Introduction
With the advancements in research on mutations in
the epidermal growth factor receptor (EGFR) kinase, as well as
fusions involving anaplastic lymphoma kinase (ALK), the treatment
of patients with lung adenocarcinoma has experienced marked
improvement (1,2). Unfortunately, the therapeutic effect
of these personalized treatment programs is less obvious in the
second most common type of lung cancer, squamous cell lung
carcinoma (SQCLC). Hence, it is necessary to identify other
significant markers as new targets for SQCLC therapy.
Angiogenin (ANG), a 14,400 Da angiogenic
ribonuclease of the RNase A superfamily, is upregulated in a
variety of human cancers and has a 33% amino acid identity and an
overall homology of 56% to that of RNase-A (3). Angiogenin was once thought to promote
cancer progression through its angiogenic activity. However, recent
research has demonstrated that ANG can directly stimulate tumor
cell proliferation and its expression and activity were found to be
upregulated significantly in a variety of human tumors including
breast, colorectal, kidney and pancreatic cancers, and this
upregulation was found to be correlated with tumor progression and
poor prognosis in patients (4). As
a transcription factor, ANG cannot be detected in normal,
non-endothelial cells (5), however,
its expression in human cancer cells and human tumors has not been
widely investigated.
More importantly, ANG is implicated in a wide range
of biological functions including cell survival, growth,
proliferation, migration, tube formation and tumor angiogenesis
(6–8).
Epithelial-mesenchymal transition (EMT), a process
in which epithelial cells lose or modify their apical-basal
polarity and are converted to a mesenchymal phenotype, is a
critical process during embryonic development and tumor metastasis
(9,10). The process of EMT is typically
characterized by losing the expression of the epithelial markers,
E-cadherin and β-catenin, which are transmembrane proteins
essential for stable adherens junctions, and acquiring the
expression of mesenchymal markers, such as N-cadherin, vimentin and
TGF-β (11,12). We, therefore, hypothesized that ANG
is an inducer of EMT in human squamous cell lung carcinoma and
enhances the metastatic potential.
Materials and methods
Tissue collection
The cancerous and corresponding non-tumor normal
specimens of SQCLC, freshly frozen for western blot analysis and
qRT-PCR, were obtained from 50 patients who underwent lung cancer
resection procedures at Changhai Hospital from October, 2012 to
October, 2015. All cancerous and matching non-cancerous samples
used for this study were provided by the Clinical and Experimental
Pathology of the Research Centre of Changhai Hospital, Shanghai,
China where initial H&E staining and histologic diagnoses were
performed after standard surgical primary tumor resection. This
study was conducted in accordance with the Declaration of Helsinki
and was approved by the Medical Ethics Committee of Changhai
Hospital, with all patients providing written informed consent.
Cell culture
SK-MES-1 cells were obtained from the Cancer Cell
Repository (Shanghai Cell Bank, Shanghai, China), and were
maintained in Dulbecco's modified Eagle's medium (DMEM)
supplemented with 10% heat-inactivated fetal bovine serum
(Gibco-BRL) and antibiotics (100 U/ml penicillin and 100 U/ml
streptomycin; Hyclone Laboratories, Inc., Logan, UT, USA) at 37°C
in a humidified atmosphere of 5% CO2.
RNA interference and ANG
overexpression
The ANG-specific short hairpin RNA adenovirus vector
(shANG) and the negative control short hairpin RNA adenovirus
vector (shScramble) were purchased from Genechem Co., Ltd.
(Shanghai, China). The sequences of shANG were: sense strand 1:
5′-CCGCGGGATGACAGATACTGTGAAGAGATTCACAGTATCTGTCATCCCG-3′ and
antisense strand 1:
5′-AACGGGATGACAGATACTGTGAATCTCTTCACAGTATCTGTCATCCCGC-3′; sense
strand 2: 5′-CCGGATCCCAGGCTCGTTCTTTGGAGACAAAGAACGAGCCTGGGATCC-3′
and antisense strand 2:
5′-AAGGATCCCAGGCTCGTTCTTTGTCTCCAAAGAACGAGCCTGGGATCC-3′. An
adenovirus vector co-expressing ANG (Ad-ANG) and a control empty
adenovirus vector (Ad-Null) were constructed using AdEasy system
(Genechem Co., Ltd., Shanghai, China).
Quantitative real-time
reverse-transcription polymerase chain reaction (qRT-PCR)
Total RNA was extracted and cDNA was synthesized
using the PrimeScript RT Reagent kit (Takara Bio). SYBR Premix EX
Taq (Takara Bio) was used for quantitative real-time polymerase
chain reaction performed using a LightCycler® 480
Real-Time PCR system (Roche Diagnostics). The primers for each gene
were as follows: 18S rRNA forward: 5′-CGGACACGGACAGGATTGAC-3′; 18S
rRNA reverse: 5′-GCATGCCAGAGTCTCGTTCG-3′; ANG forward:
5′-GGACTTGTTCTGAGGCCGAG-3′; ANG reverse:
5′-CCAGCACGAAGACCAACAAC-3′. The amplification conditions were as
follows: 1 cycle of 95°C for 30 sec and 40 cycles of 95°C for 5 sec
followed by 60°C for 30 sec. The expression levels of the target
genes relative to the control were determined using the
2−ΔΔCT method. 18S rRNA served as the internal
control.
Western blot analysis
Western blot analyses were performed as previously
described (13). The primary
antibodies included: angiogenin (Sc-74528, Santa Cruz
Biotechnology, Inc., 1:1,000), vimentin (ab92547, Abcam, 1:2,000),
TGF-β1 (ab92486, Abcam, 1:1,000), E-cadherin (ab40772, Abcam,
1:10,000) and β-catenin (60008-1-Ig, Proteintech, 1:2,000).
Immunohistochemistry
Immunohistochemical assay was performed as
previously described (14). The
primary antibodies used were: angiogenin (Sc-74528, Santa Cruz
Biotechnology, Inc., 1:100), vimentin (ab92547, Abcam, 1:200),
TGF-β1 (ab92486, Abcam, 1:100), E-cadherin (ab40772, Abcam, 1:500),
N-cadherin (ab12221, Abcam, 1:500), and β-catenin (ab32572, Abcam,
1:500).
Immunofluorescence
Paraformaldehyde-fixed SK-MES-1 cells were first
incubated with primary antibodies at 4°C overnight. After being
washed three times with PBS, the cells were incubated with
secondary antibodies for 1 h at 37°C. Nuclei were counterstained
with 4′,6-diamidino-2-phenylindole (DAPI) for 2 min. The primary
antibodies used were angiogenin (Sc-74528, Santa Cruz
Biotechnology, Inc., 1:1,000), E-cadherin (ab40772, Abcam, 1:500)
and N-cadherin (ab12221, Abcam, 1:500). The secondary antibodies
used were Alexa Fluor 498-conjugated anti-rabbit IgG (1:400,
Jackson ImmunoResearch).
Scratch assay
Cells were placed in a 24-well plate at an initial
density of 1×105 cells/well. A uniform monolayer formed
in 2–3 days. All wound-healing assays were performed in a
serum-free medium. Thirty-six hours after infection, a
micro-pipette tip was used to create a wound in the monolayer by
scraping. Images were captured at 24 h after the scratch and the
migrated cells were quantitated.
Invasion assay
The invasion assay was performed using Transwell
insert chambers with a pore size of 8 µm (Corning). The
Transwell filter inserts were coated with Matrigel;
0.5×105 cells were seeded in serum-free medium in the
upper chamber. After a 24-h incubation at 37°C, cells in the upper
chamber were carefully removed with a cotton swab and the cells
that had traversed the membrane were fixed, stained in a 0.1%
crystal violet solution and counted.
Cell proliferation
Cell proliferation was analyzed using a Cell
Counting Kit-8 (Beyotime, China). Twelve hours after being plated
into a 96-well plate at a density of 2,000 cells/well the cells
were transfected with an adenovirus. Next, the cells were incubated
for 0, 24, 48 and 72 h; then, 10 µl CCK-8 solution was added
to each well and the cultures were incubated at 37°C for 1 h.
Subsequently, the absorbance at 450 nm was measured.
Statistical analysis
All statistical analyses were performed using SPSS
version 19.0. The quantitative data were first evaluated on whether
they followed the normal distribution by the Shapiro-Wilk test. The
data with a non-normal distribution were analyzed using the
Kruskal-Wallis test. The data with a normal distribution were
assessed using the Student's t-test. A P-value <0.05 was
considered to be a statistically significant difference.
Results
ANG is expressed in SQCLC and positively
correlates with a high grade of malignancy of SQCLC
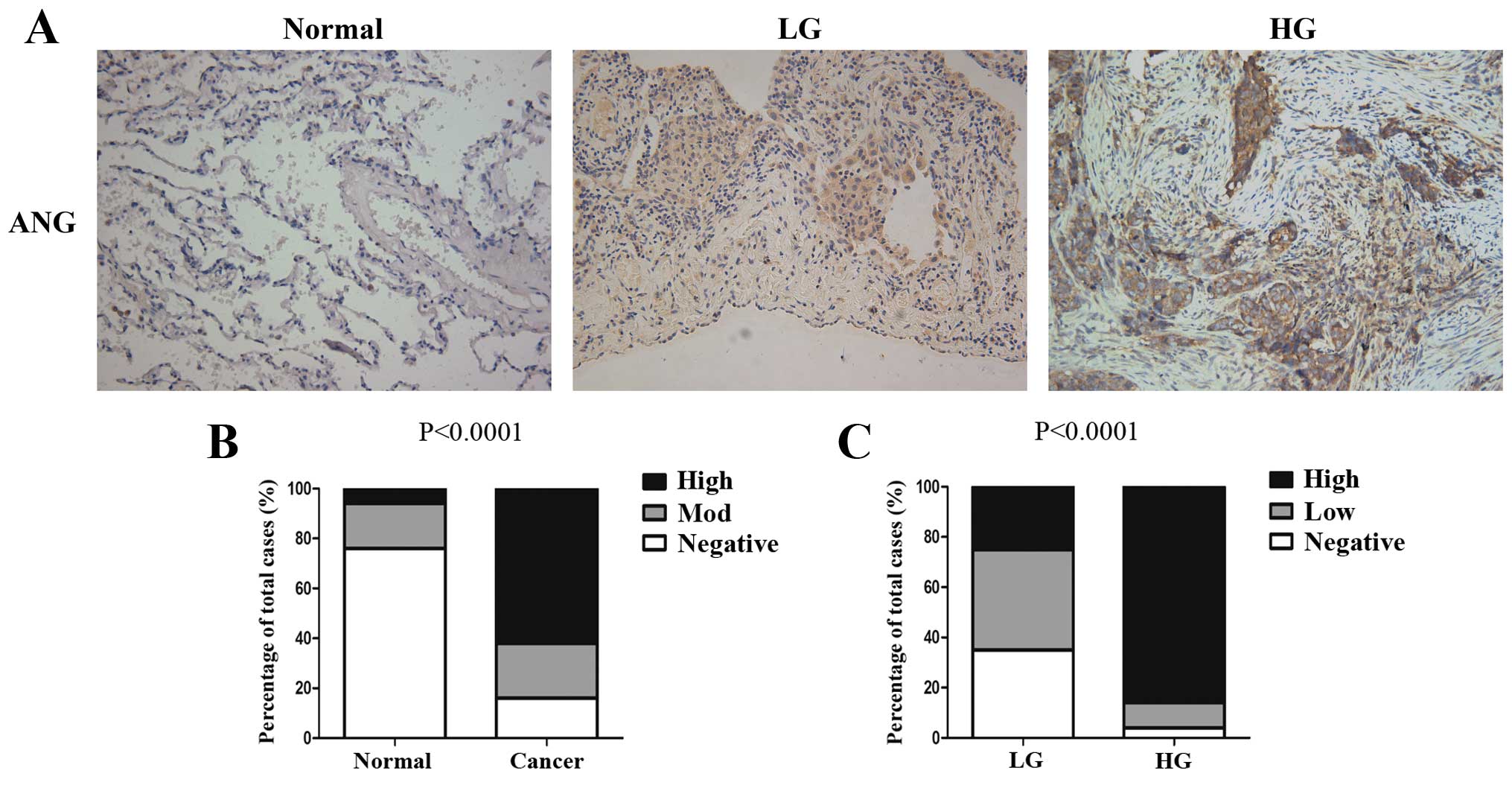
Immunohistochemical assays showed that ANG was
expressed in the SQCLC tissues, while it was not detected in the
normal lung tissues adjacent to tumors (Fig. 1A). As shown in Fig. 1B, ANG was significantly increased in
the SQCLC tissues compared with that noted in the normal lung
tissues (P<0.0001). Notably, high-grade SQCLC which indicates a
higher postoperative surgical-pathologic staging exhibited the
strongest ANG expression (Fig. 1A).
Consistently, the difference in the patterns of ANG expression in
low-grade and high-grade SQCLC was significant (low-grade vs.
high-grade, P<0.0001, Fig. 1C),
which showed that a high percentage of high-grade SQCLC cases had
strong ANG expression when compared with the expression in its
low-grade counterpart. Collectively, our results indicated that ANG
expression increased with the malignant grade of SQCLC.
Upregulation of ANG enhances
proliferation, migration and invasion of SK-MES-1 cells in
vitro
Since the local expression level of ANG was
considerably increased in the SQCLC tissues, especially in the
cancerous tissues from high-grade SQCLC patients where ANG
expression was intensely positive, as compared with the adjacent
normal tissue, we considered that ANG may play an important role in
the development, metastasis and invasiveness of SQCLC. To
investigate the effect of ANG on SQCLC, we used an in vitro
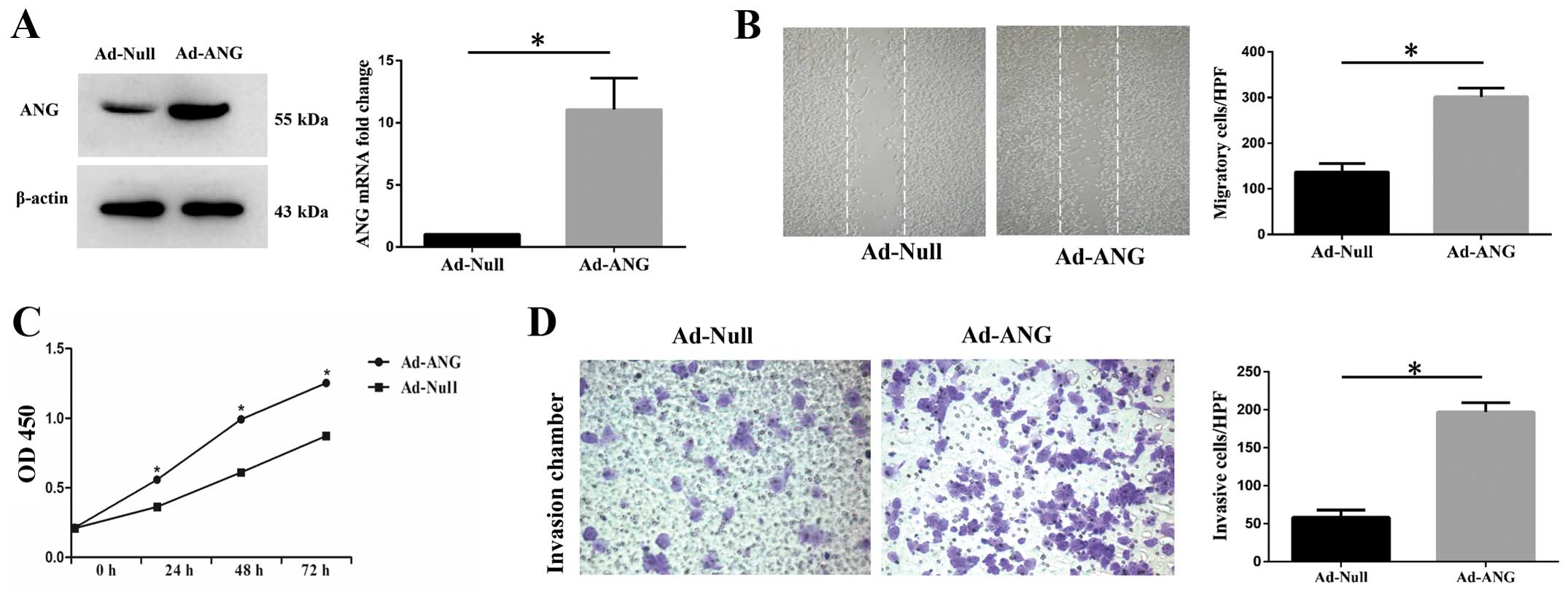
system. Firstly, we effectively infected SK-MES-1 cells with Ad-ANG
(Fig. 2A). Evaluation of cell
proliferation was performed after infection using a CCK-8 assay
kit. It was found that Ad-ANG SK-MES-1 cells manifested increased
proliferation levels at 24, 48 and 72 h after infection (Fig. 2C). With regard to SK-MES-1 cell
migration and invasion capability evaluation, scratch assay and
invasion chamber assays were applied to assess the impact of ANG on
these cell properties. Results showed that Ad-ANG SK-MES-1 cells
presented not only significantly increased migration but also
invasion capability (Fig. 2B and
D), as compared with the Ad-Null cells.
These results definitely indicated that ANG is an
important regulator of proliferation, migration and invasion in
SK-MES-1 cells.
Knockdown of ANG reduces the ability of
proliferation, migration and invasion of SK-MES-1 cells in
vitro
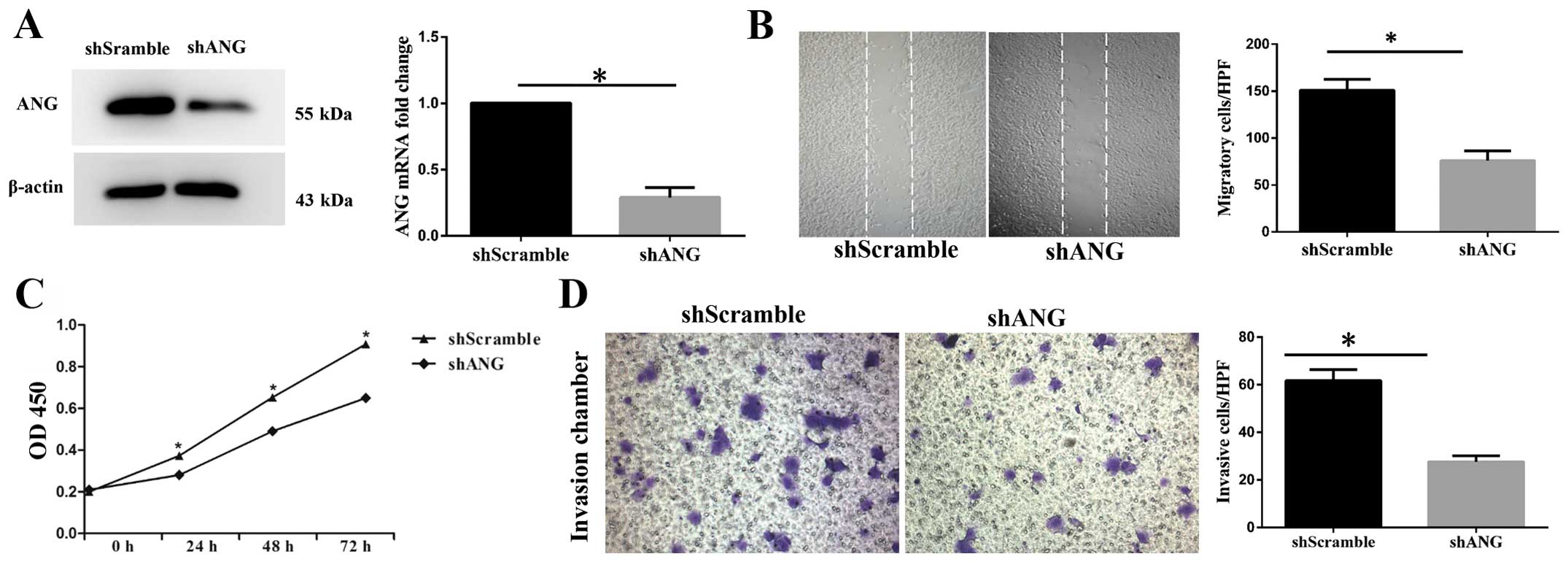
We further used shANG to knock down ANG in SK-MES-1
cells. The cells were infected with shANG and cultured for 48 h.
SK-MES-1 cells infected with shScramble were used as a negative
control. Compared with the shScramble SK-MES-1 cells, the
expression level of ANG mRNA in the shANG cells was decreased by
70% (Fig. 3A). Western blot
analysis results also confirmed lower expression level of ANG in
the shANG SK-MES-1 cells (Fig. 3A).
Furthermore, it was revealed that the shANG SK-MES-1 cells showed
decreased proliferation capability (Fig. 3C). Meanwhile, decreased migration
and invasion capacities were also observed in the shANG SK-MES-1
cells, as compared with the shScramble SK-MES-1 cells (Fig. 3B and D).
EMT is associated with the grade of
malignancy of SQCLC
Given the fact that EMT contributes to metastasis
and invasiveness of various cancers, we examined the expression of
several EMT markers in both low-grade and high-grade SQCLC tissues.
As shown in Fig. 4, compared with
low-grade SQCLC, epithelial markers including E-cadherin and
β-catenin were significantly reduced, while mesenchymal markers
including N-cadherin, vimentin and TGF-β1 were markedly enhanced in
the high-grade SQCLC, indicating that EMT may contribute to a
metastatic and aggressive phenotype of SQCLC.
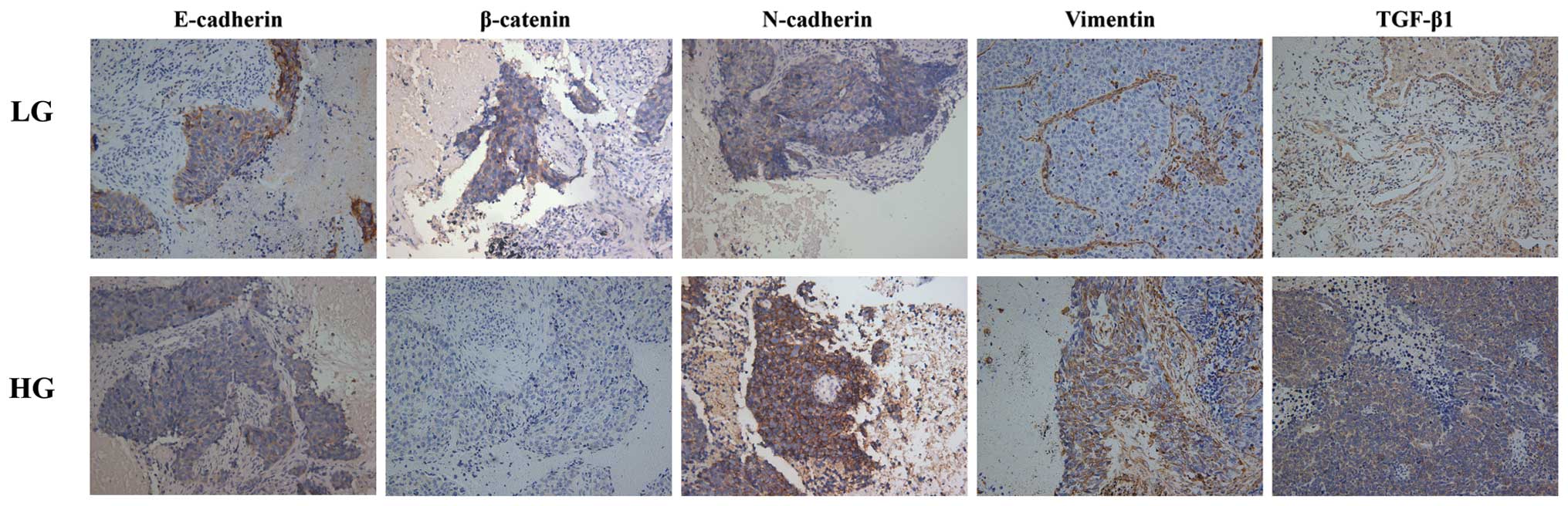 | Figure 4Immunohistochemical staining of
mesenchymal and epithelial markers, including TGF-β1, vimentin,
N-cadherin, E-cadherin and β-catenin in SQCLC. Compared with
low-grade SQCLC, epithelial markers including E-cadherin and
β-catenin were significantly reduced, while mesenchymal markers
including N-cadherin, vimentin and TGF-β1 were markedly enhanced in
the high-grade SQCLC. SQCLC, squamous cell lung carcinoma; LG,
low-grade; HG, high-grade. |
Overexpression of ANG upregulates the
expression of mesenchymal markers but downregulates epithelial
markers in the SK-MES-1 cell line
After showing that both ANG and EMT were positively
correlated with a high grade of malignancy of SQCLC, we further
tested whether ANG induces the metastatic and invasive phenotype of
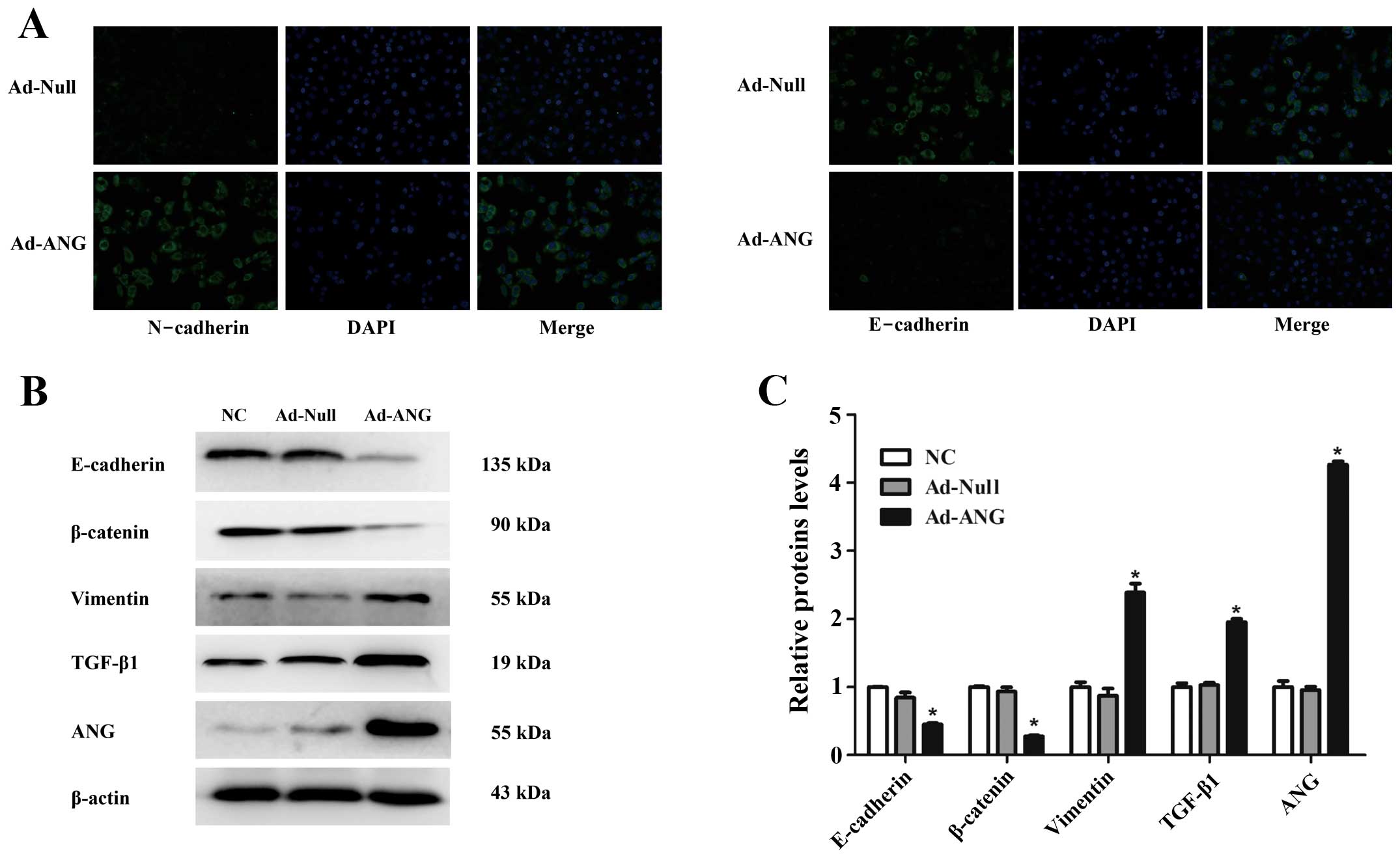
SQCLC by augmenting EMT. As shown in Fig. 5A, we found that the E-cadherin
staining was decreased while N-cadherin staining was increased in
the Ad-ANG SK-MES-1 cells as compared with the Ad-Null cells.
Furthermore, western blot analysis confirmed that the expression of
vimentin and TGF-β1 was significantly enhanced in the Ad-ANG cells.
In contrast, Ad-ANG SK-MES-1 cells exhibited a lower level of
protein expression of epithelial markers including E-cadherin and
β-catenin, as compared with the Ad-Null and negative control groups
(Fig. 5B and C, respectively;
P<0.01). These data indicated that ANG triggers the process of
EMT in SK-MES-1 cells.
Discussion
ANG, a 14.4 kDa polypeptide, was first isolated from
serum-free supernatants of a human adenocarcinoma cell line, HT-29
(15). In addition to angiogenic
activity, ANG is also implicated in a variety of biological
functions including cell growth, proliferation, migration and tube
formation (6,8,16). Of
note, some studies have reported that ANG can undergo nuclear
translocation to stimulate rRNA transcription in both cancer cells
and cancer-associated endothelial cells (17). Recent research has shown that the
expression and activity of ANG are upregulated in many human tumors
including breast, kidney, colorectal and lung cancers (18). In the present study,
immunohistochemical assays showed that ANG was expressed in SQCLC
tissues, while it was not expressed in normal lung tissues adjacent
to tumors, indicating that ANG is involved in the development of
SQCLC, which is consistant with the finding that increased ANG is
positively associated with the incidence of human tumors.
ANG has been reported as a member of the urokinase
plasminogen activator receptor interactome that participates in the
formation of plasmin and the migration of breast cancer cells,
which are necessary for tumor metastasis and invasion (19). In a clinical study, researchers
found that, among the markers representing different aspects of
cancer vascular biology and exhibiting abnormal expression in
colorectal cancer, ANG was the only index which was independently
associated with the increasing stage of colorectal cancer according
to the Dukes' and American Joint Committee on Cancer systems
(20). Similarly, in patients with
adenocarcinoma, our previous study showed that increased expression
of ANG was correlated with vascular and pleural invasion as well as
positive lymph node metastasis (6).
Consistently, we found that increased expression of ANG was
positively correlated with the high grade of SQCLC while knockdown
of ANG abrogated proliferation, migration and invasion of SK-MES-1
cells. These results from clinical specimens and cultured squamous
cell lung carcinoma cell lines supported the notion that ANG
contributes to the invasive phenotype of squamous cell lung
carcinoma cells.
Numerous studies have demonstrated that cell
invasion during tumor progression may be dependent on the
acquisition of EMT features (21).
Evidence from clinical studies also suggests that poor survival of
cancer patients and drug-resistance are associated with EMT
phenotypes in malignant cancer cells (22). EMT is a complex process during which
epithelial cells lose their polarity and cell-cell adhesion,
exhibit enhanced cell-extracellular matrix adhesion, gain invasive
properties and become mesenchymal-like cells. In both squamous cell
lung carcinoma tissues and SK-MES-1 cells, we showed that high
expression of ANG was positively correlated with the expression of
mesenchymal markers but negatively associated with epithelial
markers. These data suggest that ANG induces EMT in squamous cell
lung carcinoma. The intermediate filament protein vimentin is an
important marker of EMT and a critical regulator of mesenchymal
cell migration. Induction of vimentin is associated with increased
expression of Axl which enhanced the migratory activity of
pre-malignant breast epithelial cells (23). Cell adhesion molecules are important
to cancer invasion and metastasis. E-cadherin to N-cadherin switch
was found to promote cancer progression via TGF-β-induced EMT in
extrahepatic cholangiocarcinoma (24) and is of strong importance in the
progression of prostate cancer (25). In lung cancer cell line, H1650ER,
N-cadherin expression was also reported to be upregulated and
paralleled by the reduced expression of E-cadherin. Proliferation
and invasion of H1650ER cells were inhibited by knockdown of
N-cadherin (26). In accordance
with these previous studies, our results indicated that
overexpression of ANG led to the upregulation of vimentin,
fibronectin and N-cadherin and reduced expression of E-cadherin,
suggesting that ANG contributes to invasion and metastasis of lung
carcinoma by inducing EMT.
Many molecular and cell signaling pathways have been
reported to participate in the EMT process. For instance,
accumulating evidence indicates that the phosphatidylinositol
3-kinase (PI3K)/Akt/mTOR pathway plays an important role in EMT
(27,28). The MAPK/ERK pathway has also been
demonstrated to be critical for many features of EMT including
acquisition of invasive properties, enhanced MMP activity, and
attenuation of adherens junctions. EMT mediated by TGF-β also
involves the activation of the MAPK/ERK pathway (29). In addition, research has shown that
receptor activator of NF-κB (RANK) can induce EMT in human mammary
epithelial cells and promotes oncogenesis and metastasis (30). Notably, correlation of ANG and these
cell signaling pathways has been demonstrated in many studies.
Overexpression of ANG can activate phosphorylation of downstream
molecules of the PI3K/AKT/mTOR signaling pathway in bladder cancer
cells (31). In cultured human
umbilical vein endothelial cells, ANG induced the activation of
both ERK1/2 and SAPK/JNK (32,33).
Moreover, ANG prevented stress-induced death of P19 embryonal
carcinoma cells via upregulation of the Bcl-2 and NF-κB pathways
(34). Given these results from
extensive studies, we speculate that ANG may induce EMT via a
complex regulating network involving various cell signaling
pathways.
In summary, this study suggests that ANG promotes
the invasion and metastasis of SQCLC by enhancing proliferative,
migratory and invasive ability of squamous cell carcinoma cells
through the induction of EMT. Our results highlight the possibility
that ANG may serve as a target for the treatment of squamous cell
carcinoma of the lung.
Abbreviations:
|
ANG
|
angiogenin
|
|
EMT
|
epithelial-mesenchymal transition
|
|
EGFR
|
epidermal growth factor receptor
|
|
SQCLC
|
lung squamous cell carcinoma
|
Acknowledgments
This study was supported by the National Natural
Science Foundation of China (nos. 81272592 and 81301829).
References
|
1
|
Paez JG, Jänne PA, Lee JC, Tracy S,
Greulich H, Gabriel S, Herman P, Kaye FJ, Lindeman N, Boggon TJ, et
al: EGFR mutations in lung cancer: Correlation with clinical
response to gefitinib therapy. Science. 304:1497–1500. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Soda M, Choi YL, Enomoto M, Takada S,
Yamashita Y, Ishikawa S, Fujiwara S, Watanabe H, Kurashina K,
Hatanaka H, et al: Identification of the transforming EML4-ALK
fusion gene in non-small-cell lung cancer. Nature. 448:561–566.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li S and Hu GF: Angiogenin-mediated rRNA
transcription in cancer and neurodegeneration. Int J Biochem Mol
Biol. 1:26–35. 2010.PubMed/NCBI
|
|
4
|
Nilsson UW, Abrahamsson A and Dabrosin C:
Angiogenin regulation by estradiol in breast tissue: tamoxifen
inhibits angiogenin nuclear translocation and antiangiogenin
therapy reduces breast cancer growth in vivo. Clin Cancer Res.
16:3659–3669. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ibaragi S, Yoshioka N, Kishikawa H, Hu JK,
Sadow PM, Li M and Hu GF: Angiogenin-stimulated rRNA transcription
is essential for initiation and survival of AKT-induced prostate
intraepithelial neoplasia. Mol Cancer Res. 7:415–424. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yuan Y, Wang F, Liu XH, Gong DJ, Cheng HZ
and Huang SD: Angiogenin is involved in lung adenocarcinoma cell
proliferation and angiogenesis. Lung Cancer. 66:28–36. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ivanov P, Emara MM, Villen J, Gygi SP and
Anderson P: Angiogenin-induced tRNA fragments inhibit translation
initiation. Mol Cell. 43:613–623. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li S and Hu GF: Emerging role of
angiogenin in stress response and cell survival under adverse
conditions. J Cell Physiol. 227:2822–2826. 2012. View Article : Google Scholar :
|
|
9
|
Polyak K and Weinberg RA: Transitions
between epithelial and mesenchymal states: Acquisition of malignant
and stem cell traits. Nat Rev Cancer. 9:265–273. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mani SA, Guo W, Liao MJ, Eaton EN, Ayyanan
A, Zhou AY, Brooks M, Reinhard F, Zhang CC, Shipitsin M, et al: The
epithelial-mesenchymal transition generates cells with properties
of stem cells. Cell. 133:704–715. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zeisberg M and Neilson EG: Biomarkers for
epithelial-mesenchymal transitions. J Clin Invest. 119:1429–1437.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kokkinos MI, Wafai R, Wong MK, Newgreen
DF, Thompson EW and Waltham M: Vimentin and epithelial-mesenchymal
transition in human breast cancer-observations in vitro and in
vivo. Cells Tissues Organs. 185:191–203. 2007. View Article : Google Scholar
|
|
13
|
Kurozumi A, Kato M, Goto Y, Matsushita R,
Nishikawa R, Okato A, Fukumoto I, Ichikawa T and Seki N: Regulation
of the collagen cross-linking enzymes LOXL2 and PLOD2 by
tumor-suppressive microRNA-26a/b in renal cell carcinoma. Int J
Oncol. 48:1837–1846. 2016.PubMed/NCBI
|
|
14
|
Hu K, Tian Y, Du Y, Huang L, Chen J, Li N,
Liu W, Liang Z and Zhao L: Atrazine promotes RM1 prostate cancer
cell proliferation by activating STAT3 signaling. Int J Oncol.
48:2166–2174. 2016.PubMed/NCBI
|
|
15
|
Fett JW, Strydom DJ, Lobb RR, Alderman EM,
Bethune JL, Riordan JF and Vallee BL: Isolation and
characterization of angiogenin, an angiogenic protein from human
carcinoma cells. Biochemistry. 24:5480–5486. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Miyake M, Goodison S, Lawton A,
Gomes-Giacoia E and Rosser CJ: Angiogenin promotes tumoral growth
and angiogenesis by regulating matrix metallopeptidase-2 expression
via the ERK1/2 pathway. Oncogene. 34:890–901. 2015. View Article : Google Scholar :
|
|
17
|
Li L, Pan XY, Shu J, Jiang R, Zhou YJ and
Chen JX: Ribonuclease inhibitor up-regulation inhibits the growth
and induces apoptosis in murine melanoma cells through repression
of angiogenin and ILK/PI3K/AKT signaling pathway. Biochimie.
103:89–100. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shu J, Huang M, Tian Q, Shui Q, Zhou Y and
Chen J: Down-regulation of angiogenin inhibits the growth and
induces apoptosis in human bladder cancer cells through regulating
AKT/mTOR signaling pathway. J Mol Histol. 46:157–171. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Dutta S, Bandyopadhyay C, Bottero V,
Veettil MV, Wilson L, Pins MR, Johnson KE, Warshall C and Chandran
B: Angiogenin interacts with the plasminogen activation system at
the cell surface of breast cancer cells to regulate plasmin
formation and cell migration. Mol Oncol. 8:483–507. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ramcharan SK, Lip GY, Stonelake PS and
Blann AD: Angiogenin outperforms VEGF, EPCs and CECs in predicting
Dukes' and AJCC stage in colorectal cancer. Eur J Clin Invest.
43:801–808. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Makker A and Goel MM: Tumor progression,
metastasis, and modulators of epithelial-mesenchymal transition in
endometrioid endometrial carcinoma: An update. Endocr Relat Cancer.
23:R85–R111. 2016. View Article : Google Scholar
|
|
22
|
Mitra A, Mishra L and Li S: EMT, CTCs and
CSCs in tumor relapse and drug-resistance. Oncotarget.
6:10697–10711. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Vuoriluoto K, Haugen H, Kiviluoto S,
Mpindi JP, Nevo J, Gjerdrum C, Tiron C, Lorens JB and Ivaska J:
Vimentin regulates EMT induction by Slug and oncogenic H-Ras and
migration by governing Axl expression in breast cancer. Oncogene.
30:1436–1448. 2011. View Article : Google Scholar
|
|
24
|
Araki K, Shimura T, Suzuki H, Tsutsumi S,
Wada W, Yajima T, Kobayahi T, Kubo N and Kuwano H: E/N-cadherin
switch mediates cancer progression via TGF-β-induced
epithelial-to-mesenchymal transition in extrahepatic
cholangiocarcinoma. Br J Cancer. 105:1885–1893. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gravdal K, Halvorsen OJ, Haukaas SA and
Akslen LA: A switch from E-cadherin to N-cadherin expression
indicates epithelial to mesenchymal transition and is of strong and
independent importance for the progress of prostate cancer. Clin
Cancer Res. 13:7003–7011. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang X, Liu G, Kang Y, Dong Z, Qian Q and
Ma X: N-cadherin expression is associated with acquisition of EMT
phenotype and with enhanced invasion in erlotinib-resistant lung
cancer cell lines. PLoS One. 8:e576922013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Berg A, Hoivik EA, Mjøs S, Holst F, Werner
HM, Tangen IL, Taylor-Weiner A, Gibson WJ, Kusonmano K, Wik E, et
al: Molecular profiling of endometrial carcinoma precursor, primary
and metastatic lesions suggests different targets for treatment in
obese compared to non-obese patients. Oncotarget. 6:1327–1339.
2015. View Article : Google Scholar :
|
|
28
|
Hipp S, Walch A, Schuster T, Losko S, Laux
H, Bolton T, Höfler H and Becker KF: Activation of epidermal growth
factor receptor results in snail protein but not mRNA
overexpression in endometrial cancer. J Cell Mol Med. 13:3858–3867.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Edme N, Downward J, Thiery JP and Boyer B:
Ras induces NBT-II epithelial cell scattering through the
coordinate activities of Rac and MAPK pathways. J Cell Sci.
115:2591–2601. 2002.PubMed/NCBI
|
|
30
|
Palafox M, Ferrer I, Pellegrini P, Vila S,
Hernandez-Ortega S, Urruticoechea A, Climent F, Soler MT, Muñoz P,
Viñals F, et al: RANK induces epithelial-mesenchymal transition and
stemness in human mammary epithelial cells and promotes
tumorigenesis and metastasis. Cancer Res. 72:2879–2888. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Peng Y, Li L, Huang M, Duan C, Zhang L and
Chen J: Angiogenin interacts with ribonuclease inhibitor regulating
PI3K/AKT/mTOR signaling pathway in bladder cancer cells. Cell
Signal. 26:2782–2792. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xu Z, Monti DM and Hu G: Angiogenin
activates human umbilical artery smooth muscle cells. Biochem
Biophys Res Commun. 285:909–914. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu S, Yu D, Xu ZP, Riordan JF and Hu GF:
Angiogenin activates Erk1/2 in human umbilical vein endothelial
cells. Biochem Biophys Res Commun. 287:305–310. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li S, Yu W, Kishikawa H and Hu GF:
Angiogenin prevents serum withdrawal-induced apoptosis of P19
embryonal carcinoma cells. FEBS J. 277:3575–3587. 2010. View Article : Google Scholar : PubMed/NCBI
|