Introduction
According to the International Agency for Research
on Cancer, the number of estimated new cases of gastric cancer in
worldwide was approximately 989,600, and there were approximately
748,000 estimated deaths due to gastric cancer in the worldwide in
2008 (1). Tanshinone IIA (Tan-IIA;
C19H18O3), is one of the diterpine
quinones extracted from Danshen (Salviae miltiorrhizae
radix) (2,3), with anti-inflammatory activities
(4,5) and antioxidant properties (6,7). It is
well documented that Tanshinone IIA can inhibit many human cancer
cell lines through different molecular mechanisms (8–15).
PI3K can prevent apoptosis and promote cellular survival and
proliferation in a wide variety of cells. Akt is a
serine/threonine-specific protein kinase, which plays a key role in
apoptosis and cell proliferation. Raha et al showed that
Naringin induces autophagy-mediated growth inhibition by
downregulating the PI3K/Akt/mTOR cascade via activation of MAPK
pathways in AGS cancer cells (16).
Lee et al also showed that flavonoids isolated from
Citrus platymamma induce mitochondrial-dependent apoptosis
in AGS cells by modulation of the PI3K/AKT and MAPK pathways
(17). Zheng et al
documented that Paeoniflorin inhibits human gastric carcinoma cell
proliferation through suppression of PI3K/Akt and STAT3 signaling
(18). These documents showed that
agents can inhibit AGS cells through downregulating the
PI3K/Akt/mTOR cascade. Tan-IIA could inhibit human gastric cancer
SGC7901 cells and MKN-45 cells time- and dose-dependently through
inducing apoptosis and cell cycle phase arrest (19–21).
Tan-IIA also inhibited human gastric cancer AGS cells; one of the
molecular mechanisms may be through MAPK pathways to induce G2/M
phase arrest. The other may be through extrinsic apoptotic
signaling pathway to induce apoptosis in vitro (22,23).
Tan-IIA shows potential as an alternative therapeutic agent for
human gastric carcinoma. However, the underlying anticancer
mechanism still needs to be explored further. In the present study,
we investigated the protein expression levels of EGFR, IGFR, PI3K,
AKT, mTOR and PTEN in human gastric cancer AGS cells treated with
Tan-IIA in vitro and in vivo.
Materials and methods
The EGFR (#2239, MW 175 kDa), IGFR (#3018, MW 95
kDa), PI3K (#4292, MW 85 kDa), AKT (#3063, MW 60 kDa), mTOR (#2983,
MW 289 kDa) and PTEN (#9559, MW 54 kDa) antibodies were all
obtained from Cell Signaling Technology Inc. (Beverly, MA, USA).
Tan-IIA was obtained from Sigma-Aldrich (St. Louis, MO, USA; CAS-No
568-72-9). Fetal bovine serum (FBS), F-12K medium, glutamine and
penicillin-streptomycin were obtained from Gibco-BRL (Grand Island,
NY, USA). Triton X-100, Tris-HCl, ribonuclease-A, sodium
deoxycholate, leupeptin, sodium pyruvate, HEPES, dimethyl sulfoxide
(DMSO), 3-(4,5-dimethylthiazol-2-y1)-2,5-diphenyltetrazolium
bromide (MTT) and Tween-20, mouse anti-β-actin were obtained from
Sigma-Aldrich. Potassium phosphate and 0.2 mm PVFD membranes were
purchased from Merck Co. (Darmstadt, Germany); the AGS human
gastric adenocarcinoma cell line (BCRC number: 60102) was obtained
from the Food Industry Research and Development Institute (Hsinchu,
Taiwan). BioMax film was obtained from Kodak.
Male SCID mice
Sixty male SCID mice, weighing 10–15 g (3–4 weeks
old), were obtained from the Laboratory Animal Center, Tzu Chi
University (Hualien, Taiwan).
Cell culture
The human gastric adenocarcinoma AGS cells were
obtained from the Food Industry Research and Development Institute
(Hsinchu, Taiwan). The cell culture procedure was as described
(22,23). Briefly, the AGS cells were placed
into 75-cm2 tissue culture flasks and maintained in
F-12K contained with 10% heat-inactivated FBS (Gibco-BRL), 100 U/ml
penicillin and 100 µg/ml streptomycin. Cells were grown at
37°C in a humidified atmosphere of 95% air and 5%
CO2.
The protein expression of EGFR, IGFR,
PI3K, AKT, mTOR, PTEN and β-actin in AGS cells treated with various
concentrations of Tan-IIA
The AGS cells were treated with various
concentrations of Tan-IIA (0, 2.0, 3.7 and 5.5 µg/ml) for 24
or 48 h and then the protein expression levels of EGFR, IGFR, PI3K,
AKT, mTOR, PTEN and β-actin were evaluated by western blot
analysis.
The protein expressions of EGFR, IGFR,
PI3K, AKT, mTOR, PTEN and β-actin in AGS cells treated with Tan-IIA
for different durations
The AGS cells were treated with Tan-IIA (3.7
µg/ml) for different durations (0, 24 and 48 h) and then the
protein expression levels of VEGFR, HER2, Ras, Raf, MEK, ERK, PARP,
caspase-3 and β-actin were evaluated by western blot analysis.
Effects of Tan-IIA on the protein
expression of EGFR, IGFR, PI3K, AKT, mTOR, PTEN and β-actin in AGS
cell xenograft tumors
Three-week-old male nude SCID mice (number =60) were
xenograft with AGS cells (2x106/0.2 ml) and maintained
in a pathogen-free environment (CCH-AE-102-007; Laboratory Animal
Center of Tzu Chi University, Hualien, Taiwan). When the xenograft
tumors reached more than 0.5 cm in diameter, the mice were divided
randomly into four groups. Tan-IIA was dissolved in corn oil and
then administered to the mice at concentrations of 30, 60 and 90
mg/kg, QW1, 3, 5 by intraperitoneum injection for 8 weeks. The
control group was treated with an equal volume of corn oil. SCID
mice were scarified by CO2 inhalation and then the
xenograft tumors were dissected. All animal experiments were
approved by the Institutional Animal Care and Use Committee
(IACUC), approval no. CCH-AE-102-007). Subsequently, the protein
expressions of EGFR, IGFR, PI3K, AKT, mTOR and PTEN in the tumors
were measured by western blotting.
Protein preparation
Proteins were extracted from xenograft tumors. The
xenograft tumors were dissected and ground, then the thick liquid
(0.06 gm) was lysed in the ice-cold whole cell extract buffer
containing the protease inhibitors. The lysate was vibrated for 30
min at 4°C and centrifuged at 12,281 × g for 10 min. Protein
concentration was measured by BCA protein assay kit (Pierce,
Rockford, IL, USA).
Western blot analysis
The western blot procedures were as described
(22,23). Briefly, AGS cells were treated with
various concentrations of Tan-IIA for different durations, and then
the cells were lysed in the ice-cold whole cell extract buffer
containing the protease inhibitors. The lysate was vibrated for 30
min at 4°C and centrifuged at 12,281 x g for 10 min. Protein
concentration was measured by BCA protein assay kit (Pierce). Equal
amounts of proteins were subjected to electrophoresis using 12%
sodium dodecyl sulfate-polyacrylamide gels. To verify equal protein
loading and transfer, proteins were then transferred to
polyvinylidene difluoride membranes and the membranes were blocked
for 1 h at 4°C using blocking buffer [5% non-fat dried milk in
solution containing 50 mM Tris/HCl (pH 8.0), 2 mM CaCl2,
80 mM sodium chloride, 0.05% Tween-20 and 0.02% sodium azide]. The
membranes were then incubated for 2 h at room temperature with
specific primary antibody followed by anti-rabbit or anti-mouse
immunoglobulin G-horseradish peroxidase conjugated secondary
antibodies. The membranes were washed three times for 10 min with
washing solution. Finally, the protein bands were visualized on the
X-ray film using the enhanced chemiluminescence detection system
(PerkinElmer Life and Analytical Sciences, Boston, MA, USA).
Statistical analysis
Values are presented as the means ± SD. The
Student's t-test was used to analyze statistical significance. A
p-value of <0.05, was considered to indicate a statistically
significant difference for all the tests. P<0.05, P<0.01,
P<0.001.
Results
Effects of Tan-IIA on the viability of
AGS cells
The results revealed that Tan-IIA can inhibit AGS
cells in a time- and dose-dependent manner. The half-maximal
inhibitory concentration (IC50) was 5.5, 3.7 and 3.5
µg/ml at 24, 48 and 72 h, respectively (data not show). This
is agreement with our previous studies (22,23).
Effects of Tan-IIA on the protein
expression of EGFR, IGFR, PI3K, AKT, mTOR, PTEN and β-actin in AGS
cells
The AGS cells were treated with various
concentrations of Tan-IIA (0, 2.0, 3.7 and 5.5 µg/ml) for 24
or 48 h and then the protein expression levels of EGFR, IGFR, PI3K,
AKT, mTOR, p-TEN and β-actin were evaluated by western blot
analysis. The results showed that Tan-IIA can decrease the protein
expression levels of EGFR (Fig.
1A), IGFR (Fig. 1B), PI3K
(Fig. 1C), AKT (Fig. 1D), mTOR (Fig. 1E) and PTEN (Fig. 1F) significantly.
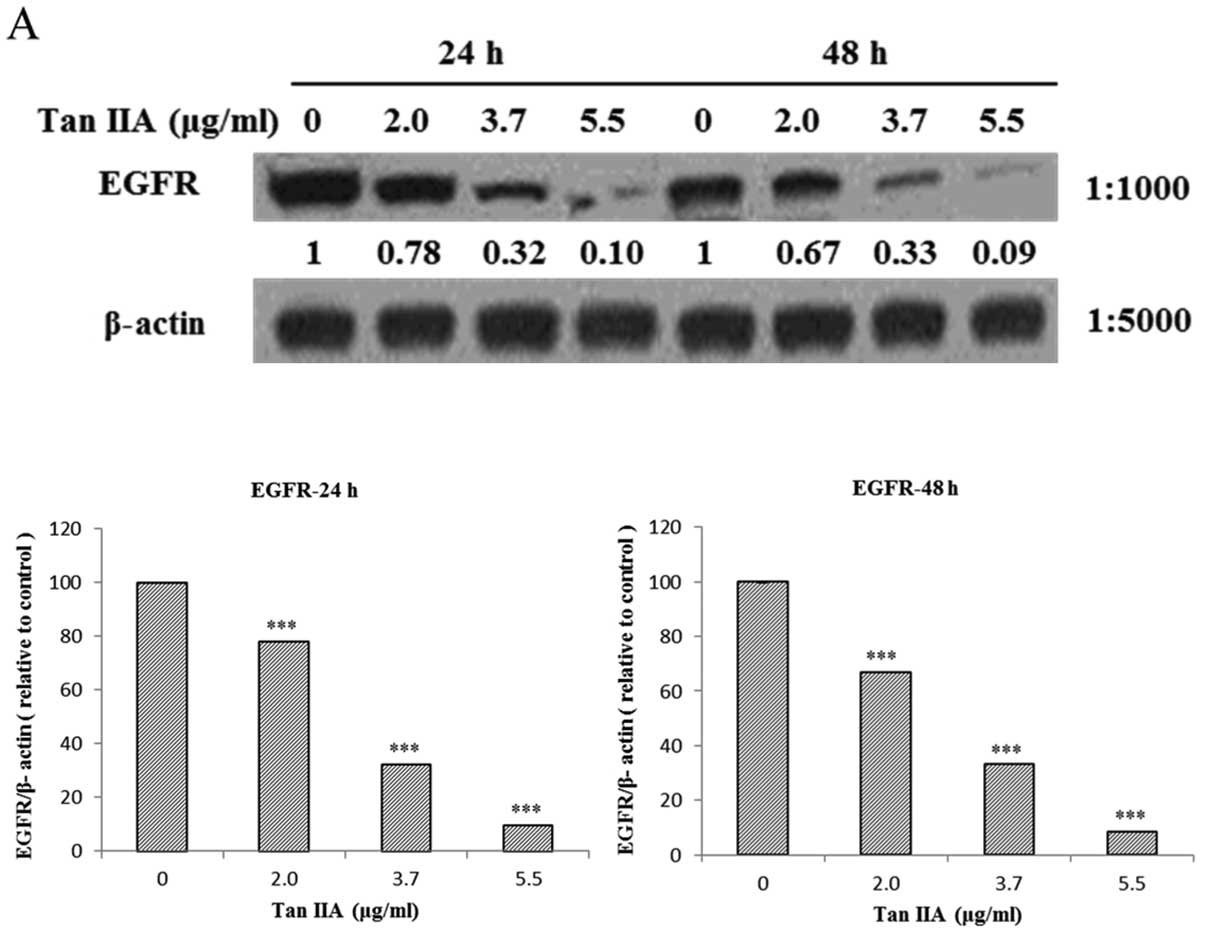 | Figure 1Effects of Tan-IIA on the protein
expression of EGFR, IGFR, PI3K, AKT, mTOR, p-TEN and β-actin in AGS
cells. The AGS cells were treated with various concentrations of
Tan-IIA (0, 2.0, 3.7 and 5.5 µg/ml) for 24 or 48 h and then
the protein expression levels were evaluated by western blot
analysis as described in Materials and methods. The results showed
that Tan-IIA can decrease the protein expression levels of EGFR
(A), IGFR (B), PI3K (C), AKT (D), mTOR (E) and p-TEN (F)
significantly and dose-dependently. *P<0.05, was
considered to indicate a statistically significant difference for
all the tests. *P<0.05, **P<0.01,
***P<0.001. |
Effects of Tan-IIA on the protein
expression of EGFR, IGFR, PI3K, AKT, mTOR, PTEN and β-actin in AGS
cells
The AGS cells were treated with Tan-IIA (3.7
µg/ml) for different durations (0, 24 and 48 h) and then the
protein expression levels of EGFR, IGFR, PI3K, AKT, mTOR, p-TEN and
β-actin were evaluated by western blot analysis. The results showed
that Tan-IIA can decrease the protein expression levels of EGFR
(Fig. 2A), IGFR (Fig. 2B), PI3K (Fig. 2C), AKT (Fig. 2D), mTOR (Fig. 2E) and PTEN (Fig. 2F) significantly.
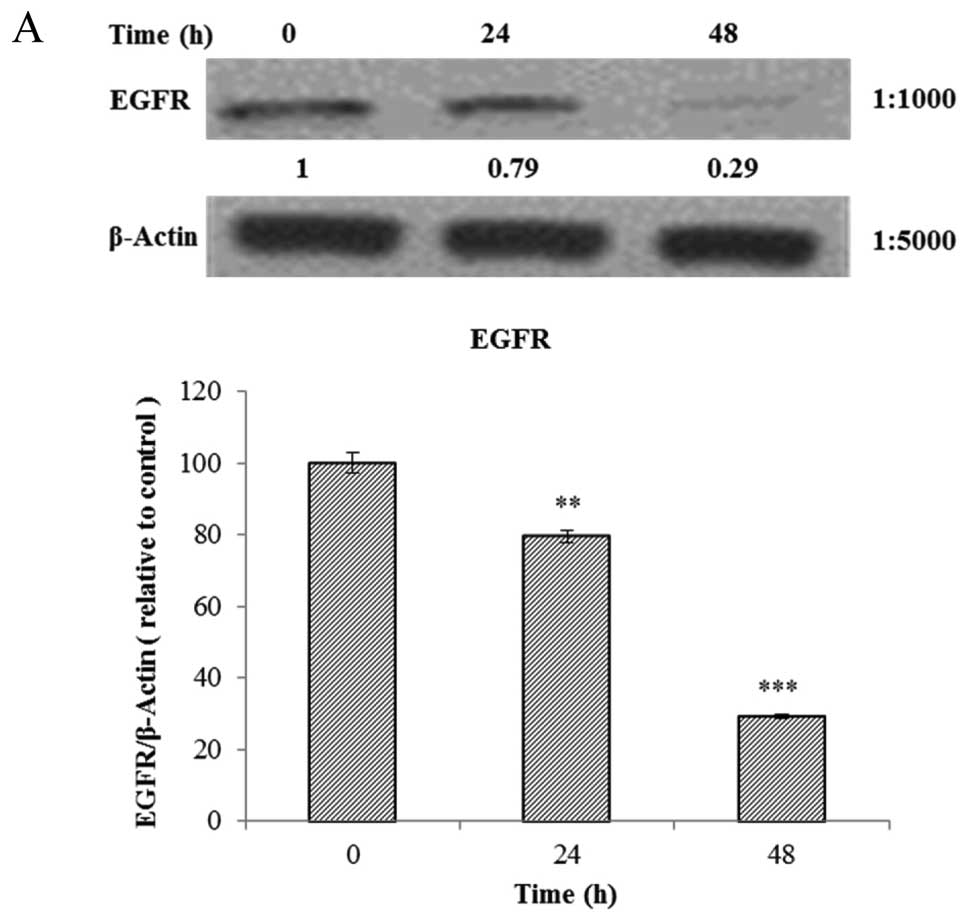 | Figure 2Effects of Tan-IIA on the protein
expression of EGFR, IGFR, PI3K, AKT, mTOR, p-TEN and β-actin in AGS
cells. The AGS cells were treated with Tan-IIA (3.7 µg/ml)
for different durations (0, 24 and 48 h) and then the protein
expression levels were evaluated by western blot analysis as
described in Materials and methods. The results showed that Tan-IIA
can decrease the protein expression levels of EGFR (A), IGFR (B),
PI3K (C), AKT (D), mTOR (E) and p-TEN (F) significantly and
dose-dependently. *P<0.05 was considered to indicate
a statistically significant difference for all the tests.
*P<0.05, **P<0.01,
***P<0.001. |
Effects of Tan-IIA on the protein
expression of EGFR, IGFR, PI3K, AKT, mTOR, PTEN and β-actin in AGS
cell xenograft tumors
The AGS cell xenograft tumor SCID mice were treated
with different doses of Tan-IIA for 8 weeks then sacrificed. The
protein expression of EGFR, IGFR, PI3K, AKT, mTOR and PTEN in
xenograft tumors were measured by western blotting as described in
Materials and methods. The results showed that Tan-IIA can decrease
the protein expression levels of EGFR (Fig. 3A), IGFR (Fig. 3B), PI3K (Fig. 3C), AKT (Fig. 3D), mTOR (Fig. 3E) and PTEN (Fig. 3F) significantly and
dose-dependently.
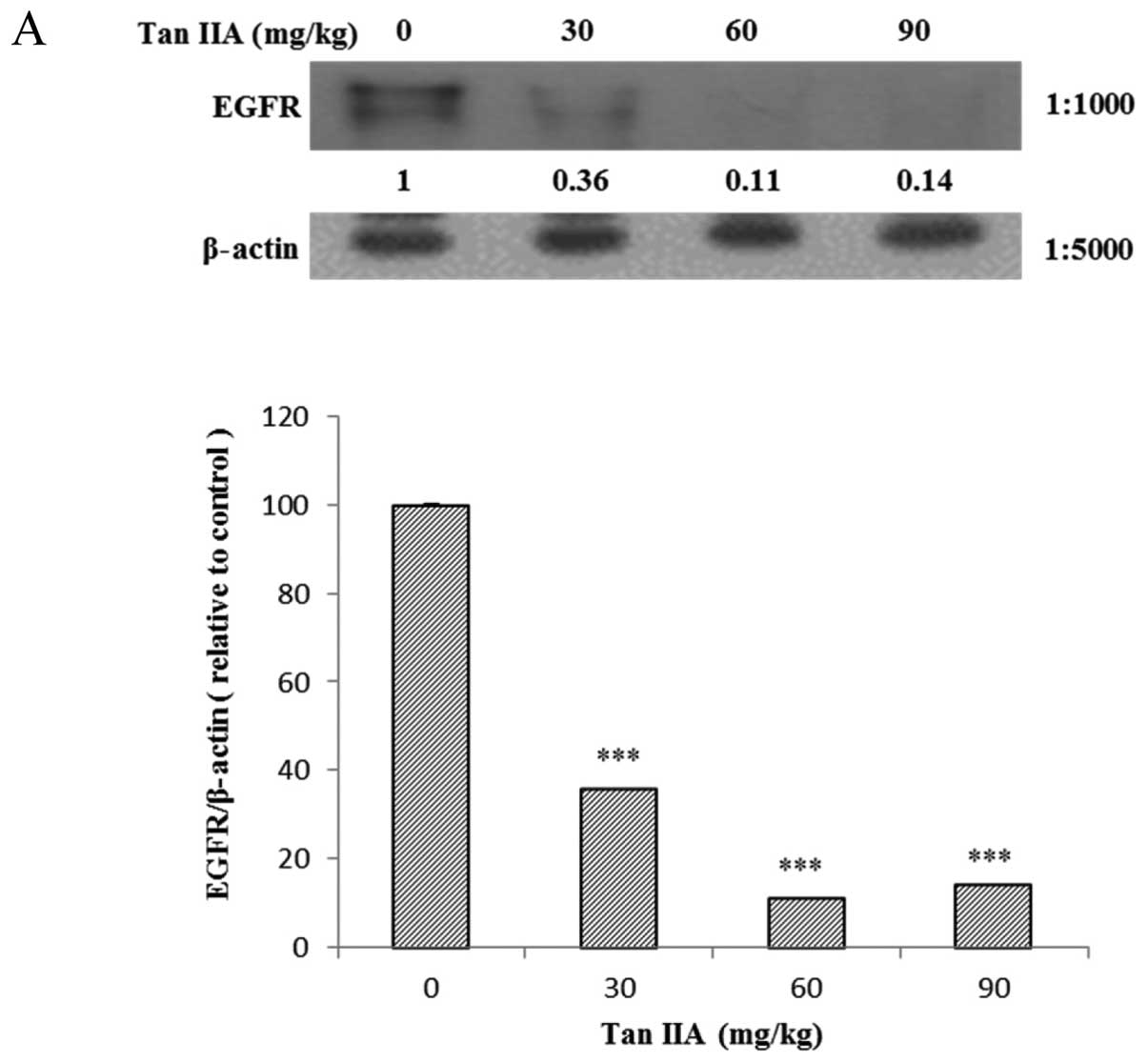 | Figure 3Effects of Tan-IIA on the protein
expressions of EGFR, IGFR, PI3K, AKT, mTOR, p-TEN and β-actin in
AGS cell xenograft tumors. AGS cell xenograft tumors were treated
with different doses of Tan-IIA then the protein expressions were
measured by western blotting as described in Materials and methods.
The results showed that Tan-IIA can decrease the protein expression
levels of EGFR (A), IGFR (B), PI3K (C), AKT (D), mTOR (E) and p-TEN
(F) significantly and dose-dependently. *P<0.05 was
considered to indicate a statistically significant difference for
all the tests. *P<0.05, **P<0.01,
***P<0.001. |
Discussion
It is well documented that PI3K/AKT/mTOR pathway is
one of the most frequently dysregulated kinase cascades in human
cancer (24,25). PI3K/PTEN/Akt/mTOR cascade inhibitors
have been investigated in pre-clinical and clinical investigations
and reported as having potential (26). The transmembrane tyrosine kinases,
such as insulin-like growth factor receptor (IGFR) or epidermal
growth factor receptor (EGFR) have been strongly implicated in the
growth, survival, and metastasis of a wide variety of human tumors
(27,28). PI3K/PTEN/Akt/mTOR pathway represents
important signal transduction mechanisms that facilitate the
proliferation and survival of cancers driven by growth factor
receptors (29).
It has been well documented that Naringin,
flavonoids and Paeoniflorin can inhibit AGS cells through
downregulating the PI3K/Akt/mTOR cascade (16–18).
Our results showed that AGS cells treated with Tan-IIA can
downregulate the protein expressions of PI3K/Akt/mTOR both in
vitro and in vivo.
Protein phosphatase activity is closely associated
with tumors. PI3K is necessary for the activation of AKT. PTEN
regulates the activity of AKT via activated phosphatidylinositol
triphosphate (PIP3) (30). Our
results showed that AGS cells treated with Tan-IIA can downregulate
the protein expression of PTEN both in vitro and in
vivo. These findings indicate Tan-IIA can inhibit the
proliferation of AGS cells, one of the molecular mechanisms may be
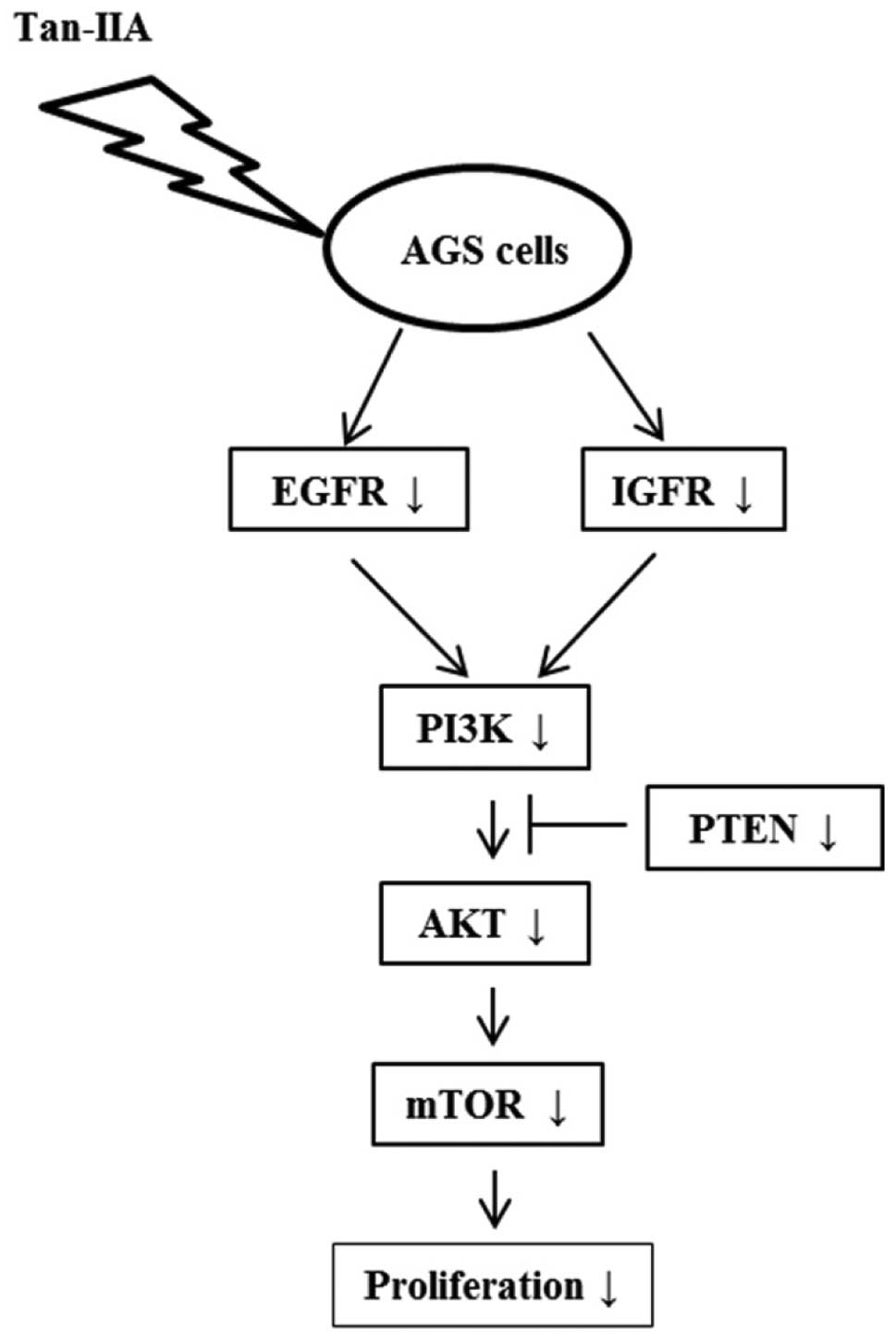
through downregulating the PI3K/Akt/mTOR cascade. The proposed
model is shown in Fig. 4. This is
the first report that Tan-IIA could inhibit gastric carcinoma AGS
cells by decreasing the protein expression of EGFR, IGFR and
blocking the PI3K/Akt/mTOR pathway in a cancer-bearing animal
model.
Acknowledgments
This study was supported by grant 103-CCH-IRP-023
from Changhua Christian Hospital from the Research Section of the
Changhua Christian Hospital (Changhua, Taiwan).
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Che AJ, Zhang JY, Li CH, Chen XF, Hu ZD
and Chen XG: Separation and determination of active components in
Radix Salviae miltiorrhizae and its medicinal preparations by
nonaqueous capillary electrophoresis. J Sep Sci. 27:569–575. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhou L, Zuo Z and Chow MS: Danshen: An
overview of its chemistry, pharmacology, pharmacokinetics, and
clinical use. J Clin Pharmacol. 45:1345–1359. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jang SI, Kim HJ, Kim YJ, Jeong SI and You
YO: Tanshinone IIA inhibits LPS-induced NF-kappaB activation in RAW
264.7 cells: Possible involvement of the NIK-IKK, ERK1/2, p38 and
JNK pathways. Eur J Pharmacol. 542:1–7. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li W, Li J, Ashok M, Wu R, Chen D, Yang L,
Yang H, Tracey KJ, Wang P, Sama AE, et al: A cardiovascular drug
rescues mice from lethal sepsis by selectively attenuating a
late-acting proinflammatory mediator, high mobility group box 1. J
Immunol. 178:3856–3864. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang AM, Sha SH, Lesniak W and Schacht J:
Tanshinone (Salviae miltiorrhizae extract) preparations attenuate
aminoglycoside-induced free radical formation in vitro and
ototoxicity in vivo. Antimicrob Agents Chemother. 47:1836–1841.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lin R, Wang WR, Liu JT, Yang GD and Han
CJ: Protective effect of tanshinone IIA on human umbilical vein
endothelial cell injured by hydrogen peroxide and its mechanism. J
Ethnopharmacol. 108:217–222. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Su CC and Lin YH: Tanshinone IIA
down-regulates the protein expression of ErbB-2 and up-regulates
TNF-α in colon cancer cells in vitro and in vivo. Int J Mol Med.
22:847–851. 2008.PubMed/NCBI
|
|
9
|
Su CC and Lin YH: Tanshinone IIA inhibits
human breast cancer cells through increased Bax to Bcl-xL ratios.
Int J Mol Med. 22:357–361. 2008.PubMed/NCBI
|
|
10
|
Chiu TL and Su CC: Tanshinone IIA induces
apoptosis in human lung cancer A549 cells through the induction of
reactive oxygen species and decreasing the mitochondrial membrane
potential. Int J Mol Med. 25:231–236. 2010.PubMed/NCBI
|
|
11
|
Cheng CY and Su CC: Tanshinone IIA may
inhibit the growth of small cell lung cancer H146 cells by
up-regulating the Bax/Bcl-2 ratio and decreasing mitochondrial
membrane potential. Mol Med Rep. 3:645–650. 2010.
|
|
12
|
Cheng CY and Su CC: Tanshinone IIA
inhibits Hep-J5 cells by increasing calreticulin, caspase 12 and
GADD153 protein expression. Int J Mol Med. 26:379–385.
2010.PubMed/NCBI
|
|
13
|
Yan MY, Chien SY, Kuo SJ, Chen DR and Su
CC: Tanshinone IIA inhibits BT-20 human breast cancer cell
proliferation through increasing caspase 12, GADD153 and
phospho-p38 protein expression. Int J Mol Med. 29:855–863.
2012.PubMed/NCBI
|
|
14
|
Huang CY, Chiu TL, Kuo SJ, Chien SY, Chen
DR and Su CC: Tanshinone IIA inhibits the growth of pancreatic
cancer BxPC 3 cells by decreasing protein expression of TCTP, MCL 1
and Bcl xL. Mol Med Rep. 7:1045–1049. 2013.PubMed/NCBI
|
|
15
|
Su CC: Tanshinone IIA could inhibit
pancreatic cancer BxPC-3 cells through increasing PERK, ATF6,
caspase-12 and CHOP expression to induce apoptosis. J BiSE.
8:149–159. 2015.
|
|
16
|
Raha S, Yumnam S, Hong GE, Lee HJ,
Saralamma VV, Park HS, Heo JD, Lee SJ, Kim EH, Kim JA, et al:
Naringin induces autophagy-mediated growth inhibition by
downregulating the PI3K/Akt/mTOR cascade via activation of MAPK
pathways in AGS cancer cells. Int J Oncol. 47:1061–1069.
2015.PubMed/NCBI
|
|
17
|
Lee HJ, Nagappan A, Park HS, Hong GE,
Yumnam S, Raha S, Saralamma VV, Lee WS, Kim EH and Kim GS:
Flavonoids isolated from Citrus platymamma induce
mitochondrial-dependent apoptosis in AGS cells by modulation of the
PI3K/AKT and MAPK pathways. Oncol Rep. 34:1517–1525.
2015.PubMed/NCBI
|
|
18
|
Zheng YB, Xiao GC, Tong SL, Ding Y, Wang
QS, Li SB and Hao ZN: Paeoniflorin inhibits human gastric carcinoma
cell proliferation through up-regulation of microRNA-124 and
suppression of PI3K/Akt and STAT3 signaling. World J Gastroenterol.
21:7197–7207. 2015.PubMed/NCBI
|
|
19
|
Hou J, He J, Jin X, Hu T and Zhang Y:
Study on optimisation of extraction process of tanshinone IIA and
its mechanism of induction of gastric cancer SGC7901 cell
apoptosis. Afr J Tradit Complement Altern Med. 10:456–458. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xu M, Cao FL, Li NY, Liu YQ, Li YP and Lv
CL: Tanshinone IIA reverses the malignant phenotype of SGC7901
gastric cancer cells. Asian Pac J Cancer Prev. 14:173–177. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Dong X, Dong J, Peng G, Hou X and Wu G:
Growth-inhibiting and apoptosis-inducing effects of Tanshinone II A
on human gastric carcinoma cells. J Huazhong Univ Sci Technolog Med
Sci. 27:706–709. 2007. View Article : Google Scholar
|
|
22
|
Su CC: Tanshinone IIA inhibits human
gastric carcinoma AGS cell growth by decreasing BiP, TCTP, Mcl 1
and Bcl xL and increasing Bax and CHOP protein expression. Int J
Mol Med. 34:1661–1668. 2014.PubMed/NCBI
|
|
23
|
Su CC: Tanshinone IIA inhibits gastric
carcinoma AGS cells through increasing p-p38, p-JNK and p53 but
reducing p-ERK, CDC2 and cyclin B1 expression. Anticancer Res.
34:7097–7110. 2014.PubMed/NCBI
|
|
24
|
Santarpia L, Lippman SM and El-Naggar AK:
Targeting the MAPK-RAS-RAF signaling pathway in cancer therapy.
Expert Opin Ther Targets. 16:103–119. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu P, Cheng H, Roberts TM and Zhao JJ:
Targeting the phosphoinositide 3-kinase pathway in cancer. Nat Rev
Drug Discov. 8:627–644. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
McCubrey JA, Steelman LS, Chappell WH,
Abrams SL, Franklin RA, Montalto G, Cervello M, Libra M, Candido S,
Malaponte G, et al: Ras/Raf/MEK/ERK and PI3K/PTEN/Akt/mTOR cascade
inhibitors: How mutations can result in therapy resistance and how
to overcome resistance. Oncotarget. 3:1068–1111. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yuen JS and Macaulay VM: Targeting the
type 1 insulin-like growth factor receptor as a treatment for
cancer. Expert Opin Ther Targets. 12:589–603. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chitnis MM, Yuen JS, Protheroe AS, Pollak
M and Macaulay VM: The type 1 insulin-like growth factor receptor
pathway. Clin Cancer Res. 14:6364–6370. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
McCubrey JA, Steelman LS, Kempf CR,
Chappell WH, Abrams SL, Stivala F, Malaponte G, Nicoletti F, Libra
M, Bäsecke J, et al: Therapeutic resistance resulting from
mutations in Raf/MEK/ERK and PI3K/PTEN/Akt/mTOR signaling pathways.
J Cell Physiol. 226:2762–2781. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Munroe ME, Businga TR, Kline JN and Bishop
GA: Anti-inflammatory effects of the neurotransmitter agonist
Honokiol in a mouse model of allergic asthma. J Immunol.
185:5586–5597. 2010. View Article : Google Scholar : PubMed/NCBI
|