1. Introduction
The rising burden of cancer places enormous strains
on public health care systems and is one of the major causes of
death in humans. To develop individualized cancer therapy and
prevention, natural agents will become important strategies in the
near future (1). Natural products,
as an attractive source of new therapeutic candidate compounds for
cancer prevention and therapy, have been playing an increasingly
important role. The analysis concerning chemotherapeutic agents and
their sources indicates that over 60% of approved drugs are derived
from natural compounds (2). In the
past few decades, through large-scale anticancer drug screening and
discovery programs, various successful anticancer agents in
clinical use or in clinical trials have been identified that are
derived from natural products and their synthetic analogues. Among
these, vincristine, etoposide and paclitaxel are plantderived
compounds; and actinomycin D, rapamycin and doxorubicin are drugs
originating from microbial sources (3). Through a variety of different
mechanisms of action, such as DNA damage, inhibition of
topoisomerases I or II, and disruption of cell signaling
transduction, these agents inhibit cancer cell proliferation and
progression (3). Carcinogenesis is
a multistep process in which an accumulation of genetic mutations
leads to progressive DNA repair, cell cycle, DNA methylation, cell
death and cell growth deregulation, and eventually to carcinoma. By
the inhibition of virus infection and the regulation of metabolism
and the immune system, cancer preventive agents are able to sustain
cancer as a chronic disease.
A cellular signaling pathway, including a variety of
proteins that consist of regulatory circles, is a complex signal
communication network that controls basic biological activities of
the cell and coordinates cell actions. Several cellular signaling
pathways, including Nf-κB, PI3K/Akt, MAPKs and p53, have been known
to regulate cell proliferation and apoptosis (4). Due to the complexity and the abnormal
alterations in cell signaling transduction, specific inhibitors
that only have one target, most often fail in cancer therapy. It
has been shown that different signaling pathways have crosstalk
with each other. The simultaneous targeting of multiple cellular
signaling pathways which control cell cycle and apoptosis to induce
cell cycle arrest and cell death is an important strategy to
control cancer cell proliferation and the formation of tumors
(5).
Several excellent review have been published on the
anticancer activity of natural compounds (2,3). In
the present review, we summarize research conducted by our
laboratory concerning various natural phenolic compounds by a
screening platform established by our laboratory. These natural
compounds include grifolin, neoalbaconol and
epigallocatechin-3-gallate (EGCG). Grifolin, by targeting ERK or
upregulating DAPK1 via p53, was found to induce the apoptosis and
cell cycle arrest in multiple cancer cell lines (6–9). We
also demonstrated that its analogue, neoalbaconol, regulated cell
metabolism and ultimately induced multiple types of cell death
in vitro and in vivo (10). Even though there are numerous
studies concerning the anticancer activity of EGCG, our group found
that it regulated EBV lytic infection through the PI3K/Akt and MAPK
pathways in EBV-positive cancer cells (11). Therefore, we review the findings of
three agents to be used as potential leading compounds in the
prevention and inhibition of tumor progression and/or EBV-related
cancer.
2. Grifolin, a promising kinase inhibitor
candidate, regulates cancer growth via targeting ERK1/2 and
DAPK1
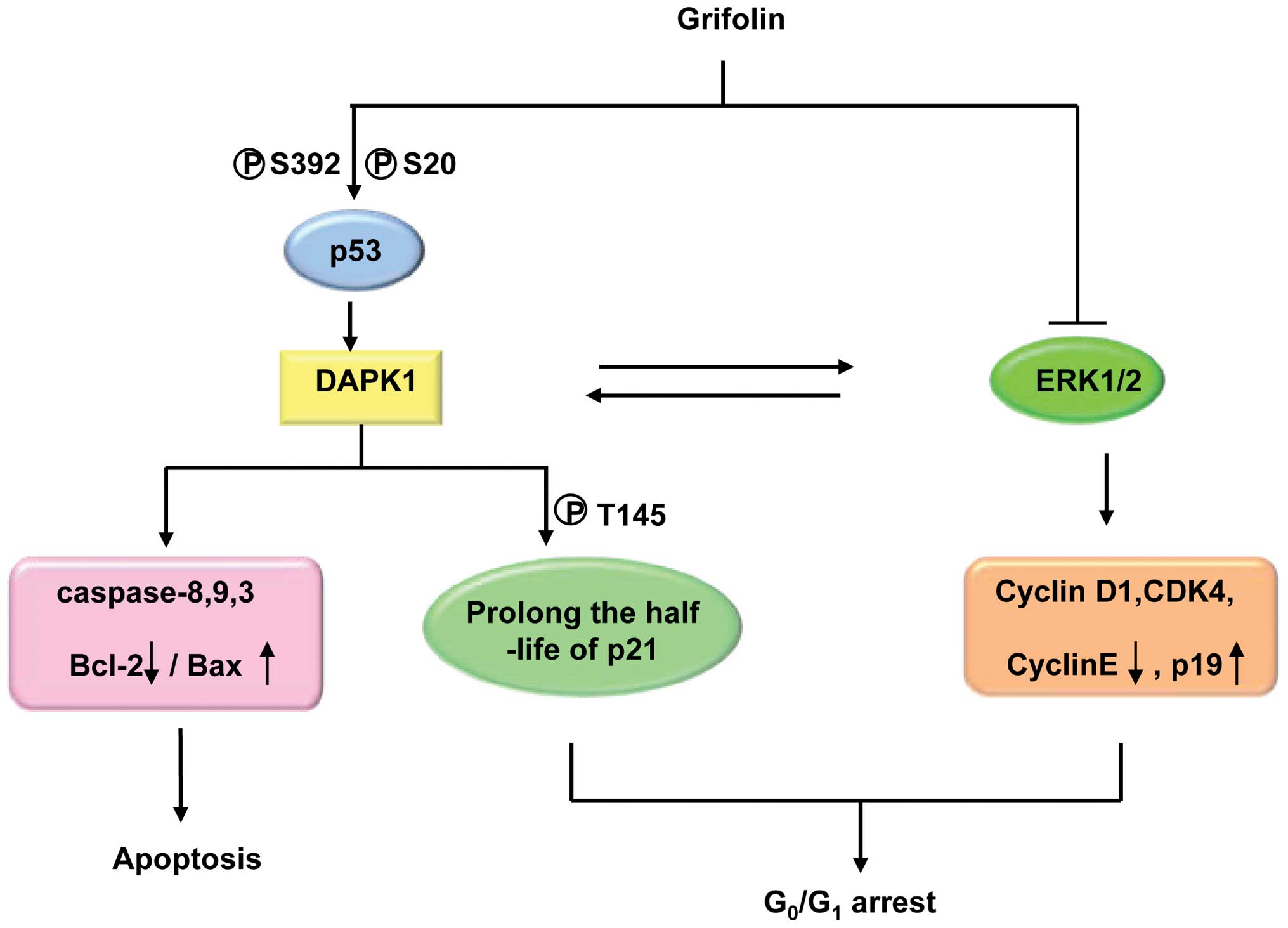
Grifolin, a farnesyl phenolic compound (fig. 1A), is a secondary metabolite
isolated from the fresh fruiting bodies of the mushroom
Albatrellus confluens. It has also been reported to
originate from the edible mushroom Boletus pseudocalopus
(12). Grifolin displays various
pharmacological and microbiological effects. Recent evidence
indicates that grifolin also possesses antioxidant and antitumor
activities (6–8,13–15).
It has been shown to inhibit the growth of various cancer cell
lines in vitro by induction of cell cycle arrest and
apoptosis in previous studies by our group (6,7).
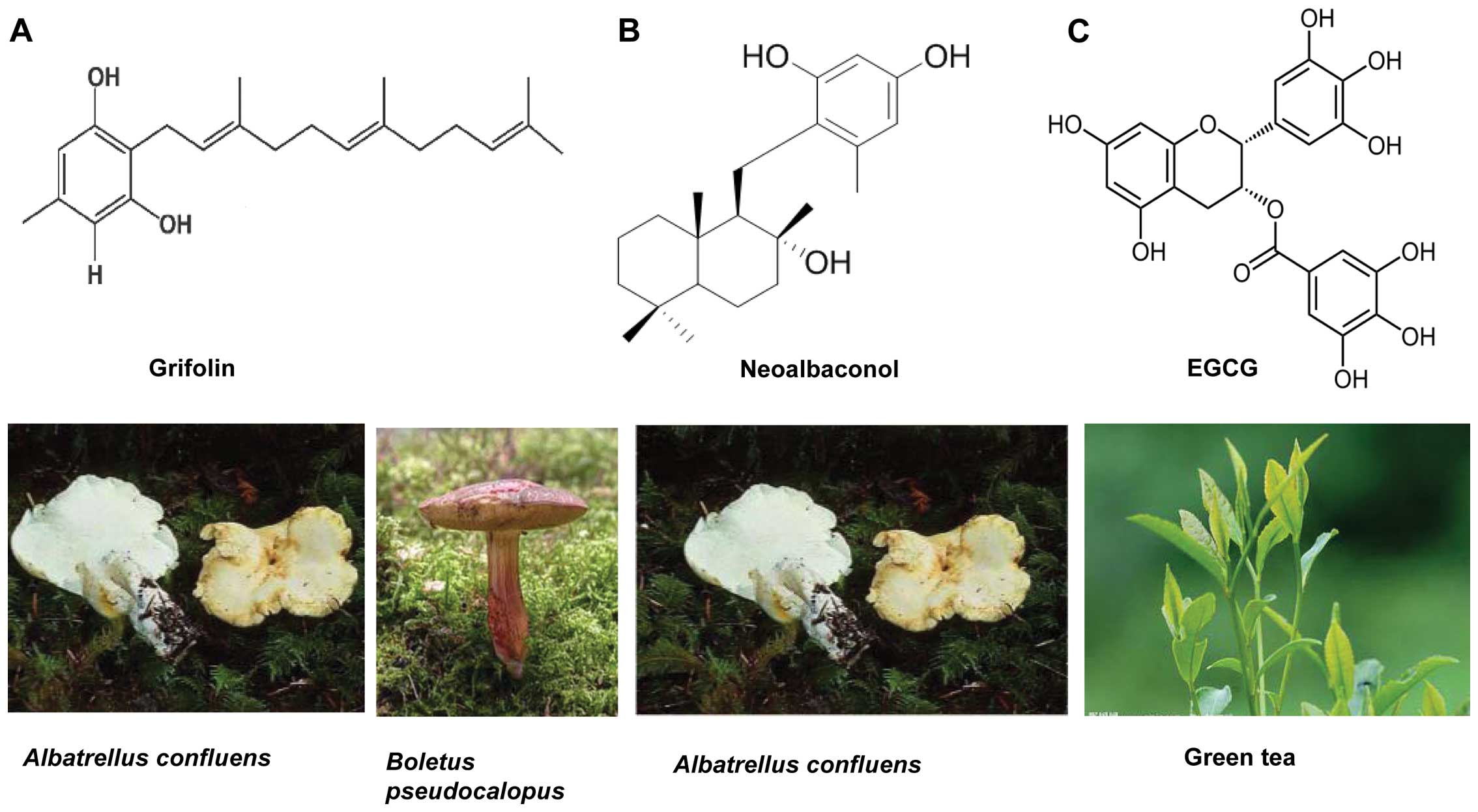 | Figure 1Molecular structures of the three
natural compounds. (A) Grifolin,
2-trans,trans-farnesyl-5-methylresorcinol. (B)
Neoalbaconol is an important analogue of grifolin, 2-{[rel-
(1R,2R,4aS,8aS)-decahydro-2-hydroxy-2,5,5,8a-tetramethylnaphthalen-1-yl]methyl}-3-methylbenzene-1,5-diol.
(C) EGCG, the ester of epigallocatechin and gallic acid, is a type
of catechin
[(2R,3R)-5,7-dihydroxy-2-(3,4,5-trihydroxyphenyl)chroman-3-yl]-3,4,5-trihydroxybenzoate. |
Our research indicates that grifolin exhibits
certain selective antitumor effects following the comparison of the
respective IC50 values of a broad spectrum of tumor cell
lines to normal or non-tumor cell lines (6,7). The
apoptotic effect of grifolin on nasopharyngeal carcinoma (NPC) cell
line CNE1 was found to be mediated by successive cascade responses,
including a decrease in the Bcl-2 level, an increase in Bax,
release of cytochrome c from mitochondria, and activation of
caspase-8, -9 and -3.
To identify the molecular targets in the signal
transduction pathways, apoptosis-related cDNA microarray analysis
was employed to investigate the mechanism of grifolin-induced cell
death at the gene expression level. Our studies identified that the
dapk1 gene was significantly upregulated. DAPK1 is an
apoptotic-positive mediator (16).
It acts as a tumor suppressor largely due to its ability to
sensitize cells to many apoptotic signals including those generated
by death receptors, cytokines, matrix detachment and
oncogene-induced hyperproliferation that are encountered as a cell
undergoes tumorigenesis. Extensive data in human primary tumors
demonstrate a significant loss of DAPK1 expression in a large
variety of tumor types (17,18).
Grifolin was found to upregulate DAPK1 mRNA and protein expression
in a dose-dependent manner in NPC cells. DAPK1 mRNA levels were
found to increase in a p53-dependent manner in various cellular
settings (19,20). We observed that grifolin promoted
the phosphorylation of p53 at Ser392 and Ser20, while it had little
effect on the phosphorylation of Ser15 or Thr81 as well as the
total p53 protein level. General consensus remains that the
phosphorylation of N-terminal regulatory sites of p53 occurs
rapidly in response to various stress stimuli to activate p53.
Reduced levels of Ser20 phosphorylation were found to attenuate p53
activity as a transcription factor (19,21).
It is currently believed that both the core DNA-binding and the
C-terminal domains of p53 possess DNA-binding activities, in that
the former primarily provides sequence specificity whereas the
latter recognizes structural features of target DNA. Using EMSA, we
further demonstrated that grifolin significantly increased the
binding activity of p53 to the dapk1 gene in vitro. ChIP
assays of the p53-dapk1 promoter complex confirmed that endogenous
p53 interacts with this region in vivo (8). We introduced an siRNA targeting DAPK1
and a scrambled siRNA as a control into CNE1 cells. Depletion of
DAPK1 reduced the apoptotic effect as well as the activation of
caspase-3 induced by grifolin. Grifolin-induced upregulation of
DAPK1 was also observed in tumor cells derived from human breast
cancer and human colon cancer. In SW480 and MCf7 cells, grifolin
also upregulated DAPK1 via the p53 pathway, and DAPK1 was found to
mediate the grifolin-induced apoptotic effect (8). Reintroduction of p53 and DAPK1 into
p53-null H1299 cells markedly enhanced the apoptotic rate, which
further confirmed the function of the p53-DAPK1 pathway in
grifolin-induced apoptosis.
Collectively, upregulation of DAPK1 expression by
grifolin may be an important mechanism contributing to its ability
to induce an apoptotic effect in tumor cells. Due to the high
frequency of loss of DAPK1 expression in a large variety of tumor
types, grifolin may rescue the pro-apoptotic function of DAPK1 via
the p53 pathway.
At high doses, both the ERK1/2 and the ERK5 pathways
may be involved in grifolin-induced cell cycle arrest. ERK has a
well-established role in regulating G1 to S phase
progression in response to mitogenic stimulation. ERK mediates
activation of multiple transcription factors including Elk1, c-Jun,
c-Myc and c-Fos. These transcription factors control the expression
of genes important for cell cycle progression, including cyclin D1
and p21WAF1/CIP1 (22).
The results obtained from evaluating the effects of grifolin on the
expression of G1-related protein suggested that cell
cycle arrest was associated with the downregulation of cyclin D1,
CDK4, cyclin E as well as the phosphorylation of pRB induced by
grifolin. In contrast, INK4 family member p19, which mediates the
inhibition of CDK4, was obviously upregulated by grifolin in a
dose-dependent manner (7).
In our previous study, we demonstrated that DAPK1
also mediated grifolin-induced G1 phase arrest effects
in NPC ells (7). DAPK1 can be
activated by several mechanisms, including dephosphorylation of
Ser308 by an unknown phosphatase, which can be activated by several
death signals (23,24). We found that grifolin induced
dephosphorylation of DAPK1 (Ser308) and subsequent phosphorylation
of p21 (Thr145). Inhibition of DAPK1 by introducing siDAPK1
reversed grifolininduced phosphorylation of p21. Furthermore, we
confirmed that grifolin increased the half-life of p21 and promoted
its stability by increased ubiquitination of p21. Normal p21
turnover is suppressed by inhibitors of the proteasome pathway, yet
degradation is independent of p21 ubiquitination of lysine residues
despite amino terminal addition (25,26).
Therefore, a different mechanism-driven ubiquitin-linkage may lead
to a different fate for the p21 protein. The ubiquitination of p21
enhances its stability and promotes its function as a cell-cycle
inhibitor. During G1-phase progression, cyclin D1/CDK4/6
complexes are activated in mid-G1 phase and are
essential for G1/S transition. The enhanced stability of
p21 induced by grifolin may effectively inhibit the activity of the
cyclin D1/CDK4/6 complex, leading to G1 phase arrest in
NPC.
Two proteins have been shown to independently
interact with the DAPK1 death domain, which include ERK protein.
Grifolin promotes the interaction of DAPK1 and ERK1/2, leading to
cytoplasmic retention of ERK1/2 (27). In summary, the activation of DAPK1
induced by grifolin thereby enhancing stability of p21 may
effectively inhibit the activity of the cyclin D1/CDK4/6 complex,
leading to G1 phase arrest in NPC. In addition, the
cytoplasmic retention of ERK1/2 synergistically enhances the
G1 phase arrest induced by grifolin (9).
Most importantly, we observed that grifolin
inhibited the kinase activity of EKR1/2 protein in vivo and
in vitro. We next used molecular modeling with the crystal
structure of ERK2 to analyze whether grifolin binds to ERK2. We
found that grifolin formed a hydrogen bond with Ile-29 at the
backbone of ERK2, and also formed hydrophobic interactions with
ERK2 at Val-37, Leu-154 and Cys-164. We further identified that
grifolin physically binds to ERK2 using affinity chromatography and
fluorescence quenching analysis (unpublished data). Grifolin, a
natural compound from mushroom Albatrellus confluens,
represents a promising kinase inhibitor candidate in the
intervention of cancer via targeting ERK1/2 and DAPK1 (fig. 2).
3. Neoalbaconol, a potential inhibitor of
PDK1, induces multiple types of cell death via the PI3K/AKT
pathway
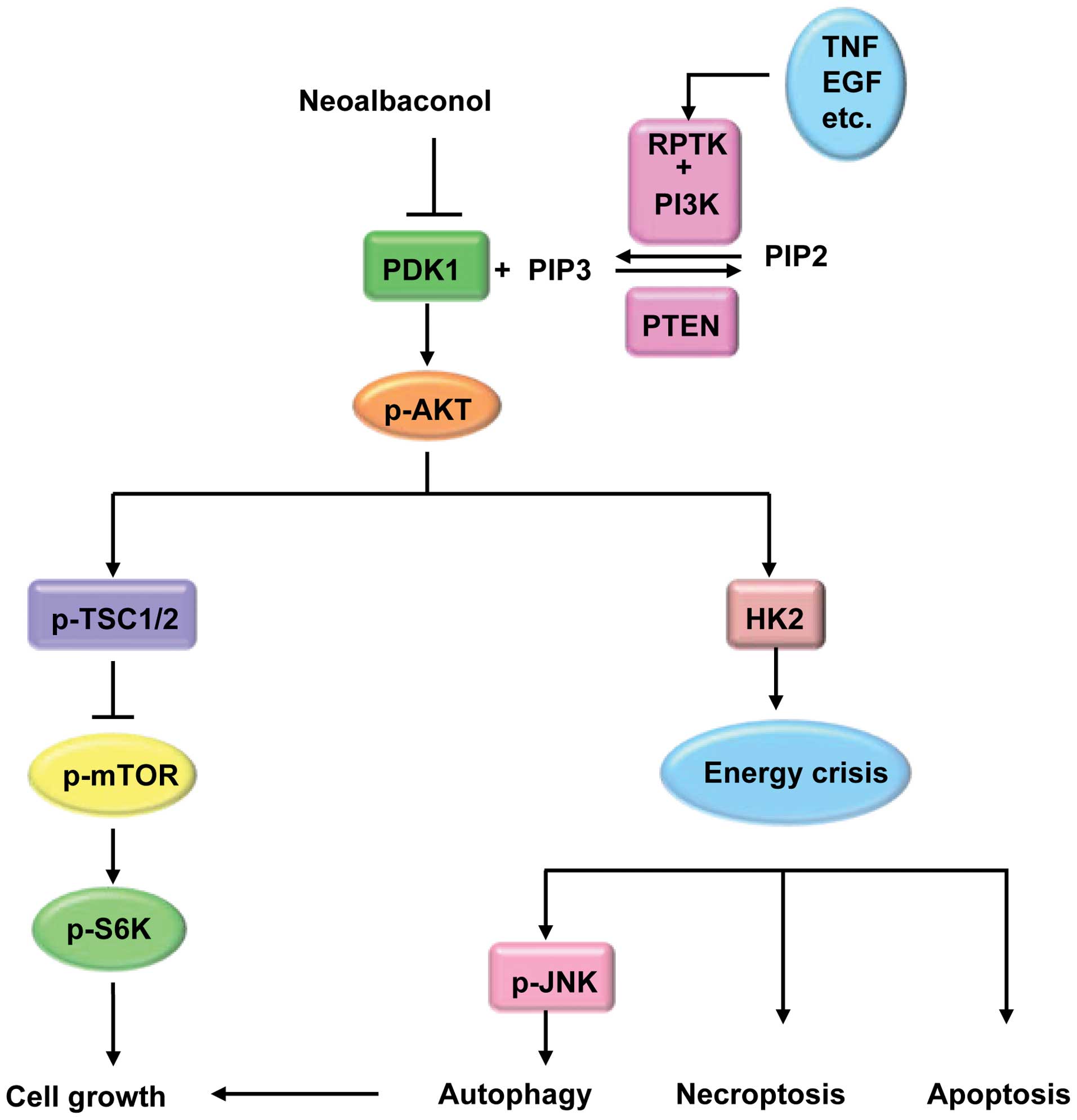
Neoalbaconol (NA), a novel small-molecular compound
with a drimane-type sesquiterpenoid structure (fig. 1B), was also isolated from the
fruiting bodies of the mushroom Albatrellus confluens.
Albatrellus confluens, mainly distributed in southeast
China, is a member of the Polyporaceae family. Several compounds,
such as grifolin and albaconol, with anticancer potential or
anti-inflammatory action have been isolated from this fungus
(28,29).
Cell death plays a key role in regulating tissue
homeostasis and physiologic processes; abnormal regulation of this
process is associated with a number of diseases, such as
neurodegeneration, ischemia and cancer (30). According to the morphological
appearance, cell death can be mainly classified into three distinct
routes: apoptosis, autophagic cell death and necrosis (31). Traditionally, apoptosis and
autophagic cell death are recognized as programmed cell death, and
necrosis is considered as accidental and unregulated. Yet, a
previous study demonstrated that necrosis is also regulated. This
form of programmed necrosis termed necroptosis can be inhibited by
small-molecular compounds such as necrostatin 1 (Nec-1) and its
execution involves the receptor interacting protein kinase (RIPK) 1
and 3 (32). Our previous studies
demonstrated that NA significantly inhibited the proliferation of
many types of cancer cell lines in a dose-dependent manner. Yet, it
did not affect the proliferation of normal immortalized cell lines
even at high doses, indicating the selectivity of NA toward cancer
cells. Using flow cytometric, protein, confocal and electron
microscope (EM) assays, our results showed that NA induced cell
apoptosis, autophagy and necroptosis (10). In our previous study, apoptosis and
necroptosis were responsible for the death-inducing efficacy of NA,
and necroptosis was found to contribute to the main effect of
NA-induced cell death in NPC cells (10). The apoptosis inhibitor zVAD-fmk and
necroptosis inhibitor Nec-1 rescued the viability of the NA-treated
cells, which confirmed the role of apoptosis and necroptosis in
NA-induced cell death (10).
As investigations have recently discovered that
autophagic vesicles are commonly observed in necroptotic cells, the
role of autophagy in cell viability is believed to be dependent on
the cell context yet remains controversial (32). It has been reported that the
suppression of pro-survival autophagy leads to necroptosis induced
by zVAD-fmk in L929 cells (33).
Yet, in proliferative T cells, Fas-associated protein with death
domain (FADD) in combination with caspase-8 limited autophagy and
protected T cells from necroptosis; while Nec-1 reduced light chain
levels in FADD−/− cells and rescued the cell cycling and
proliferation of FADD−/− T cells simultaneously
(34). We found that the
NA-mediated apoptosis, necroptosis and autophagy occurred
independent of each other (10).
Inhibition of autophagy by 3-MA enhanced cell death in NA-treated
cells, suggesting that autophagy provides a survival force in this
model. Energy metabolic reprogramming is one of the significant
features of cancer cells. The oncogenic PI3K/AKT/mTOR pathway plays
a key role in reprogramming metabolic pathways in cancer cells
(35). PDK1, an upstream regulator
of Akt/mTOR signaling, activates a group of protein kinases
belonging to the AGC kinase family including protein kinase A, G
and C (36). In response to various
cellular stimulations, PDK1 phosphorylates Akt at Ser308, resulting
in the activation of Akt and regulates energy metabolism, cell
proliferation, cell cycle progression and migration. Based on the
important functions of PDK1 in tumor cells, researchers have
recently shown that PDK1 serves as an effective therapeutic target
for anticancer treatment and several PDK1 inhibitors, such as AR-12
and GSK233447, have been developed to kill cancer cells (37). Using the phase module of the
Schrödinger molecular modeling software package, we identified PDK1
as a potential protein target of NA (10). More than three ligandbinding sites
are located in the PDK1 kinase domain, including an ATP-binding
pocket, a peptide substrate-binding site and a groove in the
N-terminal that binds to the C-terminal hydrophobic motif of the
kinase substrates (37). NA was
able to dock into the ATP binding pocket of PDK1 and form three
hydrogen bonds with the backbone of PDK1. By targeting PDK1 and
inhibiting the downstream PI3-K/Akt pathway, NA inhibited the key
energy metabolic enzyme HK2 in cancer cells (10). Thus, the final effect of NA
treatment not only significantly decreased the glucose
concentration in the medium, yet also blocked ATP generation in a
time-dependent manner.
Moreover, the necroptosis inhibitor Nec-1 rescued
the viability of the NA-treated cells. Akt overexpression decreased
LC3 expression and inhibited NA-induced autophagy, implying that
Akt inactivation and energy crisis are responsible for NA-induced
autophagy (10). We also
demonstrated the efficacy of NA in inhibiting tumor growth by
suppressing the Akt signaling pathway in vivo (10). Taken together, NA may be involved in
NA-induced apoptotic and necroptotic cell death by remodeling
cellular energy metabolism by targeting the PDK1/PI3K/Akt signaling
pathway (Fig. 3). All these
findings strongly suggest that the inhibition of tumor cell growth
by NA makes it an ideal candidate as a leading anticancer
agent.
4. EGCG, a potential chemopreventive agent,
controls EBV lytic infection by targeting the MEK/ERK1/2 and
PI3K/Akt pathways
The consumption of green tea has long been
associated with a reduced risk of cancer development. Green tea is
chemically characterized by the presence of high amounts of
polyphenolic compounds known as catechins. The most abundant
component is epigallocatechin-3-gallate (EGCG) (Fig. 1C), which appears to be the primary
active ingredient responsible for the important biological and
pharmacological properties of green tea.
Cancer chemoprevention by the natural compound,
EGCG, has been studied by many investigators and has attracted much
attention in recent years. Many mechanisms have been proposed for
the antitumor activities of EGCG. These include antioxidant
activities, cell cycle arrest, induction of apoptosis, induction or
inhibition of drug metabolic enzymes, modulation of cell signaling,
inhibition of DNA methylation, effect on miRNA expression, histone
modifications, proteases and telomerases (38,39).
Based on the study of the effects of EGCG in regulating different
key proteins in various signaling pathways, the direct interaction
of proteins with EGCG was found to be a key step in the process of
EGCG-induced effects. The eight phenolic groups of EGCG can serve
as hydrogen bond donors to many biomolecules. The identification of
proteins interacting directly with EGCG is important in
understanding the molecular mechanisms of the effects. Several
proteins that can directly bind with EGCG have been identified and
include 67-kDa laminin receptor, Bcl-2, GRP-78, insulin-like growth
factor-I receptor, G3BP1, ZAP-70, Pin1 and vimentin (40–43).
The inhibitory effects of EGCG against
carcinogenesis at different organ sites have been demonstrated in
many animal models (44,45). The cancer-preventive effects of EGCG
are also widely supported by results from clinical studies
(46). A prospective cohort study
with over 8,000 individuals revealed that the daily consumption of
green tea resulted in delayed cancer onset and a follow-up study of
breast cancer patients found that stage I and II breast cancer
patients experienced a lower recurrence rate and a longer
disease-free period (47).
Moreover, EGCG delivered in the form of green tea extract (GTE) for
12 weeks has been reported to be effective in suppressing oral
premalignant lesions (OPLs), in part through reducing angiogenic
stimulus (stromal VEGF). Higher doses of GTE may improve short-term
(12-week) OPL outcome (48). The
positive results observed in clinical trials along with significant
preclinical results indicate that strategies and the means to take
EGCG from bench to real-life situations are on the horizon.
EGCG also reportedly exhibits antibacterial,
antifungal and antiviral effects (49). Epstein-Barr virus is a human herpes
virus that infects 90% of the human population. EBV infection, as
one of many environmental factors, has been reported to be strongly
associated with the development of several human malignancies,
including Burkitt’s lymphoma, Hodgkin’s disease and NPC. Like all
other herpes viruses, EBV establishes a latent or lytic infection
in host cells. Intriguingly, evidence indicates that EBV
reactivation into the lytic cycle may play a role in the
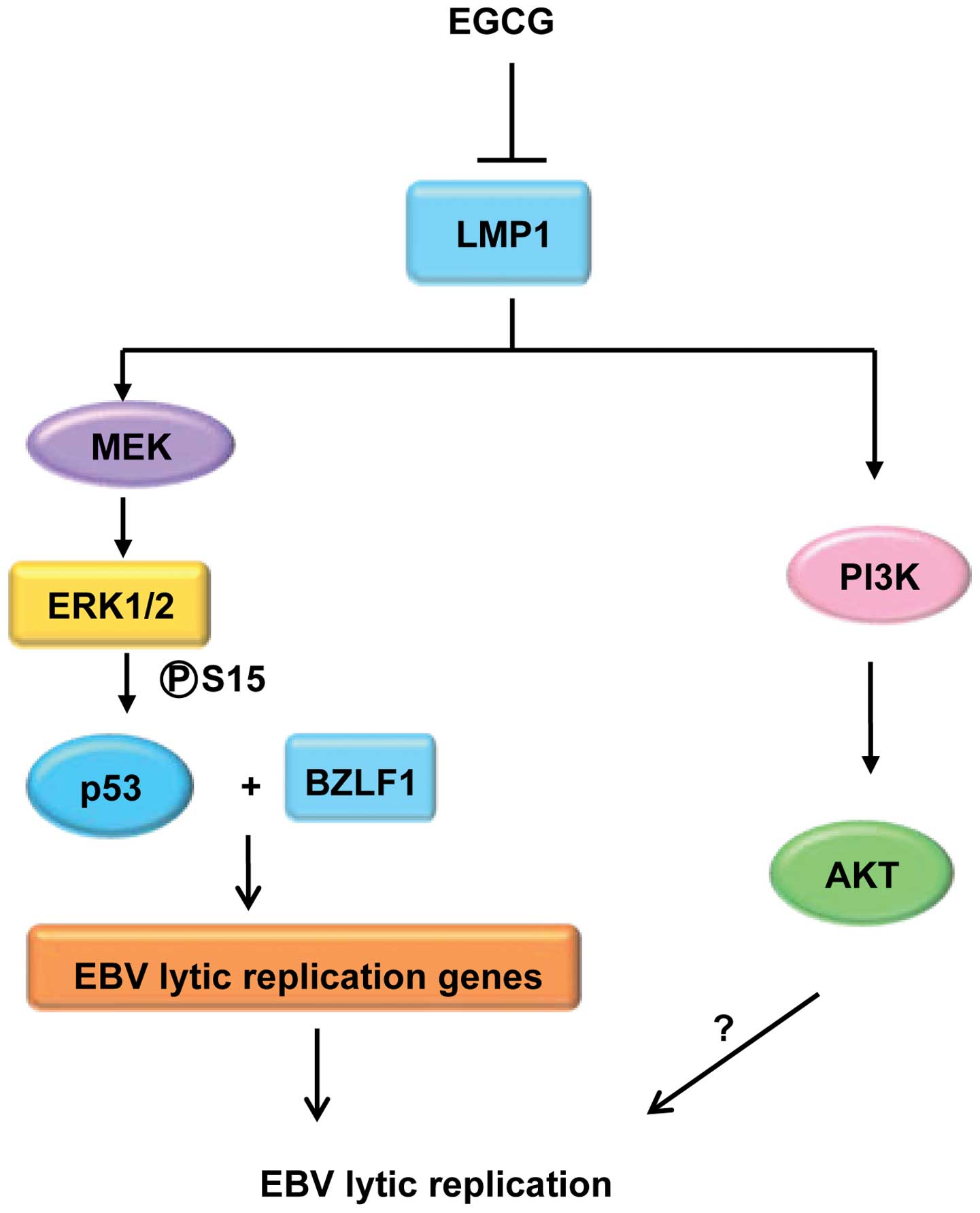
pathogenesis of malignancies. Evidence indicates that the ERK1/2,
MAPK and PI3K/Akt signaling pathways play a critical role in EBV
lytic infection (50,51). Latent membrane protein 1 (LMP1),
which is the only viral gene product with oncogenic properties
among the EBV-encoded proteins, are detected in 90% of NPC
patients. Through the cytoplasmic carboxy terminus, LMP1 triggers
multiple signal transduction cascades, including MEK/ERKs,
PI3K/Akt, JNKs and STAT3, to alter cell growth and survival
(52). Considering that EGCG can
modulate signaling pathways induced by LMP1, other groups and ours
have shown that EGCG modulates multiple signal transduction
pathways including the MAPK and PI3K/Akt pathways, thereby
imparting strong cancer chemopreventive as well as therapeutic
effects (53,54). In our recent study, we investigated
the effects of EGCG on EBV spontaneous lytic infection and the
mechanism involved in EBV-positive cells. We found that EGCG
effectively inhibited the constitutive lytic infection of EBV at
DNA, gene transcription and protein levels by decreasing the
phosphorylation and activation of ERK1/2 and Akt (11). Using signaling pathway-specific
inhibitors, we also explored the signaling mechanisms underlying
the inhibitory effects of EGCG on EBV spontaneous lytic infection
in cell models. The results showed that specific inhibitors of MEK
(PD98059) and PI3K (LY294002) markedly downregulated gene
transcription and expression of BZLf1 and BMRf1, indicating that
the MEK/ERK1/2 and PI3K/Akt pathways are involved in the EBV
spontaneous lytic cycle cascade (11). Therefore, one of the mechanisms by
which EGCG inhibits EBV spontaneous lytic infection appears to
involve the suppression of the activation of MEK/ERK1/2 and
PI3-K/Akt signaling. Since EBV lytic infection plays a critical
role in the development of EBV-associated malignancies, we
described here the potential chemopreventive activity of EGCG in
controlling EBV lytic infection. Our findings support the future
investigation of EGCG as an anticancer and chemoprevention agent
for EBV-associated malignancies (Fig.
4).
5. Conclusions and perspectives
In the present review, we summarize the anticancer
effects of three compounds, grifolin, neoalbaconol and EGCG. These
natural agents show high potential in cancer treatment and
prevention.
In the past few years, our research has revealed
that grifolin significantly increased the binding activity of p53
to the dapk1 gene, and then upregulated DAPK1 mRNA as well as
protein expression in NPC cells. This may be an important mechanism
by which to induce an apoptotic effect in tumor cells. Moreover,
the activation of DAPK1 induced by grifolin enhanced the stability
of p21. This effectively inhibited the activity of the cyclin
D1/CDK4/6 complex, leading to G1 phase arrest in NPC.
The abnormalities of epigenetics occur at an early stage in tumor
development and can be reversed by epigenetic-regulated drugs,
which provides an opportunity for cancer chemoprevention. More
recently, we found that grifolin restores the expression of genes
silenced by modulating various components such as DNA methylation,
suggesting that it acts as a DNMT inhibitor and plays a novel role
in chemoprevention (unpublished data). Together, grifolin may be a
potential leading compound in cancer prevention and therapy.
Metabolic reprogramming has been proven to be
widespread in cancer cells and is regarded as an emerging hallmark
of cancer. Studying and elucidating the relationship of metabolic
disorders and cancer should provide new ideas for molecular
intervention mechanisms and should also help to promote a new field
that can provide metabolism-based targets for cancer patients. Our
research indicated that the PDK1/PI3K/Akt signaling pathway may be
involved in NA-induced apoptotic and necroptotic cell death by
remodeling cellular energy metabolism. The mode of cell death
induced by NA is important since multiple cell death pathway
activation helps to overcome chemotherapy and radiotherapy
resistance. Consequently, induction of apoptosis and alternative
cell death-necroptosis by NA provides therapeutic benefits.
Considering the role of necroptosis in viral infection, NA is a
potential chemopreventive agent for virus-related cancer.
In regards to EGCG, we will further assess its
effect on the lytic replication of EBV in animals and humans.
Infection with EBV is universal and associates with the
pathogenesis of many types of tumors, providing a specific approach
for chemoprevention. Our in vitro experiments showed that
EGCG induces EBV lytic replication, and in vivo experiments
need to be conducted.
The above scientific research was performed on
experimental systems of tumor cells and xenografts in nude mouse
models, while suitable experimental systems of precancerous
conditions are needed to confirm the chemopreventive effects of
these compounds. Meanwhile, water solubility and biological
availability of these compounds need to be improved. Currently,
total synthesis for the two agents, grifolin and neoalbaconol, has
not been achieved. Through the collaboration of chemists, computer
experts and biologists, we hope to obtain the natural compounds by
total synthesis, validate the molecular targets of the compounds
and provide further evidence concerning the anticancer mechanisms
in vitro and in vivo.
Acknowledgments
The authors would like to thank Professor Zigang
Dong and Ann M. Bode for their kind advice. This study was
supported by the National Key Basic Research Program of China (nos.
2009CB522300 and 2011CB504305), the National Natural Science
foundation of China (nos. 81430064 and 81372393), the Hunan
Provincial Innovation foundation for Postgraduates (CX2012B082),
and the Open-End fund for the Valuable and Precision Instruments of
Central South university (CSuZC201541).
References
|
1
|
Cao Y, DePinho RA, Ernst M and Vousden K:
Cancer research: Past, present and future. Nat Rev Cancer.
11:749–754. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Harvey A: The role of natural products in
drug discovery and development in the new millennium. IDrugs.
13:70–72. 2010.PubMed/NCBI
|
|
3
|
Amin AR, Kucuk O, Khuri FR and Shin DM:
Perspectives for cancer prevention with natural compounds. J Clin
Oncol. 27:2712–2725. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sarkar FH, Li Y, Wang Z and Kong D:
Cellular signaling perturbation by natural products. Cell Signal.
21:1541–1547. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ye M, Liu JK, Lu ZX, Zhao Y, Liu SF, Li
LL, Tan M, Weng XX, Li W and Cao Y: Grifolin, a potential antitumor
natural product from the mushroom Albatrellus confluens, inhibits
tumor cell growth by inducing apoptosis in vitro. FEBS Lett.
579:3437–3443. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ye M, Luo X, Li L, Shi Y, Tan M, Weng X,
Li W, Liu J and Cao Y: Grifolin, a potential antitumor natural
product from the mushroom Albatrellus confluens, induces cell-cycle
arrest in G1 phase via the ERK1/2 pathway. Cancer Lett.
258:199–207. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Luo XJ, Li LL, Deng QP, Yu XF, Yang LF,
Luo FJ, Xiao LB, Chen XY, Ye M, Liu JK, et al: Grifolin, a potent
antitumour natural product upregulates death-associated protein
kinase 1 DAPK1 via p53 in nasopharyngeal carcinoma cells. Eur J
Cancer. 47:316–325. 2011. View Article : Google Scholar
|
|
9
|
Luo XJ, Li W, Yang LF, Yu XF, Xiao LB,
Tang M, Dong X, Deng QP, Bode AM, Liu JK, et al: DAPK1 mediates the
G1 phase arrest in human nasopharyngeal carcinoma cells induced by
grifolin, a potential antitumor natural product. Eur J Pharmacol.
670:427–434. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Deng Q, Yu X, Xiao L, Hu Z, Luo X, Tao Y,
Yang L, Liu X, Chen H, Ding Z, et al: Neoalbaconol induces energy
depletion and multiple cell death in cancer cells by targeting
PDK1-PI3-K/Akt signaling pathway. Cell Death Dis. 4:e8042013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu S, Li H, Chen L, Yang L, Li L, Tao Y,
Li W, Li Z, Liu H, Tang M, et al: (-)-Epigallocatechin-3-gallate
inhibition of Epstein- Barr virus spontaneous lytic infection
involves ERK1/2 and PI3-K/Akt signaling in EBV-positive cells.
Carcinogenesis. 34:627–637. 2013. View Article : Google Scholar
|
|
12
|
Song J, Manir MM and Moon SS: Cytotoxic
grifolin derivatives isolated from the wild mushroom Boletus
pseudocalopus (Basidiomycetes). Chem Biodivers. 6:1435–1442. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hara T, Hirasawa A, Sun Q, Sadakane K,
Itsubo C, Iga T, Adachi T, Koshimizu TA, Hashimoto T, Asakawa Y, et
al: Novel selective ligands for free fatty acid receptors GPR120
and GPR40. Naunyn Schmiedebergs Arch Pharmacol. 380:247–255. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jin S, Pang RP, Shen JN, Huang G, Wang J
and Zhou JG: Grifolin induces apoptosis via inhibition of PI3K/AKT
signalling pathway in human osteosarcoma cells. Apoptosis.
12:1317–1326. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Quang DN, Hashimoto T, Arakawa Y, Kohchi
C, Nishizawa T, Soma G and Asakawa Y: Grifolin derivatives from
Albatrellus caeruleoporus, new inhibitors of nitric oxide
production in RAW 264. 7 cells. Bioorg Med Chem. 14:164–168. 2006.
View Article : Google Scholar
|
|
16
|
Pelled D, Raveh T, Riebeling C, Fridkin M,
Berissi H, Futerman AH and Kimchi A: Death-associated protein (DAP)
kinase plays a central role in ceramide-induced apoptosis in
cultured hippocampal neurons. J Biol Chem. 277:1957–1961. 2002.
View Article : Google Scholar
|
|
17
|
Gozuacik D and Kimchi A: DAPk protein
family and cancer. Autophagy. 2:74–79. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Raval A, Tanner SM, Byrd JC, Angerman EB,
Perko JD, Chen SS, Hackanson B, Grever MR, Lucas DM, Matkovic JJ,
et al: Downregulation of death-associated protein kinase 1 (DAPK1)
in chronic lymphocytic leukemia. Cell. 129:879–890. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hupp TR, Sparks A and Lane DP: Small
peptides activate the latent sequence-specific DNA binding function
of p53. Cell. 83:237–245. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Martoriati A, Doumont G, Alcalay M,
Bellefroid E, Pelicci PG and Marine JC: dapk1, encoding an
activator of a p19ARF- p53-mediated apoptotic
checkpoint, is a transcription target of p53. Oncogene.
24:1461–1466. 2005. View Article : Google Scholar
|
|
21
|
Nichols NM and Matthews KS: Human p53
phosphorylation mimic, S392E, increases nonspecific DNA affinity
and thermal stability. Biochemistry. 41:170–178. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
MacCorkle RA and Tan TH: Mitogen-activated
protein kinases in cell-cycle control. Cell Biochem Biophys.
43:451–461. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Llambi F, Lourenço FC, Gozuacik D, Guix C,
Pays L, Del Rio G, Kimchi A and Mehlen P: The dependence receptor
UNC5H2 mediates apoptosis through DAP-kinase. EMBO J. 24:1192–1201.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shang T, Joseph J, Hillard CJ and
Kalyanaraman B: Death- associated protein kinase as a sensor of
mitochondrial membrane potential: Role of lysosome in mitochondrial
toxin-induced cell death. J Biol Chem. 280:34644–34653. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen X, Chi Y, Bloecher A, Aebersold R,
Clurman BE and Roberts JM: N-acetylation and ubiquitin-independent
proteasomal degradation of p21Cip1. Mol Cell.
16:839–847. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bloom J, Amador V, Bartolini F, DeMartino
G and Pagano M: Proteasome-mediated degradation of p21 via
N-terminal ubiquitinylation. Cell. 115:71–82. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen CH, Wang WJ, Kuo JC, Tsai HC, Lin JR,
Chang ZF and Chen RH: Bidirectional signals transduced by DAPK-ERK
interaction promote the apoptotic effect of DAPK. EMBO J.
24:294–304. 2005. View Article : Google Scholar :
|
|
28
|
Hellwig V, Nopper R, Mauler F, Freitag J,
Liu J-K, Ding Z-H and Stadler M: Activities of prenylphenol
derivatives from fruitbodies of Albatrellus spp. on the human and
rat vanilloid receptor 1 (VR1) and characterisation of the novel
natural product, confluentin. Arch Pharm. 336:119–126. 2003.
View Article : Google Scholar
|
|
29
|
Liu Q, Shu X, Wang L, Sun A, Liu J and Cao
X: Albaconol, a plant-derived small molecule, inhibits macrophage
function by suppressing NF-kappaB activation and enhancing SOCS1
expression. Cell Mol Immunol. 5:271–278. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fuchs Y and Steller H: Programmed cell
death in animal development and disease. Cell. 147:742–758. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Galluzzi L, Vitale I, Abrams JM, Alnemri
ES, Baehrecke EH, Blagosklonny MV, Dawson TM, Dawson VL, El-Deiry
WS, Fulda S, et al: Molecular definitions of cell death
subroutines: Recommendations of the Nomenclature Committee on Cell
Death 2012. Cell Death Differ. 19:107–120. 2012. View Article : Google Scholar :
|
|
32
|
Degterev A, Huang Z, Boyce M, Li Y, Jagtap
P, Mizushima N, Cuny GD, Mitchison TJ, Moskowitz MA and Yuan J:
Chemical inhibitor of nonapoptotic cell death with therapeutic
potential for ischemic brain injury. Nat Chem Biol. 1:112–119.
2005. View Article : Google Scholar
|
|
33
|
Wu YT, Tan HL, Huang Q, Kim YS, Pan N, Ong
WY, Liu ZG, Ong CN and Shen HM: Autophagy plays a protective role
during zVAD-induced necrotic cell death. Autophagy. 4:457–466.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bell BD, Leverrier S, Weist BM, Newton RH,
Arechiga AF, Luhrs KA, Morrissette NS and Walsh CM: FADD and
caspase-8 control the outcome of autophagic signaling in
proliferating T cells. Proc Natl Acad Sci USA. 105:16677–16682.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Vivanco I and Sawyers CL: The
phosphatidylinositol 3-kinase AKT pathway in human cancer. Nat Rev
Cancer. 2:489–501. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Toker A and Newton AC: Cellular signaling:
Pivoting around PDK-1. Cell. 103:185–188. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Peifer C and Alessi DR: Small-molecule
inhibitors of PDK1. Chem Med Chem. 3:1810–1838. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yang CS, Wang H, Li GX, Yang Z, Guan F and
Jin H: Cancer prevention by tea: Evidence from laboratory studies.
Pharmacol Res. 64:113–122. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Link A, Balaguer F and Goel A: Cancer
chemoprevention by dietary polyphenols: Promising role for
epigenetics. Biochem Pharmacol. 80:1771–1792. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Leone M, Zhai D, Sareth S, Kitada S, Reed
JC and Pellecchia M: Cancer prevention by tea polyphenols is linked
to their direct inhibition of antiapoptotic Bcl-2-family proteins.
Cancer Res. 63:8118–8121. 2003.PubMed/NCBI
|
|
41
|
Li M, He Z, Ermakova S, Zheng D, Tang F,
Cho YY, Zhu F, Ma WY, Sham Y, Rogozin EA, et al: Direct inhibition
of insulin-like growth factor-I receptor kinase activity by
(-)-epigal-locatechin-3-gallate regulates cell transformation.
Cancer Epidemiol Biomarkers Prev. 16:598–605. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Bode AM and Dong Z: Epigallocatechin
3-gallate and green tea catechins: united they work, divided they
fail. Cancer Prev Res. 2:514–517. 2009. View Article : Google Scholar
|
|
43
|
Urusova DV, Shim JH, Kim DJ, Jung SK,
Zykova TA, Carper A, Bode AM and Dong Z: Epigallocatechin-gallate
suppresses tumorigenesis by directly targeting Pin1. Cancer Prev
Res. 4:1366–1377. 2011. View Article : Google Scholar
|
|
44
|
Ramos S: Cancer chemoprevention and
chemotherapy: Dietary polyphenols and signalling pathways. Mol Nutr
food Res. 52:507–526. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kim JW, Amin AR and Shin DM:
Chemoprevention of head and neck cancer with green tea polyphenols.
Cancer Prev Res. 3:900–909. 2010. View Article : Google Scholar
|
|
46
|
Yang CS, Wang X, Lu G and Picinich SC:
Cancer prevention by tea: Animal studies, molecular mechanisms and
human relevance. Nat Rev Cancer. 9:429–439. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Singh BN, Shankar S and Srivastava RK:
Green tea catechin, epigallocatechin-3-gallate (EGCG): Mechanisms,
perspectives and clinical applications. Biochem Pharmacol.
82:1807–1821. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Tsao AS, Liu D, Martin J, Tang XM, Lee JJ,
El-Naggar AK, Wistuba I, Culotta KS, Mao L, Gillenwater A, et al:
Phase II randomized, placebo-controlled trial of green tea extract
in patients with high-risk oral premalignant lesions. Cancer Prev
Res. 2:931–941. 2009. View Article : Google Scholar
|
|
49
|
Steinmann J, Buer J, Pietschmann T and
Steinmann E: Anti- infective properties of
epigallocatechin-3-gallate (EGCG), a component of green tea. Br J
Pharmacol. 168:1059–1073. 2013. View Article : Google Scholar :
|
|
50
|
Satoh T, Hoshikawa Y, Satoh Y, Kurata T
and Sairenji T: The interaction of mitogen-activated protein
kinases to Epstein-Barr virus activation in Akata cells. Virus
Genes. 18:57–64. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Darr CD, Mauser A and Kenney S:
Epstein-Barr virus immediate-early protein BRLF1 induces the lytic
form of viral replication through a mechanism involving
phosphatidylino-sitol-3 kinase activation. J Virol. 75:6135–6142.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Zheng H, Li LL, Hu DS, Deng XY and Cao Y:
Role of Epstein-Barr virus encoded latent membrane protein 1 in the
carcinogenesis of nasopharyngeal carcinoma. Cell Mol Immunol.
4:185–196. 2007.PubMed/NCBI
|
|
53
|
Zhao Y, Wang H, Zhao XR, Luo FJ, Tang M
and Cao Y: Epigallocatechin-3-gallate interferes with EBV-encoding
AP-1 signal transduction pathway. Zhonghua Zhong Liu Za Zhi.
26:393–397. 2004.In Chinese. PubMed/NCBI
|
|
54
|
Kanwar J, Taskeen M, Mohammad I, Huo C,
Chan TH and Dou QP: Recent advances on tea polyphenols. Front
Biosci. 4:111–131. 2012. View
Article : Google Scholar
|