Introduction
Skin cancer has become a prevalent social and public
health issue due to the fact that the incidence and mortality rates
of skin cancer, including melanoma and non-melanoma skin cancers
(NMSCs), consisting of basal cell cardinoma (BCC) and squamous cell
carcinoma (SCC) are rising in many countries (1,2). Skin
cancer patients are associated with higher UV radiation exposure
(3) and UVB susceptibility was
reported to be significantly higher in skin cancer patients
(4). In general, UV irradiation
induces DNA damage, repair, skin inflammation and sunburn, gene
mutation, post-inflammatory immunosuppression and eventually
oncogenic pathway activity and is considered to be a major
aetiological factor for skin carcinogenesis (5,6).
UVB exposure induces cutaneous inflammation. A
single exposure to UVB may result in an acute inflammatory
response, which is characterized by erythema (redness) and edema
(swelling) due to increased vascular flow and vascular permeability
(7,8). An inflammatory microenvironment aids
in the proliferation and survival of malignant cells, promotes
angiogenesis and metastasis, subverts adaptive immune responses and
alters responses to hormones and chemotherapeutic agents, all of
which promote the development of tumors (9,10).
Inflammation is always accompanied by oxidative stress induced by
reactive oxygen species (ROS) accumulation. ROS are believed to
activate proliferative and cell survival signaling that alter
apoptotic pathways that may be involved in the pathogenesis of a
number of skin disorders. ROS act largely by driving several
important molecular pathways that play important roles in diverse
pathologic processes including inflammatory responses (11). Inflammation and ROS are inseparable
and interact in the skin cancer process.
However, UV irradiation in sunlight is the most
ubiquitous physical substance in our natural environment. Low-dose
UVB induces non-inflammation and normal non-inflammatory apoptosis
(12), which is a physiological
process and conserved feature for the non-inflammatory removal of
cells in order to protect organisms (13). Low-dose UV irradiation is also well
known to be the primary source of Vitamin D3 synthesis, which acts
as a sunscreen to prevent skin and various other types of cancer
(14–16). Skin cancer, in the United States, is
reported to account for ~20 to 30% of all neoplasms in Caucasians,
2 to 4% of all neoplasms in Asians and 1 to 2% of all neoplasms in
Blacks and Asian Indians (17–19).
This fact indicates that skin cancer is less common in persons with
skin color other than light-skin. Thus, melanin pigment is a UV
screen that protects some deleterious sunlight compositions
(20,21). Although the skin coloration in
dark-skinned ethnic groups is the result of long-term evolution
based on resistance and prevention of UVB irradiation-induced skin
injury (22,23), in the short term, skin coloration
namely pigmentation is due to solar irradiation. These studies
indicate that a low dose of UV irradiation plays a protective role
in skin cancer.
However, skin cancer still presents greater
morbidity and mortality in colored-skinned individuals (17). It appears that the morbidity and
mortality of skin cancer is not paralleled with incidence, since
UVB irradiation depletes heavily pigmented skin of Langerhans cells
(20). Once limited pigmented skin
is depleted, the skin cancerization rate greatly increases.
Therefore, the skin cancer protective role of melanin pigment is
limited or UV-relative, but not an absolute prevention for skin
cancer.
Collectively although excess UV irradiation
certainly induces skin cancer, low-dose UV irradiation exhibits a
protective role to prevent skin cancer yet is limited: i) to the
most initial stage of cutaneous lesions or ii) to relative slight
cutaneous lesions. However, we do not know how low-dose UV
irradiation prevents these cutaneous lesions.
In the present study, we performed low-dose UVB
pre-irradiation and then subsequent medium- or high-dose UVB
irradiation. We found that low-dose UVB pre-irradiation inhibited
medium-dose-induced skin inflammation, ROS production and initial
keratinocyte hypertrophy while high-dose UVB irradiation did not
induce change. This inhibition was associated with the balance of
MMP2 and its inhibitor.
Materials and methods
Mice and UV irradiation
In the present experiment C57BL/6N mice were used
and all the mice were purchased from Biocytogen Co., Ltd. (Beijing,
China). The dorsal hair was removed by a depilatory in an area of 4
cm2, and all mice were divided into 3 UV irradiation
groups and 1 control group (n=8/group). The UV irradiation groups
were exposed to low-, medium- and high-dose (100, 350 and 800
mJ/cm2) UV irradiation (FL20S. E; Toshiba, Tokyo, Japan)
for 5 min/day, 5 days every week for 4 weeks from the age of 8
weeks. Low-dose pre-irradiation was performed from the age of 6
weeks and then formal irradiation was carried out. All mice were
fed normal rodent chow and had ad libitum access to water.
The present study was approved by the Ethics Committee of the
Harbin Medical University, Harbin, China.
Cell culture and detection of
intracellular ROS
Intracellular accumulation of ROS was estimated
using the fluorescent dye H2-DCFDA (Sigma-Aldrich,
Shanghai, China), which is converted to a membrane-imper meable and
highly fluorescent compound, dichlorofluorescein diacetate (DCF),
in cells in the presence of ROS (24). The epidermal keratinocyte cell line
HacaT (Biomics Biotech, Jiangsu, China) was used in the in
vitro experiment as a source of keratinocytes. Cells were
cultured in complete DMEM (L-glutamine, pyridoxine hydrochloride,
sodium pyruvate and FCS) at 37°C in a 5% CO2/95% air
humidified atmosphere. UV irradiation groups were exposed to low-,
medium- and high-dose (10, 35 and 80 mJ/cm2) UV
irradiation as reported previously (12). After three days of low-dose UV
irradiation, an additional medium- or high-dose UVB irradiation was
re-irradiated for 30 min. Subsequently, the cells were collected
with 0.25% Trypsin and analyzed using a fluorescence
spectrophotometer (F-2500; Hitachi, Tokyo, Japan) to detect the
fluorescence of DCF inside the cells (excitation wavelength, 488
nm; emission wavelength, 521 nm).
RT-PCR analysis
Total RNA was extracted from exposed dorsal skin
tissue and relative mRNA was normalized to 18s rRNA. The primers
used are listed in Table I.
 | Table IThe primers used for the RT-PCR
analysis. |
Table I
The primers used for the RT-PCR
analysis.
| IL-1α | F: |
5′-GAAGAGACGGCTGAGTTTCAGTG-3′ |
| R: |
5′-CTGGTAGGTGTAAGGTGCTGA-3′ |
| IL-1β | F: |
5′-TCATTGTGGCTGTGGAGAAG-3′ |
| R: |
5′-CAGTTGTCTAATGGGAACGT-3′ |
| IL-6 | F: |
5′-ACAACCACGGCCTTCCCTACTT-3′ |
| R: |
5′-CACGATTTCCCAGAGAACATGTG-3′ |
| SOD1 | F: |
5′-ACTAGTATGGCGATGAAAGCGGTG-3′ |
| R: |
5′-GGATCCTGTTTACTGGGCAATCCC-3′ |
| Catalase | F: |
5′-ATGTCGGACAGTCGGGAC-3′ |
| R: |
5′-GACTGTGGAGAACCGAAC-3′ |
| 18s | F: |
5′-GTAACCCGTTGAACCCCATT-3′ |
| R: |
5′-CCATCCAATCGGTAGTAGCG-3′ |
Western blot analysis
A vertical slab gel with 12% polyacrylamide was used
for protein electrophoresis. The transfer of proteins from the
SDS-PAGE gels to the membrane was performed electrophoretically
with some modifications using a Semi Dry Electroblotter (Sartorius)
for 90 min with an electric current of 14 V. Each membrane was
treated with Block Ace™ (2.5%) for 60 min at 22°C. The first
immunological reaction was performed using a specific primary mouse
monoclonal IgG 4HNE antibody (Abnova, Taipei, Taiwan, China),
rabbit IgG MMP2 antibody (Proteintech Group Inc., Wuhan, China),
mouse monoclonal IgG TIMP2 and TIMP4 antibodies (Abcam Trading Co.
Ltd., Shanghai, China) in PBS containing 0.03% Tween-20 overnight
at 4°C. The second reaction was performed using HRP-conjugated
anti-rabbit IgG for 1 h at 22°C. The ECL Plus Western Blotting
Detection System™ was used for enhanced chemiluminescence (ECL)
detection of the protein bands.
SOD activity assay
SOD activity assay was performed as previously
described (25). In brief, skin
tissues were homogenized in cold buffer (0.1 M Tris/HCl, pH 7.4
containing 0.5% Triton X-100, 5 mM β-ME and 0.1 mg/ml PMSF) and
homogenates were centrifuged at 14,000 × g for 15 min at 4°C. The
resulting supernatant that contained cytosolic SOD was subjected to
an SOD activity assay with a commercial SOD activity assay kit
(Biovision, Palo Alto, CA, USA).
Evaluation of keratinocyte
hyperplasia
Dorsal skin tissues were excised 24 h after the last
irradiation, and each harvested tissue sample was fixed in formalin
and embedded in paraffin. Each sample was then cut into a section
at 0.05-mm intervals. Each section was then stained with H&E
staining. The thickness of the skin tissue keratinocytes was
measured by SigmaPlot (Systat Software Inc. Chicago, IL, USA), and
the average was calculated.
Gelatin zymography
To measure MMP2 activity, an extract of the
supernatant of the skin tissue was collected. The gelatin
zymography kit (Yagai Corporation, Tokyo, Japan) was used according
to the manufacturer's instructions to perform gelatin zymography.
In brief, 15 μl samples of conditioned supernatant were
subjected to electrophoresis through gels provided in the gelatin
zymography kit. Each gel was washed twice, once with each type of
washing buffer for 30 min and then incubated for 40 h at 37°C in
the reaction buffer. Each gel was then stained with Coomassie blue
and then destained three times so that the protein bands with
gelatinolytic activity were easily identified.
Statistical analysis
Data are expressed as means ± SEM of triplicate
runs. Each experiment was repeated as least twice. The Student's
t-test and analysis of variants were used to assess differences and
P<0.05 was considered to be significant.
Results
Effects of the different doses of UV
irradiation on skin inflammation and ROS production
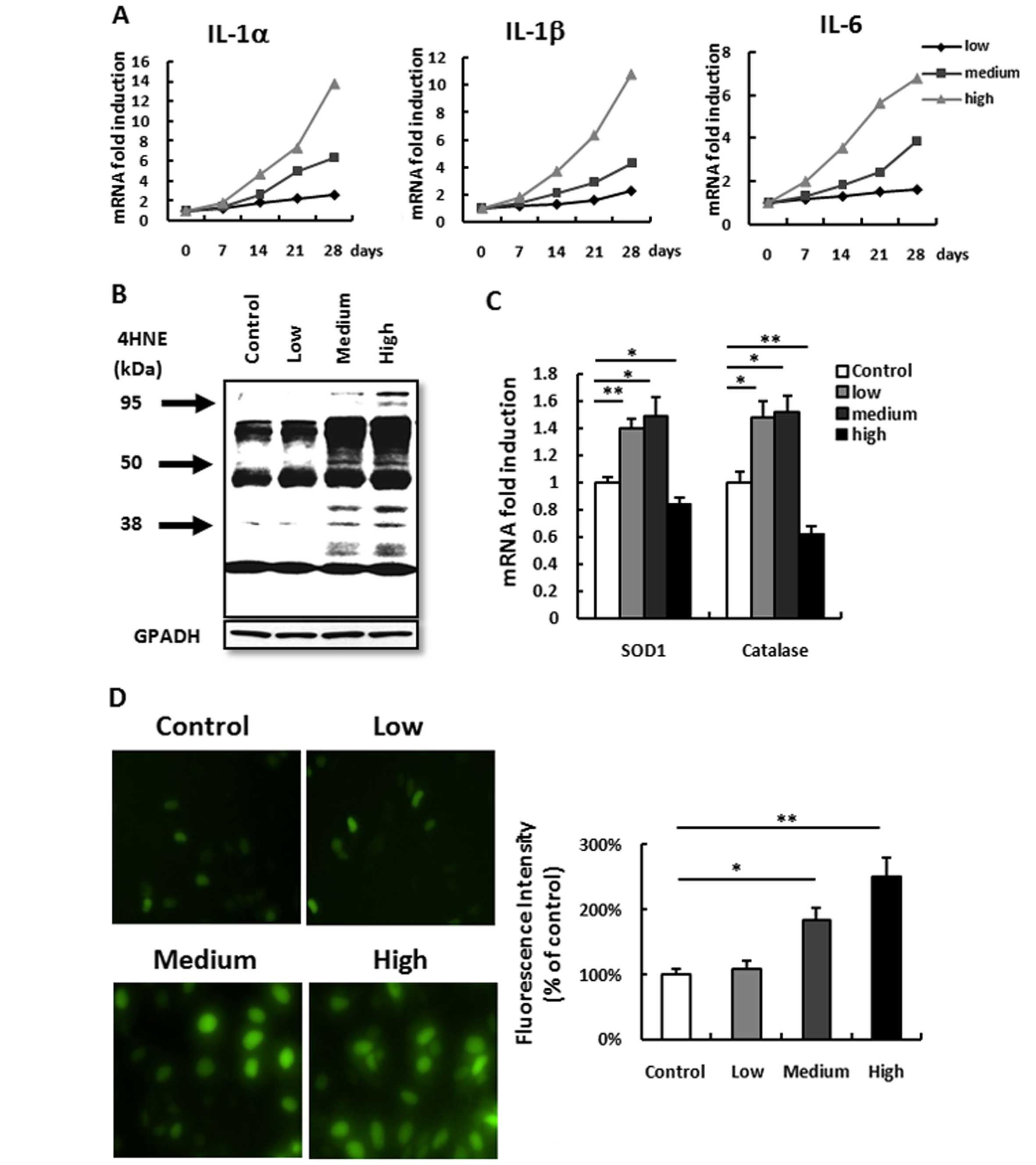
In order to confirm the role of various doses of UV
irradiation on inflammation and ROS, we detected the time course of
various classic inflammatory factors IL-1α, IL-1β and IL-6 mRNA
from mouse dorsal skin tissues exposed to the three doses of UV
irradiation. It was observed that low-dose irradiation did not
induce any inflammation after a 4-week exposure compared to that
before irradiation. A medium- or a high-dose irradiation induced a
very significant gradual increase compared to that before
irradiation (Fig. 1A). ROS
accumulation detected by 4HNE also showed a similar result.
Low-dose irradiation barely induced any ROS accumulation, while
medium- or high-dose irradiation induced a significant increase in
ROS accumulation (Fig. 1B). This
result is consistent with the results of the ROS accumulation
detection in the HacaT keratinocyte cell line (Fig. 1D). However, different from
inflammation, the difference in ROS accumulation induced between
medium- and high-dose irradiation was not significant (Fig. 1B). This result forced us to consider
that the imbalance of ROS accumulation and anti-ROS may be the
reason for the difference (26).
Thus, we next detected the anti-ROS enzyme SOD1 and catalase. Both
were increased in the low- and medium-dose irradiation groups but
decreased in the high-dose irradiation group (Fig. 1C). These results indicate that
although skin inflammation and ROS accumulation were correlated
with the dose of UV irradiation, low-dose UV induced no
inflammation and no ROS but increased anti-ROS ability, a potential
protect effect.
Low-dose pre-irradiation protects skin
epithelium against medium-dose UV irradiation-induced
hyperplasia
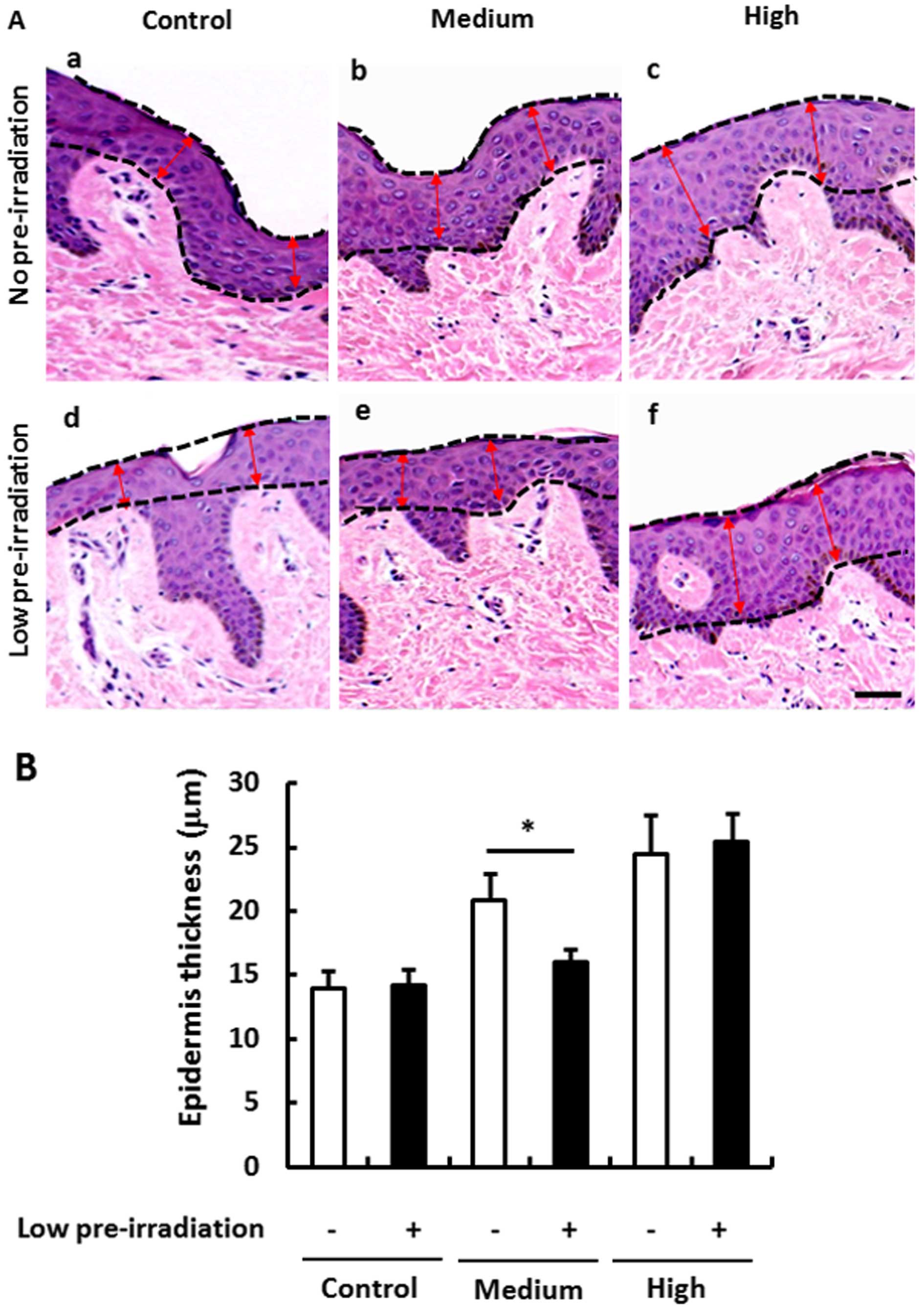
Since low-dose irradiation presented a potential
protective role, we aimed to ascertain whether low-dose UV
irradiation may also suppress the medium- and high-dose induced
skin epithelial proliferation and hypertrophy. Firstly, we
confirmed that the skin epithelial hypertrophy was dependent on the
dose of UV irradiation (Fig. 2Aa–c and
B), while low-dose UV irradiation did not induce any
hypertrophy compared with the no pre-irradiation group (Fig. 2Aa and d and B). However, compared to
the medium-dose UV irradiation-induced epithelial hypertrophy
(Fig. 2Aa and b and B), we found
that medium-dose UV irradiation did not induce hypertrophy even in
the low-dose pre-irradiation group (Fig. 2Ad and e and B). The inhibitory role
was not observed when a high-dose UV irradiation was performed
(Fig. 2Ac and f and B). These
results indicated an inhibitory role of low-dose UV irradiation on
medium-dose UV irradiation-induced epithelial hypertrophy, but not
on high-dose UV irradiation-induced hypertrophy.
Low-dose pre-irradiation suppresses
medium-dose UV irradiation-induced inflammation and ROS
accumulation
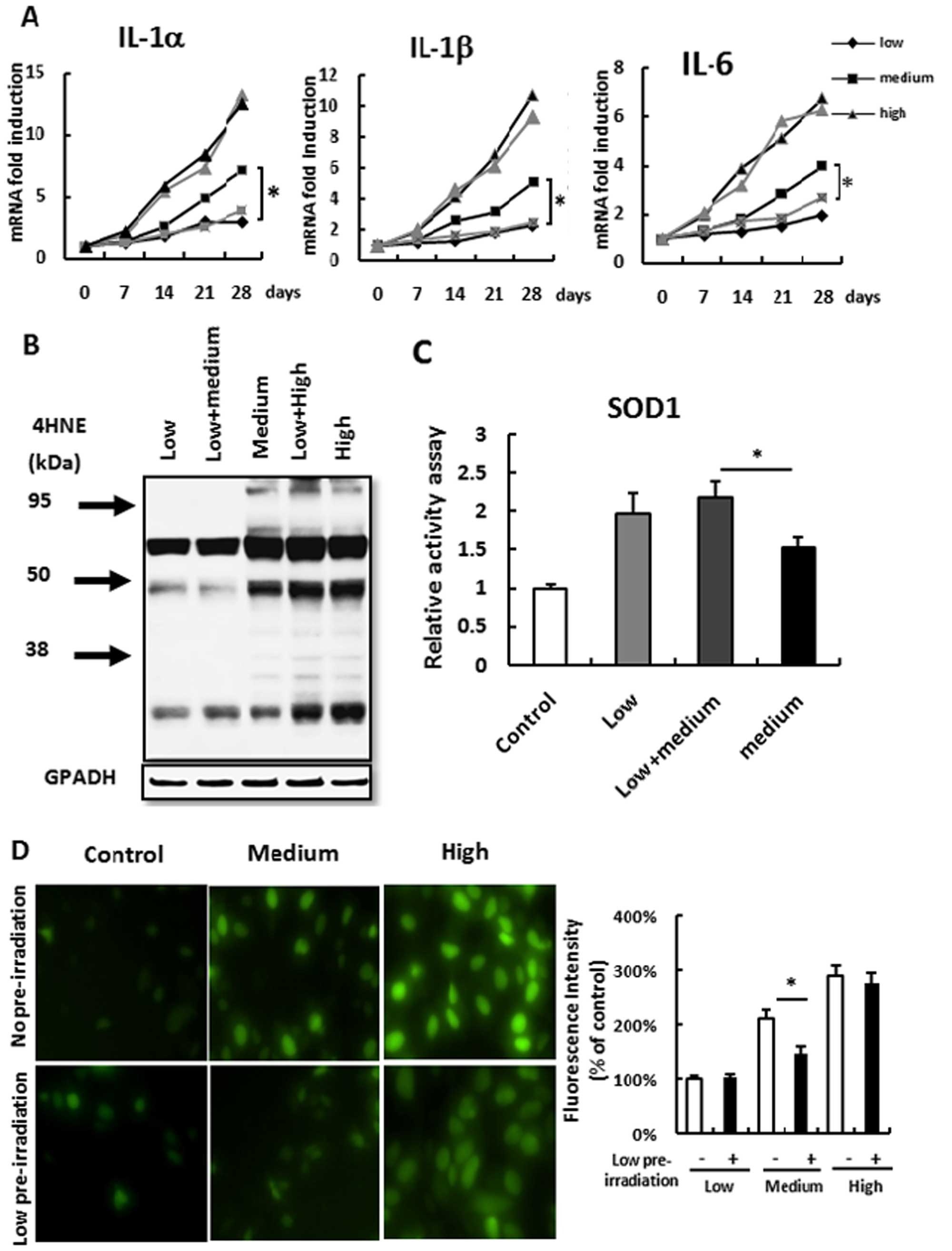
As skin inflammation and ROS may lead to epithelial
proliferation and even skin tumors (27) and low-dose pre-irradiation protected
skin epithelial proliferation, we aimed to ascertain whether
low-dose pre-irradiation suppresses UV irradiation-induced
inflammation and ROS. All the mice were administered a medium- or
high-dose re-irradiation after a low-dose UV pre-irradiation. We
found that low-dose pre-irradiation suppressed medium-dose
irradiation-induced inflammation but not high-dose
irradiation-induced inflammation (Fig.
3A). Paralleled with this result, low-dose irradiation
suppressed medium-dose irradiation-induced ROS accumulation but not
high-dose irradiation-induced ROS accumulation (Fig. 3B). The same result was also found in
the HacaT keratinocyte cell line culture. The medium-dose UV
irradiated cells after low-dose pre-irradiation presented
significant low fluorescence intensity compared with that in the
group with no pre-irradiation, but this suppression was not found
in the high-dose irradiation group (Fig. 3D).
Next, we aimed to ascertain why low-dose UV
irradiation suppresses medium-dose irradiation-induced ROS
accumulation. Although medium-dose irradiation induced anti-ROS
SOD1 expression (Fig. 1C),
medium-dose irradiation did not suppress ROS production (Fig. 1B). These results suggest that SOD1
activity was not enough to suppress extra ROS production. Thus, we
measured SOD1 activity in skin tissue after irradiation. Confirming
our hypothesis, SOD1 activity was decreased after medium-dose
irradiation when compared with low-dose irradiation, but low-dose
pre-irradiation induced elevated SOD1 activity and prevented SOD1
activity loss induced by medium-dose irradiation (Fig. 3C). These results indicated that
low-dose irradiation may elevate SOD1 activation to suppress
medium-dose irradiation-induced ROS production and
inflammation.
Low-dose pre-irradiation increases TIMP2
to inhibit MMP2 expression
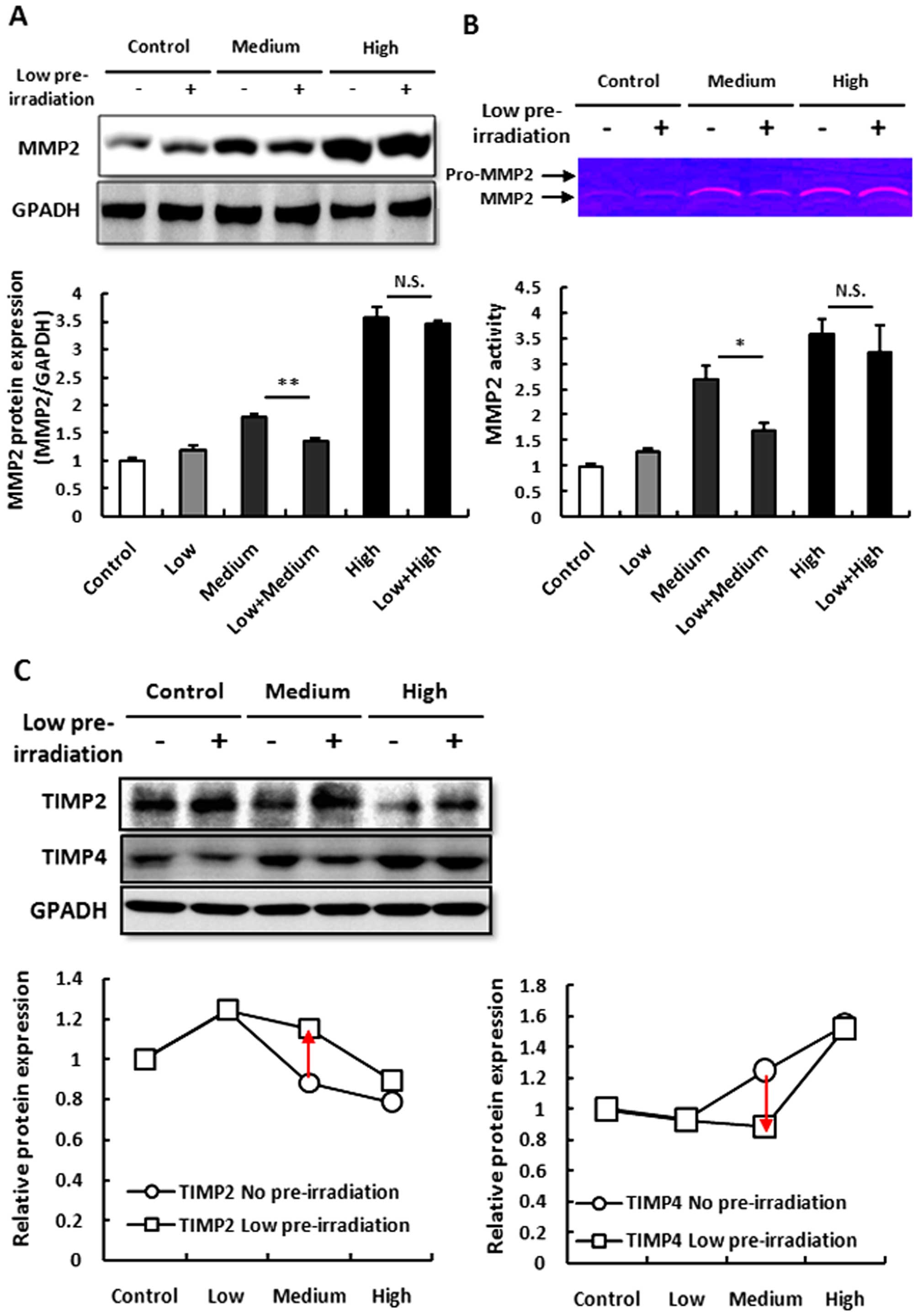
Although low-dose irradiation suppressed medium-dose
UV irradiation-induced inflammation and ROS accumulation, which are
the causes of skin epithelial hyperplasia, we did not know whether
low-dose irradiation also affects MMP2 and TIMPs which are the
direct mechanisms of skin epithelial hyperplasia (28). We found that low-dose irradiation
did not affects the MMP2 expression and its activation, however,
low-dose irradiation inhibited the increase in MMP2 induced by
medium- but not high-dose irradiation (Fig. 4A and B).
Next, we aimed to ascertain how MMP2 inhibitor,
TIMP2 and TIMP4, are affected after low-dose irradiation. We found
that TIMP2 was increased following single low-dose irradiation and
then decreased following medium- and high-dose irradiation and
TIMP4 was not significant altered following low-dose irradiation
but increased following medium- and high-dose irradiation. However,
after low-dose pre-irradiation, TIMP2 was highly expressed in the
medium-dose irradiation group and TIMP4 was decreased. Both TIMP2
and TIMP4 were not significantly altered following high-dose
irradiation (Fig. 4C). These
results indicated that low-dose irradiation may elevate TIMP2 to
inhibit MMP2 expression and activation but not TIMP4.
Discussion
In the present study, we demonstrated that a
relative non-inflammation and non-ROS-inducing low-dose UV
irradiation is a protective factor to suppress slight inflammation
and ROS accumulation, which is induced by a medium-dose UV
irradiation, further suppressing slight skin epithelial
hyperplasia. This may be contributed to elevated SOD1 activity and
suppressed skin epithelial MMP2 activity inhibited by increased
TIMP2 activity. However, this type of suppression was not observed
in high-dose UV irradiation-induced inflammation, ROS accumulation
and skin epithelial hyperplasia with MMP2 activity. These results
indicate that the protective role of low-dose UV irradiation is
only limited to the most initial stage and for very slight
lesions.
A relative low-dose UV irradiation induced no
inflammation or ROS production (Fig. 1A
and B), that was contributed to elevated anti-ROS enzymes
(Fig. 1C). Although medium-dose UV
irradiation also induced the same level of anti-ROS enzymes, which
may not suppress the continuous accumulation of ROS. But with the
increased intensity of UV irradiation, the anti-ROS system was
absolutely damaged after high-dose UV irradiation (Fig. 1B and C). This indicates the dual
character of UV irradiation on the ROS-antioxidant system (29,30).
It is very easy for us to understand how ROS are portrayed as
detrimental, as evidenced by the notable trend in the use of
dietary and cosmetic antioxidants (31). However, recently, it was reported
that increased ROS promote longevity and metabolic health, which
may be explained by mitochondrial hormesis (mitohormesis) (32,33).
ROS, derived from the mitochondrial electron transport system, may
be necessary triggering elements for a sequence of events that
result in benefits ranging from the transiently cytoprotective to
organismal-level longevity. Basis on this concept, it is not
difficult to understand that ROS also play an important role in the
beneficial alterations in cellular physiology produced by caloric
restriction, intermittent fasting, exercise and dietary
phytonutrients (34). Thus, the
balance of ROS and antioxidants seems to be a key element (30).
As the dual character of ROS, in the present study
was stimulated by UV irradiation and in light of the importance of
the ROS-antioxidant balance, it is difficult to judge whether it is
beneficial or not for health. However, it seemed that low
accumulation of ROS induced by low-dose UV irradiation, although
the low accumulation of ROS was not suppressed by elevated
antioxidants (Fig. 1C and D),
inhibited further inflammation and further ROS accumulation induced
by medium-dose UV irradiation (Fig. 3A,
B and D). In the present study, there was an interesting
phenomenon displayed. Low-dose UV irradiation inhibited medium-dose
UV irradiation. It may be explained that an accumulation of good
ROS inhibited the accumulation of bad ROS or the increasing
antioxidants (SOD and catalase) inhibited further ROS accumulation
(35). We also found another
interesting phenomenon that both low- and medium-dose UV
irradiation induced the same level of SOD expression (Fig. 1C). However, low-dose UV irradiation
induced SOD-suppressed ROS accumulation and a further ROS induced
by medium-dose UV irradiation, while the same level of SOD induced
by medium-dose UV irradiation did not suppress ROS accumulation
(Figs. 1B and 3B). This was because SOD activity was
increased after low-dose UV irradiation but damaged after
medium-dose UV irradiation (Fig.
3C). The protective role of low-dose UV irradiation contributed
to the increased antioxidant activity more than simple antioxidant
expression.
As inflammation and ROS induce skin epithelial
hyperplasia (27,36), we also demonstrated that low-dose UV
irradiation suppressed medium-dose UV irradiation induced skin
epithelial hyperplasia (Fig. 2Ab and
e), which was paralleled with its inhibition of inflammation
and ROS accumulation. Furthermore, low-dose UV irradiation did not
inhibit high-dose UV irradiation-induced inflammation and ROS, yet
low-dose UV irradiation also did not inhibit high-dose UV
irradiation-induced skin epithelial hyperplasia (Fig. 2Ac and f).
Skin epithelial hyperplasia, resulting from
degradation of the basement membrane (BM) and extracellular matrix
(ECM), is caused by epithelial cell proliferation and migration
(37) and migration was reported to
be correlated with the balance of MMP2 and its inhibitor TIMP2/4
(38,39). Although MMP9, MMP7 and MMP13 were
also reported to be correlated with keratoacanthomas and skin SCC
(40,41), we found that MMP2 was more strongly
correlated with UV-induced skin epithelial hyperplasia (Fig. 4A and B). TIMP2 and TIMP4, well known
as inhibitors of MMP2, presented different reactions to the
increased MMP2 activity (Fig. 4C).
Accompanied by decreased MMP2, TIMP2 was increased but TIMP4 was
also decreased. This indicated that the TIMP2-MMP2 system more
directly influenced UV irradiation-induced skin epithelial
hyperplasia. It may also be because TIMP2-MMP2 and TIMP4-MMP2 are
different balance systems as TIMP2 does not act synergistically
with TIMP4 (42). However,
according to the inhibitory efficiency to aggrecanase-1, it is
possible that TIMP2 has a 5-fold stronger inhibitory activity than
TIMP4 (43), since acidic residues
of Glu and Asp in the TIMP2 C-terminal tail are important for
binding to pro-MMP2, but these acidic residues are lacking in
TIMP4, in which the stability of complex formation with the MMP2
hemopexin C domain is reduced (44). Thus, although TIMP2 was increased
following medium-dose UV irradiation with pre low-dose UV
irradiation, at this time TIMP4 presented a weak inhibitory
activity due to its low expression.
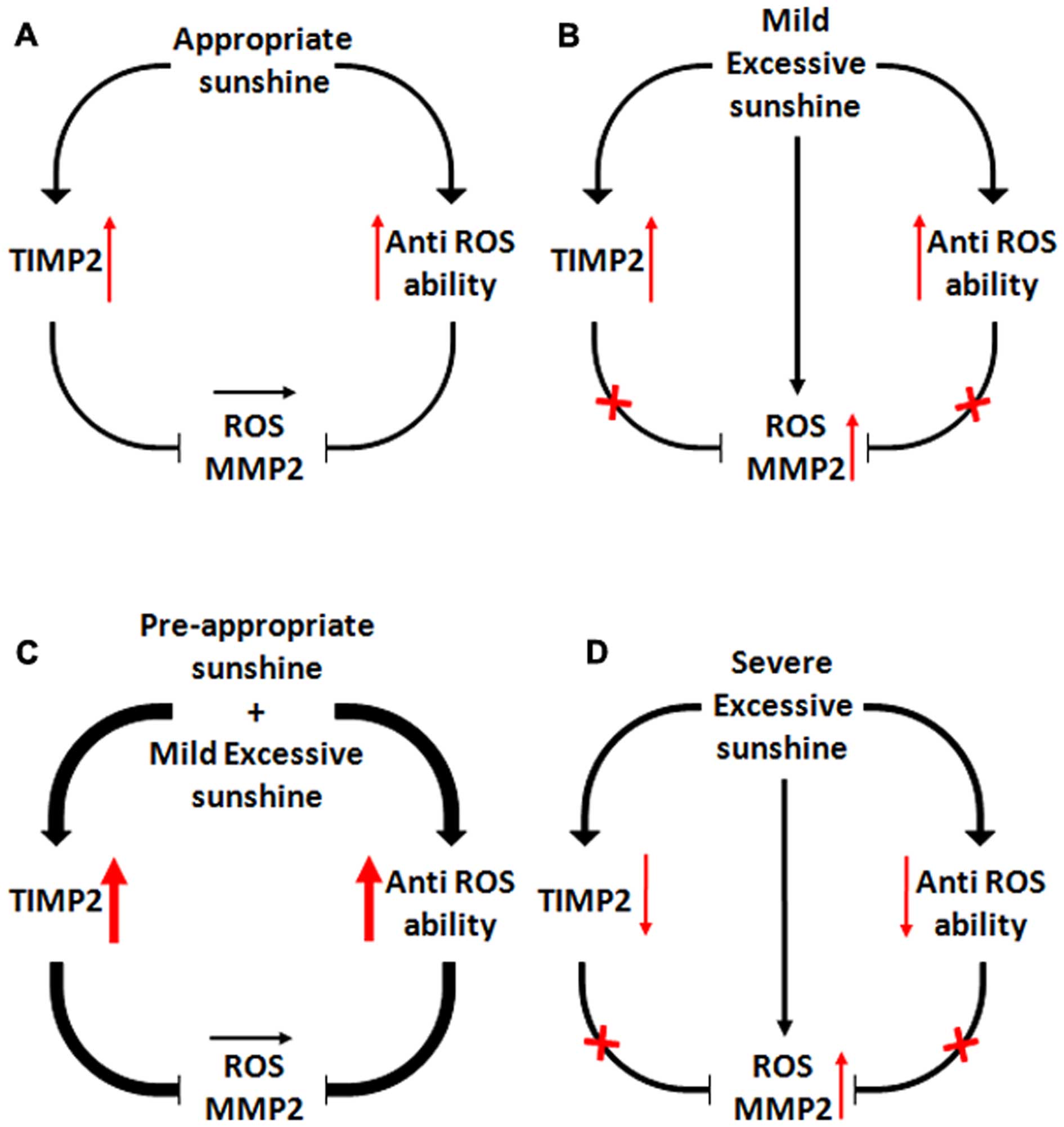
In conclusion as shown in Fig. 5, we demonstrated that low-dose UV
irradiation, which does not induce any inflammation, protected skin
against medium-dose UV irradiation-induced inflammation and ROS and
further inhibited skin epithelial hyperplasia by regulating the
balance of the TIMP2/MMP2 pathway. The present study indicates that
not all UV exposure is bad or carcinogenic and moderate UV
irradiation also has a beneficial role for increasing resistance to
prevent cancer.
References
|
1
|
Jerant AF, Johnson JT, Sheridan CD and
Caffrey TJ: Early detection and treatment of skin cancer. Am Fam
Physician. 62:357–368. 375–356. 381–352. 2000.PubMed/NCBI
|
|
2
|
Marks R: An overview of skin cancers:
Incidence and causation. Cancer. 75:607–612. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
de Gruijl FD: Skin cancer and solar UV
radiation. Eur J Cancer. 35:2003–2009. 1999. View Article : Google Scholar
|
|
4
|
Yoshikawa T, Rae V, Bruins-Slot W, Van den
Berg JW, Taylor JR and Streilein JW: Susceptibility to effects of
UVB radiation on induction of contact hypersensitivity as a risk
factor for skin cancer in humans. J Invest Dermatol. 95:530–536.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
de Gruijl FR, van Kranen HJ and Mullenders
LH: UV-induced DNA damage, repair, mutations and oncogenic pathways
in skin cancer. J Photochem Photobiol B. 63:19–27. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Melnikova VO and Ananthaswamy HN: Cellular
and molecular events leading to the development of skin cancer.
Mutat Res. 571:91–106. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Clydesdale GJ, Dandie GW and Muller HK:
Ultraviolet light induced injury: Immunological and inflammatory
effects. Immunol Cell Biol. 79:547–568. 2001. View Article : Google Scholar
|
|
8
|
Terui T, Okuyama R and Tagami H: Molecular
events occurring behind ultraviolet-induced skin inflammation. Curr
Opin Allergy Clin Immunol. 1:461–467. 2001. View Article : Google Scholar
|
|
9
|
Mantovani A, Alleven P, Sica A and
Balkwill F: Cancer-related inflammation. Nature. 454:436–444. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Diakos CI, Charles KA, McMillan DC and
Clarke SJ: Cancer-related inflammation and treatment effectiveness.
Lancet Oncol. 15:e493–e503. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bickers DR and Athar M: Oxidative stress
in the pathogenesis of skin disease. J Invest Dermatol.
126:2565–2575. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Caricchio R, McPhie L and Cohen PL:
Ultraviolet B radiation-induced cell death: Critical role of
ultraviolet dose in inflammation and lupus autoantigen
redistribution. J Immunol. 171:5778–5786. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lawrence T, Willoughby DA and Gilroy DW:
Anti-inflammatory lipid mediators and insights into the resolution
of inflammation. Nat Rev Immunol. 2:787–795. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Holick MF: Vitamin D: importance in the
prevention of cancers, type 1 diabetes, heart disease, and
osteoporosis. Am J Clin Nutr. 79:362–371. 2004.PubMed/NCBI
|
|
15
|
Holick MF: The vitamin D epidemic and its
health consequences. J Nutr. 135:2739S–2748S. 2005.PubMed/NCBI
|
|
16
|
Bikle DD: The vitamin D receptor: A tumor
suppressor in skin. Adv Exp Med Biol. 810:282–302. 2014.PubMed/NCBI
|
|
17
|
Gloster HM J and Neal K: Skin cancer in
skin of color. J Am Acad Dermatol. 55:741–760. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Halder RM and Bang KM: Skin cancer in
Blacks in the United States. Dermatol Clin. 6:397–405.
1988.PubMed/NCBI
|
|
19
|
Koh D, Wang H, Lee J, Chia KS, Lee HP and
Goh CL: Basal cell carcinoma, squamous cell carcinoma and melanoma
of the skin: Analysis of the Singapore Cancer Registry Data
1968–1997. Br J Dermatol. 148:1161–1166. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Vermeer M, Schmieder GJ, Yoshikawa T, van
den Berg JW, Metzman MS, Taylor JR and Streilein JW: Effects of
ultraviolet B light on cutaneous immune responses of humans with
deeply pigmented skin. J Invest Dermatol. 97:729–734. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gallagher RP, Hill GB, Bajdik CD, Fincham
S, Coldman AJ, McLean DI and Threlfall WJ: Sunlight exposure,
pigmentary factors, and risk of nonmelanocytic skin cancer. I Basal
cell carcinoma Arch Dermatol. 131:157–163. 1995.
|
|
22
|
Rees JL: Genetics of hair and skin color.
Annu Rev Genet. 37:67–90. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jablonski NG: The evolution of human skin
and skin color. Annu Rev Anthropol. 33:585–623. 2004. View Article : Google Scholar
|
|
24
|
Rastogi RP, Singh SP, Häder DP and Sinha
RP: Detection of reactive oxygen species (ROS) by the
oxidant-sensing probe 2′,7′-dichlorodihydrofluorescein diacetate in
the cyanobacterium Anabaena variabilis PCC 7937. Biochem Biophys
Res Commun. 397:603–607. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Luo JD, Wang YY, Fu WL, Wu J and Chen AF:
Gene therapy of endothelial nitric oxide synthase and manganese
superoxide dismutase restores delayed wound healing in type 1
diabetic mice. Circulation. 110:2484–2493. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Braga PC, Marabini L, Wang YY, Lattuada N,
Calò R, Bertelli A, Falchi M, Dal Sasso M and Bianchi T:
Characterisation of the antioxidant effects of Aesculus
hippocastanum L. bark extract on the basis of radical scavenging
activity, the chemiluminescence of human neutrophil bursts and
lipoperoxidation assay. Eur Rev Med Pharmacol Sci. 16(Suppl 3):
1–9. 2012.PubMed/NCBI
|
|
27
|
Mueller MM: Inflammation in epithelial
skin tumours: Old stories and new ideas. Eur J Cancer. 42:735–744.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Graesslin O, Cortez A, Fauvet R, Lorenzato
M, Birembaut P and Daraï E: Metalloproteinase-2, -7 and -9 and
tissue inhibitor of metalloproteinase-1 and -2 expression in
normal, hyperplastic and neoplastic endometrium: A
clinical-pathological correlation study. Ann Oncol. 17:637–645.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Acker T, Fandrey J and Acker H: The good,
the bad and the ugly in oxygen-sensing: ROS, cytochromes and
prolyl-hydroxylases. Cardiovasc Res. 71:195–207. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kawagishi H and Finkel T: Unraveling the
truth about antioxidants: ROS and disease: finding the right
balance. Nat Med. 20:711–713. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
31
|
Perera RM and Bardeesy N: Cancer: When
antioxidants are bad. Nature. 475:43–44. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ristow M and Zarse K: How increased
oxidative stress promotes longevity and metabolic health: The
concept of mitochondrial hormesis (mitohormesis). Exp Gerontol.
45:410–418. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ristow M: Unraveling the truth about
antioxidants: Mitohormesis explains ROS-induced health benefits.
Nat Med. 20:709–711. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tapia PC: Sublethal mitochondrial stress
with an attendant stoichiometric augmentation of reactive oxygen
species may precipitate many of the beneficial alterations in
cellular physiology produced by caloric restriction, intermittent
fasting, exercise and dietary phytonutrients: 'Mitohormesis' for
health and vitality. Med Hypotheses. 66:832–843. 2006. View Article : Google Scholar
|
|
35
|
Klabunde RE and Anderson DE: Role of
nitric oxide and reactive oxygen species in platelet-activating
factor-induced microvas-cular leakage. J Vasc Res. 39:238–245.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wullaert A, Bonnet MC and Pasparakis M:
NF-κB in the regulation of epithelial homeostasis and inflammation.
Cell Res. 21:146–158. 2011. View Article : Google Scholar
|
|
37
|
Thiery JP: Epithelial-mesenchymal
transitions in tumour progression. Nat Rev Cancer. 2:442–454. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Benson JM, Seagrave J, Weber WM,
Santistevan CD, Grotendorst GR, Schultz GS and March TH: Time
course of lesion development in the hairless guinea-pig model of
sulfur mustard-induced dermal injury. Wound Repair Regen.
19:348–357. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Teulière J, Faraldo MM, Deugnier MA,
Shtutman M, Ben-Ze'ev A, Thiery JP and Glukhova MA: Targeted
activation of beta-catenin signaling in basal mammary epithelial
cells affects mammary development and leads to hyperplasia.
Development. 132:267–277. 2005. View Article : Google Scholar
|
|
40
|
Coussens LM, Tinkle CL, Hanahan D and Werb
Z: MMP-9 supplied by bone marrow-derived cells contributes to skin
carcinogenesis. Cell. 103:481–490. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kuivanen TT, Jeskanen L, Kyllönen L,
Impola U and Saarialho-Kere UK: Transformation-specific matrix
metalloproteinases, MMP-7 and MMP-13, are present in epithelial
cells of keratoacanthomas. Mod Pathol. 19:1203–1212. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Toth M, Bernardo MM, Gervasi DC, Soloway
PD, Wang Z, Bigg HF, Overall CM, DeClerck YA, Tschesche H and Cher
ML: Tissue inhibitor of metalloproteinase (TIMP)-2 acts
synergistically with synthetic matrix metalloproteinase (MMP)
inhibitors but not with TIMP-4 to enhance the (Membrane type
1)-MMP-dependent activation of pro-MMP-2. J Biol Chem.
275:41415–41423. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Hashimoto G, Aoki T, Nakamura H, Tanzawa K
and Okada Y: Inhibition of ADAMTS4 (aggrecanase-1) by tissue
inhibitors of metalloproteinases (TIMP-1, 2, 3 and 4). FEBS Lett.
494:192–195. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kai HS, Butler GS, Morrison CJ, King AE,
Pelman GR and Overall CM: Utilization of a novel recombinant
myoglobin fusion protein expression system to characterize the
tissue inhibitor of metalloproteinase (TIMP)-4 and TIMP-2
C-terminal domain and tails by mutagenesis. The importance of
acidic residues in binding the MMP-2 hemopexin C-domain. J Biol
Chem. 277:48696–48707. 2002. View Article : Google Scholar : PubMed/NCBI
|