Introduction
One of the primary mechanisms responsible for cancer
chemotherapeutic failure is the development of multidrug resistance
(MDR), a phenomenon that occurs when cancer cells become resistant
to several mechanistically and structurally unrelated
antineoplastic drugs (1). One of
the major mediators of MDR is the ATP binding cassette (ABC)
transporter (1,2). These transporters significantly
decrease the intracellular accumulation of antineoplastic compounds
by increasing their efflux, thereby inducing MDR (1). Currently, 48 human ABC transporters
have been identified and characterized (3). The first identified ABC transporter,
ABCB1, is encoded in the human MDR1 gene (4,5). The
ABCB1 protein consists of two transmembrane domains (TMDs) and two
nucleotide-binding domains (NBDs) (6). The known substrates of ABCB1 include
anthracyclines, vinca alkaloids, taxanes (such as paclitaxel and
docetaxel) and epipodophyllotoxins (7). The ABCB1 protein is widely expressed
in normal human tissues, including intestinal lining epithelium,
bone marrow progenitor cells, hepatocytes, cardiac and striated
muscle and blood-brain barrier (8–10).
One pharmacological approach used to increase the
response of MDR cancer cells to antineoplastic drugs involves the
inhibition of the ABC transporters (11). Research over the past three decades
has led to the development of three generations of ABCB1 inhibitors
to re-sensitize MDR cells to ABCB1 substrates (12–14).
The first generation of ABCB1 inhibitors identified were substrates
of ABCB1, such as verapamil and cyclosporine A (CsA) (15,16)
and were essentially competitive inhibitors of the efflux activity
of different anticancer compounds by the ABCB1 transporter
(17). However, the use of these
compounds in humans was limited as a result of significant adverse
effects (18). The second
generation of ABCB1 inhibitors, such as PSC 833 and biricodar
(VX-710) (19,20), were developed and subsequent studies
indicated that compared to the first generation, these compounds
were superior as they are more potent and less toxic. However, a
number of these second generation compounds were inhibitors of
CYP3A4 and this led to a significant increase in the levels of the
substrate antineoplastic drugs and produced significant adverse and
toxic effects (21). The third
generation of modulators, such as ONT-093, tariquidar, XR9576,
zosuquidar, laniquidar, exhibited high affinity for the ABCB1
transporter but did not significantly alter the metabolism of the
antineoplastic substrate drugs and some of them are currently in
clinical trials (22–25).
β-elemene, a compound isolated from an extract of
traditional Chinese medicinal herb Rhizoma zedoariae, is a
broad spectrum antineoplastic drug which has been approved by the
state Food and Drug Administration of China for the treatment of
specific solid and malignant tumors (26,27).
However the mechanism of action of β-elemene has not been fully
elucidated. It was suggested that β-elemene significantly enhanced
the cytotoxicity of cisplatin in treating non-small cell lung
cancer cells. This aforementioned effect produced by the β-elemene
and cisplatin is due to an increase in the release of cytochrome
c from mitochondria into the cytoplasm, as well as
increasing caspase-3/7/9 activities and caspase-9 cleavage
(28). It was previously reported
that β-elemene significantly increased the intracellular
accumulation of the antineoplastic drug adriamycin (ADM) in MCF/ADM
cells (29). Since ADM is a
substrate of ABCB1 and breast cancer resistance protein
(BCRP/ABCG2), it is possible that the β-elemene-induced increase in
ADM levels was due to its inhibition of the ABCB1 and/or BCRP
transporters (30,31). However, the present study did not
determine which transporter or transporters were affected by
β-elemene or the mechanism by which β-elemene increased the
intracellular levels of ADM. Therefore, in this in vitro
study, we examined the effect of the compound β-elemene (see
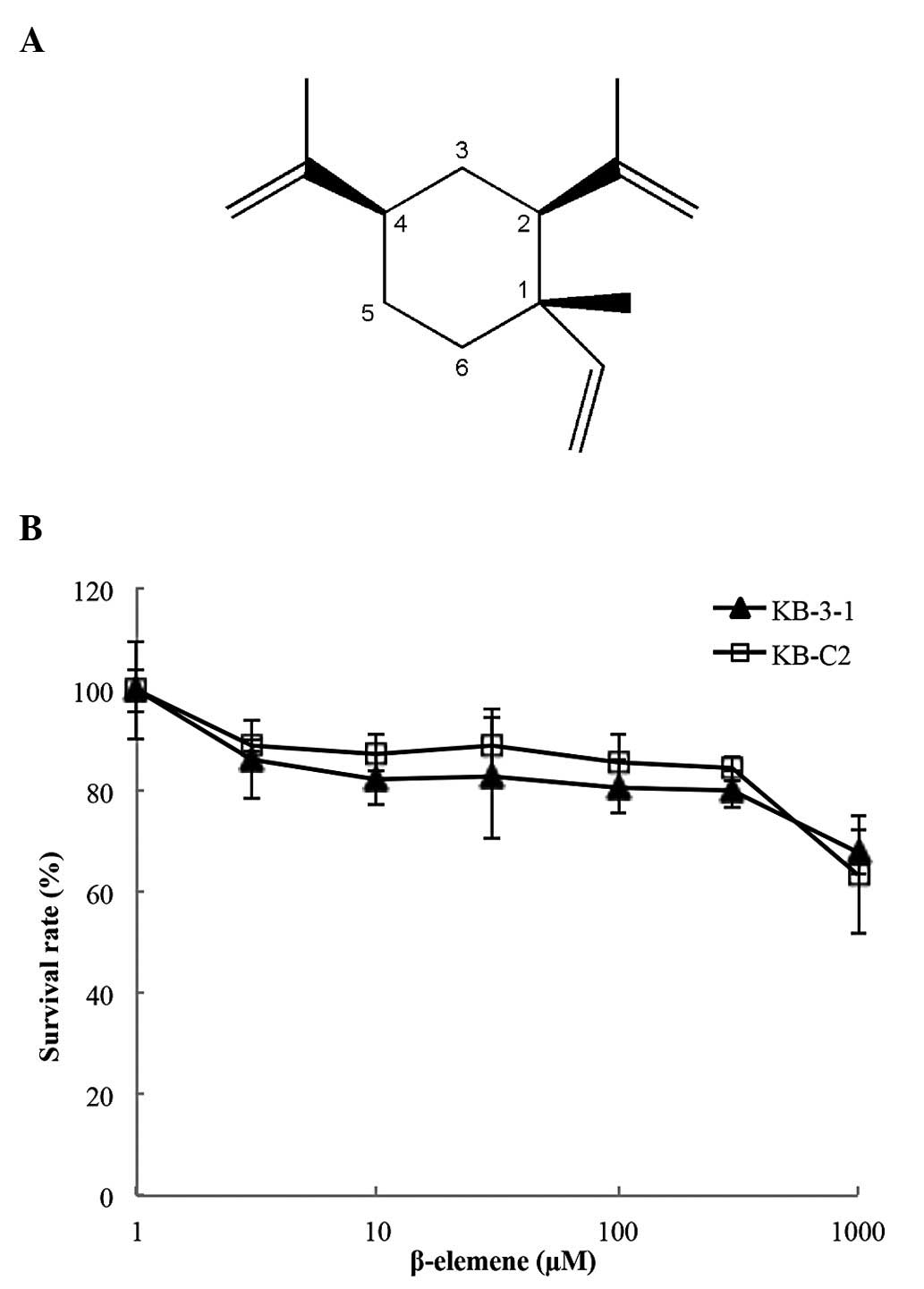
Fig. 1 for structure), on the
response of cells to substrates for the ABCB1 (P-glycoprotein/P-gp,
MDR), multidrug resistance protein 1 [(MRP1), ABCC1] and BCRP
(ABCG2/MXR). In addition, we examined the effect of β-elemene on
the efflux of [3H]-paclitaxel and on the expression of
the ABCB1 protein in cells overexpressing the ABCB1 transporter.
Finally, using docking analysis, the interaction of β-elemene with
the ABCB1 transporter was elucidated.
Materials and methods
Chemicals and equipment
β-elemene was provided by Dalian Jingang
Pharmaceutical Co. Ltd. (Dalian, Liaoning, China). Dulbecco’s
modified Eagle’s medium (DMEM), fetal bovine serum,
penicillin/streptomycin and trypsin were purchased from HyClone
Thermo Scientific (Logan, UT, USA). The monoclonal mouse antibody
C219 (against ABCB1) and the secondary horseradish
peroxidase-labeled anti-goat or anti-mouse IgG were purchased from
Signet Laboratory (Dedham, MA, USA). Paclitaxel, vincristine,
verapamil, vinblastine, cochicine, cisplatin,
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
were purchased from Sigma-Aldrich Chemical Co. (St. Louis, MO,
USA). [3H]-paclitaxel (37.9 Ci/mmol) was purchased from
Moravek Biochemicals Inc. (Brea, CA, USA). Nilotinib was purchased
from Chemie Tek Co. (Indianapolis, IN). Opsys microplate reader was
purchased from Dynex Technologies Inc. (Chantilly, VA, USA).
Cell lines
The ABCB1-overexpressing KB-C2 cells were cloned in
cell culture medium by a step-wise selection of the parental human
epidermoid carcinoma cell line KB-3-1, using colchicine at
concentrations up to 2 μg/ml (32).
NCI-H460/MX20 and lung cancer cell lines NCI-H460 (parental),
HEK293/pcDNA3.1 and wild-type HEK293/ABCB1-transfected cells were
kindly provided by Drs Susan Bates and Robert Robey (NCI, NIH,
Bethesda, MD, USA). HEK293/ABCB1 cells were obtained by
transfecting HEK293 cell lines with a pcDNA3.1 vector containing a
full-length cDNA of ABCB1. All cell lines were grown in
DMEM, supplemented with 10% bovine serum, 100 U/ml penicillin, and
100 mg/ml streptomycin in a humidified incubator with 5%
CO2 at 37°C.
MTT assays
Cell lines were seeded evenly in (160 μl/well)
96-well plates and were cultured for 24 h (33). β-elemene with different
concentrations (10, 30, 100 μM) was added into wells 1 h before
adding the appropriate antineoplastic drug colchicine, vinblastine
and paclitaxel that were diluted to various concentrations and
incubated with cells for 72 h. Subsequently, 20 μl of MTT was added
to each well. The plates were placed in an incubator at 37°C for 4
h. In order to avoid agitating the adhesive cell, the medium and
MTT were gently discarded. Subsequently, 100 μl DMSO was added into
each well. The absorbance of each sample at 570 nm was determined
using an Opsys microplate reader. The magnitude of resistance was
calculated by dividing the IC50 (concentration required
to inhibit cellular proliferation by 50%) for the MDR cells in the
absence or presence of inhibitors by the IC50 of the
parental cells in the absence of the reversal compound.
[3H]-paclitaxel accumulation
assays
HEK293/pcDNA3.1 and HEK293/ABCB1 were seeded into
two T75 flasks and incubated in DMEM at 37°C for 24 h (34). Cells were harvested upon reaching
70–95% confluence and two aliquots from each cell line were
suspended in DMEM. Subsequently, HEK293/pcDNA3.1 and HEK293/ABCB1
were incubated with 0.1 μM [3H]-paclitaxel in the
presence or absence of the β-elemene at 37°C for 2 h. Cells were
washed 3 times with cold phosphate-buffered saline (PBS) and 200 μl
of lysis buffer (pH 7.4, containing 1% Triton X-100 and 0.2% SDS).
The sample was placed in scintillation fluid and radioactivity was
determined using a Packard Tri-Carb 1900CA liquid scintillation
analyzer (Packard Instrument Company, Inc. (Downers Grove, IL,
USA).
[3H]-paclitaxel efflux
assays
HEK293/pcDNA3.1- and HEK293/ABCB1-transfected cells
were prepared as described for the accumulation experiments
(34,35). Cells were exposed to the same
procedure as stated for the drug accumulation experiment. The cells
were rinsed 3 times with cold PBS and incubated with or without 100
μM β-elemene at 37°C. The cells were lysed at various time points
(0, 30, 60, 120 min) and radioactivity in the lysate was determined
using liquid scintillation counter.
Western blotting
NCI-H460/MX20, NCI-H460, KB-3-1 and KB-C2 were
incubated with 100 μM of β-elemene for 0, 24, 48 or 72 h (36). The cells were harvested and washed 3
times with PBS. The cell extracts were incubated with
radioimmunoprecipitation assay (RIPA) buffer (PBS with 0.1% SDS, 1%
Nonidet P-40, 0.5% sodium deoxycholate and 100 mg/ml
p-aminophenylmethylsulfonyl fluoride) for 30 min and the cells were
centrifuged at 12,000 × g for 15 min at 4°C. The total cell lysate
was dissolved in SDS-PAGE and electrophoretically transferred onto
polyvinylidene fluoride (PVDF) membranes. The membranes were
incubated in blocking solution containing 5% non-fat milk in TBST
buffer [10 mM Tris-HCL (pH 8.0), 150 mM NaCl and 0.1% Tween-20] for
1 h at room temperature. Subsequently, the membranes were incubated
for 12 h with monoclonal antibodies against ABCB1 protein at 1:200
dilution, and then further incubated for 3 h at room temperature
with a horseradish peroxide (HRP)-conjugated secondary anti-mouse
antibody (1:1,000 dilution). The presence of the protein-antibody
complex was detected by chemiluminescence. The expression level of
the protein was measured by determining the area and density of
bands, using Scion Image software (Scion Co., Frederick, MD,
USA).
Immunofluorescence assays
KB-3-1 and KB-C2 cells were seeded in 24-well plates
and 100 μM of β-elemene was added to each well and the samples were
incubated for 0 and 72 h at 37°C. The cells were washed with PBS
and fixed in 4% paraformaldehyde for 15 min at room temperature and
rinsed 3 times with PBS. The cells were incubated with BSA (2
mg/ml) for 1 h at 37°C. Subsequently, cells were incubated with a
polyclonal antibody against ABCB1 (1:200; Santa Cruz Biotechnology,
Inc., Santa Cruz, CA, USA) for 12 h, followed by a goat anti-mouse
IgG (1:100; Molecular Probes, Eugene, OR, USA) for 1 h.
2-(4-Amidinophenyl)-6-indolecarbamidine dihydrochloride (DAPI) (1
ml/well) was applied to counterstain the nuclei. Finally, images
were captured by an IX70 microscope with IX-FLA fluorescence and
CCD camera (36).
Ligand structure preparation
The structure of β-elemene was constructed using the
fragment dictionary of Maestro v9.0 and the energy was minimized by
a MacroModel program v9.7 (Schrödinger, Inc., New York, NY, USA,
2009) using the OPLS-AA force-field with the steepest descent
followed by truncated Newton conjugate gradient protocol. The
low-energy 3D structures of β-elemene were generated by LigPrep
v2.3. Ligand structures representing β isomer were further used for
generating 100 ligand conformations using the default parameters of
mixed torsional/low-mode sampling function. The conformations were
filtered with a maximum relative energy difference of 5 kcal/mol to
exclude redundant conformers. The output conformational search
(Csearch) file containing at most 100 unique conformers of
β-elemene were used as input for docking simulations into each of
the binding sites of human ABCB1 (37).
Protein structure preparation
The X-ray crystal structure of mouse ABCB1 in the
apoprotein state (PDB ID: 3G5U), and in complex with the ABCB1
inhibitors QZ59-RRR (PDB ID: 3G60), QZ59-SSS (PDB ID:
3G61) (37), were obtained from the
RCSB Protein Data Bank and were used as the templates to generate
homology models for human ABCB1 for sites 1–4 (38). The protocol for the homology
modeling was essentially the same as reported previously (38). The refined human ABCB1 homology
model was used to generate different receptor grids for different
sites (sites 1–4) through selection of bound QZ59-RRR
(site-1) and QZ59-SSS (site-2) ligands, all amino acid
residues known to contribute to verapamil binding (site-3), two
residues known to be common to three previously determined sites
(site-4) as previously reported (38).
Docking protocol
The conformational library of β-elemene was docked
at each of the generated grids (sites 1–4) by using the XP mode of
Glide 5.0 (Schrödinger, Inc., 2009) and the default parameters. The
top-scoring pose was used for graphical analysis. All computations
were performed with a Dell Precision 470n dual-processor computer
with the Linux operating system (Red Hat Enterprise WS 4.0).
Statistical analysis
All experiments were performed in triplicate. The
data were analyzed using two-tail Student’s t-test. The a
priori significance level was set at p<0.05.
Results
Effect of β-elemene on the sensitivity of
ABCB1-, MRP1- and BCRP-overexpressing cells to antineoplastic
drugs
In order to determine if β-elemene alone produces
significant in vitro cytotoxicity, we examined its effect on
the survival of KB-3-1 and KB-C2 cell lines using the MTT assay.
The results indicated that up to a concentration of 100 μM,
β-elemene did not significantly alter the survival rate of either
KB-3-1 or KB-C2 cells (Fig. 1B).
Based on these results, the highest concentration of β-elemene we
used was 100 μM. Subsequently, we determined the effect of
β-elemene at concentrations of 10, 30 or 100 μM on the response of
KB-3-1 or KB-C2 cells to the antineoplastic drugs paclitaxel,
vinblastine and colchicine. As shown in Table I, at a concentration of 10 μM,
β-elemene significantly decreased the IC50 values for
the ABCB1 substrates paclitaxel, vinblastine and colchicine in the
ABCB1-overexpressing cell line KB-C2. However there was no
significant change in the cytotoxicity of these drugs for parental
cell line KB-3-1. The response to paclitaxel, vinblastine and
colchicine in KB-C2 cells was 648-, 11.7- and 720-fold lower,
respectively, than that for the parental KB-3-1 cell line (Table I). The incubation of KB-C2 cells
with β-elemene significantly increased their sensitivity to
paclitaxel, vinblastine and colchicine. For example, in the
presence of 10 μM β-elemene, the response of KB-C2 to paclitaxel,
vinblastine and colchicine was 52.2-, 3.4- and 113.3-fold lower,
respectively (Table I). The
magnitude of drug resistance in the presence of 30 μM of β-elemene
to paclitaxel, vinblastine and colchicine was 16.8-, 2.1- and
28-fold lower, respectively, which was significantly lower than
that for the 10 μM β-elemene concentration (Table I). The 100 μM concentration of
β-elemene significantly lowered the magnitude of resistance to
paclitaxel, vinblastine and colchicine (4.6-, 5.4- and 1.1-fold,
respectively), when compared to the 10 and 30 μM concentrations
(Table I). In fact, the 100 μM
concentration of β-elemene reduced resistance to a level comparable
to that for verapamil (5 μM), a known inhibitor of the ABCB1
transporter (Table I). β-elemene,
at 100 μM, did not significantly alter the response of KB-C2 to
cisplatin, a compound that is not a substrate for ABCB1.
 | Table IThe reversal effect of β-elemene on
ABCB1-mediated drug resistance.a |
Table I
The reversal effect of β-elemene on
ABCB1-mediated drug resistance.a
| IC50 ±
SDb (μM) |
|---|
|
|
|---|
| Compounds | KB-3-1 | KB-C2 |
|---|
| Paclitaxel
(μM) | 0.0050±0.0007
(1.0)c | 3.240±0.901
(648.0) |
| +β-elemene
(10) | 0.0058±0.0015
(1.2) | 0.261±0.019e (52.2) |
| +β-elemene
(30) | 0.0051±0.0011
(1.0) | 0.084±0.014e (16.8) |
| +β-elemene
(100) | 0.0047±0.0021
(0.9) | 0.023±0.005e (4.6) |
| +Verapamil
(10) | 0.0035±0.0007
(0.7) | 0.010±0.003e (2.0) |
| Colchicine
(μM) | 0.0054±0.0015
(1.0)c | 3.891±0.531
(720.6) |
| +β-elemene
(10) | 0.0052±0.0014
(1.0) | 0.612±0.201e (113.3) |
| +β-elemene
(30) | 0.0048±0.0021
(0.9) | 0.152±0.050e (28.1) |
| +β-elemene
(100) | 0.0047±0.0018
(0.9) | 0.029±0.013e (5.4) |
| +Verapamil
(10) | 0.0036±0.0011
(0.7) | 0.025±0.008e (4.6) |
| Vinblastine
(μM) | 0.0411±0.0012
(1.0)c | 0.482±0.111
(11.7) |
| +β-elemene
(10) | 0.0442±0.0113
(1.1) | 0.138±0.073e (3.4) |
| +β-elemene
(30) | 0.0418±0.0087
(1.0) | 0.087±0.009e (2.1) |
| +β-elemene
(100) | 0.0370±0.0096
(0.9) | 0.046±0.011e (1.1) |
| +Verapamil
(10) | 0.0313±0.0003
(0.8) | 0.045±0.008e (1.1) |
| Cisplatin (μM) | 2.444±0.192
(1.0)c | 2.628±0.196
(1.1) |
| +β-elemene
(100) | 2.023±0.157
(0.8) | 2.140±0.287
(0.9) |
| +Verapamil
(10) | 2.361±0.087
(1.0) | 2.639±0.061
(1.0) |
We also determined the response of HEK293 cells
overexpressing ABCB1 transporters to paclitaxel in the presence of
β-elemene. β-elemene, at 100 μM, did not significantly alter the
sensitivity of the empty vector-transfected cell line
HEK293/pcDNA3.1 to paclitaxel (Table
II). In contrast, 30 and 100 μM of β-elemene significantly
increased the sensitivity of HEK293/ABCB1 cells to paclitaxel and
the effect produced by 100 μM of β-elemene was almost identical to
that produced by verapamil (10 μM) (Table II).
| Table IIThe effect of β-elemene and verapamil
on the response of ABCB1-transfected cells to paclitaxel and
cisplatin.a |
Table II
The effect of β-elemene and verapamil
on the response of ABCB1-transfected cells to paclitaxel and
cisplatin.a
| IC50 ±
SDb (μM) |
|---|
|
|
|---|
| Treatment |
HEK293/pcDNA3.1 | HEK293/ABCB1 |
|---|
| Paclitaxel
(μM) | 0.0222±0.0008
(1.0)c | 2.5981±0.1583
(117.2) |
| +β-elemene
(10) | 0.0212±0.0003
(1.0) |
0.2431±0.0721e (11.0) |
| +β-elemene
(30) | 0.0203±0.0039
(0.9) |
0.0628±0.0838e (2.8) |
| +β-elemene
(100) | 0.0206±0.0021
(0.9) |
0.0315±0.0040e (1.4) |
| +Verapamil
(10) | 0.0187±0.0020
(0.8) |
0.0323±0.0030e (1.5) |
| Cisplatin (μM) | 1.9972±0.1916
(1.0)c | 1.9918±0.2714
(1.0) |
| +β-elemene
(100) | 1.9187±0.1446
(1.0) | 1.8500±0.0687
(0.9) |
| +Verapamil
(10) | 1.8445±0.0662
(0.9) | 1.8326±0.2062
(0.9) |
We further determined if β-elemene significantly
reverses multidrug resistance in cell lines that overexpress BCRP
or MRP1. In the parental NCI-H460 and the mitoxantrone-resistant
cells NCI-H460/MX20, which overexpress BCRP, 100 μM of β-elemene
did not significantly alter the sensitivity of the parental
NCI-H460 cells (Table III). In
contrast, 100 μM of β-elemene significantly enhanced the
sensitivity of cells overexpressing the BCRP transporter to
mitoxantrone, a known substrate of BCRP (Table III). In addition, β-elemene (100
μM) did not significantly alter the sensitivity of HEK293 cells
overexpressing MRP1 (HEK293/MRP1) to the MRP1 substrate,
vincristine (Table III).
| Table IIIThe reversal effect of β-elemene on
HEK293/pcDNA3.1, HEK293/MRP1, NCI-H460 and NCI-H460/MX20
cells.a |
Table III
The reversal effect of β-elemene on
HEK293/pcDNA3.1, HEK293/MRP1, NCI-H460 and NCI-H460/MX20
cells.a
| IC50 ±
SDb (μM) |
|---|
|
|
|---|
| Treatment | NCI-H460 | NCI-H460/MX20 |
|---|
| Mitoxantrone
(μM) | 0.15±0.06
(1.0)c | 18.8±1.30
(125.0) |
| +β-elemene
(100) | 0.10±0.01
(0.8) | 9.20±1.40e (61.3) |
| +Nilotinib
(5) | 0.13±0.04
(0.9) | 0.24±0.07f (1.6) |
|
| IC50 ±
SDb (μM) |
|
|
| Treatment |
HEK293/pcDNA3.1 | HEK293/MRP1 |
|
| Vincristine
(μM) | 0.004±0.0003
(1.0)d | 0.055±0.0120
(13.8) |
| +β-elemene
(100) | 0.004±0.0006
(1.0) | 0.050±0.0009
(12.7) |
| +ONO-1078 (5) | 0.0037±0.0006
(0.9) |
0.006±0.0011f
(1.6) |
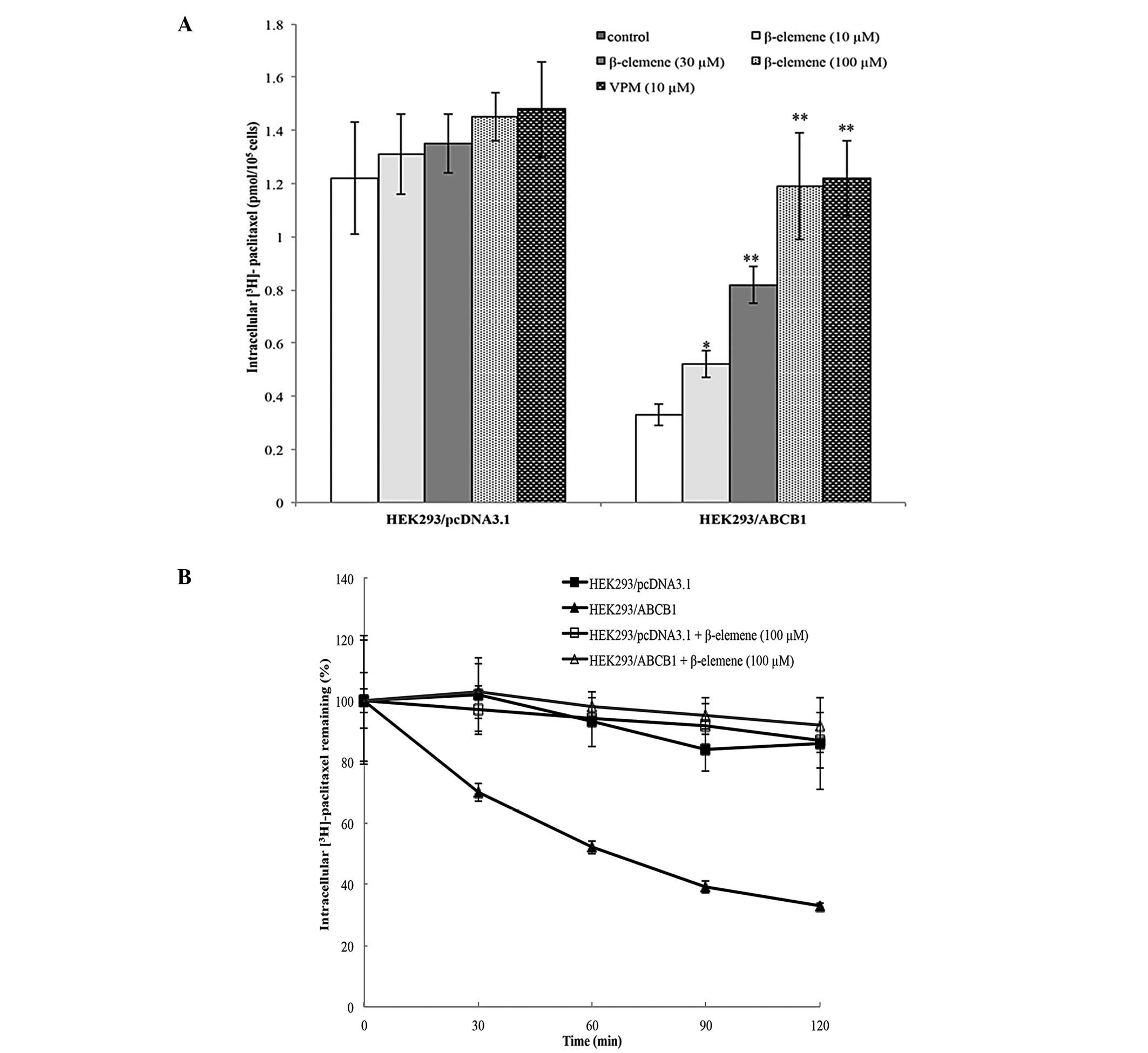
Effect of β-elemene on the accumulation
of [3H]-paclitaxel in HEK293/ABCB1 cells
In order to understand the mechanism of
β-elemene-induced sensitization of MDR cell lines to antineoplastic
drugs, we examined the effect of β-elemene on the intracellular
accumulation of [3H]-paclitaxel in HEK293 cells
overexpressing the ABCB1 transporter. Overall, in the absence of
β-elemene, the accumulation of [3H]-paclitaxel was 88%
lower in the HEK293/ABCB1 cells when compared to the
HEK293/pcDNA3.1 cells that do not overexpress ABCB1 (Fig. 2A). However, the incubation of
HEK293/ABCB1 cells with β-elemene (100 μM) completely reversed the
accumulation of [3H]-paclitaxel to a level comparable to
that of the parental cell line (Fig.
2A). These results suggested that β-elemene may potentiate the
cytotoxicity of paclitaxel by increasing the intracellular
accumulation of antineoplastic drugs.
Effect of β-elemene on the efflux of
[3H]-paclitaxel in HEK293/ABCB1 cells
In these experiments, we determined the effect of
β-elemene on the efflux of [3H]-paclitaxel in the
parental HEK293/pcDNA3.1 and the ABCB1-overexpressing cell line
HEK293/ABCB1. The efflux of [3H]-paclitaxel from the
ABCB1-overexpressing cells was significantly greater than that of
the non-overexpressing parental cells (Fig. 2B). However, in the presence of 100
μM of β-elemene, the efflux of [3H]-paclitaxel from the
HEK293/ABCB1 cells was significantly reduced when compared to
controls (Fig. 2B). These results
indicated that β-elemene sensitized ABCB1-overexpressing cells to
paclitaxel by blocking its efflux via the ABCB1 transporter.
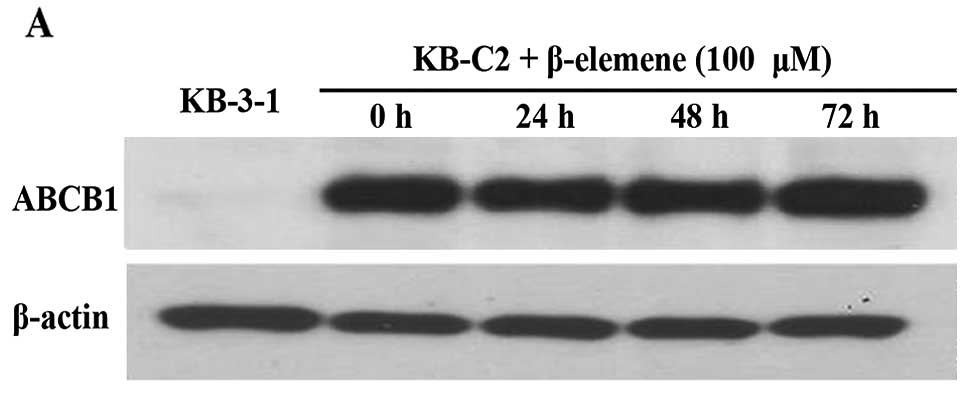
Effect of β-elemene on the expression of
ABCB1 and BCRP protein levels
It is possible that β-elemene increases the
intracellular accumulation of [3H]-paclitaxel by
downregulating the expression of the ABCB1 protein as opposed to
simply blocking efflux activity of ABCB1. To further delineate the
mechanism of action of β-elemene, we examined its effect on the
level of ABCB1 protein expression in KB-3-1 and KB-C2 cells. The
incubation of the cell lines with 100 μM of β-elemene for either
24, 48 or 72 h did not significantly alter the expression level of
the ABCB1 protein (Fig. 3A). Thus,
it is unlikely that the β-elemene-induced increase in the
intracellular levels of [3H]-paclitaxel is due to the
downregulation of expression of the ABCB1 transporter.
Effect of β-elemene on the subcellular
localization of the ABCB1 transporter
In these experiments, we sought to ascertain if
β-elemene altered the cellular localization of the ABCB1
transporter. The incubation of KB-3-1 and KB-C2 cells with 100 μM
of β-elemene for 72 h did not alter the cellular distribution of
the ABCB1 transporter when compared to cells exposed only to
incubation solution (Fig. 3C).
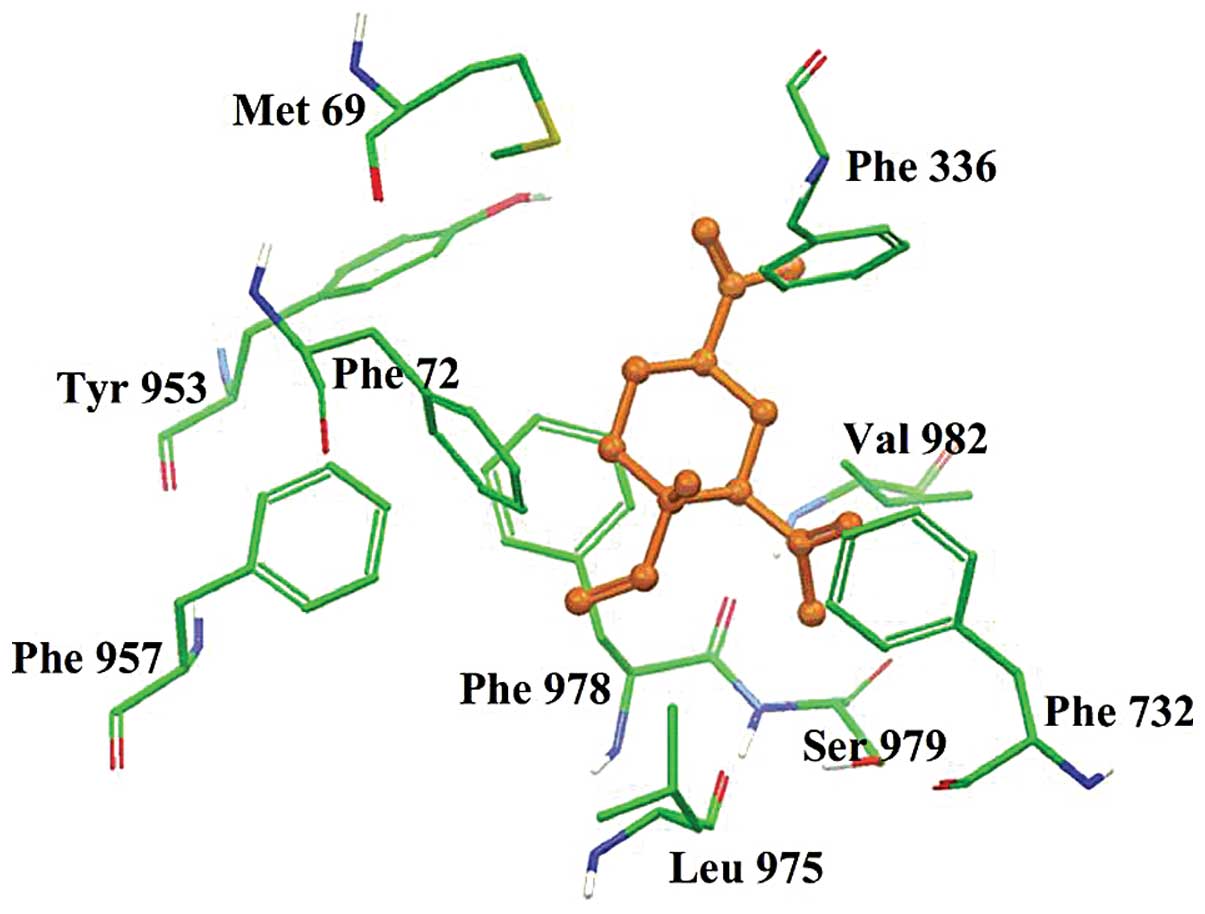
Calculation of the interaction between
β-elemene and the ABCB1 protein using docking analysis
The interaction between β-elemene and the ABCB1
transporter was determined using the docking analysis technique. A
docking score was calculated for each of the 4 possible binding
sites on the ABCB1 transporter for β-elemene. The docking score
(site-1, -7.4 kcal/mol; site-2, -3.7 kcal/mol; site-3, -4.8
kcal/mol and site-4, -5.5 kcal/mol) clearly indicated that site-1
of ABCB1 is the best docking or binding site for β-elemene.
Therefore, the XP-Glide predicted binding model for β-elemene at
site-1 of ABCB1 is discussed below and shown in Fig. 4.
Since β-elemene contains only hydrocarbon atoms, the
primary factor contributing to the binding of β-elemene to the
ABCB1 protein is via hydrophobic interactions. The 3 carbon-carbon
double bonds located at the C4, C2 and
C1 (atom nos. shown in Fig.
1A) side chains of β-elemene are stabilized by π-π interactions
with the phenyl rings of Phe336, Phe732 and Phe978, respectively.
The side chain at the C1 atom is stabilized by Leu975,
Phe978 and Ser979, whereas the side chain at the C2 atom
is stabilized by Phe732, Ser979 and Val982. The cyclohexyl ring of
β-elemene is positioned in the large hydrophobic cavity formed by
the side chains of Met69, Phe72, Tyr953, Phe957 and Val982.
Discussion
One of the major findings of the present in
vitro study is that in cell lines overexpressing the ABCB1
transporter, β-elemene significantly increased the sensitivity to
the ABCB1 substrates paclitaxel, colchicine and vinblastine. This
effect was specific, as β-elemene did not significantly alter the
response of cells to cisplatin, which is not a substrate for ABCB1.
Furthermore, β-elemene did not significantly affect the sensitivity
of the ABCB1-negative cell lines KB-3-1 and HEK/293pcDNA3.1.
In contrast to the result of a previous study
(29), β-elemene produced a
significant reversal of MDR by overexpression of the ABCB1
transporter (see Tables I and
II). The discrepancy between the
present study and that of Hu et al may be due to use of
different cell lines (MCF-7/ADM and MCF vs. KB-C2 and HEK293
overexpressing ABCB1). In the present study, 100 μM of β-elemene
significantly reduced the IC50 value of paclitaxel to
0.23 μM, a result that is comparable to that of the TKI lapatinib
(IC50 of 0.18 μM), an established ABCB1 inhibitor
(39). However, β-elemene
moderately enhanced the sensitivity or response to mitoxantrone in
cell lines overexpressing the BCRP transporter (NCI-H460/MX20) and
did not significantly reverse MRP1-mediated vincristine resistance.
Therefore, β-elemene selectively increases the response of cell
lines overexpressing the ABCB1 transporter to certain
antineoplastic drugs.
Our results indicated that at 10 μM, β-elememe was
less potent than vardenafil (a PDE5 and ABCB1 inhibitor) in
decreasing the fold-resistance value of paclitaxel (12.4 vs. 54.1)
and the colchicine (184 vs. 6.4) in KB-C2 cells overexpressing the
ABCB1 transporter (40). However,
in HEK293-transfected cells overexpressing the ABCB1 transporter,
the fold-resistance value of paclitaxel for 10 μM of β-elememe
(10.6) was greater than that for vardenafil (8.7) (40). In contrast, in KB-C2 cells, the
fold-resistance for paclitaxel for 10 μM of tadalafil (a PDE5
inhibitor) was lower than that for 10 μM of β-elememe (4.3 vs.
12.4) and there was no difference in the fold-resistance value for
colchicine between 10 μM of vardenafil (6.38) (42) and 10 μM of β-elememe (6.36).
Another finding of the present study was that 100 μM
of β-elememe significantly enhanced (3.6-fold) the intracellular
accumulation of [3H]-paclitaxel in HEK293 cells
transfected with the ABCB1 gene (Fig. 2A). These findings are similar to
those reported for HEK293/ABCB1 cells incubated with other ABCB1
inhibitors, such as PD173074, pargueren (5.6-fold increase) and
vardenafil (3.5-fold increase) (40–42).
Additional experiments indicated that the intracellular increase in
[3H]-paclitaxel levels is due to β-elemene’s inhibition
of the efflux function of the ABCB1 transporter.
Previously, it was reported that 30 μM of β-elemene
significantly downregulated the levels of the ABCB1 protein in
vitro in the MCF-7 cell line (43). In contrast, our results indicated
that β-elemene (100 μM for 24, 48 and 72 h) did not significantly
alter the level of ABCB1 protein expression. The exact explanation
for this discrepancy is uncertain but it may be due, in part, to
differences in the cell lines used in the present study (KB-C2 and
HEK293/ABCB1) when compared to that of Xu et al (MCF-7/DOX)
(43). In addition, the results of
immunofluorescence assays indicated that the incubation of KB-3-1
and KB-C2 cells with 100 μM of β-elemene for 72 h did not
significantly alter the cellular distribution of the ABCB1
transporter when compared to cells exposed only to the incubation
solution (Fig. 3C). Overall, our
results indicated that β-elemene significantly increased the
sensitivity of the cell lines we used by inhibiting the efflux
function of the ABCB1 transporter.
In order to characterize the binding interaction of
β-elemene with ABCB1, docking study was carried out to describe the
site on which β-elemene functioned. The docking simulations of
β-elemene on ABCB1 were performed on a homology model of human
ABCB1 due to the lack of the co-crystal structures of human ABCB1.
To identify the predicted binding site of β-elemene, docking
studies were performed at 4 possible binding sites of ABCB1 as
described by Aller et al (37). The most favorable binding site is
identified as site-1 by exploring docking scores of β-elemene at
all of the binding sites. The drug-binding pocket located in
transmembrane domains of human ABCB1 contains predominantly
hydrophobic and aromatic residues as illustrated by Aller et
al (37). Then, the relatively
high affinity of the docking pose showed 3 terminal located double
bonds of β-elemene forming π-π interactions with aromatic residues.
These interactions may be critical in restricting the ligand at the
correct position of binding cavity of ABCB1. Overall, these results
suggest that β-elemene directly interacts with the substrate
binding pocket of the ABCB1 transporter. However, further studies,
using other approaches, must be conducted to confirm that β-elemene
directly binds to the ABCB1 protein.
In conclusion, in the present in vitro study,
β-elemene significantly increased the cytotoxicity of paclitaxel,
colchicine and vinblastine in cell lines overexpressing the ABCB1
transporter. β-elemene’s reversal of MDR is most likely due to its
inhibition of the efflux activity of the ABCB1 transporter. The
in vivo effect of β-elemene on the action of paclitaxel
remains to be determined.
Acknowledgements
We thank Dr Michael M. Gottesman (NCI, NIH,
Bethesda, MD, USA) for providing us with KB-3-1 cells, Dr Shin-Ichi
Akiyama (Kagoshima University, Japan) for providing the KB-C2 cell
line and ONO-1078, Drs Susan E. Bates and Robert W. Robey (NIH) for
giving us NCI-H460/MX20 cells and FTC, and Dr Suresh V. Ambudkar
(NIH) for the ABCB1-transfected HEK/ABCB1 and HEK/MRP1 cell lines.
Grant support of the present study was by St. John’s University
Research Seed Grant (no. 579-1110-7002) to Z.S.C, Jie-Ping Wu grant
to H.G.
References
|
1
|
Gottesman MM and Pastan I: Biochemistry of
multidrug resistance mediated by the multidrug transporter. Annu
Rev Biochem. 62:385–427. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Goldstein LJ, Gottesman MM and Pastan I:
Expression of the MDR1 gene in human cancers. Cancer Treat
Res. 57:101–119. 1991.
|
|
3
|
Vasiliou V, Vasiliou K and Nebert DW:
Human ATP-binding cassette (ABC) transporter family. Hum Genomics.
3:281–290. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ambudkar SV, Kimchi-Sarfaty C, Sauna ZE
and Gottesman MM: P-glycoprotein: from genomics to mechanism.
Oncogene. 22:7468–7485. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Perez-Victoria JM, Di Pietro A, Barron D,
Ravelo AG, Castanys S and Gamarro F: Multidrug resistance phenotype
mediated by the P-glycoprotein-like transporter in Leishmania: a
search for reversal agents. Curr Drug Targets. 3:311–333. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sauna ZE, Kim IW and Ambudkar SV: Genomics
and the mechanism of P-glycoprotein (ABCB1). J Bioenerg Biomembr.
39:481–487. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dean M and Annilo T: Evolution of the
ATP-binding cassette (ABC) transporter superfamily in vertebrates.
Annu Rev Genomics Hum Genet. 6:123–142. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Janssen IJ, Krücken J, Demeler J, Basiaga
M, Kornaś S and von Samson-Himmelstjerna G: Genetic variants and
increased expression of Parascaris equorum P-glycoprotein-11
in populations with decreased ivermectin susceptibility. PLoS One.
8:e616352013.PubMed/NCBI
|
|
9
|
Shen DW, Lu YG, Chin KV, Pastan I and
Gottesman MM: Human hepatocellular carcinoma cell lines exhibit
multidrug resistance unrelated to MRD1 gene expression. J
Cell Sci. 98:317–322. 1991.PubMed/NCBI
|
|
10
|
Cordon-Cardo C, O’Brien JP, Boccia J,
Casals D, Bertino JR and Melamed MR: Expression of the multidrug
resistance gene product (P-glycoprotein) in human normal and
tumor tissues. J Histochem Cytochem. 38:1277–1287. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wu CP, Calcagno AM and Ambudkar SV:
Reversal of ABC drug transporter-mediated multidrug resistance in
cancer cells: evaluation of current strategies. Curr Mol Pharmacol.
1:93–105. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Robert J and Jarry C: Multidrug resistance
reversal agents. J Med Chem. 46:4805–4817. 2003. View Article : Google Scholar
|
|
13
|
Tan B, Piwnica-Worms D and Ratner L:
Multidrug resistance transporters and modulation. Curr Opin Oncol.
12:450–458. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dantzig AH, de Alwis DP and Burgess M:
Considerations in the design and development of transport
inhibitors as adjuncts to drug therapy. Adv Drug Deliv Rev.
55:133–150. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tsuruo T, Iida H, Tsukagoshi S and Sakurai
Y: Potentiation of vincristine and adriamycin effects in human
hemopoietic tumor cell lines by calcium antagonists and calmodulin
inhibitors. Cancer Res. 43:2267–2272. 1983.PubMed/NCBI
|
|
16
|
de Souza PS, da Cunha Vasconcelos F, Silva
LF and Maia RC: Cyclosporine A enables vincristine-induced
apoptosis during reversal of multidrug resistance phenotype in
chronic myeloid leukemia cells. Tumour Biol. 33:943–956.
2012.PubMed/NCBI
|
|
17
|
Shukla S, Wu CP and Ambudkar SV:
Development of inhibitors of ATP-binding cassette drug
transporters: present status and challenges. Expert Opin Drug Metab
Toxicol. 4:205–223. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Leonard GD, Polgar O and Bates SE: ABC
transporters and inhibitors: new targets, new agents. Curr Opin
Investig Drugs. 3:1652–1659. 2002.PubMed/NCBI
|
|
19
|
Boesch D, Gavériaux C, Jachez B,
Pourtier-Manzanedo A, Bollinger P and Loor F: In vivo circumvention
of P-glycoprotein-mediated multidrug resistance of tumor cells with
SDZ PSC 833. Cancer Res. 51:4226–4233. 1991.PubMed/NCBI
|
|
20
|
Germann UA, Shlyakhter D, Mason VS, et al:
Cellular and biochemical characterization of VX-710 as a
chemosensitizer: reversal of P-glycoprotein-mediated multidrug
resistance in vitro. Anticancer Drugs. 8:125–140. 1997. View Article : Google Scholar
|
|
21
|
Krishna R and Mayer LD: Multidrug
resistance (MDR) in cancer. Mechanisms, reversal using modulators
of MDR and the role of MDR modulators in influencing the
pharmacokinetics of anticancer drugs. Eur J Pharm Sci. 11:265–283.
2000.PubMed/NCBI
|
|
22
|
Müller H, Pajeva IK, Globisch C and Wiese
M: Functional assay and structure-activity relationships of new
third-generation P-glycoprotein inhibitors. Bioorg Med Chem.
16:2448–2462. 2008.PubMed/NCBI
|
|
23
|
Morschhauser F, Zinzani PL, Burgess M,
Sloots L, Bouafia F and Dumontet C: Phase I/II trial of a
P-glycoprotein inhibitor, Zosuquidar.3HCl trihydrochloride
(LY335979), given orally in combination with the CHOP regimen in
patients with non-Hodgkin’s lymphoma. Leuk Lymphoma. 48:708–715.
2007.PubMed/NCBI
|
|
24
|
van Zuylen L, Sparreboom A, van der Gaast
A, et al: Disposition of docetaxel in the presence of
P-glycoprotein inhibition by intravenous administration of R101933.
Eur J Cancer. 38:1090–1099. 2002.PubMed/NCBI
|
|
25
|
Guns ES, Bullock PL, Reimer ML, Dixon R,
Bally M and Mayer LD: Assessment of the involvement of CYP3A in the
vitro metabolism of a new modulator of MDR in cancer chemotherapy,
OC144–193, by human liver microsomes. Eur J Drug Metab
Pharmacokinet. 26:273–282. 2001.PubMed/NCBI
|
|
26
|
Zhou HY, Shen JK, Hou JS, Qiu YM and Luo
QZ: Experimental study on apoptosis induced by elemene in glioma
cells. Ai Zheng. 22:959–963. 2003.(In Chinese).
|
|
27
|
Zou L, Liu W and Yu L: beta-elemene
induces apoptosis of K562 leukemia cells. Zhonghua Zhong Liu Za
Zhi. 23:196–198. 2001.(In Chinese).
|
|
28
|
Li QQ, Wang G, Zhang M, Cuff CF, Huang L
and Reed E: β-Elemene, a novel plant-derived antineoplastic agent,
increases cisplatin chemosensitivity of lung tumor cells by
triggering apoptosis. Oncol Rep. 22:161–170. 2009.
|
|
29
|
Hu J, Jin W and Yang PM: Reversal of
resistance to adriamycin in human breast cancer cell line MCF-7/ADM
by β-elemene. Zhonghua Zhong Liu Za Zhi. 26:268–270. 2004.(In
Chinese).
|
|
30
|
Biedler JL and Riehm H: Cellular
resistance to actinomycin D in Chinese hamster cells in vitro:
cross-resistance, radioautographic, and cytogenetic studies. Cancer
Res. 30:1174–1184. 1970.PubMed/NCBI
|
|
31
|
Juliano RL and Ling V: A surface
glycoprotein modulating drug permeability in Chinese hamster ovary
cell mutants. Biochim Biophys Acta. 455:152–162. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Akiyama S, Fojo A, Hanover JA, Pastan I
and Gottesman MM: Isolation and genetic characterization of human
KB cell lines resistant to multiple drugs. Somat Cell Mol Genet.
11:117–126. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Carmichael J, DeGraff WG, Gazdar AF, Minna
JD and Mitchell JB: Evaluation of a tetrazolium-based semiautomated
colorimetric assay: assessment of radiosensitivity. Cancer Res.
47:943–946. 1987.PubMed/NCBI
|
|
34
|
Sodani K, Tiwari AK, Singh S, et al:
GW583340 and GW2974, human EGFR and HER-2 inhibitors, reverse
ABCG2- and ABCB1-mediated drug resistance. Biochem Pharmacol.
83:1613–1622. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tiwari AK, Sodani K, Dai CL, et al:
Nilotinib potentiates anticancer drug sensitivity in murine ABCB1-,
ABCG2-, and ABCC10-multidrug resistance xenograft models. Cancer
Lett. 328:307–317. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sun YL, Chen JJ, Kumar P, et al: Reversal
of MRP7 (ABCC10)-mediated multidrug resistance by tariquidar. PLoS
One. 8:e555762013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Aller SG, Yu J, Ward A, et al: Structure
of P-glycoprotein reveals a molecular basis for poly-specific drug
binding. Science. 323:1718–1722. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Shi Z, Tiwari AK, Shukla S, et al:
Sildenafil reverses ABCB1- and ABCG2-mediated chemotherapeutic drug
resistance. Cancer Res. 71:3029–3041. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Dai CL, Tiwari AK, Wu CP, et al: Lapatinib
(Tykerb, GW572016) reverses multidrug resistance in cancer cells by
inhibiting the activity of ATP-binding cassette subfamily B member
1 and G member 2. Cancer Res. 68:7905–7914. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ding PR, Tiwari AK, Ohnuma S, et al: The
phosphodiesterase-5 inhibitor vardenafil is a potent inhibitor of
ABCB1/P-glycoprotein transporter. PLoS One. 6:e193292011.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Huang XC, Sun YL, Salim AA, Chen ZS and
Capon RJ: Parguerenes: marine red alga bromoditerpenes as
inhibitors of P-glycoprotein (ABCB1) in multidrug resistant human
cancer cells. Biochem Pharmacol. 85:1257–1268. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Patel A, Tiwari AK, Chufan EE, et al:
PD173074, a selective FGFR inhibitor, reverses ABCB1-mediated drug
resistance in cancer cells. Cancer Chemother Pharmacol. 72:189–199.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Xu HB, Li L, Fu J, Mao XP and Xu LZ:
Reversion of multidrug resistance in a chemoresistant human breast
cancer cell line by β-elemene. Pharmacology. 89:303–312.
2012.PubMed/NCBI
|