Introduction
Tumor marker genes earn their functional importance
from the fact that they play an essential role during disease
development. Thus, their detection is fundamental for outcome
monitoring and targeted treatment. Different types of tumor marker
genes have been proposed including prognostic (1), diagnostic (2), predictive (3), pharmacodynamic (4) and recurrence (5) markers. In different tumor types,
reduced oxygen supply to the tumor microenvironment, namely
hypoxia, plays an important role during disease development to more
severe stages (6).
Hypoxia-inducible factor-1α (HIF-1α) is known to play an essential
role in the regulation of genes induced or upregulated by the
hypoxic tumor microenvironment (7,8). The
cytoplasmic protein N-myc downstream-regulated gene 1 (NDRG1), also
known as CAP43, non-race specific disease specific protein 1
(NDR1), differentiation-related gene 1 protein (Drg-1),
nickel-specific induction protein (Cap43), protein regulated by
OXYgen-1 (PROXY1) and NMSL protein, belong to the α/β-hydrolase
superfamily (9). NDRG1 is involved
in a diversity of cellular characteristics such as specific stress
responses, hormone responses, cell growth and differentiation.
NDRG1 gene mutations cause the neuropathy Charcot-Marie-Tooth
disease type 4D (10). Its
expression may also possess the potential to be used as a
prognostic factor for some types of cancer (11). Cell differentiation signals
upregulate NDRG1 and result in metastasis suppression (9). However, observations regarding the
role of NDRG1 in human cancer development are controversial. In
hepatocellular carcinoma (HCC) upregulation of NDRG1 has been shown
to correlate with tumor aggressiveness and patient survival
(11); in colon cancer cell lines
NDRG1 downregulation was observed and its overexpression led to
induction of cell differentiation (12). Moreover, a low NDRG1 protein
expression level was found to correlate with unfavorable patient
prognosis in glioma (13),
colorectal cancer (14), esophageal
squamous cell carcinoma (15),
pancreatic ductal adenocarcinoma (16), prostate (17,18)
and breast cancer (19).
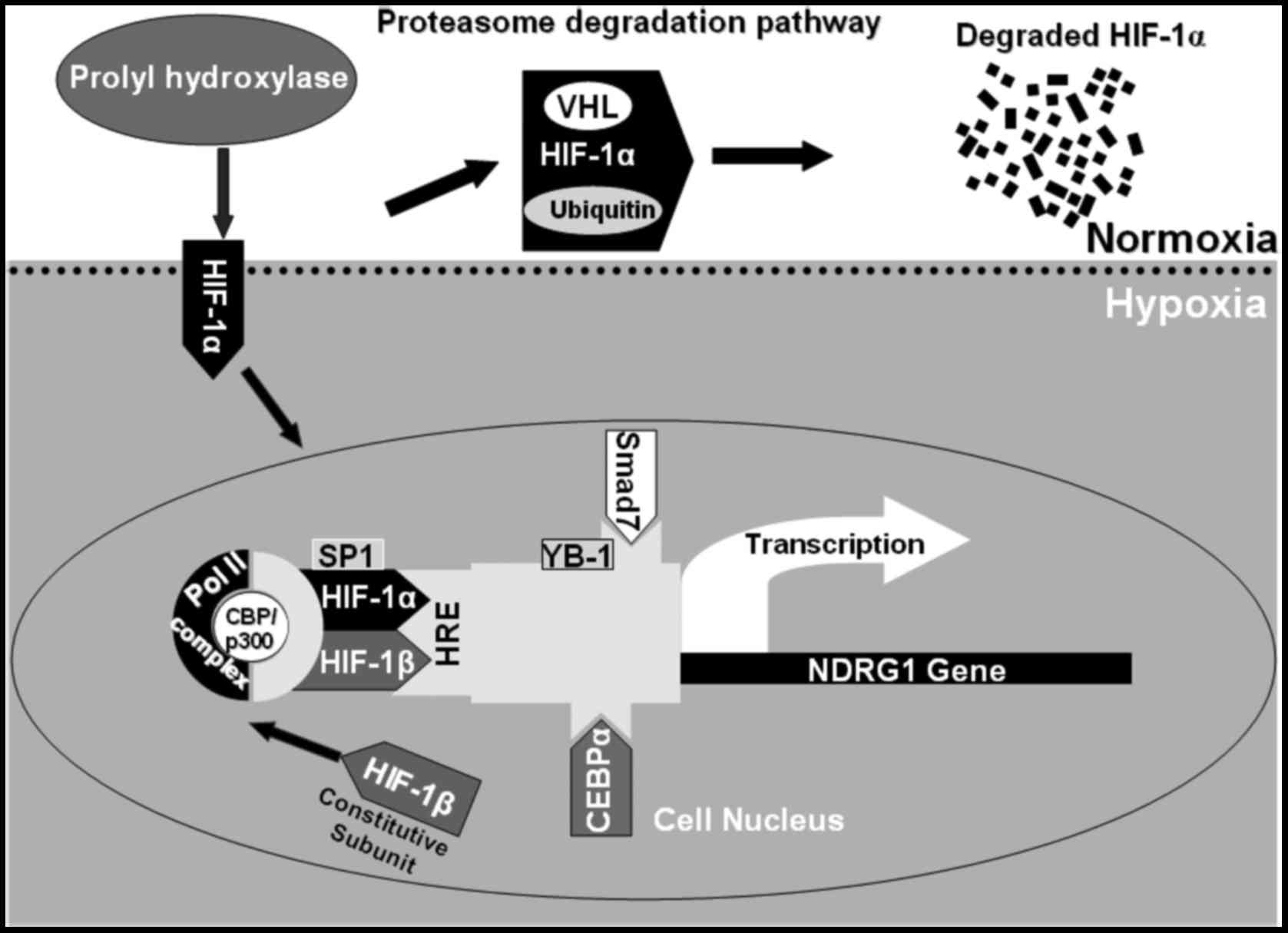
Under hypoxia, accumulation of HIF-1α and its
translocation to the nucleus along with the co-activator CBP/p300
results in its binding to hypoxia-responsive elements (HREs) of
target gene promoters and subsequent regulation of their
transcription (22,23). Studies on the NDRG1 promoter
identified HREs in the upstream promoter region and suggest
regulation of NDRG1 by HIF-1α (24). However, these studies have revealed
controversial results for the manner of NDRG1 regulation by HIF-1α
under hypoxia. Various of these studies reported a marked
upregulation of the NDRG1 expression level under hypoxic
conditions, whereas others have shown downregulated NDRG1 under the
same conditions (8,13–15,18,19,25).
Consequently, the regulation of NDRG1 under hypoxia is highly
complex and its cellular function is still a matter of debate.
However, it is established that besides HIF-1α several other
transcription factors involved in tumorigenesis are implicated in
its regulation (8,20,21).
Thus, it is important to clearly define the role of NDRG1 during
carcinogenesis as well as the regulatory events steering this
process.
It has been shown that different signal transduction
pathways are modulated and controlled by different factors such as
cytokines and growth factors or environmental conditions such as
hypoxia for example (26).
Transforming growth factor-β1 (TGF-β1) is one such signaling
protein. Pathways involving Smad family members are also important
regulators (27). Such proteins are
crucial for the pathophysiological regulation and involved in the
development of various diseases including fibrosis (28) and cirrhosis, leading to HCCs
(29). Particularly, one member of
the Smad family, Smad7, abrogates the TGF-β1 signaling pathway by
initiating a negative feedback loop (30). Such a regulation may be involved in
the process of cancer manifestation as seen for HCC (31). Smad7 is regulated by different
pathways (32). One of these
involves the transcription factor YB-1, a member of the cold shock
family of proteins (30).
Specific transcription factor 1 (SP1) is another
protein involved in the regulation of many cancer-related genes
(33). In addition, it is involved
in the active regulation of vascular endothelial growth factor
(VEGF) which is another HIF-1α regulated hypoxia gene (34). Since the HIF-1α promoter contains
SP1-specific transcription elements (35), SP1 may also be an important factor
in NDRG1 regulation.
The transcription factor CCAAT enhancer binding
protein α (CEBPα), which belongs to the BLZIP family (36), plays an important role in governing
metabolic processes relevant for disease development including
cancer (37). Although HIF-1α is
well recognized as a main regulator during hypoxia (38), the role of other transcription
factors during hypoxic conditions in the tumor microenvironment
have not yet been clearly established. Therefore, we aimed to
examine NDRG1 protein and mRNA expression under different
oxygenation conditions in human glioblastoma multiforme (GBM) cell
lines to gain further insight into its biological role. In
addition, the expression levels of potential NDRG1 gene-regulating
transcription factors HIF-1α, SP1, CEBPα Smad7 and YB-1 were
investigated. At least to a certain extent, we planned to address,
whether these factors may have the potential to serve as tumor
markers.
Materials and methods
Cell lines, cell culture and hypoxia
treatment
The human malignant glioma cell lines U87-MG and
U373 were originally purchased from the American Type Culture
Collection (ATCC; Rockville, MD, USA). GaMG is an established cell
line from a patient with GBM (Gade Institute, University Bergen,
Bergen, Norway) (39). Cell lines
were grown on glass Petri dishes in Dulbecco's modified Eagle's
medium (DMEM), supplemented with 10% fetal bovine serum (FBS),
penicillin (100 IU/ml), streptomycin (100 µg/ml) and 2 mM
L-glutamine in a humidified incubator with 5% CO2 at
37°C. For hypoxia treatment, the cells were exposed to 0.1%
O2 for 1, 6 and 24 h in a Ruskinn hypoxic workstation
(Ruskinn Technology Ltd., Cincinnati, OH, USA) as previously
described (8). Cells incubated
under aerobic conditions served as the negative control; cells
treated with 100 µM of the chelating agent desferrioxamine (DFO)
under aerobic conditions served as the positive control.
Nuclear extract preparation
Nuclear extracts were prepared according to
previously described protocols with minor modifications (8,40). A
total of 5×106 cells were used for each experiment. The
cells were scratched-off the Petri dishes using 10 ml
phosphate-buffered saline (PBS) and a pellet was obtained by
centrifugation. The supernatant was discarded. Subsequently, the
cells were transferred into a pre-chilled micro-centrifuge tube,
re-suspended in 1 ml PBS and centrifuged at 14,000 × g for 45 sec
at 4°C. The cells were then gently re-suspended in 400 µl ice cold
hypotonic buffer [10 µM/l HEPES pH 7.9, 10 µM/l KCl, 0.1 µM/l
ethylenediaminetetraacetic acid (EDTA), 0.1 µM/l ethylene glycol
tetraacetic acid (EGTA), 1 µM/l phenylmethylsulfonyl fluoride
(PMSF), 10 µl complete protease inhibitor cocktail (Roche,
Mannheim, Germany) and 1 µmol/l dithiothreitol (DTT)] by pipetting
up and down several times, and then incubated on ice for 15 min.
The cells were lysed by adding 25 µl of 10% NP40 with successive
vortexing for 10 sec at the highest setting. The homogenate was
centrifuged for 10 min at 3,000 × g at 4°C. The supernatant,
containing the cytoplasmic fraction, was transferred and saved. The
pellet, containing the nuclear extract, was re-suspended in 50 µl
complete cell extraction buffer [20 µM/l HEPES pH 7.9, 0.4 M/l
NaCl, 1 µM/l EDTA, 1 µM/l EGTA, 1 µM/l PMSF and 0.1 µl protease
inhibitor cocktail (Roche)] by shaking it for 20 min at 4°C in a
tube shaker followed by centrifugation at 14,000 × g and 4°C for 5
min. The supernatant was transferred to a clean micro-centrifuge
tube and stored at −80°C. The protein concentration was quantitated
using the Bradford method protein Quantitation Assay kit 5000001
(Bio-Rad, Munich, Germany).
Whole-cell lysate preparation and
western blotting
Whole-cell lysates were prepared with 0.1 ml RIPA
buffer (Amresco, Vienna, Austria), 0.5% sodium deoxycholate, 0.1%
SDS, protease inhibitors pepstatin A 1.4 µM, aprotinin 0.15 µM and
leupeptin 2.3 µM and 100 µM PMSF (all from Sigma-Aldrich, Munich,
Germany). Phosphatase inhibitor mix (Sigma-Aldrich) was added. The
lysates were transferred to micro-centrifuge tubes, followed by 30
min incubation on ice. Subsequently, cell lysates were cleared by
centrifugation at 15,000 × g for 12 min at 4°C.
Western blotting was performed using 20 µg of
protein lysates as previously described (40,41).
The protein lysates were size-fractionated on 8% polyacrylamide
gels (NuPAGE; Life Technologies Carlsbad, CA, USA) by
electrophoresis, transferred onto nitrocellulose membranes (Protran
BA85; Schleicher & Schuell, Dassel, Germany), and incubated
with antibodies directed against HIF-1α (BD Biosciences,
Heidelberg, Germany), CEBPα (Abcam Plc., Cambridge, UK), Egr1, SP1
(both from Santacruz Biotech, Heidelberg, Germany), NDRG1, YB-1,
MADH7 (all from Abcam Plc.), β-actin and β-tubulin antibodies (both
from Sigma-Aldrich). Bound antibodies were detected by developing
the membrane using the ECL Plus western blotting detection system
for 5 min with subsequent development of the Hyperfilm (both from
Amersham Biosciences, Cambridge, UK).
Total RNA isolation and
semi-quantitative reverse transcription-polymerase chain reaction
(RT-PCR) analysis
Total RNA was isolated from GBM cell lines as
previously described (40,41). The RNA (1–5 µg) was reverse
transcribed to cDNA using the RevertAid Reverse Transcription kit
1691 (Thermo Fisher Scientific GmbH, Schwerte, Germany) in a 20 µl
reaction mixture for 1 h at 42°C according to the manufacturer's
instructions. RT-PCR analysis then was performed utilizing PCR
systems and reagents from Promega™ (Mannheim, Germany) according to
the manufacturer's instructions. The primers were designed in
flanking exons with Primer3 software (available online http://frodo.wi.mit.edu/cgibin/primer3/primer3_www.cgi)
(Table I). The PCR products were
separated on 1% agarose gels (Sigma-Aldrich) and visualized by
staining with 0.07 µg/ml ethidium bromide (Bio-Rad).
 | Table I.Examined genes and primers used. |
Table I.
Examined genes and primers used.
|
| Gene | Gene Bank accession
no. | Forward primer
(FP) | Reverse primer
(RP) | PCR product size
(bp) |
|---|
| 1 | SP1 | NM138473.2 |
5′-ATGGCAAGACCTCTCACCTG-3′ |
5′-TCTCTTGGACCCATGCTACC-3′ | 526 |
| 2 | Smad7 | NM_005904.2 |
5′-CTCGGTGCTCAAGAAACTGA-3′ |
5′-AATCCATCGGGTATCTGGAG-3′ | 362 |
| 3 | CEBPα | NM_004364.2 |
5′-AACCTTGTGCCTTGGAAATG-3′ |
5′-CCCTATGTTTCCACCCCTTT-3′ | 246 |
| 4 | YB-1 | NM_004559.3 |
5′-GGAGATGAGACCCAAGGTCA-3′ |
5′-GGTGCTTGCAGTTTGTTGAC-3′ | 551 |
| 5 | Egr-1 | NM_001964.2 |
5′-AGCTGGAGGAGATGATGCTG-3′ |
5′-ACAAGGTGTTGCCACTGTT-3′ | 348 |
| 6 | HIF-1α | NM_001530.2 |
5′-TTACAGCAGCCAGACGATCA-3′ |
5′-CCCTGCAGTAGGTTTCTGCT-3′ | 233 |
| 7 | NDRG1 | NM_006096.3 |
5′-CTCTGTTCACGTCACGCTGT-3′ |
5′-CTCCACCATCTCAGGGTTGT-3′ | 593 |
| 8 | β-actin | NM_001101.3 |
5′-CGTGCGTGACATTAAGGAGA-3 |
5′-CACCTTCACCGTTCCAGTTT'-3 | 668 |
Densitometric evaluation and
statistical analysis
Western blotting band densities and signal strength
of RT-PCR were analyzed with 1D Kodak Image Analysis Software
(8). DNA or protein amounts were
calculated as previously described (21). Statistical data analysis was
performed using SPSS 15.0 (SPSS, Inc., Chicago, IL, USA). All
experiments were conducted as triplets. Differences between groups
were analyzed with either the Student's-test (unpaired, two-tailed)
or the Mann-Whitney U test. Statistical significance was considered
at p<0.05.
Results
Time- and oxygen
concentration-dependent protein expression of NDRG1 and its
potential transcription factors
Long-term but not short-term hypoxia leads to the
induction of NDRG1 in human glioma cell lines. Previous studies
have shown that such a change is associated with tumor progression
and an unfavorable prognosis of glioma patients (8). Since hypoxia plays an important role
in the regulation of NDRG1 expression, we examined the NDRG1 mRNA
and protein expression levels and its regulatory pathways under
extreme hypoxia and re-oxygenation in the human glioma cell lines
U87-MG, U373 and GaMG. We assumed that the hypoxia-induced
transcription factors HIF-1α, CEBPα, Egr-1, SP1, YB-1 and Smad7 may
be involved in NDRG1 regulation. Therefore, we examined the protein
levels of NDRG1 and of the above-mentioned transcription factors
under normoxia (21% O2) and under extreme hypoxic
conditions (0.1% O2), which were applied for 1, 6 and 24
h. An additional analysis of protein levels was conducted 24 and 48
h, after re-oxygenation. NDRG1 protein expression was not
detectable under normoxic or hypoxic conditions for 1 h in all cell
lines analyzed (Fig. 1). Only
U87-MG cells exhibited low NDRG1 protein expression under these
conditions. However, this cell line exhibited an increase in NDRG1
protein expression after re-oxygenation (Fig. 1B). U373 cells displayed moderate
NDRG1 protein expression after 6 h of hypoxia (Fig. 1C), whereas NDRG1 expression was
increased in the GaMG cells under extended hypoxia of 24 h
(Fig. 1A). In all cell lines, high
NDRG1 expression remained stable after re-oxygenation (Fig. 1). Among the analyzed transcription
factors, Smad7 displayed a notably alternating response to hypoxia
and re-oxygenation. The Smad7 protein expression was undetectable
under normoxia, very low after 1 h of 0.1% O2,
disappeared again after 6 h hypoxia and then increased to the
highest concentration after 24 h hypoxia in GaMG cells (Fig. 1A). Re-oxygenation after 24 h of
hypoxia led to a decrease in the Smad7 protein level. In the U87-MG
cells, no Smad7 protein expression was detectable, except after 24
h of hypoxia, which led to moderate expression (Fig. 1A). Notably, U373 cells displayed a
different expression pattern. Smad7 expression increased after 1 h
of hypoxia with a moderate decrease after 6 h and a complete
disappearance after 24 h (Fig. 1C).
Under re-oxygenation or normoxia no Smad7 expression was detectable
in this cell line (Fig. 1C).
Hypoxic conditions caused an early increase of SP1 protein already
after 1 h in all cell lines, although the magnitude varied from
strong (U373 and U87-MG) to moderate (GaMG) (Fig. 1). Moreover, the SP1 protein
expression continued to increase and reached a peak after 6 h, to
be reduced again after 24 h of hypoxia. After re-oxygenation SP1
expression disappeared in the U373 cells, was low in GaMG and
moderate in U87-MG cells, resembling the normoxic expression levels
of these cell lines (Fig. 1C).
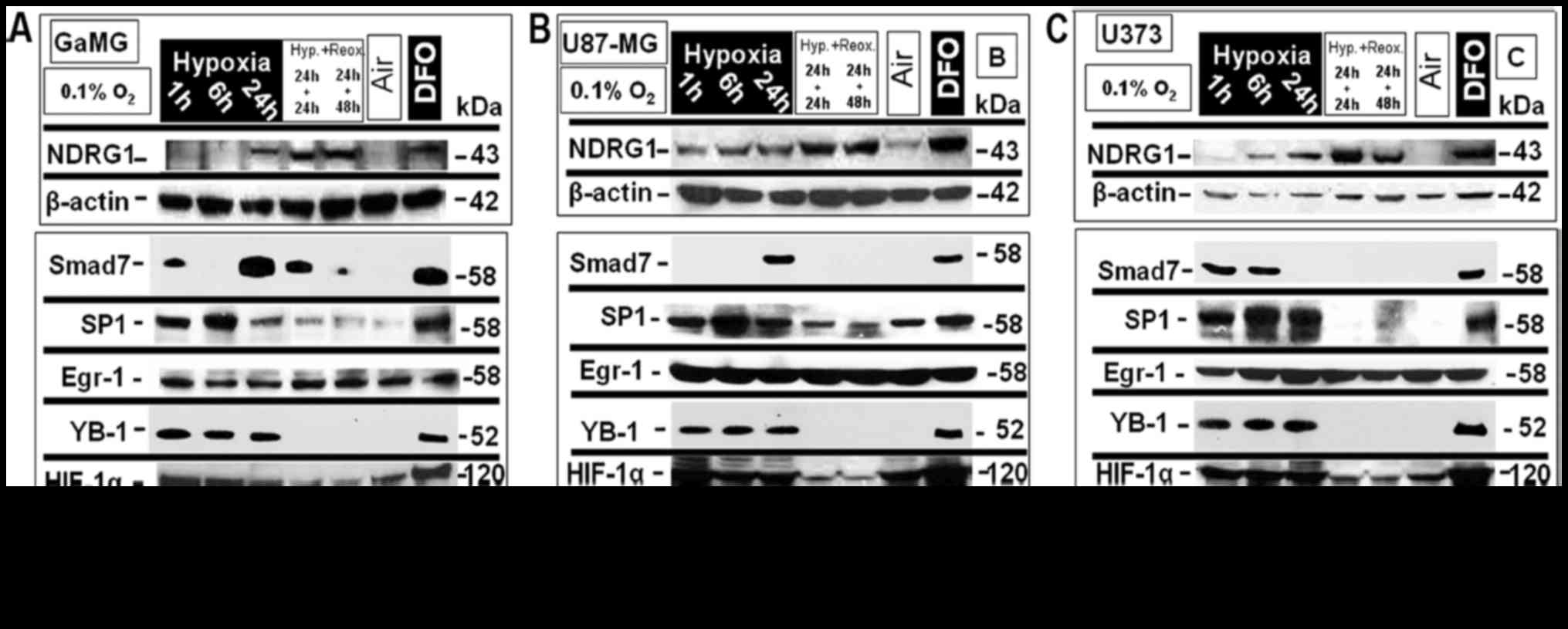 | Figure 1.In vitro protein expression
analysis in glioblastoma multiforme (GBM) cells under different
oxygenation conditions. The GBM cell lines, GaMG, U87-MG and U373,
were exposed to extreme hypoxic oxygenation conditions (0.1%
O2) for up to 24 h and then re-oxygenated for 24 h and
48 h. Normoxia and treatment with desferoxamine (DFO; 100 µM) for
24 h served as negative and positive controls, respectively. The
protein expression of NDRG1 and of the transcription factors Smad7,
SP1, Egr-1, YB-1, HIF-1α and CEBPα was analyzed by western
blotting. A clear O2 concentration- and time-dependent
expression regulation and also cell line-specific differences were
observed. (A) In GaMG, NDRG1 protein expression increase was only
significant after 24 h and at the highest expression level and
relatively stable upon re-oxygenation for a period of up to 48 h
after 24 h of hypoxia. Smad7 protein expression (3rd row from
above) pattern displayed a significant protein expression level
increase after only 1 h of exposure to extreme hypoxia which
decreased to the basal expression level after 6 h of hypoxia and
again increased to a maximum level after 24 h of hypoxia, while,
re-oxygenation resulted in a significant decrease in the expression
level. SP-1 expression pattern (4th row from above) showed a
significant increase in the expression level after only 1 h of
extreme hypoxia that reached its maximum after 6 h of hypoxia and
decreased to expression levels below that of the previous ones, but
still higher than the basal expression level and of 24 h of
hypoxia, while re-oxygenation up to 48 h after 24 h of hypoxia
resulted in a decrease to a lower level slightly higher than those
at basal expression level. Egr-1 expression level was stable in
response to the different oxygenation conditions. YB-1 expression
(6th row from above) was significantly increased after 1 h of
hypoxia and was relatively stable up to 24 h of extreme hypoxia;
while re-oxygenation for 24 and 48 h, respectively after 24 h of
hypoxia resulted in the decrease to an expression level similar to
the basal expression level. HIF-1α protein expression level (7th
row from above) increased after 1 h of extreme hypoxic exposure and
was stable up to 24 h of extreme hypoxia and its level decreased to
a similar pattern as the basal expression level under normoxic
conditions. CEBPα showed an expression pattern similar to that of
YB-1 expression with a lower protein level expressed. (B) In
U87-MG, NDRG1 expression increased significantly after 1 h of
extreme hypoxia and continued to increase in its expression level
up to 24 h of hypoxia and 24 and 48 h of re-oxygenation after 24 h
of hypoxia and returned to basal expression level under nomoxic
oxygenation conditions. Smad7 expression increased only after 24 h
of hypoxia reaching its maximum. SP-1 expression pattern was
similar to that in GaMG, but with a higher protein expression
level. Egr-1 expression level was stable in response to the
different oxygenation conditions examined. YB-1 and HIF-1α
expression was displayed in a comparable pattern as in GaMG, with a
higher protein level of HIF-1α, while CEBPα expression increased
significantly only after 1 h of hypoxia and was at a stable level
for up to 24 h of hypoxia but it decreased significantly upon
re-oxygenation after 24 h of hypoxia. (C) In U373, NDRG1 expression
was significantly increased after 1 h of extreme hypoxia and
maintained an increase up to 24 h of hypoxia and 24 and 48 h of
re-oxygenation. Smad7 expression level increased after only 1 h of
hypoxia and was stable at 6 h of hypoxia but started to decrease to
basal expression level after 24 h of hypoxia. SP-1 was expressed at
a significant level after only 1 h of hypoxia and increased in its
expression level gradually until 24 h, followed by a decline to
basal level after 24 h of re-oxygenation and a relative increase
after 48 h of re-oxygenation. Egr-1 again showed no change in its
displayed pattern in response to the alternating oxygenation
conditions. YB-1 and HIF-1α expression patterns were the same as
the corresponding protein as was the case in U87-MG with a higher
protein expression rate for HIF-1α, and CEBPα expression level
increased significantly after 1 h of extreme hypoxic exposition and
decreased after 6 h and increased again after 24 h of extreme
hypoxia to its maximum level, and decreased again upon
re-oxygenation to its basic expression level. One representative
experiment out of three independent repetitions is shown. The
housekeeping genes β-actin and β-tubulin served as loading
controls. |
All three examined cells lines expressed Egr-1
protein with no alterations caused by hypoxia or re-oxygenation
(Fig. 1).YB-1 was not expressed
under normoxia or after re-oxygenation, but hypoxic conditions
induced YB-1 protein expression already after 1 h, which remained
stable for at least 24 h in all three cell lines (Fig. 1). Hypoxic conditions led to an early
increase in HIF-1α protein expression in all three cell lines.
Re-oxygenation then caused a considerable decrease of protein
expression, but not in its complete disappearance (Fig. 1).
CEBPα displayed a similar pattern under hypoxic
conditions with higher expression levels in U87-MG and U373 cells
compared to GaMG. Re-oxygenation led to the disappearance of
detectable CEBPα protein in GaMG cells, while in U373 and U87-MG
residual protein still was detectable with highest concentrations
in the U87-MG cells (Fig. 1B).
Desferoxamine (DFO) was used as a control and significantly induced
the protein expression of all proteins analyzed (Fig. 1).
Time- and oxygen
concentration-dependent mRNA expression of NDRG1 and its potential
transcription factors
The mRNA expression levels of CEBPα, HIF-1α, EGR-1,
Smad7, YB-1 and SP1 in the human glioma cell lines U87-MG, U373 and
GaMG cultivated under different oxygenation conditions are
illustrated in Fig. 2.
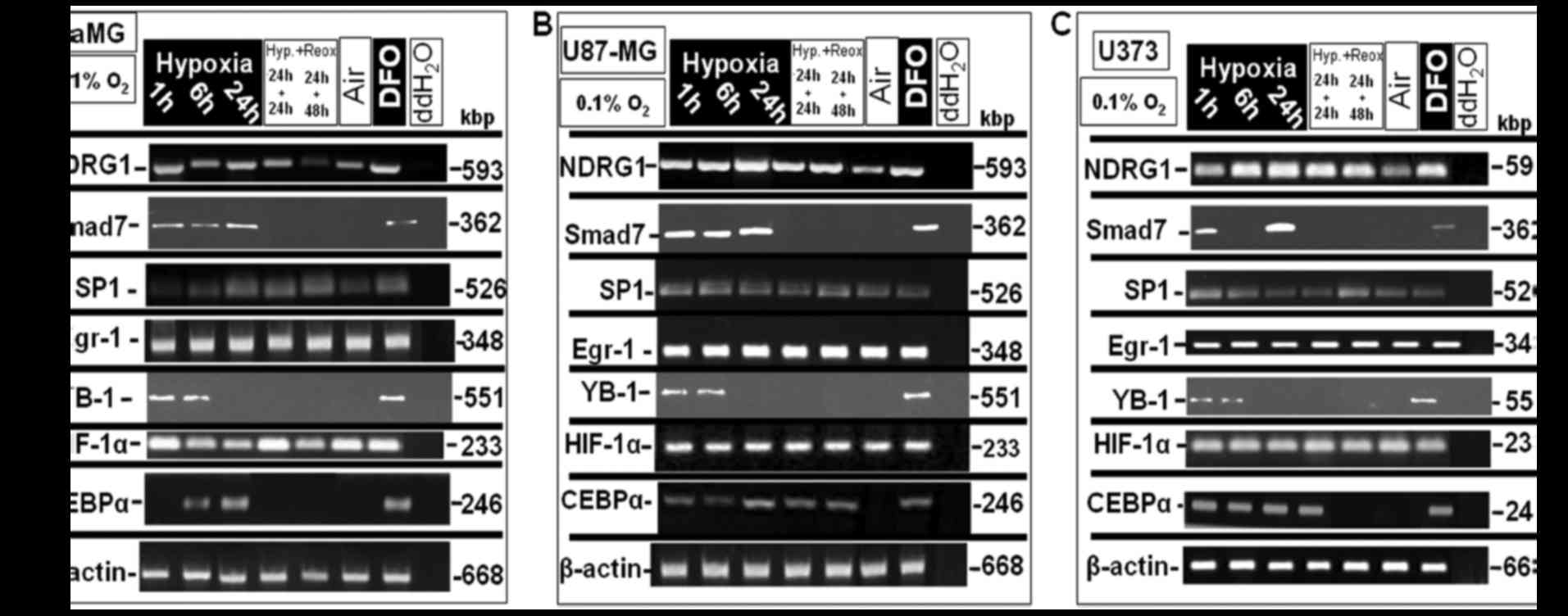 | Figure 2.In vitro mRNA expression
analysis in glioblastoma multiforme (GBM) cells under different
oxygenation conditions. The GBM cell lines GaMG, U87-MG and U373
were exposed to extreme hypoxic oxygenation conditions (0.1%
O2) for up to 24 h and then were re-oxygenated for 24
and 48 h. Normoxia and treatment with 100 µM DFO for 24 h served as
negative and positive controls, respectively. mRNA-lysates were
then analyzed by semi-quantitative RT-PCR for the expression of
NDRG1 and of the transcription factors Smad7, SP1, Egr-1, YB-1,
HIF-1α and CEBP-α. (A) In GaMG, NDRG1 mRNA expression (1st row,
left column) increased after just 1 h of extreme hypoxic exposure
and remained at a stable level up to 24 h, and 24 h of
re-oxygenation after 24 h of hypoxia, but declined to an expression
level nearly to that expressed under normoxia. Smad7 mRNA
expression level (2nd row) increased after 1 h of hypoxia and was
stable up to 24 h of hypoxia and declined upon re-oxygenation after
24 h to the basal expression level. SP-1 expression (3rd row),
increased to its maximum level after 24 h of hypoxia and was stable
upon re-oxygenation for 24 and 48 h, respectively, after 24 h of
hypoxia. Egr-1 expression (4th row) was at a stable level under the
different conditions examined. YB-1 expression level (5th row, left
column) reached its maximum level after just 1 h of hypoxic
exposition and declined to an expression level similar to that
noted under normoxia. HIF-1α mRNA (6th row) was stable under the
different aeration conditions examined. CEBPα expression (7th row)
increased significantly after 6 h of hypoxia reaching its maximum
after 24 h and declined to the basal expression level upon
re-oxygenation. (B) In U87-MG, NDRG1 (1st row, middle column)
reached its maximum mRNA expression level after only 1 h of hypoxia
and was at a stable level under all examined aeration levels and
declined to a basal expression level under normoxia. Smad7 mRNA
expression (2nd row) behaved in a similar way as in GaMG cells with
a higher expression rate. SP-1 mRNA showed a significant expression
pattern (3rd row), starting with a significant increase in to a
high expression level after 1 h reaching its maximum level after 6
h with a relative decline after 24 h of hypoxia and 24 h of
re-oxygenation and after hypoxia and again a slight increase at 48
h of re-oxygenation after 24 h of hypoxia. Egr-1 expression (4th
row) was stable with a uniform pattern during the different
aeration conditions examined. YB-1 (5th row) reached its maximum
expression level after just 1 h of hypoxic exposition, and was
stable after 6 h and declined to a basal expression level after 24
h of hypoxia and re-oxygenation after hypoxia. HIF-1α (6th row) was
expressed at a stable level under all conditions examined. CEBPα
mRNA expression increased significantly after only 1 h of hypoxia,
then declined to one half of the expression after 6 h of hypoxia
and reached its maximum after 24 h and declined to an expression
level similar to a basal expression level upon re-oxygenation after
hypoxia. (C) In U373, NDRG1 mRNA (1st row), reached its maximum
level after only 6 h of hypoxia and was stable under the different
aeration conditions examined. Smad7 mRNA (2nd row) showed a notable
expression pattern, showing a significant increase in the
expression level after 1 h of hypoxia, that declined after 6 h and
increased again to its maximum expression level after 24 h.
Re-oxygenation after hypoxia resulted in minimization to a basal
expression level. SP-1 expression (3rd row) was also interesting as
it reached its maximum level after only 1 h of hypoxia and
continuing with a lower expression rate for 6 and 24 h of hypoxia
and 24 h re-oxygenation after 24 h of hypoxia and reaching again
the maximal expression level after 48 h of re-oxygenation after 24
h of hypoxia. Egr-1 mRNA (4th row) displayed a constant expression
pattern under all oxygenation conditions examined. YB-1mRNA (5th
row) reached it maximum after 1 h and was stable up to 6 h and
declined after 24 h as well after re-oxygenation. HIF-1α (6th row)
showed a constant expression pattern under the different
oxygenation conditions examined. CEBPα mRNA (7th row), was
significantly expressed at its maximum expression level after only
1 h of hypoxia which kept constant up to 24 h of hypoxia and 24 h
of re-oxygenation after hypoxia and declined towards the basal
expression level upon re-oxygenation for 48 h after 24 h of
hypoxia. One representative experiment out of three independent
repetitions is shown. The housekeeping gene β-actin served as a
loading control. |
NDRG1 mRNA was detectable under all conditions in
all cell lines analyzed. However, hypoxia procured a relative
increase of mRNA levels with time. Re-oxygenation led to a slight
reduction after 48 h. NDRG1 mRNA expression was highest in U373
cells, followed by U87-MG and lowest in GaMG, which also was most
sensitive to re-oxygenation (Fig.
2).
Smad7 mRNA expression was clearly dependent upon the
oxygenation conditions. It was not detectable under normoxia or
after re-oxygenation, but highly induced already after 1 h of
hypoxia. Notably, Smad7 mRNA expression disappeared after 6 h of
hypoxia in the U373 cells and re-appeared after 24 h to a higher
level then after 1 h (Fig. 2C).
U87-MG cells displayed the strongest mRNA expression and GaMG the
lowest level (Fig. 2A and B).
SP1 mRNA expression also showed significant
features. In U87-MG cells, SP1 mRNA was moderately expressed and
without changes under all tested conditions (Fig. 2B). However, in GaMG cells there was
an increase over time under hypoxia, which remained stable at a
high level after re-oxygenation (Fig.
2A). In contrast, the U373 cells had increased SP1 mRNA
expression noticeably after 1 h of hypoxia, which then decreased
back to basal levels over 24 h. The same basal expression was
visible after 24 h of hypoxia followed by 24 h of re-oxygenation.
Surprisingly, 48 h re-oxygenation caused the strongest SP1 mRNA
expression in this experiment (Fig.
2C).
The same Egr-1 mRNA expression strength was
detectable under all conditions in all three cell lines examined
(Fig. 2).
In contrast, YB-1 mRNA expression was induced by 1
and 6 h of hypoxia, but disappeared after 24 h of hypoxia. It was
not detectable under normoxia or after re-oxygenation.
HIF-1α mRNA was expressed at a constant level under
all oxygenation conditions in the three cell lines, which is a
common expression feature of HIF-1α (Fig. 2).
CEBPα displayed a cell line-specific expression
pattern. In all three cell lines there was expression under
normoxia (Fig. 2). Hypoxia induced
CEBPα expression. In the GaMG cells there was very low expression
detectable after 1 h hypoxia, which considerably increased after 6
h and reached the highest levels after 24 h. CEBPα expression then
disappeared under re-oxygenation (Fig.
2A). In contrast, the U87-MG cells exhibited increased CEBPα
mRNA expression strongly already after 1 h of hypoxia, then after 6
h the cells showed a decrease in expression by 50% to increase the
expression again after 24 h. The expression remained high even
after re-oxygenation (Fig. 2B). In
the U373 cells, there was a significant increase in CEBPα mRNA
expression after 1 h of hypoxia, which remained constant over 24 h
and only disappeared after 48 h of re-oxygenation (Fig. 2C).
DFO was able to significantly induce the mRNA
expression of all tested factors in the three cell lines examined
(Fig. 2).
Discussion
The present study showed that NDRG1 expression was
associated with long-term hypoxia, but not with short-term hypoxia
in human GBM cells. NDRG1 expression may be under the control of
HIF-1α in addition to the other transcription factors examined in a
so-called expression-regulated time point. The significant increase
in NDRG1 expression in hypoxia-exposed GBM cells confirms our
previous findings of NDRG1 as a hypoxia-associated molecule with
overexpression in tumor cells (21). A common survival strategy of tumor
cells at the beginning of hypoxia includes Pasteur-effect
remission, in this case it appears that HIF-1α binds to HER
sequences of hypoxia-responsive genes (42) and thereby stimulates glycolytic
transporter proteins and enzyme expression (43).
The NDRG1 gene contains two HREs within its
non-coding sequence (24),
indicating that its regulation is HIF-1α-dependent (44). These data support our previous
findings that NDRG1 expression is induced by HIF-1α in human tumor
cells (8). In addition, the HIF-1α
expression precedes NDRG1 expression, indicating that it may be a
prerequisite for NDRG1 upregulation since it is its major
transcriptional regulator (8,21,45,46).
The same holds true for other HIF-1α-regulated genes such as CA9
(47). Therefore, we suggest NDRG1
as a marker for hypoxic regions within a tumor mass (8). It may also be of prognostic relevance,
since the survival rate of patients without NDRG1 expression was
lower than that of patients bearing NDRG1-positive cells (13). In addition, high expression level of
NDRG1 does not correlate with the overall patient survival
(48).
In two of the examined GBM cell lines, NDRG1 protein
expression was undetectable under normoxic conditions. Although
increased NDRG1 protein expression has been reported for different
types of tumor cells such as human HCC and colorectal tumors
(11,49), other studies showed a decrease in
its expression level such as in metastasizing colon cancer cell
lines (50). Low-grade astrocytoma
(LGA) displayed a lower level of NDRG1 mRNA and protein expression
than GBM samples from patients (8).
However, another study showed that the NDRG1 mRNA and protein
expression was reduced in GBM compared to normal brain tissue
(13). These observations suggest
that the regulation of NDRG1 may be cancer cell-specific and also
dependent on the oxygenation status of the tumor tissue.
In the present study, we showed that NDRG1 mRNA
expression was downregulated upon re-oxygenation, which is in
agreement with a previous study concerning decreased NDRG1 mRNA
expression in the breast cancer cell line MCF-7 after
re-oxygenation (51). Therefore,
NDRG1 is considered to be an O2-responsive gene, linked
to tumor adaptation towards re-oxygenation (51,52).
However, our western blot results did not support this notion, and
suggest posttranscriptional regulation of NDRG1 expression. The
stability of NDRG1 after re-oxygenation following long-time
exposure to hypoxic oxygenation conditions may be related to both,
post transcriptional and post translational modifications. Based on
our results and other research data we can postulate that a post
transcriptional modification may be achieved by NDRG1 interaction
with the HSP90 protein (53). The
mTOR signaling pathway may also be involved, since the downstream
regulator CCR4-NOT may upregulate NDRG1 (54,55).
Such post transcriptional regulation may act along with post
translational modification, such as e.g. sumoylation of HIF-1α.
HIF-1α is destabilized and as a consequence the transcription rate
of NDRG1 is reduced. In contrast, SUMO protease-1 contributes to
HIF-1α stability and thereby increases the NDRG1 transcription rate
(55–57). Such modifications may contribute to
the modulatory process that in the end renders NDRG1 mRNA for
protein synthesis.
Egr-1 displayed a constant expression pattern at the
mRNA and protein level, irrespective of the tested oxygenation
conditions. This suggests a constitutive functional role for this
transcription factor under various physiological conditions in
cancer cells (58).
In contrast, the expression of the transcription
factors HIF-1α, SP1, CEBPα, YB-1 and Smad7 widely correlated with
those of NDRG1. Particularly, the nuclear mRNA levels of HIF-1α,
SP1 and C/EBP-α were closely correlated with those of NDRG1. It has
been shown that hypoxic conditions raise the expression of these
transcription factors in nuclear extracts (59), suggesting that the
hypoxia-stimulated upregulation of NDRG1 may be mainly mediated by
a combined effect of these factors and to a lesser extend by Smad7
and YB-1. A cooperative mechanism, as suggested in the present
study, has been shown to promote the regulation of genes necessary
for cell adaptation to varying microenvironments such as during
starvation and hypoxia (60).
Indeed, NDRG1 promoter studies revealed that a SP1 site partially
is responsible for the VHL-induced suppression of NDRG1 (61,62).
The analysis of the murine NDRG1 promoter revealed
an overlap between Egr-1 and SP1 binding motifs (62). Although, it has previously been
shown that NDRG1 expression during hypoxia is initiated by HIF-1α
(63), SP1 triggers the hypoxic
promoter activation independently (64) or by interaction with HIF-1α
(65). In the present study, SP1
protein expression was increased during hypoxia in the human glioma
cell lines. The highest level was detected after 6 h of extreme
hypoxia. Most of the transcription factors regulating metabolic
events in cancer, including HIF-1α, have a binding site for SP1 in
their promoter region (66). HIF-1α
expression has been shown to be SP1-dependent in human prostate
cancer cells (67), suggesting a
crucial regulatory role in cancer metabolism as well as during
hypoxia.
We showed an upregulation of CEBPα in human GBM
cells in response to hypoxia. However, these results are not
consistent with a previous study using breast cancer cells, which
showed that the exposure of these cells to 1% O2
downregulates the CEBPα production (68) and transcription (69). These differences may be explained by
the severity of hypoxia, as in our experiments the GBM cells were
exposed to extreme hypoxic conditions (0.1% O2) for up
to 24 h. It has been reported that the HIF-1α-bound HRE within the
CEBPα promoter region is essential for CEBPα transcriptional
downregulation under hypoxic conditions (62). In contrast, we observed a lack of
CEBPα expression in GBM cells under normoxic conditions. CEBPα
downregulation under normoxia/moderate hypoxia and its upregulation
during extreme hypoxia may play a role during progression.
Different signal transduction pathways are
controlled and modulated by different growth factors and
environmental events such as hypoxic oxygenation conditions
(26). The TGF-β pathway involves
the Smad protein family (27).
These proteins play an important role in triggering diseases
including for example fibrosis (28) and cirrhosis, leading to HCC
(29). NDRG1 overexpression could
inhibit TGF-β-induced effects via reducing both Smad2 and Smad3
expression (63). However, Smad7,
another member of this protein family, abrogates the TGF-β
signaling pathway by a regulative negative feedback loop (30), thereby, paving the physiological
conditions towards cancer disease manifestation, as HCC is an
example when this negative feedback loop is hindered, while the
transition to fibrosis, cirhosis and as a consequence HCC is
hindered when this negative feedback loop of Smad7 remains active
(31). Smad7 is regulated by
different signaling pathways (32).
IFNγ contributes to this regulatory scheme by inducing a member of
the cold shock protein family, the transcription factor YB-1
(30,70–73)
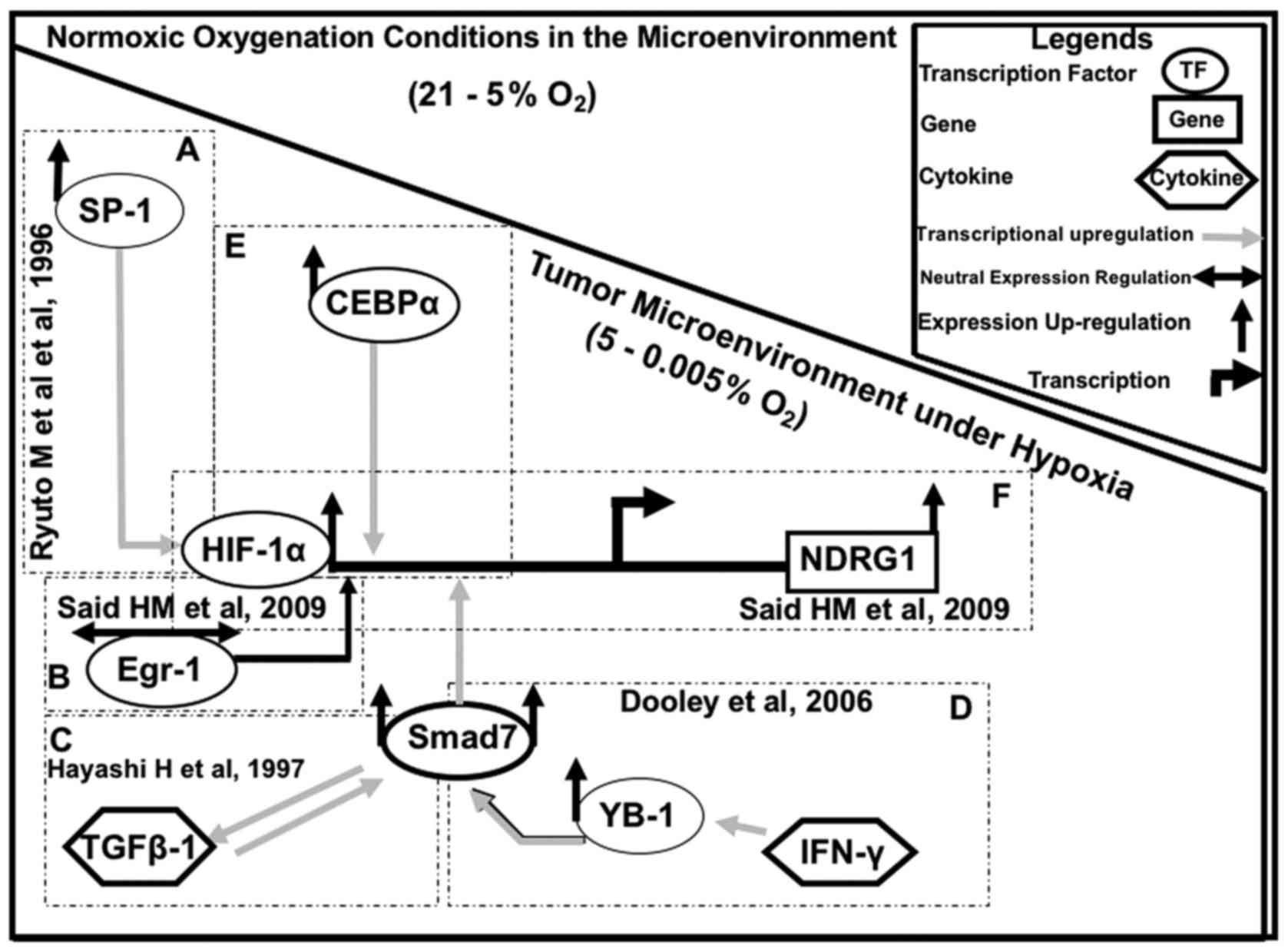
(Fig. 3).
Our results suggest that HIF-1α regulates NDRG1
expression in GBM cells in conjunction with SP1, CEBPα, YB-1 and
Smad7 under different physiological conditions, whereas Egr-1 may
act as a cofactor (Fig. 4). A very
similar regulatory mechanism may also exist for other
hypoxia-induced genes (7), such as
CA9, Epo and VEGF in human brain cancer (40,64).
These factors are also controlled by HIF-1α. However, other
co-factors may be involved. The knowledge concerning such
regulatory processes during cancer development is important for the
development and application of cancer treatment modalities.
In conclusion, we showed for the first time that
NDRG1 protein expression was significantly increased in GBM cells
under hypoxia and that there was a correlation with the protein
expression of HIF-1α, SP1, CEBPα, YB-1 and Smad7. We suggest that
SP1 regulates NDRG1 in GBM cells under different oxygenation
conditions in conjunction with HIF1α, CEBPα and, to a certain
extent, YB-1 and Smad7. NDRG1 and its potential transcription
factors therefore may represent target proteins for complementary
treatment approaches. Combined, they also may serve as potential
hypoxia tumor markers in gene expression analyses.
Acknowledgements
The authors are indebted to the Dokuz Eylul
University, Graduate Institute of Health Sciences (SBE) for
support. The present study was partially funded by means of the
University of Würzburg, Medical Faculty, Department of Radiation
Oncology. The authors are also grateful to the College of Medicine
and Medical Sciences, Arabian Gulf University, Manama, Kingdom of
Bahrain. We would like to thank Stefanie Gerngras and Siglinde
Kühnel for their technical assistance.
References
|
1
|
Theunissen PH and Blaauw G: Proliferating
cell nuclear antigen immunostaining and survival in cerebral
astrocytoma. Histopathology. 23:75–79. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kaye SB, Bagshawe KD, McElwain TJ and
Peckham MJ: Brain metastases in malignant teratoma: A review of
four years' experience and an assessment of the role of tumour
markers. Br J Cancer. 39:217–223. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Molendini L, Benassi MS, Magagnoli G,
Merli M, Sollazzo MR, Ragazzini P, Gamberi G, Ferrari C, Balladelli
A, Bacchini P, et al: Prognostic significance of cyclin expression
in human osteosarcoma. Int J Oncol. 12:1007–1011. 1998.PubMed/NCBI
|
|
4
|
Nam DH, Park K, Park C, Im YH, Kim MH, Lee
S, Hong SC, Shin HJ, Kim JH, Eoh W, et al: Intracranial inhibition
of glioma cell growth by cyclooxygenase-2 inhibitor celecoxib.
Oncol Rep. 11:263–268. 2004.PubMed/NCBI
|
|
5
|
Xie SS, Tan M, Lin HY, Xu L, Shen CX, Yuan
Q, Song XL and Wang CH: Overexpression of adenylate
cyclase-associated protein 1 may predict brain metastasis in
non-small cell lung cancer. Oncol Rep. 33:363–371. 2015.PubMed/NCBI
|
|
6
|
Angst E, Sibold S, Tiffon C, Weimann R,
Gloor B, Candinas D and Stroka D: Cellular differentiation
determines the expression of the hypoxia-inducible protein NDRG1 in
pancreatic cancer. Br J Cancer. 95:307–313. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Semenza GL: Involvement of oxygen-sensing
pathways in physiologic and pathologic erythropoiesis. Blood.
114:2015–2019. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Said HM, Stein S, Hagemann C, Polat B,
Staab A, Anacker J, Schoemig B, Theobald M, Flentje M and
Vordermark D: Oxygen-dependent regulation of NDRG1 in human
glioblastoma cells in vitro and in vivo. Oncol Rep. 21:237–246.
2009.PubMed/NCBI
|
|
9
|
Ellen TP, Ke Q, Zhang P and Costa M:
NDRG1, a growth and cancer related gene: Regulation of gene
expression and function in normal and disease states.
Carcinogenesis. 29:2–8. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Berger P, Young P and Suter U: Molecular
cell biology of Charcot-Marie-Tooth disease. Neurogenetics. 4:1–15.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chua MS, Sun H, Cheung ST, Mason V,
Higgins J, Ross DT, Fan ST and So S: Overexpression of NDRG1 is an
indicator of poor prognosis in hepatocellular carcinoma. Mod
Pathol. 20:76–83. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wangpu X, Yang X, Zhao J, Lu J, Guan S, Lu
J, Kovacevic Z, Liu W, Mi L, Jin R, et al: The metastasis
suppressor, NDRG1, inhibits ‘stemness’ of colorectal cancer via
down-regulation of nuclear β-catenin and CD44. Oncotarget.
6:33893–33911. 2015.PubMed/NCBI
|
|
13
|
Sun B, Chu D, Li W, Chu X, Li Y, Wei D and
Li H: Decreased expression of NDRG1 in glioma is related to tumor
progression and survival of patients. J Neurooncol. 94:213–219.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Strzelczyk B, Szulc A, Rzepko R, Kitowska
A, Skokowski J, Szutowicz A and Pawelczyk T: Identification of
high-risk stage II colorectal tumors by combined analysis of the
NDRG1 gene expression and the depth of tumor invasion. Ann Surg
Oncol. 16:1287–1294. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ando T, Ishiguro H, Kimura M, Mitsui A,
Kurehara H, Sugito N, Tomoda K, Mori R, Takashima N, Ogawa R, et
al: Decreased expression of NDRG1 is correlated with tumor
progression and poor prognosis in patients with esophageal squamous
cell carcinoma. Dis Esophagus. 19:454–458. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Maruyama Y, Ono M, Kawahara A, Yokoyama T,
Basaki Y, Kage M, Aoyagi S, Kinoshita H and Kuwano M: Tumor growth
suppression in pancreatic cancer by a putative metastasis
suppressor gene Cap43/NDRG1/Drg-1 through modulation of
angiogenesis. Cancer Res. 66:6233–6242. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nakamura K, Yasunaga Y, Segawa T, Ko D,
Moul JW, Srivastava S and Rhim JS: Curcumin down-regulates AR gene
expression and activation in prostate cancer cell lines. Int J
Oncol. 21:825–830. 2002.PubMed/NCBI
|
|
18
|
Bandyopadhyay S, Pai SK, Gross SC, Hirota
S, Hosobe S, Miura K, Saito K, Commes T, Hayashi S, Watabe M, et
al: The Drg-1 gene suppresses tumor metastasis in prostate cancer.
Cancer Res. 63:1731–1736. 2003.PubMed/NCBI
|
|
19
|
Bandyopadhyay S, Pai SK, Hirota S, Hosobe
S, Tsukada T, Miura K, Takano Y, Saito K, Commes T, Piquemal D, et
al: PTEN up-regulates the tumor metastasis suppressor gene Drg-1 in
prostate and breast cancer. Cancer Res. 64:7655–7660. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cangul H: Hypoxia upregulates the
expression of the NDRG1 gene leading to its overexpression in
various human cancers. BMC Genet. 5:272004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Said HM, Polat B, Staab A, Hagemann C,
Stein S, Flentje M, Theobald M, Katzer A and Vordermark D: Rapid
detection of the hypoxia-regulated CA-IX and NDRG1 gene expression
in different glioblastoma cells in vitro. Oncol Rep. 20:413–419.
2008.PubMed/NCBI
|
|
22
|
Arany Z, Huang LE, Eckner R, Bhattacharya
S, Jiang C, Goldberg MA, Bunn HF and Livingston DM: An essential
role for p300/CBP in the cellular response to hypoxia. Proc Natl
Acad Sci USA. 93:pp. 12969–12973. 1996; View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kallio PJ, Okamoto K, O'Brien S, Carrero
P, Makino Y, Tanaka H and Poellinger L: Signal transduction in
hypoxic cells: Inducible nuclear translocation and recruitment of
the CBP/p300 coactivator by the hypoxia-inducible factor-1alpha.
EMBO J. 17:6573–6586. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Le NT and Richardson DR: Iron chelators
with high antiproliferative activity up-regulate the expression of
a growth inhibitory and metastasis suppressor gene: A link between
iron metabolism and proliferation. Blood. 104:2967–2975. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Salnikow K, Costa M, Figg WD and
Blagosklonny MV: Hyperinducibility of hypoxia-responsive genes
without p53/p21-dependent checkpoint in aggressive prostate cancer.
Cancer Res. 60:5630–5634. 2000.PubMed/NCBI
|
|
26
|
Kockar F, Yildrim H, Sagkan RI, Hagemann
C, Soysal Y, Anacker J, Hamza AA, Vordermark D, Flentje M and Said
HM: Hypoxia and cytokines regulate carbonic anhydrase 9 expression
in hepatocellular carcinoma cells in vitro. World J Clin Oncol.
3:82–91. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kusanagi K, Inoue H, Ishidou Y, Mishima
HK, Kawabata M and Miyazono K: Characterization of a bone
morphogenetic protein-responsive Smad-binding element. Mol Biol
Cell. 11:555–565. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yamaguchi T, Matsuzaki K, Inokuchi R,
Kawamura R, Yoshida K, Murata M, Fujisawa J, Fukushima N, Sata M,
Kage M, et al: Phosphorylated Smad2 and Smad3 signaling: Shifting
between tumor suppression and fibro-carcinogenesis in chronic
hepatitis C. Hepatol Res. 43:1327–1342. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Neuzillet C, Tijeras-Raballand A, de
Mestier L, Cros J, Faivre S and Raymond E: MEK in cancer and cancer
therapy. Pharmacol Ther. 141:160–171. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Dooley S, Hamzavi J, Breitkopf K,
Wiercinska E, Said HM, Lorenzen J, Ten Dijke P and Gressner AM:
Smad7 prevents activation of hepatic stellate cells and liver
fibrosis in rats. Gastroenterology. 125:178–191. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Del Vecchio F, Gallo F, Di Marco A,
Mastroiaco V, Caianiello P, Zazzeroni F, Alesse E and Tessitore A:
Bioinformatics approach to predict target genes for dysregulated
microRNAs in hepatocellular carcinoma: Study on a
chemically-induced HCC mouse model. BMC Bioinformatics. 16:4082015.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu Y, Meyer C, Müller A, Herweck F, Li Q,
Müllenbach R, Mertens PR, Dooley S and Weng HL: IL-13 induces
connective tissue growth factor in rat hepatic stellate cells via
TGF-β-independent Smad signaling. J Immunol. 187:2814–2823. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Szalad A, Katakowski M, Zheng X, Jiang F
and Chopp M: Transcription factor Sp1 induces ADAM17 and
contributes to tumor cell invasiveness under hypoxia. J Exp Clin
Cancer Res. 28:1292009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ryuto M, Ono M, Izumi H, Yoshida S, Weich
HA, Kohno K and Kuwano M: Induction of vascular endothelial growth
factor by tumor necrosis factor alpha in human glioma cells.
Possible roles of SP-1. J Biol Chem. 271:28220–28228. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Minet E, Ernest I, Michel G, Roland I,
Remacle J, Raes M and Michiels C: HIF1A gene transcription is
dependent on a core promoter sequence encompassing activating and
inhibiting sequences located upstream from the transcription
initiation site and cis elements located within the 5′UTR. Biochem
Biophys Res Commun. 261:534–540. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Osada S, Takano K, Nishihara T, Suzuki T,
Muramatsu M and Imagawa M: CCAAT/enhancer-binding proteins alpha
and beta interact with the silencer element in the promoter of
glutathione S-transferase P gene during hepatocarcinogenesis. J
Biol Chem. 270:31288–31293. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Darlington GJ, Wang N and Hanson RW: C/EBP
alpha: A critical regulator of genes governing integrative
metabolic processes. Curr Opin Genet Dev. 5:565–570. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hubbi ME and Semenza GL: Regulation of
cell proliferation by hypoxia-inducible factors. Am J Physiol Cell
Physiol. 309:C775–C782. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Akslen LA, Andersen KJ and Bjerkvig R:
Characteristics of human and rat glioma cells grown in a defined
medium. Anticancer Res. 8:797–803. 1988.PubMed/NCBI
|
|
40
|
Said HM, Staab A, Hagemann C, Vince GH,
Katzer A, Flentje M and Vordermark D: Distinct patterns of hypoxic
expression of carbonic anhydrase IX (CA IX) in human malignant
glioma cell lines. J Neurooncol. 81:27–38. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Said HM, Katzer A, Flentje M and
Vordermark D: Response of the plasma hypoxia marker osteopontin to
in vitro hypoxia in human tumor cells. Radiother Oncol. 76:200–205.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Madan A and Curtin PT: A 24-base-pair
sequence 3′ to the human erythropoietin gene contains a
hypoxia-responsive transcriptional enhancer. Proc Natl Acad Sci
USA. 90:pp. 3928–3932. 1993; View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Payen VL, Brisson L, Dewhirst MW and
Sonveaux P: Common responses of tumors and wounds to hypoxia.
Cancer J. 21:75–87. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kitowska A and Pawełczyk T: N-myc
downstream regulated 1 gene and its place in the cellular
machinery. Acta Biochim Pol. 57:15–21. 2010.PubMed/NCBI
|
|
45
|
Wang Q, Li LH, Gao GD, Wang G, Qu L, Li JG
and Wang CM: HIF-1α up-regulates NDRG1 expression through binding
to NDRG1 promoter, leading to proliferation of lung cancer A549
cells. Mol Biol Rep. 40:3723–3729. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Salnikow K, Davidson T, Zhang Q, Chen LC,
Su W and Costa M: The involvement of hypoxia-inducible
transcription factor-1-dependent pathway in nickel carcinogenesis.
Cancer Res. 63:3524–3530. 2003.PubMed/NCBI
|
|
47
|
Said HM, Hagemann C, Staab A, Stojic J,
Kühnel S, Vince GH, Flentje M, Roosen K and Vordermark D:
Expression patterns of the hypoxia-related genes osteopontin, CA9,
erythropoietin, VEGF and HIF-1alpha in human glioma in vitro and in
vivo. Radiother Oncol. 83:398–405. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Blaes J, Weiler M, Sahm F, Hentschel B,
Osswald M, Czabanka M, Thomé CM, Schliesser MG, Pusch S, Luger S,
et al: NDRG1 prognosticates the natural course of disease in WHO
grade II glioma. J Neurooncol. 117:25–32. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wang Z, Wang F, Wang WQ, Gao Q, Wei WL,
Yang Y and Wang GY: Correlation of N-myc downstream-regulated gene
1 overexpression with progressive growth of colorectal neoplasm.
World J Gastroenterol. 10:550–554. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Guan RJ, Ford HL, Fu Y, Li Y, Shaw LM and
Pardee AB: Drg-1 as a differentiation-related, putative metastatic
suppressor gene in human colon cancer. Cancer Res. 60:749–755.
2000.PubMed/NCBI
|
|
51
|
Lai LC, Su YY, Chen KC, Tsai MH, Sher YP,
Lu TP, Lee CY and Chuang EY: Down-regulation of NDRG1 promotes
migration of cancer cells during reoxygenation. PLoS One.
6:e243752011. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Luo EC, Chang YC, Sher YP, Huang WY,
Chuang LL, Chiu YC, Tsai MH, Chuang EY and Lai LC: MicroRNA-769-3p
down-regulates NDRG1 and enhances apoptosis in MCF-7 cells during
reoxygenation. Sci Rep. 4:59082014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Banz VM, Medová M, Keogh A, Furer C,
Zimmer Y, Candinas D and Stroka D: Hsp90 transcriptionally and
post-translationally regulates the expression of NDRG1 and
maintains the stability of its modifying kinase GSK3beta. Biochim
Biophys Acta. 1793:1597–1603. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Dominick G, Bowman J, Li X, Miller RA and
Garcia GG: mTOR regulates the expression of DNA damage response
enzymes in long-lived Snell dwarf, GHRKO, and PAPPA-KO mice. Aging
Cell. 16:52–60. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Khaliullina H, Love NK and Harris WA:
Nutrient-deprived retinal progenitors proliferate in response to
hypoxia: Interaction of the HIF-1 and mTOR pathway. J Dev Biol.
4:172016. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Berta MA, Mazure N, Hattab M, Pouysségur J
and Brahimi-Horn MC: SUMOylation of hypoxia-inducible factor-1alpha
reduces its transcriptional activity. Biochem Biophys Res Commun.
360:646–652. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Cheng J, Kang X, Zhang S and Yeh ET:
SUMO-specific protease 1 is essential for stabilization of
HIF1alpha during hypoxia. Cell. 131:584–595. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Gosslar U, Schmid RM and Holzmann B:
Regulation of Egr-1-dependent gene expression by the C-terminal
activation domain. Biochem Biophys Res Commun. 255:208–215. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Huang RP, Fan Y, Ni Z, Mercola D and
Adamson ED: Reciprocal modulation between Sp1 and Egr-1. J Cell
Biochem. 66:489–499. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Koizume S, Ito S, Nakamura Y, Yoshihara M,
Furuya M, Yamada R, Miyagi E, Hirahara F, Takano Y and Miyagi Y:
Lipid starvation and hypoxia synergistically activate ICAM1 and
multiple genes in an Sp1-dependent manner to promote the growth of
ovarian cancer. Mol Cancer. 14:772015. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Turkoglu SA and Kockar F: SP1 and USF
differentially regulate ADAMTS1 gene expression under normoxic and
hypoxic conditions in hepatoma cells. Gene. 575:48–57. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Zhang P, Tchou-Wong KM and Costa M: Egr-1
mediates hypoxia-inducible transcription of the NDRG1 gene through
an overlapping Egr-1/Sp1 binding site in the promoter. Cancer Res.
67:9125–9133. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Salnikow K, Kluz T, Costa M, Piquemal D,
Demidenko ZN, Xie K and Blagosklonny MV: The regulation of hypoxic
genes by calcium involves c-Jun/AP-1, which cooperates with
hypoxia-inducible factor 1 in response to hypoxia. Mol Cell Biol.
22:1734–1741. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Lee M, Song SU, Ryu JK and Suh JK:
Sp1-dependent regulation of the tissue inhibitor of
metalloproteinases-1 promoter. J Cell Biochem. 91:1260–1268. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Miki N, Ikuta M and Matsui T:
Hypoxia-induced activation of the retinoic acid receptor-related
orphan receptor alpha4 gene by an interaction between
hypoxia-inducible factor-1 and Sp1. J Biol Chem. 279:15025–15031.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Archer MC: Role of sp transcription
factors in the regulation of cancer cell metabolism. Genes Cancer.
2:712–719. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Koshikawa N, Hayashi J, Nakagawara A and
Takenaga K: Reactive oxygen species-generating mitochondrial DNA
mutation up-regulates hypoxia-inducible factor-1alpha gene
transcription via phosphatidylinositol 3-kinase-Akt/protein kinase
C/histone deacetylase pathway. J Biol Chem. 284:33185–33194. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Seifeddine R, Fulchignoni-Lataud MC and
Massaad-Massade L: Down-regulation of C/EBPalpha in breast cancer
cells by hypoxia-estrogen combination is mainly due to hypoxia.
Anticancer Res. 29:1227–1231. 2009.PubMed/NCBI
|
|
69
|
Seifeddine R, Dreiem A, Blanc E,
Fulchignoni-Lataud MC, Le Frère Belda MA, Lecuru F, Mayi TH, Mazure
N, Favaudon V, Massaad C, et al: Hypoxia down-regulates
CCAAT/enhancer binding protein-alpha expression in breast cancer
cells. Cancer Res. 68:2158–2165. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Gery S, Tanosaki S, Bose S, Bose N,
Vadgama J and Koeffler HP: Down-regulation and growth inhibitory
role of C/EBPalpha in breast cancer. Clin Cancer Res. 11:3184–3190.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Chen Z, Zhang D, Yue F, Zheng M, Kovacevic
Z and Richardson DR: The iron chelators Dp44mT and DFO inhibit
TGF-β-induced epithelial-mesenchymal transition via up-regulation
of N-Myc downstream-regulated gene 1 (NDRG1). J Biol Chem.
287:17016–17028. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Hayashi M, Martinez OM, Krams SM, Burns W
and Esquivel CO: Cytokines are involved in the rejection of small
intestine allografts. Transplant Proc. 29:18021997. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Dooley S, Said HM, Gressner AM, Floege J,
En-Nia A and Mertens PR: Y-box protein-1 is the crucial mediator of
antifibrotic interferon-gamma effects. J Biol Chem. 281:1784–1795.
2006. View Article : Google Scholar : PubMed/NCBI
|