Introduction
Interferon-λ (IFN-λ), type III IFNs, the newest IFN
family, includes 3 subtypes, IFN-λ1, -λ2 and -λ3, also known as
interleukin-29 (IL-29), IL-28A and IL-28B. All of the IFN-λs bind
the same heterodimeric receptor complexes composed of the IL-10
receptor β (IL-10Rβ) and a novel IL-28 receptor α (IL-28Rα)
(1,2). Interferons (IFNs) have been described
as agents mediating antiviral responses for many years. Type I
IFNs, IFN-α/β, have been clinically applied for treating certain
types of malignancies for more than 30 years, yet a wide range of
adverse effects hampered further clinical applications (3,4).
However, further investigations are required to clarify the
biological properties and the mechanisms responsible for their
functions. Type III IFNs, IFN-λs, have similar signaling pathways
as IFN-α/β and inhibit the proliferation of tumor cells through
cell cycle arrest or apoptosis.
Gastric carcinoma is an intractable disease, with a
high incidence and mortality rate, as it frequently develops in the
aged (5–7). Extensive surgery decreases the quality
of life of patients, and radical chemotherapy and radiotherapy are
not often appropriate for aged patients (8). A novel therapeutic strategy is thereby
required to improve prognosis. More effective, preventive and
therapeutic approaches are urgently needed. Type I IFNs have been
clinically examined in various types of malignanies including
gastric carcinoma. Yet, type I IFNs are associated with numerous
serious adverse effects, e.g. major depression. Type III IFNs have
various immunological actions similar to type I IFNs. However,
IFN-λ-mediated growth suppression has not yet been well
explored.
Research has shown that the IFN-λs may have unique
cellular functions distinct from their role as secreted cytokines
(9). Yan et al found that
the IFN-λs could inhibit lung carcinoma cell growth and produce
antitumor effects (10). The same
observation was also made by Steen et al that IFN-λs could
sharply inhibit the proliferation of gastric carcinoma cells
(11). These results indicate that
IFN-λs may be a new therapeutic option for tumors. However, whether
IFN-λs inhibit the growth of gastric carcinoma cells remains
unknown.
In the present study, we examined the potential
antitumor action of IFN-λ1 in human gastric carcinoma cell lines
and the possible interaction between IFN-λ1 and human gastric
carcinoma cells. The results indicated that IFN-λ1 plays a crucial
role in inhibiting gastric carcinoma cells and provide the
foundation for the use of IFN-λ1 in tumor therapy.
Materials and methods
Antibodies and reagents
Mouse anti-human monoclonal primary antibodies for
β-actin, Bax, cytochrome c, apoptosis-inducing factor (AIF),
p21, p27 and horseradish peroxidase-conjugated goat anti-mouse IgG
antibodies were purchased from Cell Signaling (Beverly, MA, USA).
RPMI-1640 and fetal bovine serum (FBS) were purchased from Gibco
(Invitrogen, USA). Propidium iodide (PI) was purchased from
Sigma-Aldrich (St. Louis, MO, USA). IFN-λ1 was purchased from
R&D Systems (Minneapolis, MN, USA). Viable cells were measured
with a cell proliferation assay kit
[3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide; MTT]
(Roche, USA). The Annexin V-FITC/PI apoptosis kit was purchased
from Biosea Biotechnology Co. Ltd. (Beijing, China). All other
reagents were purchased from Sigma-Aldrich unless otherwise
mentioned.
Cell culture
Human gastric carcinoma HGC-27 and SGC-7901 cells
were obtained from the Chinese Type Culture Collection (Institute
of Cell Biology, Chinese Academy of Science, Shanghai, China) and
cultured in RPMI-1640 medium containing 10% FBS, 100 IU/ml
penicillin and 100 mg/ml streptomycin at 37°C in a humidified
atmosphere of 5% CO2 and 95% air. After reaching 80–90%
confluency, cells were collected by 0.25% trypsin detachment and
centrifugation and then replanted into 96- or 6-well plates at an
appropriate density according to the different experimental
requirements. After a 48-h culture, all experiments were
performed.
Cell viability assay
Cell viability was evaluated using the MTT assay.
Cells were seeded in a 96-well plate at a density of
5×103 cells/well. The total volume was adjusted to 200
μl with growth medium. At 24 h after the seeding, cells were
treated with different concentrations of IFN-λ1 (0, 10, 100, 1000
ng/ml). Cell viability was examined after 48 h using a standard MTT
method. Drug effect was expressed as a percentage relative to the
controls. The rate of cell viability was calculated as follows:
(absorbance of treated group - absorbance of blanks)/(absorbance of
control - absorbance of blanks) × 100 expressed as a
percentage.
Flow cytometry and cell cycle
analysis
The apoptotic cells were detected using the Annexin
V-FITC apoptosis kit. Briefly, the treated cells were collected by
0.25% trypsin and centrifuged, then washed with ice-cold phosphate
buffer solution and resuspended in 200 μl binding buffer at a
density of 1×106/ml. Ten microliters of Annexin V-FITC
antibody was added, and cells were incubated for 30 min at room
temperature and supplemental 300 μl of binding buffer was added and
cells were stained with 5 μl PI for 5 min. The apoptotic cells were
measured by flow cytometry (FACScan, Becton-Dickinson, Mountain
View, CA, USA). For synchronization at G1 phase, cells were
cultured with IFN-λ1 and fixed in ice-cold 70% ethanol, treated
with RNase (50 μg/ml) and stained with PI (50 μg/ml). Cell cycle
distribution was analyzed with FACScan and Cell Quest software.
Detection of mitochondrial membrane
potential (ΔΨm)
The mitochondrial membrane potential ΔΨm was
determined using the KeyGEN mitochondrial membrane JC-1 kit
(KeyGEN, China) according to the manufacturer’s instructions
(12). Briefly, after cells were
infected with viral particles for the indicated times, the cells
were pelleted, washed with PBS and resuspended in 0.5 ml of diluted
JC-1 reagent (1 μmol/ml in incubation buffer) for 20 min. Then 0.2
ml of incubation buffer was added, and the cells were centrifuged,
washed and resuspended in 1 ml PBS for laser scanning confocal
microscopy (Olympus, Japan) and flow cytometry.
Western blot analysis
Cells were treated with IFN-λ1 for 48 h and washed
with phosphate-buffered saline and then resuspended in RIPA buffer
(25 mM Tris-HCl, pH 7.6, 150 mM NaCl, 1% Nonidet P-40, 1% sodium
deoxycholate, 0.1% SDS) with a protease inhibitor cocktail
(Sigma-Aldrich). SDS-PAGE was performed with 12% polyacrylamide
gels, and portions (25 μg) of protein of the cell lysate were used.
After electrotransfer, membranes were blocked for 1 h at room
temperature with 6% Quick-Blocker (Chemicon, Temecula, CA, USA)
dissolved in TBST [20 mM Tris-HCl, 137 mM NaCl, 0.1% (v/v)
Tween-20, pH 7.6], then incubated overnight at 4°C with the
different primary antibodies at the appropriate dilution in TBS-T
buffer. The following monoclonal antibodies were used: anti-p21
antibody (Ab), anti-p27 Ab, anti-Bax Ab, anti-Bcl-2 Ab,
anti-caspase-3 Ab, anti-caspase-9 Ab, anti-cytochrome c Ab
and anti-apoptosis-inducing factor (AIF) Ab (Cell Signaling,
Beverly, MA, USA), followed by incubation with a horseradish
peroxidase-conjugated secondary antibody. The membranes were
developed with the ECL system (GE Healthcare, Little Chalfont, UK)
and exposed to X-ray film. The anti-β-actin antibody was used as an
internal control for sample loading.
Statistical analysis
The results from the in vivo studies are
representative of at least 2–3 independent experiments and are
expressed as mean ± SD. The differences between the groups were
examined using one-way analysis of variance (ANOVA) using the
Statistical Package for the Social Sciences (SPSS) 14.0 software
(SPSS Inc., Chicago, IL, USA). Statistical significance was assumed
at P<0.05 in all cases.
Results
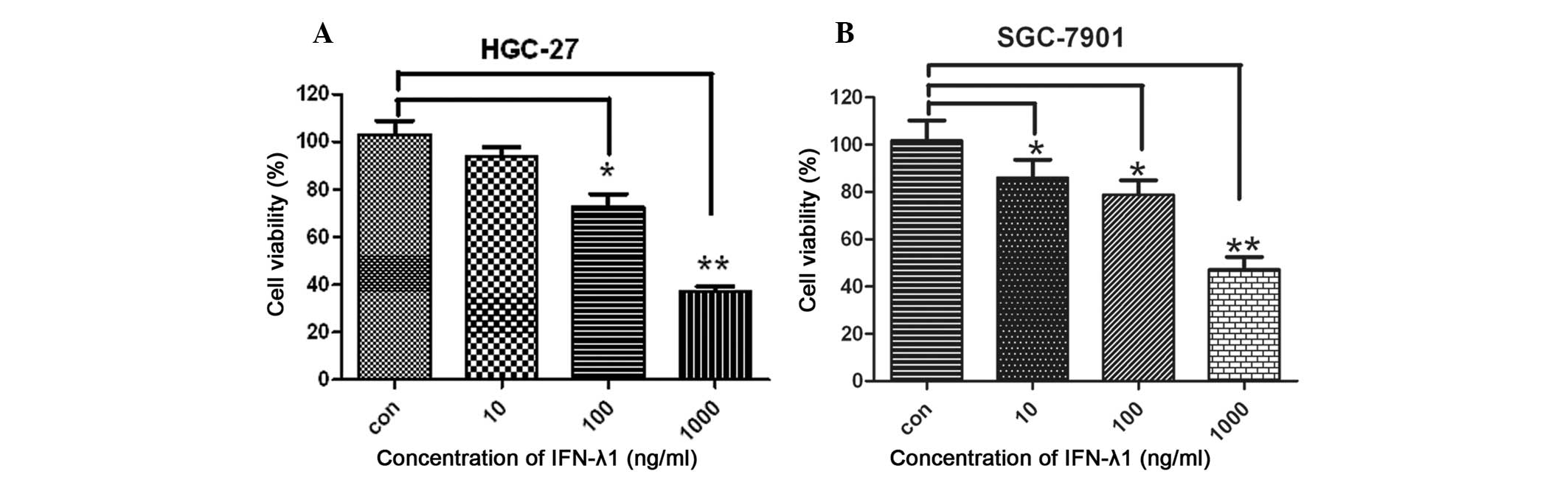
IFN-λ1-mediated growth suppression
We examined whether IFN-λ1 could suppress the growth
of the gastric carcinoma cells. We treated the HGC-27 and SGC-7901
cells with IFN-λ1 and examined the viability and the cell
proliferation rates. Compared with the control group, IFN-λ1
treatment significantly inhibited the proliferation of the gastric
carcinoma cells in a concentration-dependent manner (Fig. 1).
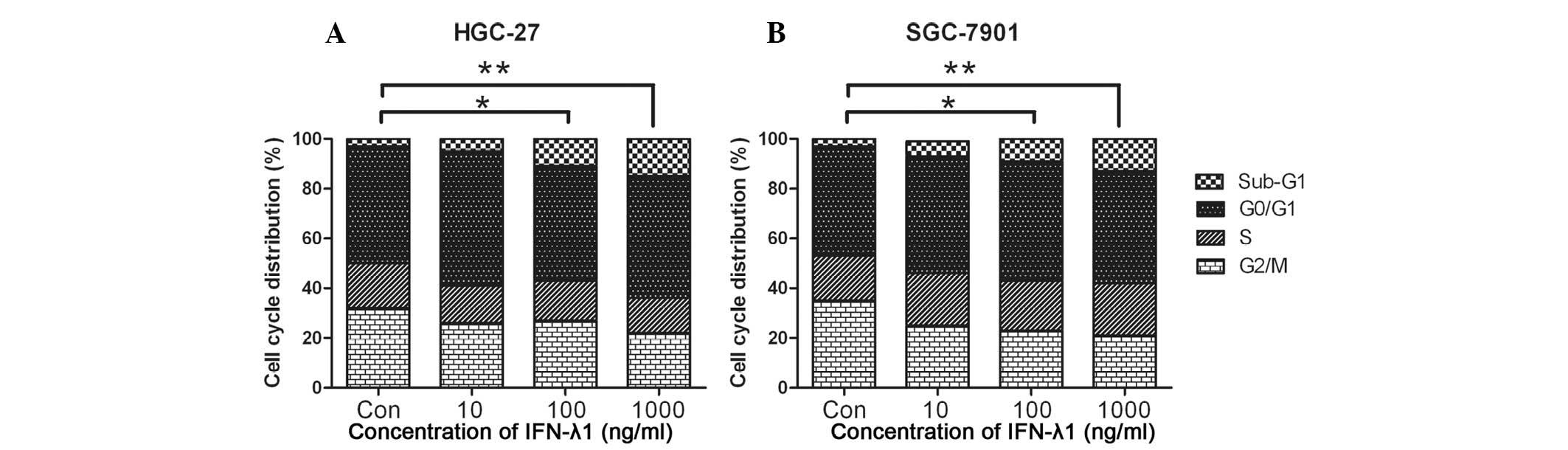
G1 phase arrest induced by IFN-λ1
To explain the mechanism of cell growth inhibition
by IFN-λ1 concerning cell cycle change, we analyzed the effects of
IFN-λ1 treatment on the cell cycle. As shown in Table I, the percentage of cells in the G1
phase increased, whereas the percentage of cells in the G2 and S
phases decreased (Fig. 2). These
results demonstrated that IFN-λ1 induced G1 phase arrest in the
gastric carcinoma cells.
 | Table ICell cycle distribution after
Ad/IFN-λ1 treatment. |
Table I
Cell cycle distribution after
Ad/IFN-λ1 treatment.
| | Cell cycle
distribution (%) |
|---|
| |
|
|---|
| Cells | Treatment
(ng/ml) | Sub-G1 | G0/G1 | S | G2/M |
|---|
| HGC-27 | Control | 2.7±0.7 | 47.5±1.4 | 17.6±0.7 | 33.7±0.9 |
| 10 | 4.9±0.9 | 53.7±1.2a | 15.2±0.3 | 28.1±0.8 |
| 100 | 11.0±0.3a | 46.1±0.8 | 15.9±0.2 | 27.1±1.2 |
| 1000 | 15.3±0.7b | 49.2±0.4 | 14.7±0.1 | 23.2±0.9 |
| SGC-7901 | Control | 3.1±0.8 | 44.0±0.3 | 18.3±0.6 | 34.6±0.6 |
| 10 | 5.6±0.6 | 47.5±0.9 | 20.9±1.4 | 26.5±0.7 |
| 100 | 8.9±0.9a | 48.8±0.6 | 20.1±0.2 | 22.9±1.2 |
| 1000 | 13.4±0.4b | 45.1±0.7 | 21.3±1.0 | 22.1±0.8 |
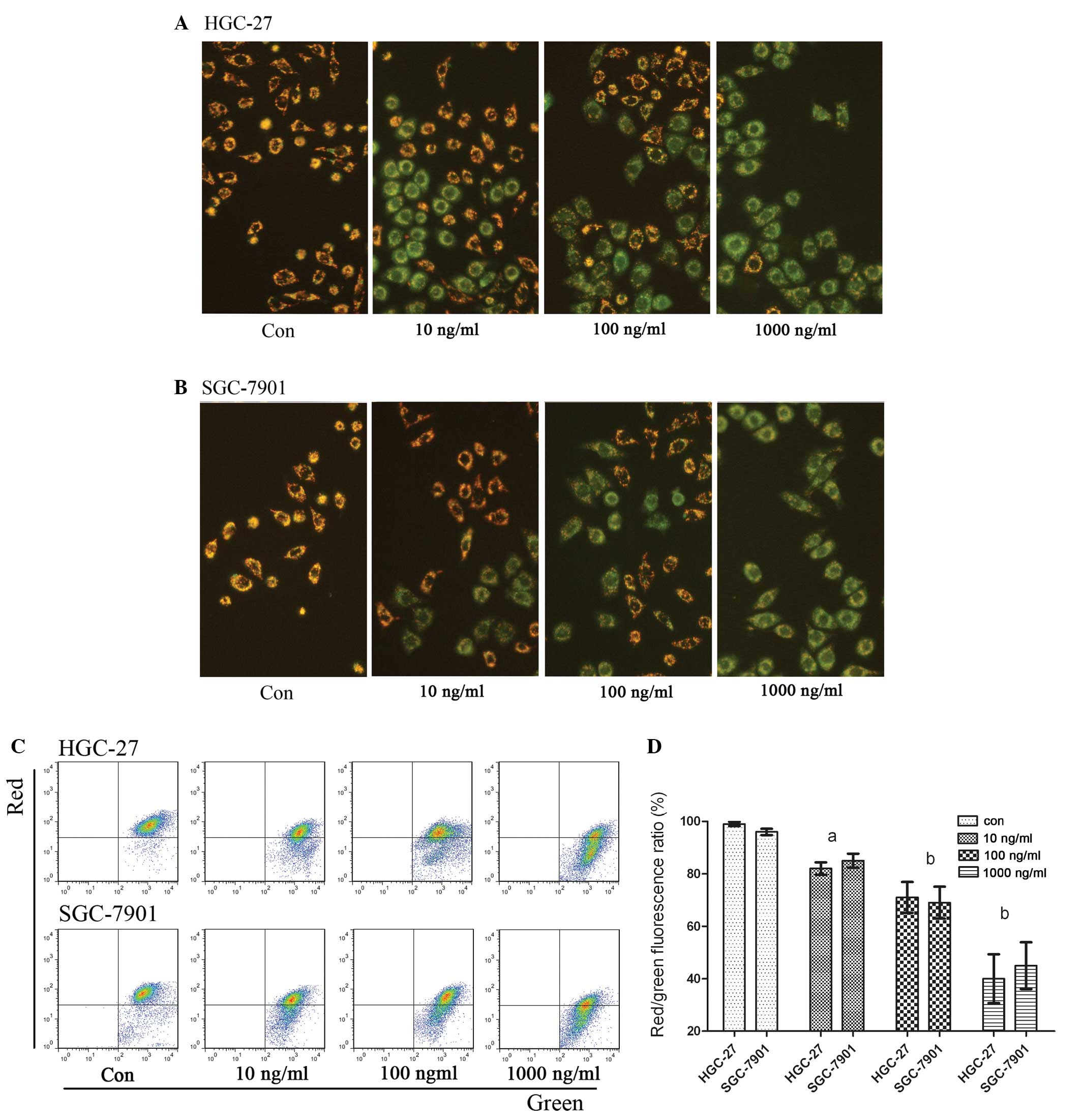
Mitochondrial membrane potential ΔΨm
To confirm the effects of IFN-λ1 on gastric
carcinoma cell mitochondria in vitro, gastric carcinoma
cells were treated with different concentrations of IFN-λ1. Changes
in mitochondrial membrane potential of the cells were detected
using laser scanning confocal microscopy (Fig. 3A and 3B) and flow cytometry
(Fig. 3C). It was obvious that the
mitochondrial membrane potential was decreased after treatment with
IFN-λ1 (Fig. 3D). Based on the
results above, we hypothesized that IFN-λ1 may induce apoptosis
through pathways that downregulate mitochondrial membrane
potential.
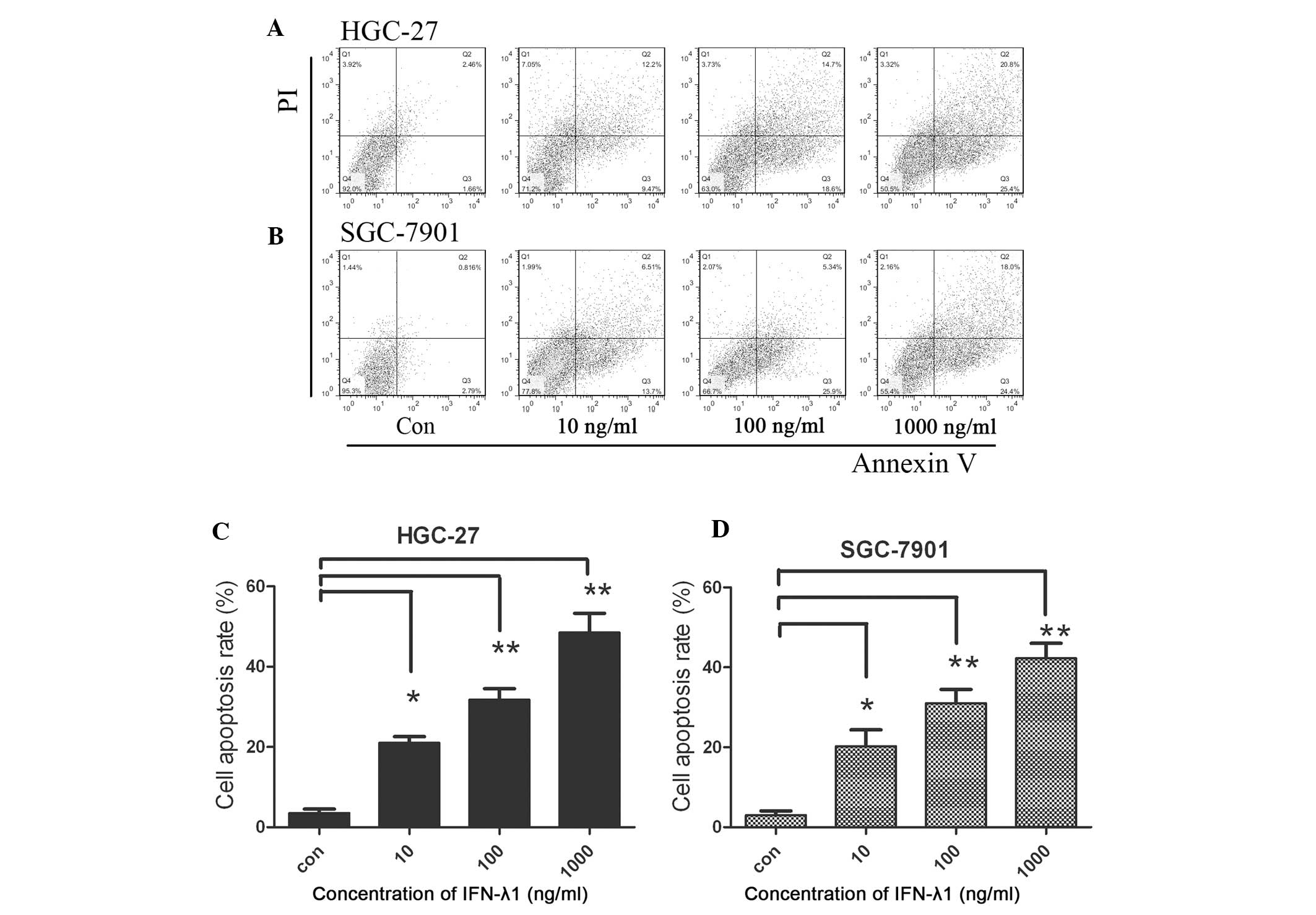
Apoptosis induction by IFN-λ1
treatment
Apoptosis was confirmed by Annexin V and PI
staining. As shown in Fig 4, the
Annexin V+/PI fraction and subsequently the Annexin
V+/PI+ fraction of the gastric carcinoma
cells were increased following IFN-λ1 treatment. These data
collectively suggest an apoptotic effect of IFN-λ1 in gastric
carcinoma cells.
Expression of proteins involved in cell
cycle progression and apoptosis
To understand the mechanism by which IFN-λ1 caused
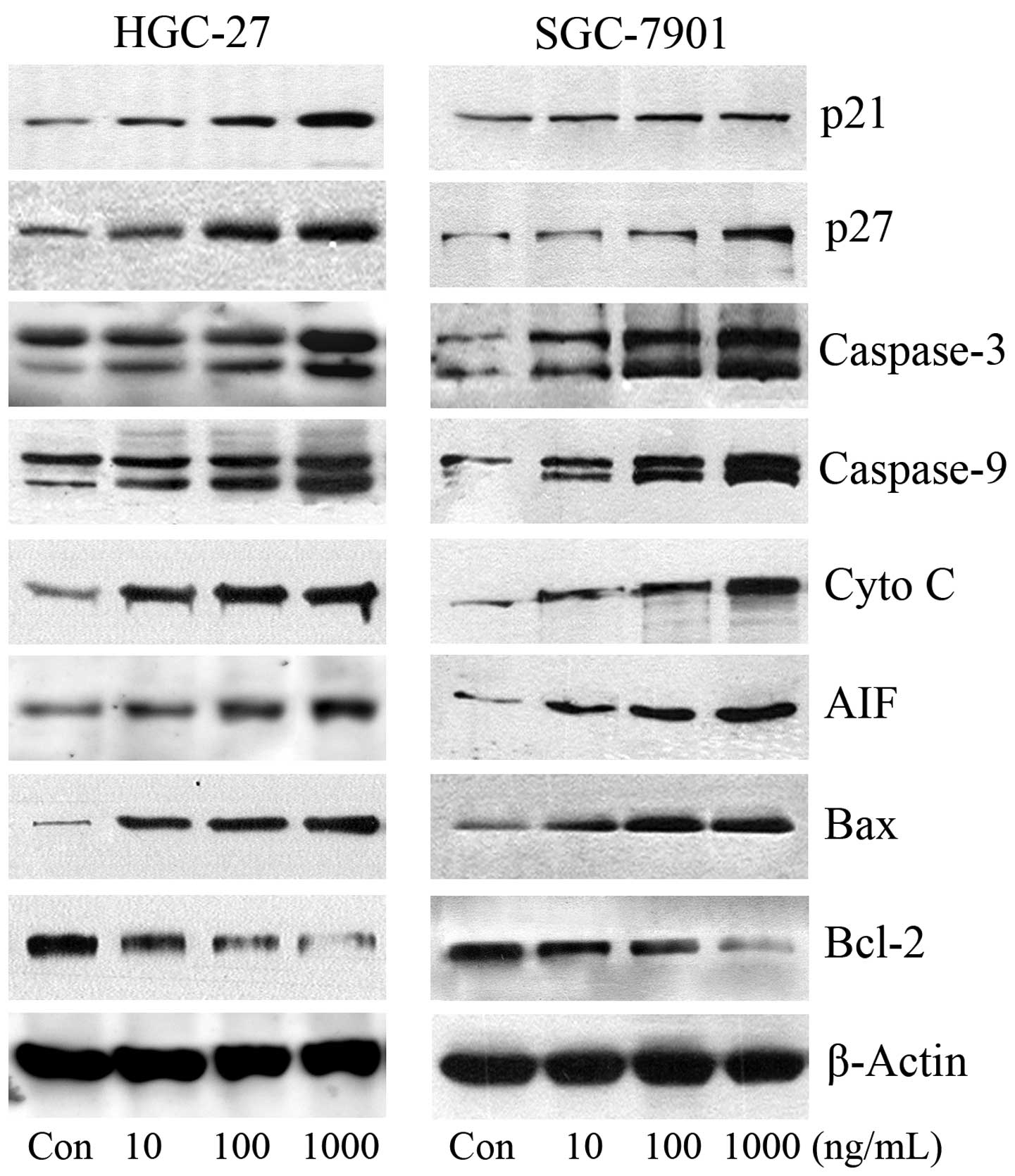
G1-phase cell cycle arrest, we examined the effects of IFN-λ1 on
the induction of p21 and p27, which regulate the G1-phase
checkpoint (13–15). IFN-λ1 treatment induced p21 and p27
expression in a concentration-dependent manner (P<0.05)
(Fig. 5), suggesting that IFN-λ1
may regulate cell cycle arrest through p21 and p27.
Disruption of the mitochondrial membrane potential
is a critical event in triggering mitochondrial-mediated apoptosis.
Mitochondrial dysfunction is induced by various cellular stimuli,
causing the translocation of Bax from the cytosol to the
mitochondria, which subsequently triggers the release of cytochrome
c and AIF from the mitochondria into the cytosol and finally
induces apoptosis via either caspase-dependent or
caspase-independent pathways. To further clarify the mechanisms of
apoptosis in the gastric carcinoma cells triggered by IFN-λ1, we
targeted apoptotic proteins in the mitochondrial pathway. IFN-λ1
significantly increased Bax expression in the mitochondria
(Fig. 5). IFN-λ1 also
concentration-dependently upregulated the expression of cytochrome
c and AIF proteins (P<0.05).
Discussion
In recent years, increasing research has focused on
the relationship between cytokines and tumorigenesis. It has been
suggested that cytokines may be a new therapeutic option for tumors
(16–20). In our experiments, we demonstrated
that IFN-λ1 inhibited the growth of gastric carcinoma cells in a
concentration-dependent manner. These data suggest that IFN-λ1 may
be a potential antitumor agent for the treatment of gastric
cancer.
Impaired apoptotic induction and dysregulated cell
cycle progression are important factors in cancer development.
Accordingly, inhibition of cell cycle regulation is particularly
useful in the treatment of cancer. In our in vitro study, we
demonstrated the apoptosis-inducing effects of IFN-λ1 in gastric
carcinoma cells using PI cell cycle analysis, Annexin V and PI
staining as well as activated caspase-3. Our study showed that
IFN-λ1 induced G1 phase arrest and apoptosis in the gastric
carcinoma cells.
Experimental evidence suggests that apoptosis can be
mediated by several different pathways and that there are numerous
regulatory molecules associated with these pathways. Proteins of
the Bcl-2 family include both pro- and anti-apoptotic members that
elicit opposing effects on mitochondria (21–23)
Enhancement of pro-apoptotic Bax over Bcl-2 proteins can enhance
the permeability of the mitochondrial membrane, which in turn
results in the release of apoptogenic factors. Repression of
anti-apoptotic members of this family, including Bcl-2 and Bcl-xl,
preserves the integrity of the mitochondria. This blocks the
release of soluble inter-membrane factors such as cytochrome
c that activate effectors of apoptosis. In the present
study, we found that treatment of gastric carcinoma cells with
IFN-λ1 upregulated pro-apoptotic Bax protein and downregulated
anti-apoptotic protein Bcl-2.
The alteration in the Bax/Bcl-2 ratio is critical
for apoptosis and causes the release of cytochrome c from
mitochondria into the cytosol after loss of mitochondrial membrane
potential. Cytosolic cytochrome c can bind to Apaf-1 and
activate caspase-9 in the apoptosomes in response to diverse
inducers of cell death. Caspase-9 is activated and finally an
increase in the activation or cleavage of caspase-3 results
(24,25). It has been recognized that control
of cell cycle progression in cancer cells is an effective strategy
to inhibit tumor growth as the molecular analyses of human cancers
have revealed that cell cycle regulators are frequently deregulated
in most common malignancies. Our in vitro data demonstrated
that treatment of gastric carcinoma cells with IFN-λ1 induced G1
phase arrest of cell cycle progression, indicating that one of the
mechanisms by which IFN-λ1 inhibits the proliferation of gastric
cancer cells is inhibition of cell cycle progression.
Regulation of the cell cycle at the G1 checkpoint is
complex and involves multiple molecular processes. p21 appears to
be important since introduction of nonfunctional p21 or a p21
antisense oligonucleotide diminished the G1 arrest in a variety of
cancer cells (26,27). Apart from directly inhibiting
cyclin-dependent kinase activity to promote cell cycle arrest, p21
interacts with proliferating cell nuclear antigens. Our results
demonstrated elevated p21 and p27 levels in gastric carcinoma cells
in response to IFN-λ1. It is possible that G1 arrest upon IFN-λ1
treatment here was caused by p21 and p27. Exposure to IFN-λ1
elevated the Bax level in the mitochondria, and stimulated the
release of cytochrome c and AIF from the mitochondria into
the cytosol, suggesting that IFN-λ1-induced apoptosis in gastric
carcinoma cells is mediated by the mitochondrial pathway.
In conclusion, we demonstrated that IFN-λ1 induced
G1 phase arrest and apoptotic cell death in gastric carcinoma cells
through the activation of p21/p27 and the mitochondrial-mediated
apoptotic pathway. In the present study, although the exact
mechanism remains unclear, IFN-λ1 as a chemotherapeutic agent
induces cell death and may be a potential therapeutic option for
gastric cancers.
References
|
1
|
Lopušná K, Režuchová I, Betáková T, et al:
Interferons lambda, new cytokines with antiviral activity. Acta
Virol. 57:171–179. 2013.PubMed/NCBI
|
|
2
|
Tagawa M, Kawamura K, Li Q, et al: A
possible anticancer agent, type III interferon, activates cell
death pathways and produces antitumor effects. Clin Dev Immunol.
2011:4790132011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li Q, Kawamura K, Okamoto S, et al:
Adenoviruses-mediated transduction of human oesophageal carcinoma
cells with the interferon-λ genes produced anti-tumour effects. Br
J Cancer. 105:1302–1312. 2011.PubMed/NCBI
|
|
4
|
Donnelly RP and Kotenko SV:
Interferon-lambda: a new addition to an old family. J Interferon
Cytokine Res. 30:555–564. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tamura W and Fukami N: Early gastric
cancer and dysplasia. Gastrointest Endosc Clin N Am. 23:77–94.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hu Y, Fang JY and Xiao SD: Can the
incidence of gastric cancer be reduced in the new century? J Dig
Dis. 14:11–15. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Iravani O, Tay BW, Chua PJ, et al:
Claudins and gastric carcinogenesis. Exp Biol Med. 238:344–349.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Saito H, Takaya S, Fukumoto Y, et al:
Clinicopathologic characteristics and prognosis of gastric cancer
in young patients. Yonago Acta Med. 55:57–61. 2012.PubMed/NCBI
|
|
9
|
Wongthida P, Diaz RM, Galivo F, et al:
Type III IFN interleukin-28 mediates the antitumor efficacy of
oncolytic virus VSV in immune-competent mouse models of cancer.
Cancer Res. 70:4539–4549. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yan Y, Zhang J, Liu Y, et al: Inhibition
of lung adenocarcinoma transfected with interleukin 28A recombinant
adenovirus (Ad- mIFN-λ2) in vivo. Cancer Biother Radiopharm.
28:124–130. 2013.PubMed/NCBI
|
|
11
|
Steen HC and Gamero AM: Interferon-lambda
as a potential therapeutic agent in cancer treatment. J Interferon
Cytokine Res. 30:597–602. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ding F, Shao ZW, Yang SH, et al: Role of
mitochondrial pathway in compression-induced apoptosis of nucleus
pulposus cells. Apoptosis. 17:579–590. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mitrea DM, Yoon MK, Ou L, et al:
Disorder-function relationships for the cell cycle regulatory
proteins p21 and p27. Biol Chem. 393:259–274. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang L, Wang G, Yang D, et al: Euphol
arrests breast cancer cells at the G1 phase through the modulation
of cyclin D1, p21 and p27 expression. Mol Med Rep. 8:1279–1285.
2013.PubMed/NCBI
|
|
15
|
Zhang Z, Du GJ, Wang CZ, et al: Compound
K, a ginsenoside metabolite, inhibits colon cancer growth via
multiple pathways encluding p53-p21 interactions. Int J Mol Sci.
14:2980–2995. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Abushahba W, Balan M, Castaneda I, et al:
Antitumor activity of type I and type III interferons in BNL
hepatoma model. Cancer Immunol Immunother. 59:1059–1071. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang L, Wei J and He S: Integrative
genomic analyses on interferon-lambdas and their roles in cancer
prediction. Int J Mol Med. 25:299–304. 2010.PubMed/NCBI
|
|
18
|
Li Q, Kawamura K, Ma G, et al:
Interferon-lambda induces G1 phase arrest or apoptosis in
oesophageal carcinoma cells and produces anti-tumour effects in
combination with anti-cancer agents. Eur J Cancer. 46:180–190.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lasfar A, Abushahba W, Balan M, et al:
Interferon lambda: a new sword in cancer immunotherapy. Clin Dev
Immunol. 2011:3495752011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
George PM, Badiger R, Alazawi W, et al:
Pharmacology and therapeutic potential of interferons. Pharmacol
Ther. 135:44–53. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Renault TT, Teijido O, Antonsson B, et al:
Regulation of Bax mitochondrial localization by Bcl-2 and Bcl-x
(L): keep your friends close but your enemies closer. Int J Biochem
Cell Biol. 45:64–67. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Low IC, Kang J and Pervaiz S: Bcl-2: a
prime regulator of mitochondrial redox metabolism in cancer cells.
Antioxid Redox Signal. 15:2975–2987. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shamas-Din A, Kale J, Leber B, et al:
Mechanisms of action of Bcl-2 family proteins. Cold Spring Harb
Perspect Biol. 5:a0087142013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Imesch P, Scheiner D, Szabo E, et al:
Conjugates of cytochrome c and antennapedia peptide
activates apoptosis and inhibit proliferation of HeLa cancer cells.
Exp Ther Med. 6:786–790. 2013.
|
|
25
|
Liang J, Yu Y, Wang B, et al: Ginsenoside
Rb1 attenuates oxygen-glucose deprivation-induced apoptosis in
SH-SY5Y cells via protection of mitochondria and inhibition of AIF
and cytochrome c release. Molecules. 18:12777–12792. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Park C, Jeong NY, Kim GY, et al:
Momilactone B induces apoptosis and G1 arrest of the cell cycle in
human monocytic leukemia U937 cells through downregulation of pRB
phosphorylation and induction of the cyclin-dependent kinase
inhibitor p21Waf1/Cip1. Oncol Rep. 31:1653–1660. 2014.
|
|
27
|
Koyama M, Sowa Y, Hitomi T, et al:
Perillyl alcohol causes G1 arrest through p15(INK4b) and
p21(WAF1/Cip1) induction. Oncol Rep. 29:779–784. 2013.PubMed/NCBI
|