Introduction
Liver cancer remains the fifth most common cancer in
men and the seventh in women worldwide (1). The curative effect of current
treatment for liver cancer is less than satisfactory. There is a
great need to develop new approaches for liver cancer treatment. In
China, Traditional Chinese Medicine (TCM) has long been used for
liver cancer treatment, and has been confirmed to effectively
control cancer progression, improve the quality of life, and
prolong survival times to some extent in liver cancer patients
(2–5). Based on different syndrome patterns,
TCM therapeutic principles, such as invigorating spleen and
regulating Qi, clearing heat-dampness, dissipating stasis,
softening hardness, or tonifying liver and kidney can be used to
treat cancer (5–8).
The fruit of Ligustrum lucidum Ait.
(Nü-zhen-zi in Chinese) is one of the most frequently used
liver/kidney Yin tonifying herbs in liver cancer patients for
clinical syndrome amelioration (9).
As a tonic Chinese herb, Ligustrum lucidum fruit has been
confirmed to be effective in improving chemotherapy-induced
myelosuppression, alopecia and immunosuppression (10–12).
Ligustrum lucidum fruit has also been used to enhance the
therapeutic effects of chemotherapy in TCM clinical practice.
Ligustrum lucidum fruit has been demonstrated to display
antiproliferative potential against lung and pancreatic carcinoma,
breast and prostate adenocarcinoma, glioma and colorectal carcinoma
(13–15). However, the effect of Ligustrum
lucidum fruit on liver cancer cells remains unknown.
The present study aimed to evaluate the effect of an
aqueous extract of Ligustrum lucidum fruit on
hepatocarcinoma cells. We observed that Ligustrum lucidum
fruit extract (LLFE) induced apoptosis in human hepatocellular
carcinoma Bel-7402 cells through activation of caspases. LLFE also
induced cell senescence accompanied by upregulation of p21 and
downregulation of RB phosphorylation. In addition, silencing of p21
by RNA interference partially abrogated LLFE-induced apoptosis, and
significantly abrogated LLFE-induced cell senescence.
Materials and methods
Chemicals and reagents
The Cell Counting Kit-8 (CCK-8) was obtained from
Dojindo (Kumamoto, Japan). Colorimetric CaspACE™ Assay System was
the product of Promega (Madison, WI, USA). Z-VAD-FMK, Caspase-8 and
Caspase-9 Colorimetric Assay kits were purchased from R&D
Systems (Minneapolis, MN, USA). Antibodies against p53, p16, p21,
RB, pRB and β-actin, and the Senescence β-Gal staining kit were the
products of Cell Signaling Technology (Danvers, MA, USA). Small
interfering RNA (siRNA) against p21 and control siRNA were procured
from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Lipofectamine™
2000 was from Invitrogen (Carlsbad, CA, USA).
Extraction of Ligustrum lucidum
fruit
An aqueous extract of Ligustrum lucidum fruit
was prepared as a lyophilized-dry powder as previously described
(16,17). Authentic Ligustrum lucidum
fruit herb material was obtained from Longhua Hospital.
Ligustrum lucidum fruit was soaked for 1 h, and decocted
twice with an 8-fold volume of boiling distilled water for 2 h. The
decoction was filtered and centrifuged twice at 12,000 rpm for 30
min to remove the insoluble ingredients. The supernatants were
mixed with an equal volume of ethanol and kept at 4°C overnight,
and centrifuged at 12,000 rpm for 30 min to remove the insoluble
ingredients. The resultant supernatants were lyophilized, weighed,
dissolved in RPMI-1640 medium and adjusted to a concentration of
400 mg/ml, and were sequentially passed through 0.45- and 0.22-μm
filters and sterilized.
Cell culture
Human hepatocellular carcinoma Bel-7402 cells and
human hepatocyte HL-7702 cells were obtained from the Cell Bank of
the Type Culture Collection of the Chinese Academy of Sciences.
Bel-7402 and HL-7702 cells were grown in RPMI-1640 medium with 10%
FBS and 1% pen-strep, and maintained at 37°C in a humidified
incubator with a 5% CO2 atmosphere.
Cell proliferation assay
Cells in logarithmic growth phase were seeded into a
96-well plate (4×103 cells/well) and allowed to attach
for 24 h before treatment. The cells were exposed to various doses
of LLFE for 72 h, and cell viability was evaluated every 24 h by
using the CCK-8 colorimetric assay according to the manufacturer’s
instructions. The cell survival rate was calculated as follows:
Cell survival rate (%) = experimental OD value/control OD value ×
100%.
Flow cytometric analysis
LLFE-treated Bel-7402 cells were collected, stained
with Annexin V-FITC and PI as recommended by the manufacturer, and
detected using a FACSCalibur flow cytometer (Becton-Dickinson). For
cell cycle analysis, LLFE-treated Bel-7402 cells were stained with
PI (50 μg/ml) and analyzed using a FACSCalibur flow cytometer.
Caspase activity assay
After treatment with different concentrations of
LLFE, caspase-3, -8 and -9 activities were measured by the cleavage
of the specific chromogenic substrate according to the
manufacturer’s instructions. For caspase inhibition, cells
pretreated with Z-VAD-FMK (50 μmol/l, 2 h) were incubated with LLFE
for another 72 h.
Senescence-activated β-galactosidase
staining
Bel-7402 cells (3×104) were plated in
35-mm-diameter plates and treated with different doses of LLFE for
5 days. Senescence-activated expression of β-galactosidase activity
(18) was detected by the
Senescence β-Gal staining kit according to the manufacturer’s
protocol, and observed under a microscope.
Western blotting
Western blot analyses were performed as previously
described (16,17). Briefly, collected cells were lysed
and subjected to 8–12% SDS-PAGE gel, and transferred onto a
nitrocellulose membrane (Amersham Biosciences, Buckinghamshire,
UK). The transferred membranes were blocked with 5% non-fat milk,
washed, and probed with the indicated antibodies. Blots were then
washed and incubated with IRDye 700- and IRDye 800-conjugated
secondary antibodies (Rockland Immunochemicals, Gilbertsville, PA,
USA), and visualized in the Odyssey Infrared Imaging System (LI-COR
Biosciences, Lincoln, NE, USA).
siRNA treatment
For siRNA transfection, Bel-7402 cells were cultured
on a 6-well plate to 60% confluency, and 80 pmol of specific or
non-specific control siRNA was introduced into the cells using
Lipofectamine™ 2000 according to the manufacturer’s
recommendations. After 24 h of transfection, cells were treated
with 200 μg/ml of LLFE or the same volume of RPMI-1640, and
harvested for apoptosis assay or senescence β-Gal staining.
Statistical analyses
Results are expressed as means ± standard deviation
of at least two independent experiments, each conducted in
triplicate. Differences between the control and LLFE treatment were
analyzed by one-way ANOVA. Differences were considered to indicate
a statistically significant result at P<0.05.
Results
LLFE inhibits the proliferation of
Bel-7402 cells
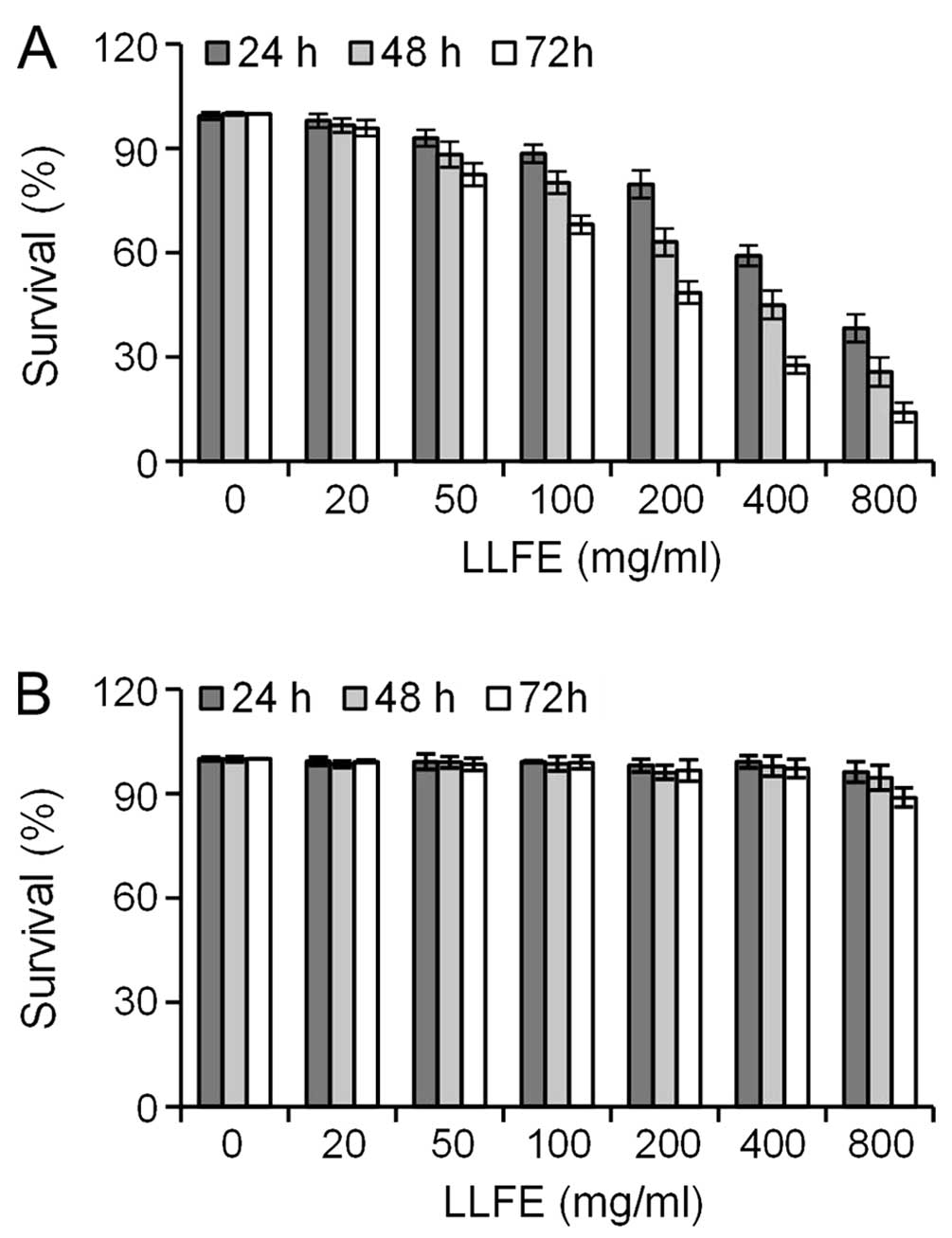
The effect of LLFE on the proliferation of Bel-7402
cells was detected by CCK-8 assay. At final concentrations of
50–800 μg/ml, LLFE significantly inhibited the proliferation of
Bel-7402 cells in a dose- and time-dependent manner (Fig. 1A) (P<0.05). In contrast, LLFE had
no significant effect on the proliferation of human normal
hepatocyte HL-7702 cells even at concentrations that were highly
toxic to the Bel-7402 cells (Fig.
1B). These observations were consistent with a previous report
that aqueous extracts of Ligustrum lucidum are inactive on
normal human mammary epithelial cells (13).
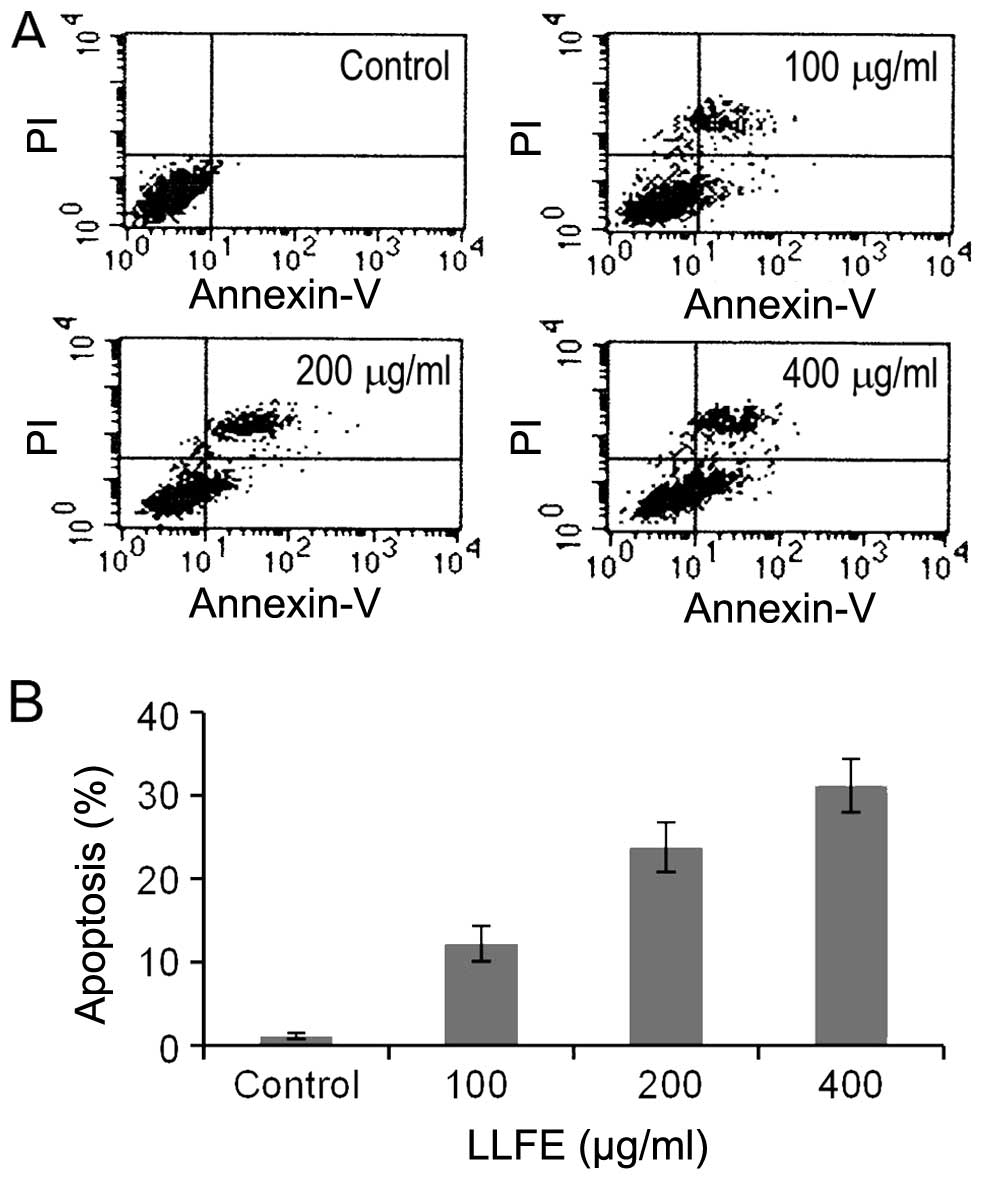
LLFE induces the apoptosis of Bel-7402
cells
Apoptosis, an evolutionarily conserved cell suicide
process elicited by physiological, pathological or pharmacological
stimuli, has been recognized as a major anticancer treatment
response (19,20). Thus, we determined the effects of
LLFE on the apoptosis of Bel-7402 cells. As shown in Fig. 2, treatment with 100–400 μg/ml of
LLFE for 72 h induced significant apoptosis in the Bel-7402 cells
in a dose-dependent manner (P<0.01).
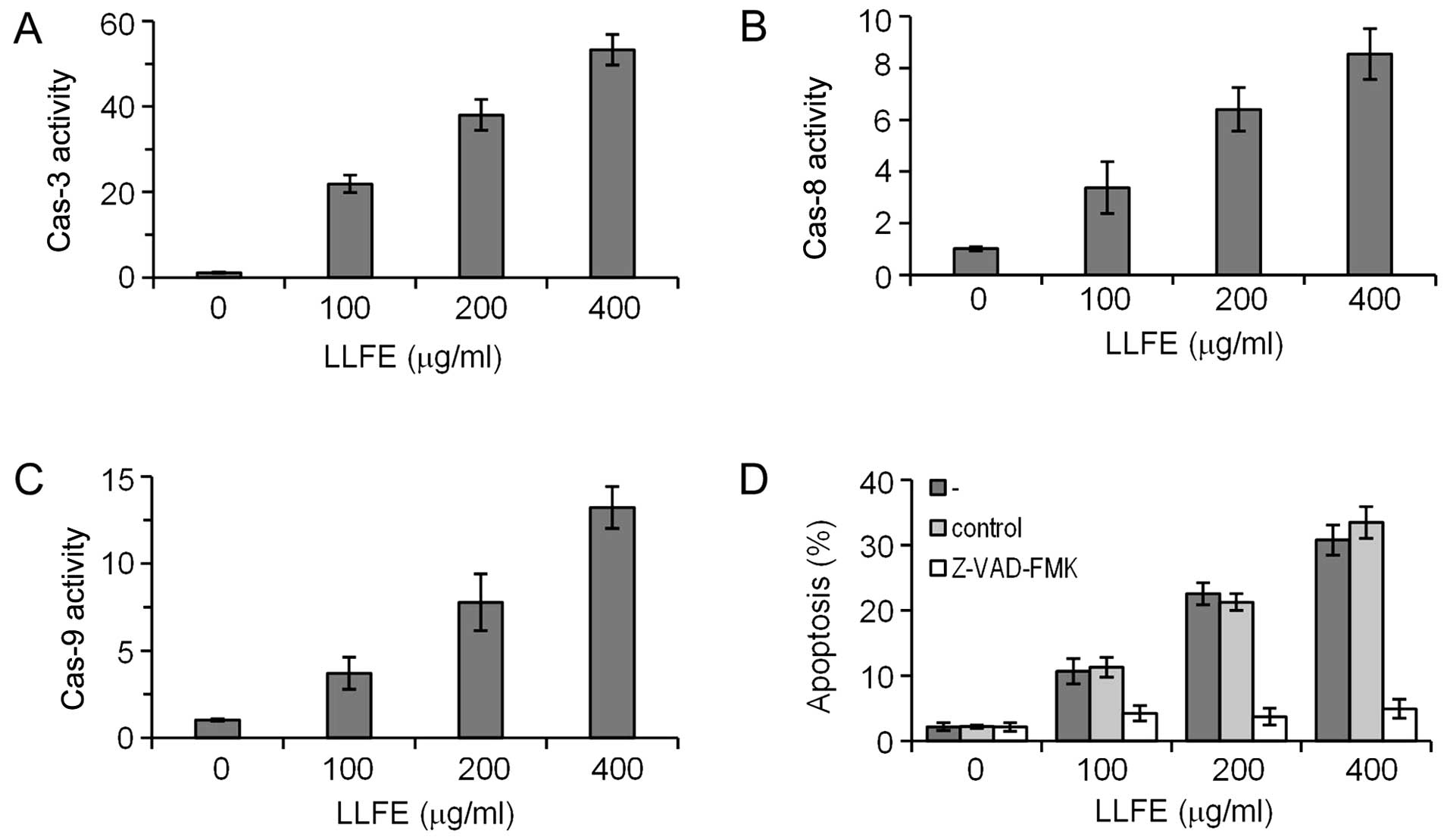
LLFE activates caspases in Bel-7402
cells
To determine whether caspases contribute to the
LLFE-induced apoptosis of Bel-7402 cells, activities of caspases
were measured by the cleavage of the specific substrate. Caspase
activity assays showed that LLFE activated caspase-3, -8 and -9 in
the Bel-7402 cells in a dose-dependent manner (Fig. 3A–C) (P<0.01). In addition,
LLFE-induced apoptosis of Bel-7402 cells was completely abrogated
by the pan-caspase inhibitor Z-VAD-FMK (Fig. 3D) (P<0.01), suggesting that
LLFE-induced apoptosis is associated with the caspase cascade.
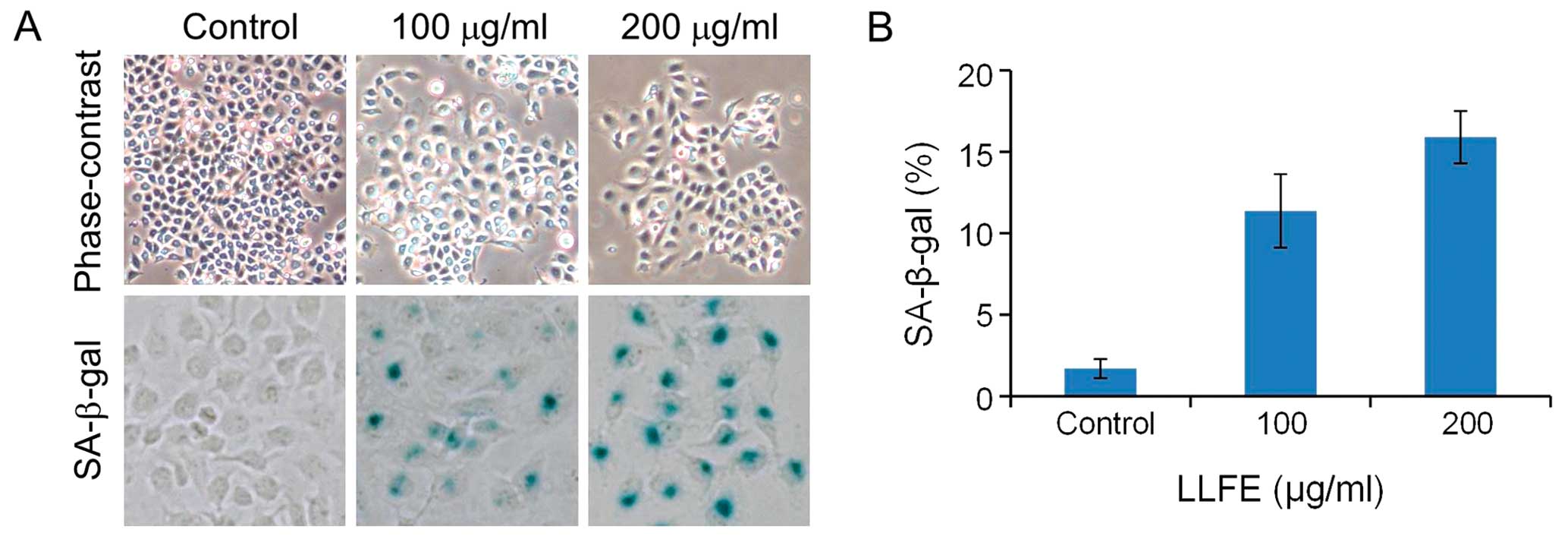
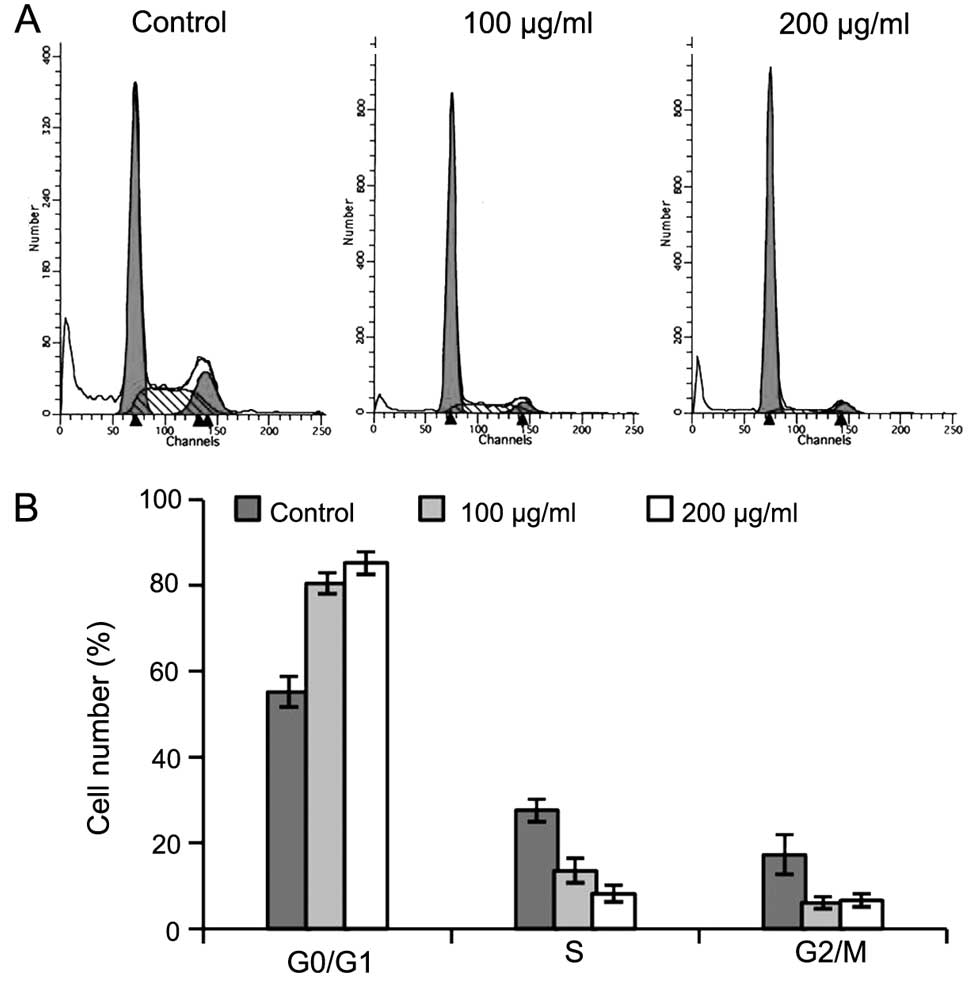
LLFE induces cell senescence in Bel-7402
cells
Upon treatment with low doses of LLFE, the Bel-7402
cells gradually exhibited a large and flattened morphology,
indicative of cell senescence (Fig.
4). Thus, we further performed senescence-activated
β-galactosidase (SA-β-gal) staining. As shown in Fig. 4, LLFE treatment resulted in a higher
percentage of cells with SA-β-gal-positive staining, compared with
the controls (P<0.01). In addition, flow cytometric analysis
revealed that the cell cycle of LLFE-treated Bel-7402 cells was
arrested in the G0/G1 phase (Fig. 5) (P<0.01). These observations
suggest that LLFE induces senescence in Bel-7402 cells.
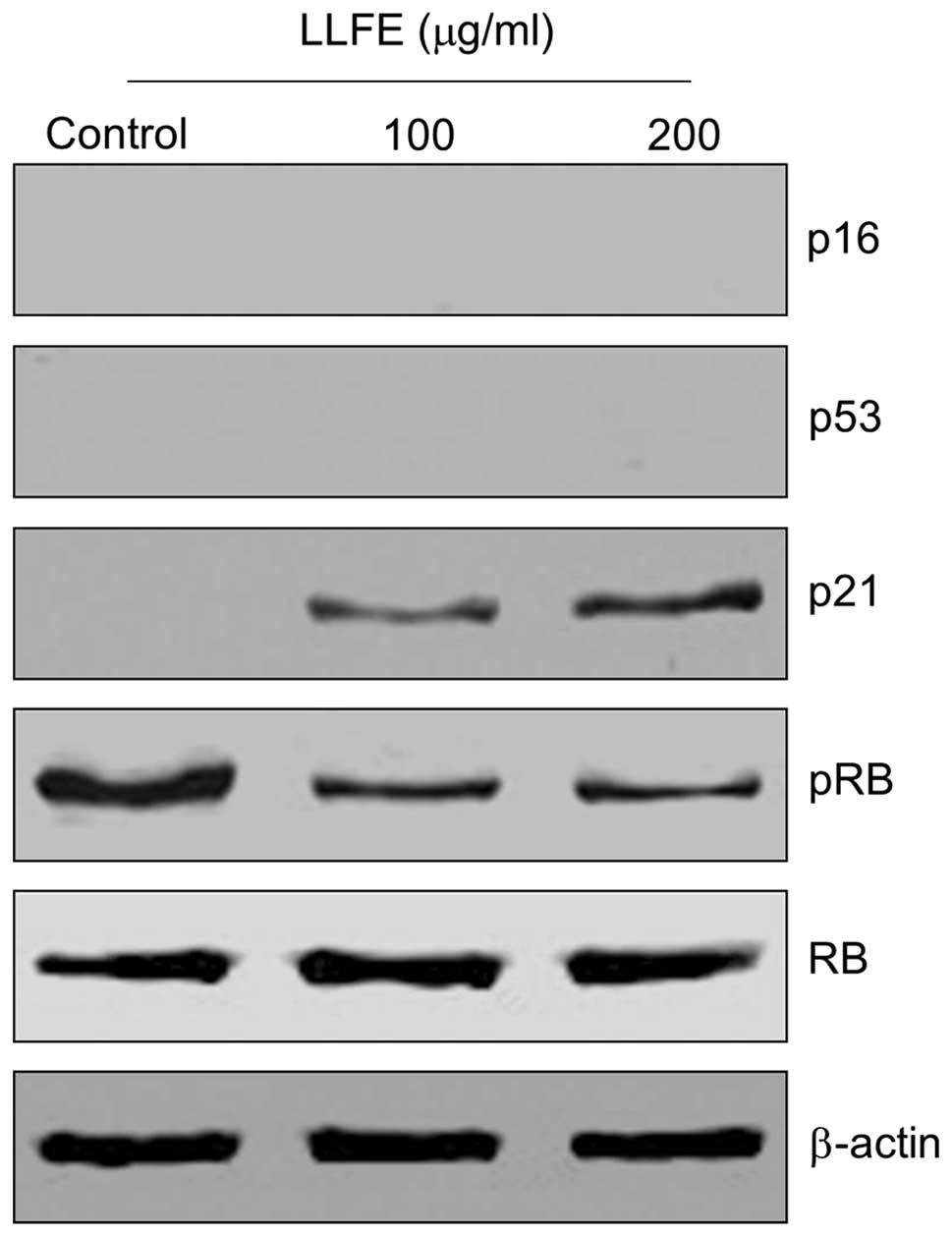
Effects of LLFE on the expression of
senescence regulatory genes
It has been reported that cell senescence is
regulated by the CDKN1a (p21WAF-1/Cip1)/pRB or the
CDKN2a (p16INK4A)/pRB signaling pathway (21,22).
We examined the effects of LLFE on the expression of senescence
regulatory genes in the Bel-7402 cells by western blotting. As
shown in Fig. 6, treatment with low
doses of LLFE caused an upregulation in the p21 expression, and
downregulation of RB phosphorylation. However, expression of p53
and p16 was not detected in the Bel-7402 cells.
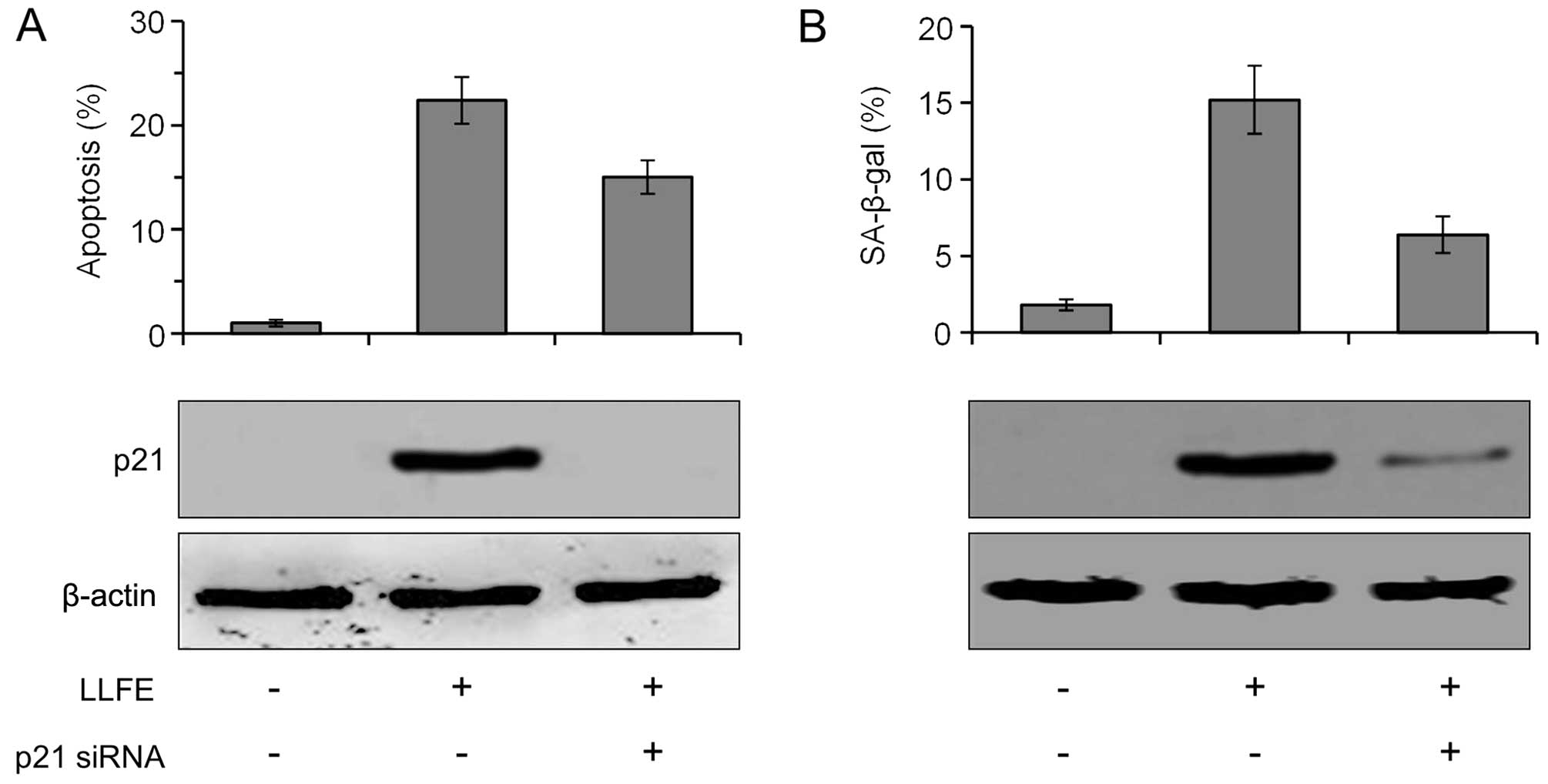
Role of p21 in LLFE-induced apoptosis and
cell senescence
Since p21 expression is upregulated by LLFE, we
further determined the role of p21 in LLFE-induced apoptosis and
cell senescence. As shown in Fig.
7, the expression of p21 was significantly inhibited by
specific siRNA. Specific knockdown of p21 expression partially
abrogated LLFE-induced apoptosis (Fig.
7A), and significantly abrogated LLFE-induced cell senescence
(Fig. 7B). These observations
suggest that p21 may contribute to LLFE-induced apoptosis and cell
senescence.
Discussion
The present study demonstrated that LLFE inhibited
proliferation in a dose- and time-dependent manner, and induced
apoptosis in Bel-7402 cells. The initiation and execution of
apoptosis are dependent on the activation of the extrinsic and/or
intrinsic death pathways (19,20,23).
The extrinsic or death receptor pathways are associated with the
oligomerization of cell-surface death receptors by their ligands,
resulting in recruitment and activation of caspase-8 followed by
activation of executioner caspase-3. On the other hand, intrinsic
or the mitochondrial pathway involves signals to the mitochondria
that lead to the release of cytochrome c and Apaf-1, forming
an apoptosome that activates the initiating protease caspase-9,
which in turn activates caspase-3, causing the cell to undergo
apoptosis. The present study showed that LLFE activates caspase-8,
-9 and -3, and LLFE-induced apoptosis was blocked by a caspase
inhibitor. These observations indicate that LLFE-induced apoptosis
in Bel-7402 cells is through the extrinsic and intrinsic
pathways.
In addition to apoptosis, cell senescence plays an
important role in suppressing tumorigenesis, and may contribute to
the outcome of cancer therapy (19,24–26).
Cell senescence is a state of stable irreversible cell cycle arrest
provoked by a variety of stimuli. Senescent cells maintain some
metabolic activity, but can no longer proliferate, even stimulated
with mitogens. Cell senescence is usually characterized by large
and flattened morphology, an increase in intracellular granules,
elevated SA-β-gal activity, and cell cycle arrest (18,24).
It has been reported that chemotherapeutic agents such as
cisplatin, doxorubicin, SN-38, and camptothecin can inhibit cancer
cell growth via cell senescence (26–29).
In the present study, we observed that a low dose of LLFE treatment
caused large and flat morphologic cellular changes, positive
SA-β-gal staining, and G0/G1 phase cell cycle
arrest, suggesting that LLFE treatment induces cell senescence in
Bel-7402 cells.
Cell senescence is closely related to the activation
of the CDKN1a (p21WAF-1/Cip1)/pRB or the CDKN2a
(p16INK4A)/pRB signaling pathway (21,22).
p21, an important cell cycle regulator, can inhibit a variety of
cyclin/CDK complexes and induce the hypophosphorylation or
dephosphorylation of protein Rb. Hypophosphorylated Rb binds to E2F
and prevents it from activating target genes that are essential in
the cell cycle, usually leading to cell cycle arrest.
Overexpression of p21 may induce a senescence-like state in cancer
cells (30). In addition, p21 also
contributes to drug-induced apoptosis in cancer cells (31,32).
Expression of p21 in cancer cells may be dependent or independent
of p53 (33–36). The present study showed that LLFE
treatment resulted in cell senescence accompanied by upregulation
of p21 and downregulation of RB phosphorylation, suggesting that
p21/RB may contribute to LLFE-induced cell senescence. Since p53
was not expressed in the Bel-7402 cells, p21 induced by LLFE may be
independent of p53. Further observations revealed that knockdown of
p21 expression significantly abrogated LLFE-induced cell
senescence, suggesting that the effects of LLFE on cell senescence
may depend on p21. LLFE-induced apoptosis was partially abrogated
by p21 silencing suggesting that the effects of LLFE on apoptosis
may partially involve p21.
In summary, the present study demonstrated that LLFE
activates caspases to induce apoptosis in human hepatocellular
carcinoma Bel-7402 cells with the participation of p21. LLFE also
induced cell senescence in Bel-7402 cells, which may correlate with
upregulation of p21 and downregulation of RB phosphorylation. These
observations suggest that Nü-zhen-zi is a potential anticancer herb
and support the traditional use of Nü-zhen-zi for liver cancer
treatment. Nevertheless, further studies are needed to determine
the upstream signal transduction of p21 upregulation, and to
address which chemical(s) are responsible for the LLFE-induced
anticancer effects.
Acknowledgements
This study was partially supported by the National
Natural Science Foundation of China (81273726), the Three-year
Action Program of Shanghai Municipality for Traditional Chinese
Medicine (ZYSNXD-CC-ZDYJ017), the Xing-Lin Scholars Program, the
Long-Yi Scholars and Research Team Program from the State Clinical
Research Center of TCM in Longhua Hospital (LYTD-04), and the
Shanghai Shen Kang Platform Grant (SHDC12007206). We also thank the
Shanghai Key Laboratory of Tissue Engineering for technical
assistance.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar
|
|
2
|
Yu Y, Lang Q, Chen Z, Li B, Yu C, Zhu D,
Zhai X and Ling C: The efficacy for unresectable hepatocellular
carcinoma may be improved by transcatheter arterial
chemoembolization in combination with a traditional Chinese herbal
medicine formula: a retrospective study. Cancer. 115:5132–5138.
2009. View Article : Google Scholar
|
|
3
|
Meng MB, Cui YL, Guan YS, Ying Z, Zheng
MH, Yuan CK and Zhang RM: Traditional Chinese medicine plus
transcatheter arterial chemoembolization for unresectable
hepatocellular carcinoma. J Altern Complement Med. 14:1027–1042.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shu X, McCulloch M, Xiao H, Broffman M and
Gao J: Chinese herbal medicine and chemotherapy in the treatment of
hepatocellular carcinoma: a meta-analysis of randomized controlled
trials. Integr Cancer Ther. 4:219–229. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Du Q, Hu B, Shen KP and An HM:
Pathogenesis and treatment law of Traditional Chinese Medicine in
hepatocarcinoma. World J Integr Tradit West Med. 5:814–817.
2010.(In Chinese).
|
|
6
|
Yan JG and Yao SK: The study of TCM
syndrome differentiation and treatment in liver cancer. Chin J
Integr Tradit West Med Liver Dis. 20:189–190. 2010.(In
Chinese).
|
|
7
|
Pan MQ and Tian H: Employ the methods of
invigorating spleen and regulating Qi, clearing toxic heat,
dissipating stasis and softening hardness to treat primary hepatic
cancer. J Tradit Chinese Med. 34:239–240. 1993.(In Chinese).
|
|
8
|
Li YL and Zhang SP: The effects of
invigorating spleen and regulating Qi in patients with primary
hepatic cancer: an analysis of 127 cases. Tianjin Tradit Chinese
Med. 17:12–13. 2000.(In Chinese).
|
|
9
|
Liu Q, Zhang YB, Ma CH, Yue XQ and Ling
CQ: Analysis of literature on therapeutic methods and medicines of
traditional Chinese medicine for primary liver cancer. Zhong Xi Yi
Jie He Xue Bao. 3:260–262. 2005.(In Chinese).
|
|
10
|
Bo X, Zhao XM, Liu HT, Ma WG and Li H: A
study on 4 kinds of Chinese medicines reducing arrest of bone
marrow by cyclophosphomidum. J Beijing Union Univ (Natural
Science). 8:58–61. 1994.(In Chinese).
|
|
11
|
Zhao YL and Fan WX: The effects of
Nü-zhen-zi formula on chemotherapy induced alopecia. Acta Univ Med
Nanjing (Natural Science). 24:305–306. 2004.(In Chinese).
|
|
12
|
Wu XF, Han SY, Zhu LS, Bai J and Liu SM:
The effects of Ligustrum lucidum Ait. fruits extract on
cyclophosphomidum induced immunosuppression. J North China Coal Med
Coll. 10:303–304. 2008.(In Chinese).
|
|
13
|
Shoemaker M, Hamilton B, Dairkee SH, Cohen
I and Campbell MJ: In vitro anticancer activity of twelve Chinese
medicinal herbs. Phytother Res. 19:649–651. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jeong JC, Kim JW, Kwon CH, Kim TH and Kim
YK: Fructus ligustri lucidi extracts induce human glioma
cell death through regulation of Akt/mTOR pathway in vitro and
reduce glioma tumor growth in U87MG xenograft mouse model.
Phytother Res. 25:429–434. 2011.
|
|
15
|
Zhang JF, He ML, Dong Qi, Xie WD, Chen YC,
Lin MC, Leung PC, Zhang YO and Kung HF: Aqueous extracts of Fructus
Ligustri Lucidi enhance the sensitivity of human colorectal
carcinoma DLD-1 cells to doxorubicin-induced apoptosis via Tbx3
suppression. Integr Cancer Ther. 10:85–91. 2011. View Article : Google Scholar
|
|
16
|
Hu B, An HM, Shen KP, Xu L, Du Q, Deng S
and Wu Y: Modified Yi Guan Jian, a Chinese herbal formula, induces
anoikis in Bel-7402 human hepatocarcinoma cells in vitro.
Oncol Rep. 26:1465–1470. 2011.PubMed/NCBI
|
|
17
|
Hu B, Shen KP, An HM, Wu Y and Du Q:
Aqueous extract of Curcuma aromatica induces apoptosis and
G2/M arrest in human colon carcinoma LS-174-T cells independent of
p53. Cancer Biother Radiopharm. 26:97–104. 2011.
|
|
18
|
Dimri GP, Lee X, Basile G, Acosta M, Scott
G, Roskelley C, Medrano EE, Linskens M, Rubelj I, Pereira-Smith O,
Peacocke M and Campisi J: A biomarker that identifies senescent
human cells in culture and in aging skin in vivo. Proc Natl Acad
Sci USA. 92:9363–9367. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chiantore MV, Vannucchi S, Mangino G,
Percario ZA, Affabris E, Fiorucci G and Romeo G: Senescence and
cell death pathways and their role in cancer therapeutic outcome.
Curr Med Chem. 16:287–300. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Brown JM and Attardi LD: The role of
apoptosis in cancer development and treatment response. Nat Rev
Cancer. 5:231–237. 2005.PubMed/NCBI
|
|
21
|
Roninson IB: Tumor cell senescence in
cancer treatment. Cancer Res. 63:2705–2715. 2003.PubMed/NCBI
|
|
22
|
Dimri GP: What has senescence got to do
with cancer? Cancer Cell. 7:505–512. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Meier P and Vousden KH: Lucifer’s
labyrinth - ten years of path finding in cell death. Mol Cell.
28:746–754. 2007.
|
|
24
|
Schmitt CA: Cellular senescence and cancer
treatment. Biochim Biophys Acta. 1775:5–20. 2007.PubMed/NCBI
|
|
25
|
Gewirtz DA, Holt SE and Elmore LW:
Accelerated senescence: an emerging role in tumor cell response to
chemotherapy and radiation. Biochem Pharmacol. 76:947–957. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Nardella C, Clohessy JG, Alimonti A and
Pandolfi PP: Pro-senescence therapy for cancer treatment. Nat Rev
Cancer. 11:503–511. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Di X, Shiu RP, Newsham IF and Gewirtz DA:
Apoptosis, autophagy, accelerated senescence and reactive oxygen in
the response of human breast tumor cells to adriamycin. Biochem
Pharmacol. 77:1139–1150. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
te Poele RH, Okorokov AL, Jardine L,
Cummings J and Joel SP: DNA damage is able to induce senescence in
tumor cells in vitro and in vivo. Cancer Res. 62:1876–1883.
2002.PubMed/NCBI
|
|
29
|
Han Z, Wei W, Dunaway S, Darnowski JW,
Calabresi P, Sedivy J, Hendrickson EA, Balan KV, Pantazis P and
Wyche JH: Role of p21 in apoptosis and senescence of human colon
cancer cells treated with camptothecin. J Biol Chem.
277:17154–17160. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kagawa S, Fujiwara T, Kadowaki Y, Fukazawa
T, Sok-Joo R, Roth JA and Tanaka N: Overexpression of the
p21sdi1 gene induces senescence-like state
in human cancer cells: implication for senescence-directed
molecular therapy for cancer. Cell Death Differ. 6:765–772.
1999.
|
|
31
|
Ahmad N, Adhami VM, Afaq F, Feyes DK and
Mukhtar H: Resveratrol causes WAF-1/p21-mediated
G1-phase arrest of cell cycle and induction of apoptosis
in human epidermoid carcinoma A431 cells. Clin Cancer Res.
7:1466–1473. 2001.PubMed/NCBI
|
|
32
|
Liu WK, Cheung FW, Liu BP, Li C, Ye W and
Che CT: Involvement of p21 and FasL in induction of cell cycle
arrest and apoptosis by neochamaejasmin A in human prostate LNCaP
cancer cells. J Nat Prod. 71:842–846. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Thaler S, Hähnel PS, Schad A, Dammann R
and Schuler M: RASSF1A mediates p21Cip1/Waf1-dependent
cell cycle arrest and senescence through modulation of the
Raf-MEK-ERK pathway and inhibition of Akt. Cancer Res.
69:1748–1757. 2009.PubMed/NCBI
|
|
34
|
Peng CC, Chen KC, Peng RY, Chyau CC, Su CH
and Hsieh-Li HM: Antrodia camphorata extract induces
replicative senescence in superficial TCC, and inhibits the
absolute migration capability in invasive bladder carcinoma cells.
J Ethnopharmacol. 109:93–103. 2007. View Article : Google Scholar
|
|
35
|
Wakoh T, Uekawa N, Terauchi K, Sugimoto M,
Ishigami A, Shimada J and Maruyama M: Implication of p53-dependent
cellular senescence related gene, TARSH in tumor suppression.
Biochem Biophys Res Commun. 380:807–812. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lodygin D, Menssen A and Hermeking H:
Induction of the Cdk inhibitor p21 by LY83583 inhibits tumor cell
proliferation in a p53-independent manner. J Clin Invest.
110:1717–1727. 2002. View Article : Google Scholar : PubMed/NCBI
|