation remains essential to discover additional mechanisms
and identify strategies for treating hyperactive PPP signaling in
human cancers.
Not applicable.
The present study was supported by The National
Natural Sciences Foundation of China (grant no. 81672685).
Not applicable.
LJ drafted the manuscript. YZ revised the
manuscript. All authors have read and approved the final version of
this manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interest.
|
1
|
Vazquez A, Kamphorst JJ, Markert EK, Schug
ZT, Tardito S and Gottlieb E: Cancer metabolism at a glance. J Cell
Sci. 129:3367–3373. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Weber GF: Metabolism in cancer metastasis.
Int J Cancer. 138:2061–2066. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lu J, Tan M and Cai Q: The Warburg effect
in tumor progression: Mitochondrial oxidative metabolism as an
anti-metastasis mechanism. Cancer Lett. 356:156–164. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Vander Heiden MG, Cantley LC and Thompson
CB: Understanding the Warburg effect: The metabolic requirements of
cell proliferation. Science. 324:1029–1033. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Martinez-Outschoorn UE, Peiris-Pagés M,
Pestell RG, Sotgia F and Lisanti MP: Cancer metabolism: A
therapeutic perspective. Nat Rev Clin Oncol. 14:11–31. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ramos-Martinez JI: The regulation of the
pentose phosphate pathway: Remember Krebs. Arch Biochem Biophys.
614:50–52. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Patra KC and Hay N: The pentose phosphate
pathway and cancer. Trends Biochem Sci. 39:347–354. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kruger NJ and von Schaewen A: The
oxidative pentose phosphate pathway: Structure and organisation.
Curr Opin Plant Biol. 6:236–246. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pavlova NN and Thompson CB: The emerging
hallmarks of cancer metabolism. Cell Metab. 23:27–47. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Stincone A, Prigione A, Cramer T, Wamelink
MM, Campbell K, Cheung E, Olin-Sandoval V, Grüning NM, Krüger A,
Tauqeer Alam M, et al: The return of metabolism: Biochemistry and
physiology of the pentose phosphate pathway. Biol Rev Camb Philos
Soc. 90:927–963. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang C, Zhang Z, Zhu Y and Qin S:
Glucose-6-phosphate dehydrogenase: A biomarker and potential
therapeutic target for cancer. Anticancer Agents Med Chem.
14:280–289. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kathagen-Buhmann A, Schulte A, Weller J,
Holz M, Herold-Mende C, Glass R and Lamszus K: Glycolysis and the
pentose phosphate pathway are differentially associated with the
dichotomous regulation of glioblastoma cell migration versus
proliferation. Neuro Oncol. 18:1219–1229. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ju HQ, Lu YX, Wu QN, Liu J, Zeng ZL, Mo
HY, Chen Y, Tian T, Wang Y, Kang TB, et al: Disrupting
G6PD-mediated Redox homeostasis enhances chemosensitivity in
colorectal cancer. Oncogene. 36:6282–6292. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Au SW, Gover S, Lam VM and Adams MJ: Human
glucose-6-phosphate dehydrogenase: The crystal structure reveals a
structural NADP(+) molecule and provides insights into enzyme
deficiency. Structure. 8:293–303. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cohen P and Rosemeyer MA: Subunit
interactions of glucose-6-phosphate dehydrogenase from human
erythrocytes. Eur J Biochem. 8:8–15. 1969. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jiang P, Du W, Wang X, Mancuso A, Gao X,
Wu M and Yang X: p53 regulates biosynthesis through direct
inactivation of glucose-6-phosphate dehydrogenase. Nat Cell Biol.
13:310–316. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ma X, Wang L, Huang, Li Y, Yang D, Li T,
Li F, Sun L, Wei H, He K, et al: Polo-like kinase 1 coordinates
biosynthesis during cell cycle progression by directly activating
pentose phosphate pathway. Nat Commun. 8:15062017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rao X, Duan X, Mao W, Li X, Li Z, Li Q,
Zheng Z, Xu H, Chen M, Wang PG, et al: O-GlcNAcylation of G6PD
promotes the pentose phosphate pathway and tumor growth. Nat
Commun. 6:84682015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Thiepold AL, Lorenz NI, Foltyn M, Engel
AL, Divé I, Urban H, Heller S, Bruns I, Hofmann U, Dröse S, et al:
Mammalian target of rapamycin complex 1 activation sensitizes human
glioma cells to hypoxia-induced cell death. Brain. 140:2623–2638.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang X, Zhang X, Li Y, Shao Y, Xiao J,
Zhu G and Li F: PAK4 regulates G6PD activity by p53 degradation
involving colon cancer cell growth. Cell Death Dis. 8:e28202017.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yang X, Peng X and Huang J: Inhibiting
6-phosphogluconate dehydrogenase selectively targets breast cancer
through AMPK activation. Clin Transl Oncol. 20:1145–1152. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bhanot H, Weisberg EL, Reddy MM, Nonami A,
Neuberg D, Stone RM, Podar K, Salgia R, Griffin JD and Sattler M:
Acute myeloid leukemia cells require 6-phosphogluconate
dehydrogenase for cell growth and NADPH-dependent metabolic
reprogramming. Oncotarget. 8:67639–67650. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zheng W, Feng Q, Liu J, Guo Y, Gao L, Li
R, Xu M, Yan G, Yin Z, Zhang S, et al: Inhibition of
6-phosphogluconate dehydrogenase reverses cisplatin resistance in
ovarian and lung cancer. Front Pharmacol. 8:4212017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shan C, Elf S, Ji Q, Kang HB, Zhou L,
Hitosugi T, Jin L, Lin R, Zhang L, Seo JH, et al: Lysine
acetylation activates 6-phosphogluconate dehydrogenase to promote
tumor growth. Mol Cell. 55:552–565. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lin R, Elf S, Shan C, Kang HB, Ji Q, Zhou
L, Hitosugi T, Zhang L, Zhang S, Seo JH, et al: 6-Phosphogluconate
dehydrogenase links oxidative PPP, lipogenesis and tumour growth by
inhibiting LKB1-AMPK signalling. Nat Cell Biol. 17:1484–1496. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hitosugi T, Zhou L, Elf S, Fan J, Kang HB,
Seo JH, Shan C, Dai Q, Zhang L, Xie J, et al: Phosphoglycerate
mutase 1 coordinates glycolysis and biosynthesis to promote tumor
growth. Cancer Cell. 22:585–600. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yao P, Sun H, Xu C, Chen T, Zou B, Jiang P
and Du W: Evidence for a direct cross-talk between malic enzyme and
the pentose phosphate pathway via structural interactions. J Biol
Chem. 292:17113–17120. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Qiu Z, Guo W, Wang Q, Chen Z, Huang S,
Zhao F, Yao M, Zhao Y and He X: MicroRNA-124 reduces the pentose
phosphate pathway and proliferation by targeting PRPS1 and RPIA
mRNAs in human colorectal cancer cells. Gastroenterology.
149:1587–1598.e11. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chou YT, Jiang JK, Yang MH, Lu JW, Lin HK,
Wang HD and Yuh CH: Identification of a noncanonical function for
ribose-5-phosphate isomerase A promotes colorectal cancer formation
by stabilizing and activating β-catenin via a novel C-terminal
domain. PLoS Biol. 16:e20037142018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ciou SC, Chou YT, Liu YL, Nieh YC, Lu JW,
Huang SF, Chou YT, Cheng LH, Lo JF, Chen MJ, et al:
Ribose-5-phosphate isomerase A regulates hepatocarcinogenesis via
PP2A and ERK signaling. Int J Cancer. 137:104–115. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Benito A, Polat IH, Noé V, Ciudad CJ,
Marin S and Cascante M: Glucose-6-phosphate dehydrogenase and
transketolase modulate breast cancer cell metabolic reprogramming
and correlate with poor patient outcome. Oncotarget.
8:106693–106706. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xu IM, Lai RK, Lin SH, Tse AP, Chiu DK,
Koh HY, Law CT, Wong CM, Cai Z, Wong CC and Ng IO: Transketolase
counteracts oxidative stress to drive cancer development. Proc Natl
Acad Sci USA. 113:E725–E734. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kamenisch Y, Baban TSA, Schuller W, von
Thaler AK, Sinnberg T, Metzler G, Bauer J, Schittek B, Garbe C,
Rocken M and Berneburg M: UVA-irradiation induces melanoma invasion
via the enhanced Warburg effect. J Invest Dermatol. 136:1866–1875.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu H, Huang D, McArthur D, Boros L,
Nissen N and Heaney A: Fructose induces transketolase flux to
promote pancreatic cancer growth. Cancer Res. 70:6368–6376. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Coy JF, Dressler D, Wilde J and Schubert
P: Mutations in the transketolase-like gene TKTL1: Clinical
implications for neurodegenerative diseases, diabetes and cancer.
Clin Lab. 51:257–273. 2005.PubMed/NCBI
|
|
36
|
Coy JF, Dübel S, Kioschis P, Thomas K,
Micklem G, Delius H and Poustka A: Molecular cloning of
tissue-specific transcripts of a transketolase-related gene:
Implications for the evolution of new vertebrate genes. Genomics.
32:309–316. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Langbein S, Zerilli M, Zur Hausen A,
Staiger W, Rensch-Boschert K, Lukan N, Popa J, Ternullo MP,
Steidler A, Weiss C, et al: Expression of transketolase TKTL1
predicts colon and urothelial cancer patient survival: Warburg
effect reinterpreted. Br J Cancer. 94:578–585. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lange CA, Tisch-Rottensteiner J, Böhringer
D, Martin G, Schwartzkopff J and Auw-Haedrich C: Enhanced TKTL1
expression in malignant tumors of the ocular adnexa predicts
clinical outcome. Ophthalmology. 119:1924–1929. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lin CC, Chen LC, Tseng VS, Yan JJ, Lai WW,
Su WP, Lin CH, Huang CY and Su WC: Malignant pleural effusion cells
show aberrant glucose metabolism gene expression. Eur Respir J.
37:1453–1465. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Xu X, Zur Hausen A, Coy JF and Löchelt M:
Transketolase-like protein 1 (TKTL1) is required for rapid cell
growth and full viability of human tumor cells. Int J Cancer.
124:1330–1337. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kočevar N, Odreman F, Vindigni A, Grazio
SF and Komel R: Proteomic analysis of gastric cancer and immunoblot
validation of potential biomarkers. World J Gastroenterol.
18:1216–1228. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wang C, Guo K, Gao D, Kang X, Jiang K, Li
Y, Sun L, Zhang S, Sun C, Liu X, et al: Identification of
transaldolase as a novel serum biomarker for hepatocellular
carcinoma metastasis using xenografted mouse model and clinic
samples. Cancer Lett. 313:154–166. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Hsieh BS, Huang LW, Su SJ, Cheng HL, Hu
YC, Hung TC and Chang KL: Combined arginine and ascorbic acid
treatment induces apoptosis in the hepatoma cell line HA22T/VGH and
changes in redox status involving the pentose phosphate pathway and
reactive oxygen and nitrogen species. J Nutr Biochem. 22:234–241.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Li Z, Zhang B, Yao W, Zhang C, Wan L and
Zhang Y: APC-Cdh1 regulates neuronal apoptosis through modulating
glycolysis and pentose-phosphate pathway after oxygen-glucose
deprivation and reperfusion. Cell Mol Neurobiol. 39:123–135. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Senyilmaz D and Teleman A: Chicken or the
egg: Warburg effect and mitochondrial dysfunction. F1000Prime Rep.
7:412015. View
Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wilson JE: Isozymes of mammalian
hexokinase: Structure, subcellular localization and metabolic
function. J Exp Biol. 206:2049–2057. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Gu J, Singh A, Xue K, Mavis C, Barth M,
Yanamadala V, Lenz P, Grau M, Lenz G, Czuczman MS and
Hernandez-Ilizaliturri FJ: Up-regulation of hexokinase II
contributes to rituximab-chemotherapy resistance and is a
clinically relevant target for therapeutic development. Oncotarget.
9:4020–4033. 2017.PubMed/NCBI
|
|
48
|
Patra KC, Wang Q, Bhaskar PT, Miller L,
Wang Z, Wheaton W, Chandel N, Laakso M, Muller WJ, Allen EL, et al:
Hexokinase 2 is required for tumor initiation and maintenance and
its systemic deletion is therapeutic in mouse models of cancer.
Cancer Cell. 24:213–228. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
An MX, Li S, Yao HB, Li C, Wang JM, Sun J,
Li XY, Meng XN and Wang HQ: BAG3 directly stabilizes Hexokinase 2
mRNA and promotes aerobic glycolysis in pancreatic cancer cells. J
Cell Biol. 216:4091–4105. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Hoppe-Seyler K, Honegger A, Bossler F,
Sponagel J, Bulkescher J, Lohrey C and Hoppe-Seyler F: Viral E6/E7
oncogene and cellular hexokinase 2 expression in HPV-positive
cancer cell lines. Oncotarget. 8:106342–106351. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Ju HQ, Zhan G, Huang A, Sun Y, Wen S, Yang
J, Lu WH, Xu RH, Li J, Li Y, et al: ITD mutation in FLT3 tyrosine
kinase promotes Warburg effect and renders therapeutic sensitivity
to glycolytic inhibition. Leukemia. 31:2143–2150. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Wang J, Duan Z, Nugent Z, Zou JX, Borowsky
AD, Zhang Y, Tepper CG, Li JJ, Fiehn O, Xu J, et al: Reprogramming
metabolism by histone methyltransferase NSD2 drives endocrine
resistance via coordinated activation of pentose phosphate pathway
enzymes. Cancer Lett. 378:69–79. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Ha JH, Radhakrishnan R, Jayaraman M, Yan
M, Ward JD, Fung KM, Moxley K, Sood AK, Isidoro C, Mukherjee P, et
al: LPA induces metabolic reprogramming in ovarian cancer via a
pseudohypoxic response. Cancer Res. 78:1923–1934. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Moon JS, Kim HE, Koh E, Park SH, Jin WJ,
Park BW, Park SW and Kim KS: Krüppel-like factor 4 (KLF4) activates
the transcription of the gene for the platelet isoform of
phosphofructokinase (PFKP) in breast cancer. J Biol Chem.
286:23808–23816. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Park YY, Kim SB, Han HD, Sohn BH, Kim JH,
Liang J, Lu Y, Rodriguez-Aguayo C, Lopez-Berestein G, Mills GB, et
al: Tat-activating regulatory DNA-binding protein regulates
glycolysis in hepatocellular carcinoma by regulating the platelet
isoform of phosphofructokinase through microRNA 520. Hepatology.
58:182–191. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Cabrera R, Baez M, Pereira HM, Caniuguir
A, Garratt RC and Babul J: The crystal complex of
phosphofructokinase-2 of Escherichia coli with
fructose-6-phosphate: Kinetic and structural analysis of the
allosteric ATP inhibition. J Biol Chem. 286:5774–5783. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Yi W, Clark PM, Mason DE, Keenan MC, Hill
C, Goddard WA III, Peters EC, Driggers EM and Hsieh-Wilson LC:
Phosphofructokinase 1 glycosylation regulates cell growth and
metabolism. Science. 337:975–980. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Wang S, Peng Z, Wang S, Yang L, Chen Y,
Kong X, Song S, Pei P, Tian C, Yan H, et al: KRAB-type zinc-finger
proteins PITA and PISA specifically regulate p53-dependent
glycolysis and mitochondrial respiration. Cell Res. 28:572–592.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Lee JH, Liu R, Li J, Zhang C, Wang Y, Cai
Q, Qian X, Xia Y, Zheng Y, Piao Y, et al: Stabilization of
phosphofructokinase 1 platelet isoform by AKT promotes
tumorigenesis. Nat Commun. 8:9492017. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Wang H, Nicolay BN, Chick JM, Gao X, Geng
Y, Ren H, Gao H, Yang G, Williams JA, Suski JM, et al: The
metabolic function of cyclin D3-CDK6 kinase in cancer cell
survival. Nature. 546:426–430. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Kim NH, Cha YH, Lee J, Lee SH, Yang JH,
Yun JS, Cho ES, Zhang X, Nam M, Kim N, et al: Snail reprograms
glucose metabolism by repressing phosphofructokinase PFKP allowing
cancer cell survival under metabolic stress. Nat Commun.
8:143742017. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Yamamoto T, Takano N, Ishiwata K, Ohmura
M, Nagahata Y, Matsuura T, Kamata A, Sakamoto K, Nakanishi T, Kubo
A, et al: Reduced methylation of PFKFB3 in cancer cells shunts
glucose towards the pentose phosphate pathway. Nat Commun.
5:34802014. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Gui DY, Lewis CA and Vander Heiden MG:
Allosteric regulation of PKM2 allows cellular adaptation to
different physiological states. Sci Signal. 263:pe72013.
|
|
64
|
Fukuda S, Miyata H, Miyazaki Y, Makino T,
Takahashi T, Kurokawa Y, Yamasaki M, Nakajima K, Takiguchi S, Mori
M and Doki Y: Pyruvate kinase M2 modulates esophageal squamous cell
carcinoma chemotherapy response by regulating the pentose phosphate
pathway. Ann Surg Oncol. 22 (Suppl 3):S1461–S1468. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Israelsen WJ, Dayton TL, Davidson SM,
Fiske BP, Hosios AM, Bellinger G, Li J, Yu Y, Sasaki M, Horner JW,
et al: PKM2 isoform-specific deletion reveals a differential
requirement for pyruvate kinase in tumor cells. Cell. 155:397–409.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Keller KE, Tan IS and Lee YS: SAICAR
stimulates pyruvate kinase isoform M2 and promotes cancer cell
survival in glucose-limited conditions. Science. 338:1069–1072.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Yang W, Xia Y, Hawke D, Li X, Liang J,
Xing D, Aldape K, Hunter T, Alfred Yung WK and Lu Z: PKM2
phosphorylates histone H3 and promotes gene transcription and
tumorigenesis. Cell. 150:685–696. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Anastasiou D, Poulogiannis G, Asara JM,
Boxer MB, Jiang JK, Shen M, Bellinger G, Sasaki AT, Locasale JW,
Auld DS, et al: Inhibition of pyruvate kinase M2 by reactive oxygen
species contributes to cellular antioxidant responses. Science.
334:1278–1283. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Luo W, Hu H, Chang R, Zhong J, Knabel M,
O'Meally R, Cole RN, Pandey A and Semenza GL: Pyruvate kinase M2 is
a PHD3-stimulated coactivator for hypoxia-inducible factor 1. Cell.
145:732–744. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Chaneton B, Hillmann P, Zheng L, Martin
ACL, Maddocks ODK, Chokkathukalam A, Coyle JE, Jankevics A, Holding
FP, Vousden KH, et al: Serine is a natural ligand and allosteric
activator of pyruvate kinase M2. Nature. 491:458–462. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Goh GB, Li JW, Chang PE, Chow KY and Tan
CK: Deciphering the epidemiology of hepatocellular carcinoma
through the passage of time: A study of 1,401 patients across 3
decades. Hepatol Commun. 1:564–571. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Busby J, Mills K, Zhang S, Liberante F and
Cardwell C: Postdiagnostic Calcium channel blocker use and breast
cancer mortality: A population-based cohort study. Epidemiology.
29:407–413. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Nanavaty P, Alvarez MS and Alberts WM:
Lung cancer screening: Advantages, controversies, and applications.
Cancer Control. 21:9–14. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Lu M, Lu L, Dong Q, Yu G, Chen J, Qin L,
Wang L, Zhu W and Jia H: Elevated G6PD expression contributes to
migration and invasion of hepatocellular carcinoma cells by
inducing epithelial-mesenchymal transition. Acta Biochim Biophys
Sin (Shanghai). 50:370–380. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Kowalik M, Guzzo G, Morandi A, Perra A,
Menegon S, Masgras I, Trevisan E, Angioni MM, Fornari F, Quagliata
L, et al: Metabolic reprogramming identifies the most aggressive
lesions at early phases of hepatic carcinogenesis. Oncotarget.
7:32375–32393. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Kong DH, Li S, Du ZX, Liu C, Liu BQ, Li C,
Zong ZH and Wang HQ: BAG3 elevation inhibits cell proliferation via
direct interaction with G6PD in hepatocellular carcinomas.
Oncotarget. 7:700–711. 2016.PubMed/NCBI
|
|
77
|
Hong X, Song R, Song H, Zheng T, Wang J,
Liang Y, Qi S, Lu Z, Song X, Jiang H, et al: PTEN antagonises
Tcl1/hnRNPK-mediated G6PD pre-mRNA splicing which contributes to
hepatocarcinogenesis. Gut. 63:1635–1647. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Maruyama H, Kleeff J, Wildi S, Friess H,
Büchler MW, Israel MA and Korc M: Id-1 and Id-2 are overexpressed
in pancreatic cancer and in dysplastic lesions in chronic
pancreatitis. Am J Pathol. 155:815–822. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Yin X, Tang B, Li JH, Wang Y, Zhang L, Xie
XY, Zhang BH, Qiu SJ, Wu WZ and Ren ZG: ID1 promotes hepatocellular
carcinoma proliferation and confers chemoresistance to oxaliplatin
by activating pentose phosphate pathway. J Exp Clin Cancer Res.
36:1662017. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
DeWaal D, Nogueira V, Terry AR, Patra KC,
Jeon SM, Guzman G, Au J, Long CP, Antoniewicz MR and Hay N:
Hexokinase-2 depletion inhibits glycolysis and induces oxidative
phosphorylation in hepatocellular carcinoma and sensitizes to
metformin. Nat Commun. 9:4462018. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Lin YH, Wu MH, Huang YH, Yeh CT, Cheng ML,
Chi HC, Tsai CY, Chung IH, Chen CY and Lin KH: Taurine up-regulated
gene 1 functions as a master regulator to coordinate glycolysis and
metastasis in hepatocellular carcinoma. Hepatology. 67:188–203.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Jin F, Wang Y, Zhu Y, Li S, Liu Y, Chen C,
Wang X, Zen K and Li L: The miR-125a/HK2 axis regulates cancer cell
energy metabolism reprogramming in hepatocellular carcinoma. Sci
Rep. 7:30892017. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Li M, Jin R, Wang W, Zhang T, Sang J, Li
N, Han Q, Zhao W, Li C and Liu Z: STAT3 regulates glycolysis via
targeting hexokinase 2 in hepatocellular carcinoma cells.
Oncotarget. 8:24777–24784. 2017.PubMed/NCBI
|
|
84
|
Lee H, Kim H, Son T, Jeong Y, Kim SU, Dong
SM, Park YN, Lee JD, Lee JM and Park JH: Regulation of HK2
expression through alterations in CpG methylation of the HK2
promoter during progression of hepatocellular carcinoma.
Oncotarget. 7:41798–41810. 2016.PubMed/NCBI
|
|
85
|
Dong T, Kang X, Liu Z, Zhao S, Ma W, Xuan
Q, Liu H, Wang Z and Zhang Q: Altered glycometabolism affects both
clinical features and prognosis of triple-negative and neoadjuvant
chemotherapy-treated breast cancer. Tumour Biol. 37:8159–8168.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Pu H, Zhang Q, Zhao C, Shi L, Wang Y, Wang
J and Zhang M: Overexpression of G6PD is associated with high risks
of recurrent metastasis and poor progression-free survival in
primary breast carcinoma. World J Surg Oncol. 13:3232015.
View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Tseng CW, Kuo WH, Chan SH, Chan HL, Chang
KJ and Wang LH: Transketolase regulates the metabolic switch to
control breast cancer cell metastasis via the α-ketoglutarate
signaling pathway. Cancer Res. 78:2799–2812. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Gao Y, Yang Y, Yuan F, Huang J, Xu W, Mao
B, Yuan Z and Bi W: TNFα-YAP/p65-HK2 axis mediates breast cancer
cell migration. Oncogenesis. 6:e3832017. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Gallagher LE, Radhi OA, Abdullah MO,
McCluskey AG, Boyd M and Chan EYW: Lysosomotropism depends on
glucose: A chloroquine resistance mechanism. Cell Death Dis.
8:e30142017. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Marini C, Bianchi G, Buschiazzo A, Ravera
S, Martella R, Bottoni G, Petretto A, Emionite L, Monteverde E,
Capitanio S, et al: Divergent targets of glycolysis and oxidative
phosphorylation result in additive effects of metformin and
starvation in colon and breast cancer. Sci Rep. 6:195692016.
View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Doménech E, Maestre C, Esteban-Martínez L,
Partida D, Pascual R, Fernández-Miranda G, Seco E, Campos-Olivas R,
Pérez M, Megias D, et al: AMPK and PFKFB3 mediate glycolysis and
survival in response to mitophagy during mitotic arrest. Nat Cell
Biol. 17:1304–1316. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Ge X, Lyu P, Gu Y, Li L, Li J, Wang Y,
Zhang L, Fu C and Cao Z: Sonic hedgehog stimulates glycolysis and
proliferation of breast cancer cells: Modulation of PFKFB3
activation. Biochem Biophys Res Commun. 464:862–868. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Dasgupta S, Rajapakshe K, Zhu B, Nikolai
BC, Yi P, Putluri N, Choi JM, Jung SY, Coarfa C, Westbrook TF, et
al: Metabolic enzyme PFKFB4 activates transcriptional coactivator
SRC-3 to drive breast cancer. Nature. 556:249–254. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Zhou Z, Li M, Zhang L, Zhao H, Şahin Ö,
Chen J, Zhao JJ, Songyang Z and Yu D: Oncogenic kinase-induced PKM2
tyrosine 105 phosphorylation converts non-oncogenic PKM2 to a tumor
promoter and induces cancer stem-like cells. Cancer Res.
78:2248–2261. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Singh S, Narayanan S, Biswas K, Gupta A,
Ahuja N, Yadav S, Panday RK, Samaiya A, Sharan SK and Shukla S:
Intragenic DNA methylation and BORIS-mediated cancer-specific
splicing contribute to the Warburg effect. Proc Natl Acad Sci USA.
114:11440–11445. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Giatromanolaki A, Sivridis E, Arelaki S
and Koukourakis M: Expression of enzymes related to glucose
metabolism in non-small cell lung cancer and prognosis. Exp Lung
Res. 43:167–174. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Nagashio R, Oikawa S, Yanagita K, Hagiuda
D, Kuchitsu Y, Igawa S, Naoki K, Satoh Y, Ichinoe M, Murakumo Y, et
al: Prognostic significance of G6PD expression and localization in
lung adenocarcinoma. Biochim Biophys Acta Proteins Proteom.
1867:38–46. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Hong W, Cai P, Xu C, Cao D, Yu W, Zhao Z,
Huang M and Jin J: Inhibition of Glucose-6-phosphate dehydrogenase
reverses cisplatin resistance in lung cancer cells via the redox
system. Front Pharmacol. 9:432018. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Chan B, VanderLaan P and Sukhatme VP:
6-Phosphogluconate dehydrogenase regulates tumor cell migration in
vitro by regulating receptor tyrosine kinase c-Met. Biochem Biophys
Res Commun. 439:247–251. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Kayser G, Sienel W, Kubitz B, Mattern D,
Stickeler E, Passlick B, Werner M and Zur Hausen A: Poor outcome in
primary non-small cell lung cancers is predicted by transketolase
TKTL1 expression. Pathology. 43:719–724. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Lu H and Zhu H: Effect of siRNA-mediated
gene silencing of transketolase on A549 lung cancer cells. Oncol
Lett. 14:5906–5912. 2017.PubMed/NCBI
|
|
102
|
Wang H, Wang L, Zhang Y, Wang J, Deng Y
and Lin D: Inhibition of glycolytic enzyme hexokinase II (HK2)
suppresses lung tumor growth. Cancer Cell Int. 16:92016. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
De Rosa V, Iommelli F, Monti M, Fonti R,
Votta G, Stoppelli MP and Del Vecchio S: Reversal of Warburg effect
and reactivation of oxidative phosphorylation by differential
inhibition of EGFR signaling pathways in non-small cell lung
cancer. Clin Cancer Res. 21:5110–5120. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Su H, Bodenstein C, Dumont RA, Seimbille
Y, Dubinett S, Phelps ME, Herschman H, Czernin J and Weber W:
Monitoring tumor glucose utilization by positron emission
tomography for the prediction of treatment response to epidermal
growth factor receptor kinase inhibitors. Clin Cancer Res.
12:5659–5667. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Zhang K, Zhang M, Jiang H, Liu F, Liu H
and Li Y: Down-regulation of miR-214 inhibits proliferation and
glycolysis in non-small-cell lung cancer cells via down-regulating
the expression of hexokinase 2 and pyruvate kinase isozyme M2.
Biomed Pharmacother. 105:545–552. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Cheng X, Liu F, Liu H, Wang G and Hao H:
Enhanced glycometabolism as a mechanism of NQO1 potentiated growth
of NSCLC revealed by metabolomic profiling. Biochem Biophys Res
Commun. 496:31–36. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Minchenko OH, Ogura T, Opentanova IL,
Minchenko DO, Ochiai A, Caro J, Komisarenko SV and Esumi H:
6-Phospho-fructo-2-kinase/fructose-2,6-bisphosphatase gene family
overexpression in human lung tumor. Ukr Biokhim Zh (1999).
77:46–50. 2005.PubMed/NCBI
|
|
108
|
Yang J, Li J, Le Y, Zhou C, Zhang S and
Gong Z: PFKL/miR-128 axis regulates glycolysis by inhibiting AKT
phosphorylation and predicts poor survival in lung cancer. Am J
Cancer Res. 6:473–485. 2016.PubMed/NCBI
|