Introduction
Gestational trophoblastic disease (GTD) is a group
of diseases that has its origins in trophoblasts. The common
characteristics of GTD are abnormal proliferation of atypical
trophoblasts and secretion of human chorionic gonadotropin (hCG).
GTD consists of hydatidiform mole, invasive mole, choriocarcinoma,
placental site trophoblastic tumour (PSTT) and epithelioid
trophoblastic tumour (ETT) (1).
Hydatidiform mole is an abnormal pregnancy resulting from impaired
fertilization. Other GTDs are grouped under gestational
trophoblastic neoplasia (GTN) which has the potential for local
invasion and metastasis (1,2). Common GTNs are invasive mole and
choriocarcinoma which respond to chemotherapy. However, ~10% of
choriocarcinoma patients have a poor prognosis, particularly when
they have metastasis other than pulmonary metastasis (3,4). hCG
is a unique reliable marker for monitoring GTD (2,4).
Structural changes of branched N-glycans on
certain glycoproteins are mediated by a group of enzymes named
N-acetylglucosaminyltransferases (GnTs) which add
N-acetylglucosamine (GlcNAc) branch antennae to the mannose
core of N-glycans. These modifications of N-glycans
alter the biological functions of glycoproteins and eventually, the
properties of cells containing these glycoproteins (5). N-glycan modifications mediated
by GnTs greatly influence the invasion and migration abilities of
cancer cells (6,7). N-acetylglucosaminyltransferase
IV (GnT-IV) catalyses the formation of β1,4GlcNAc branches on the
mannose core of N-glycans (8,9). Many
previous studies showed that GnT-IV is involved in the progression
of cancers of the liver, pancreas, kidney and colon (10–13).
hCG is a glycoprotein hormone composed of α- and
β-subunits. α-subunit has two asparagine-linked (N-linked)
glycosylation sites and β-subunit has two N-linked and four
serine-linked (O-linked) glycosylation sites (14). Previous studies revealed that
N-glycans of hCG have many important biological functions
(14–16) and that they are differentially
modified in physiological and pathological states (17,18).
hCG from normal pregnant women and patients with hydatidiform mole
contains only mono- and bi-antennary branched N-glycans. On
the other hand, additional tri-antennary branched N-glycans
are present on hCG of invasive mole and choriocarcinoma patients,
and abnormal bi-antennary N-glycan is detected on hCG from
choriocarcinoma. The tri-antennary and abnormal bi-antennary
branched N-glycans are biosynthesized by the action of
GnT-IV on mono- and bi-antennary N-glycans. Takamatsu et
al (19) confirmed the elevated
activity of GnT-IV and higher expression level of GnT-IVa in
choriocarcinoma cells compared to those in normal placentas,
whereas the mRNA of GnT-IVb, another isoenzymatic form, was
expressed at the same level in choriocarcinoma and placentas. Niimi
et al (20) reported that
GnT-IVa protein expression was strong in GTN (invasive mole and
choriocarcinoma) and suggested that GnT-IVa might be involved in
invasion of choriocarcinoma, using GnT-IVa suppression model. For
further understanding of the role of GnT-IVa in choriocarcinoma, we
established a choriocarcinoma cell line which overexpresses GnT-IVa
and studied its target proteins for GnT-IVa glycosylation which
contribute to the malignancy of choriocarcinoma.
Materials and methods
Tissue collection and processing
The present study was approved by the ethics
committee of Nagoya University Graduate School of Medicine. Written
informed consent was obtained before surgery from all patients with
GTD for the use of clinical samples. Tissue samples from four cases
of invasive mole and seven cases of choriocarcinoma were used for
this study. Tissue samples were fixed in 10% formaldehyde, embedded
in paraffin and routinely stained with haematoxylin and eosin for
histological examination. All sections were histologically assessed
by pathologists in our institute.
Cell lines and culture
Human choriocarcinoma cell line Jar was purchased
from the American Type Culture Collection (ATCC; Manassas, VA, USA)
and maintained in RPMI-1640 medium (Sigma-Aldrich, St. Louis, MO,
USA) supplemented with 10% fetal bovine serum (FBS; Gibco, Grand
Island, NY, USA), penicillin (100 U/ml), streptomycin (100 µg/ml),
at 37°C in a humidified atmosphere of 5% CO2 and 95%
air.
Immunohistochemistry
Immunohistochemical staining for GnT-IVa protein was
performed using a monoclonal anti-GnT-IVa antibody (sc-100785;
Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA) at a dilution
of 1:400 as previously described (20,21).
Immunohistochemical staining for Griffonia simplicifolia
lectin-II (GSL-II) was performed by a previous method with
modification (22). Sections of 4
µm thickness were deparaffinised, rehydrated and washed with
phosphate-buffered saline (PBS). Endogenous peroxidase activity was
blocked by incubating the sections in 0.3% hydrogen peroxide in
100% methanol. The sections were washed twice with PBS and once
with 0.05 M Tris-buffered saline (TBS). For antigen retrieval, the
sections were treated with 0.1% trypsin solution in TBS containing
0.1% CaCl2. Non-specific lectin-protein binding was
blocked by incubating the sections in 5% skim milk in PBS. The
sections were then incubated overnight with biotinylated GSL-II
(B-1215; Vector Laboratories, Inc., Burlingame, CA, USA) at a
dilution of 1:400 in TBS containing 1% bovine serum albumin (BSA)
and 10 mM CaCl2 at 4°C. After rinsing, the sections were
immunostained using Vectastain Elite ABC kit (Vector Laboratories)
for 30 min according to the manufacturers instructions.
3,3-diaminobenzidine tetrahydrochloride reagent (Nichirei, Corp.,
Tokyo, Japan) was used for colour development.
Construction of an overexpression
vector of GnT-IVa and retroviral transduction
Construction of GnT-IVa-encoding retroviral vector
was performed as previously described (23). Briefly, GnT-IVa was PCR
amplified using primers (F, 5-AA AAAGCAGGCTCCACCATGAGGCTCCGCAAT
GG-3 and R, 5-AGAAAGCTGGGTCAGTTGGTGGCTTTTTTAATA TGAATC-3) from the
vector GnT-IVa/pSVL which was provided by Ogri et al
(24). After confirming the
sequence by subcloning the product into an entry vector,
GnT-IVa was transferred to the destination vector,
pCLMSCV-puro-Rfa, using the Gateway system (Invitrogen-Life
Technologies, Carlsbad, CA, USA). The retroviral vector of GnT-IVa
(pCLMSCV-GnT-IVa/puro) was transfected into 293T cells with
packaging vectors encoding viral gag-pol and envelope protein
(pCL-GALV) as previously described (23), and the retroviral supernatant was
transfected into Jar cells (Jar-GnT4a cells). Control vectors were
transfected into Jar cells (mock cells) by the same methods.
Jar-GnT4a and mock cells were selected by adding 1 µg/ml
puromycin.
Western blot and lectin blot
analysis
Whole cell lysate was prepared by lysing collected
mock or Jar-GnT4a cells in Tris-NaCl-EDTA (TNE) buffer supplemented
with protease inhibitor cocktail (Roche Diagnostics, Indianapolis,
IN, USA). Secreted proteins from mock or Jar-GnT4a cells were
obtained by concentrating the serum-free conditioned media in which
they were cultured with Amicon® Ultra-4 Centrifugal
Filter Units (Merck Millipore Ltd., Cork, Ireland). The protein
concentrations were determined using BCA protein assay kit (Pierce,
Rockford, IL, USA). Equal amounts of prepared proteins or
immunoprecipitated proteins were separated by 10% sodium dodecyl
sulphate-polyacrylamide gel electrophoresis (SDS-PAGE). Proteins on
the gels were transferred onto polyvinylidene difluoride membranes.
The membranes were blocked with 5% skim milk in PBS, or 3% BSA in
TBS containing 0.05% Tween-20 (TBS-T) at room temperature for 1 h,
for western blot or lectin blot analysis, respectively. The
membranes were incubated with a monoclonal anti-GnT-IVa antibody
(sc-100785, diluted 1:1,000; Santa Cruz Biotechnology), a
monoclonal anti-β-actin antibody (A2228, diluted 1:1,000;
Sigma-Aldrich), a polyclonal anti-integrin β1 antibody (ab183666,
diluted 1:5,000; Abcam plc., Cambridge, UK) and a monoclonal
anti-lysosome-associated membrane protein-2 (LAMP-2) antibody
(sc-18822, diluted 1:400; Santa Cruz Biotechnology) for western
blot analyses, or biotinylated GSL-II (B-1215, diluted 1:1,000;
Vector Laboratories) and Datura stramonium lectin (DSA)
(B-1185, diluted 1:1,000; Vector Laboratories) for lectin blot
analyses. The membranes were then incubated with horseradish
peroxidase-conjugated anti-mouse IgG secondary antibody (#7076S;
Cell Signaling Technology, Inc., Danvers, MA, USA) or
streptavidin-horseradish peroxidase (Vector Laboratories). Protein
bands were visualized by developing the blots using ECL reagents
(GE Healthcare, Buckinghamshire, UK) and ImageQuant LAS 4000 mini
(GE Healthcare).
Cell proliferation assay
Cell proliferation was determined using modified
tetrazolium salt (MTS) assay using the CellTiter 96®
Aqueous One Solution Cell Proliferation assay (Promega, Madison,
WI, USA). Cells (5×103) in 100 µl of growth medium were
plated in 96-well plates and incubated at 37°C for 72 h. At 24, 48
and 72 h, 20 µl of MTS reagent was added to each well, followed by
incubation for 2 h and measurement of absorbance at 490 nm by
microplate reader (Multiskan Bichromatic; Labsystems, Helsinki,
Finland).
Transwell migration and invasion
assay
Migration assay and invasion assay were performed
using the Transwell inserts (Corning Inc., Corning, NY, USA) with a
filter of 6.5 mm diameter and 8 µm pore size for 24-well plates as
described earlier (20,21). Invasion assay was performed using
the upper chambers coated with Matrigel (Collaborative Biomedical
Products, Bedford, MA, USA). Cells (4×104) in serum-free
medium were plated in the upper chambers, while medium with 10% FBS
was added to the lower chambers as chemo-attractant. Cells on the
upper side of the filters were scraped off after 24 h of incubation
at 37°C, and the filters were fixed and stained. The migration and
invasion activities were evaluated by counting the number of
migrated cells on the lower surface of the filters at ×100
magnification.
Cell adhesion assay
Cell adhesion assay was performed as previously
reported (20,25). Cells (4×104) were plated
in 100 ml of medium in 96-well plates coated with fibronectin,
collagen type I or collagen type IV (Becton-Dickinson, Franklin
Lakes, NJ, USA). The plates were centrifuged at 200 × g for 15 sec,
and the cells were allowed to attach to each matrix at 37°C for 30
min. After washing with PBS twice for eliminating non-adherent
cells, absorbance at 492 nm (A492nm) was read using a
microplate reader. The rate of cell adherence was calculated as
follows: [A492nm (matrix) - A492nm (no
matrix)]/A492nm (no matrix) (25).
Mouse studies
All animal procedures were reviewed and approved by
the Nagoya University institutional Animal Experimentation
Committee and performed in accordance with the institutional
guidelines of Nagoya University Division of Experimental Animals.
Five-week-old female nude mice (Balb/c slc nu/nu) were purchased
from Japan SLC, Inc. (Shizuoka, Japan) and were injected
subcutaneously with 5×105 mock or Jar-GnT4a cells in 200
µl of growth medium (n=8, each). Tumour size and health status of
mice were monitored, and when the minor axis of tumours exceeded 5
mm, tumour formation was considered to have occurred.
Immunoprecipitation
Whole cell lysate proteins or secreted proteins
(200–1,000 µg) from mock or Jar-GnT4a cells were incubated with
antibodies (10 µg) or biotinylated lectins (80 µg) in TNE buffer
containing protease protein inhibitor at 4°C overnight with rotary
shaking. The immuno-complex was then incubated with resins at 4°C
for 1 h with rotary shaking. Resins used were Protein G Sepharose 4
Fast Flow (GE Healthcare) for immunoprecipitation or Streptavidin
Agarose Ultra Performance™ (Solulink Bioscience, Inc., San Diego,
CA, USA) for lectin precipitation. The resins were used after
equilibration with TBS-T. Resin bound immnune-complexes were washed
five times with TBS-T and denatured by boiling at 100°C in SDS
sample buffer.
Nano-LC/MS/MS
The immunoprecipitates were separated by SDS-PAGE
and stained using Silver Stain MS kit (Wako Pure Chemical
Industries, Ltd., Osaka, Japan). The protein bands were excised
from the gels. Subsequent procedures and nano-LC/MS/MS analysis
were conducted by Japan Proteomics (Sendai, Japan) using a standard
protocol.
Flow cytometric analysis
For flow cytometric analysis, 1×105 cells
were suspended in PBS containing 1% BSA and were incubated for 30
min with 0.2 µg of a monoclonal anti-LAMP-2 antibody (sc-18822;
Santa Cruz Biotechnology). The cells were incubated with 2 µg of an
anti-mouse IgG (H+L) antibody Alexa Fluor® 488 conjugate
(A-11029; Invitrogen/Life Technologies) at 4°C for 30 min. After
washing with 3 ml of PBS, flow cytometric analysis was performed
using Attune® Acoustic Focusing Cytometer and
Attune® Cytometric software (Applied Biosystems,
Carlsbad, CA, USA). Mouse IgG1 isotype (MAB002; R&D
Systems, Inc., Minneapolis, MN, USA) was used for a negative
control.
Statistical analysis
JMP PRO version 11.0.0 (SAS Institute, Inc., Cary,
USA) was used for statistical analysis. Error bars in graphical
data represent mean ± standard deviation (SD). Differences were
considered statistically significant at P<0.05 in a two-tailed
Students t-test and log-rank test.
Results
Highly branched N-glycans present in
invasive mole and choriocarcinoma
We first examined the expression of GnT-IVa and
glycosylation level in invasive mole and choriocarcinoma by
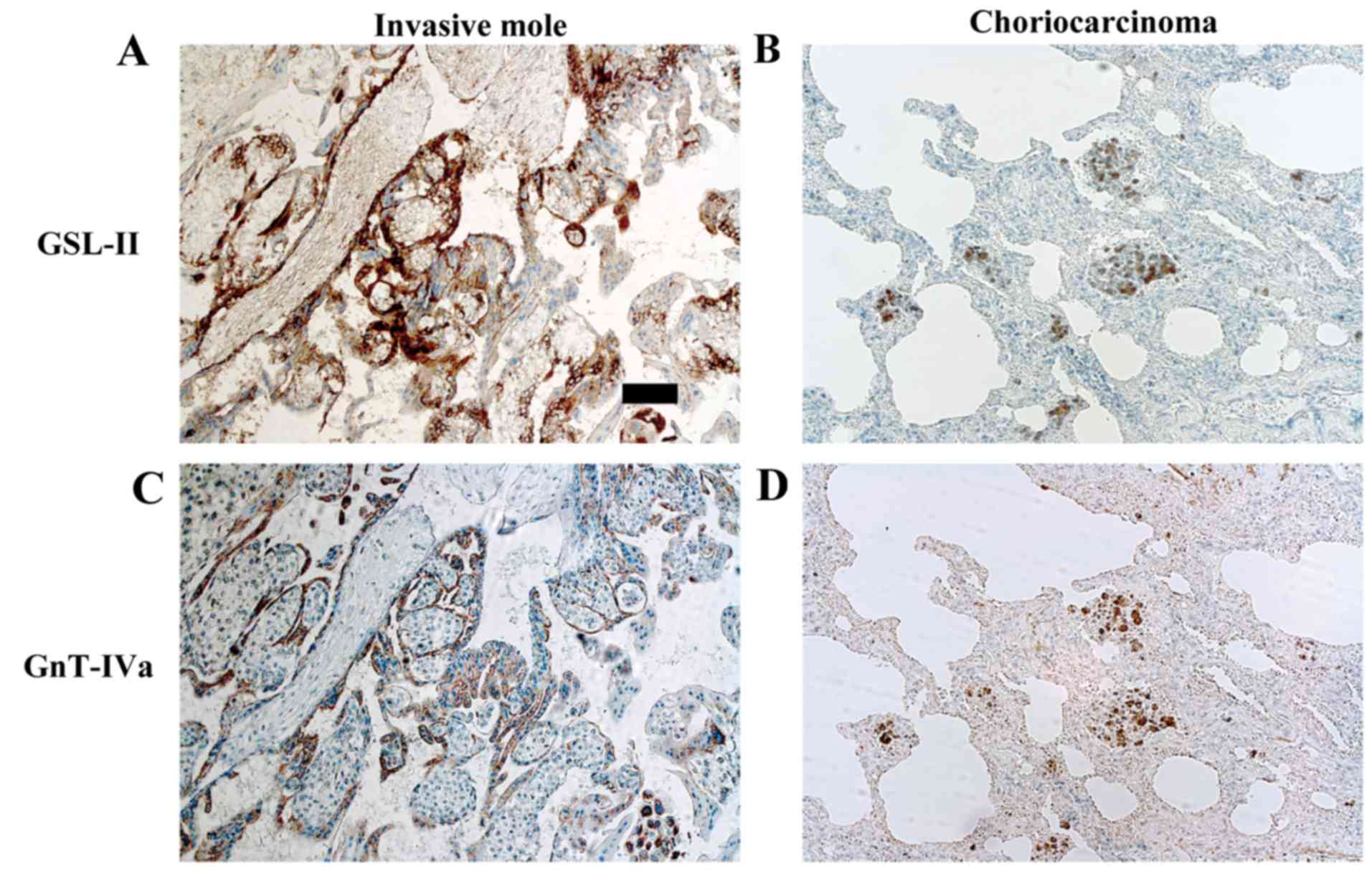
immunohistochemical staining of surgical specimens. GSL-II is a
lectin which recognizes tri- or tetra-antennary branched
N-glycans modified by GnT-IVa (26). Immunostaining with GSL-II was strong
in the invasive mole and choriocarcinoma and corresponded to that
with anti-GnT-IVa antibody (Fig.
1). These results indicated that highly branched
N-glycans modified by GnT-IVa are present in trophoblastic
cells of invasive mole and choriocarcinoma.
GnT-IVa overexpression increases cell
adhesion, migration and invasion in choriocarcinoma cells
To investigate the function of GnT-IVa in
choriocarcinoma cells, we established a stable cell line that
overexpressed GnT-IVa protein (Jar-GnT4a) using the choriocarcinoma
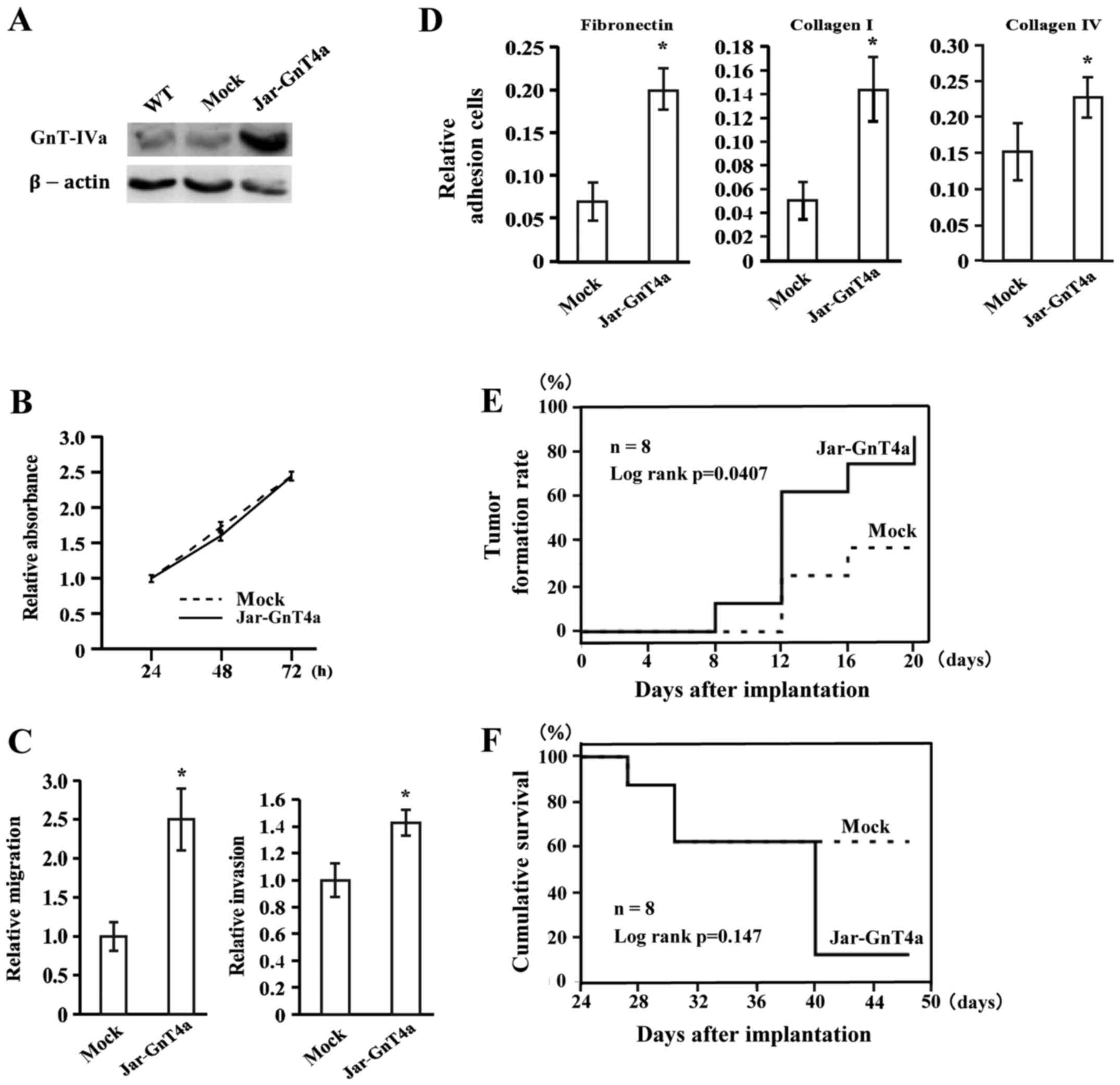
cell line Jar. The expression of GnT-IVa protein was dramatically
increased in Jar-GnT4a cells in comparison to that in parent Jar
(WT) or transfectants with control gene (mock cells, Fig. 2A). We observed no significant
difference between Jar-GnT4a cells and mock cells in cell
proliferation for 72 h, by MTS assay (Fig. 2B). The migration and invasion assay
showed that the migratory and invasive abilities of Jar-GnT4a cells
increased 2.5- and 1.4-fold, respectively, compared to those of
mock cells (Fig. 2C). Next, we
investigated the effect of GnT-IVa overexpression on cell adhesion
to extracellular matrix (ECM). Cell adhesion assay showed that
Jar-GnT4a cells had significantly greater adherence potential to
ECM than that of mock cells (Fig.
2D). The number of Jar-GnT4a cells that adhered to fibronectin,
collagen type I and collagen type IV were 2.88, 2.86 and 1.48 times
higher, respectively, than that of mock cells.
In vivo effects of GnT-IVa
overexpression on choriocarcinoma engraftment
We examined whether GnT-IVa overexpression increases
malignant potential of choriocarcinoma cells in vivo. We
subcutaneously injected Jar-GnT4a or mock cells into 5-week-old
Balb/c slc nu/nu nude mice. The rate of tumour formation in mice
injected with Jar-GnT4a cells was significantly higher than that in
mice injected with mock cells (P=0.0407; Fig. 2E). All of five mice and one of two
mice that developed tumours injected with Jar-GnT4a cells and mock
cells, respectively, died. The cumulative survival rate of mice
with Jar-GnT4a cells was relatively lower (62.5 vs. 12.5%),
although the difference was not statistically significant (Fig. 2F). Considered together, these
results suggest that GnT-IVa overexpression promotes cell adhesion,
migration and invasion of choriocarcinoma.
GnT-IVa overexpression increases
highly branched N-glycans on integrin β1
To investigate how GnT-IVa overexpression increased
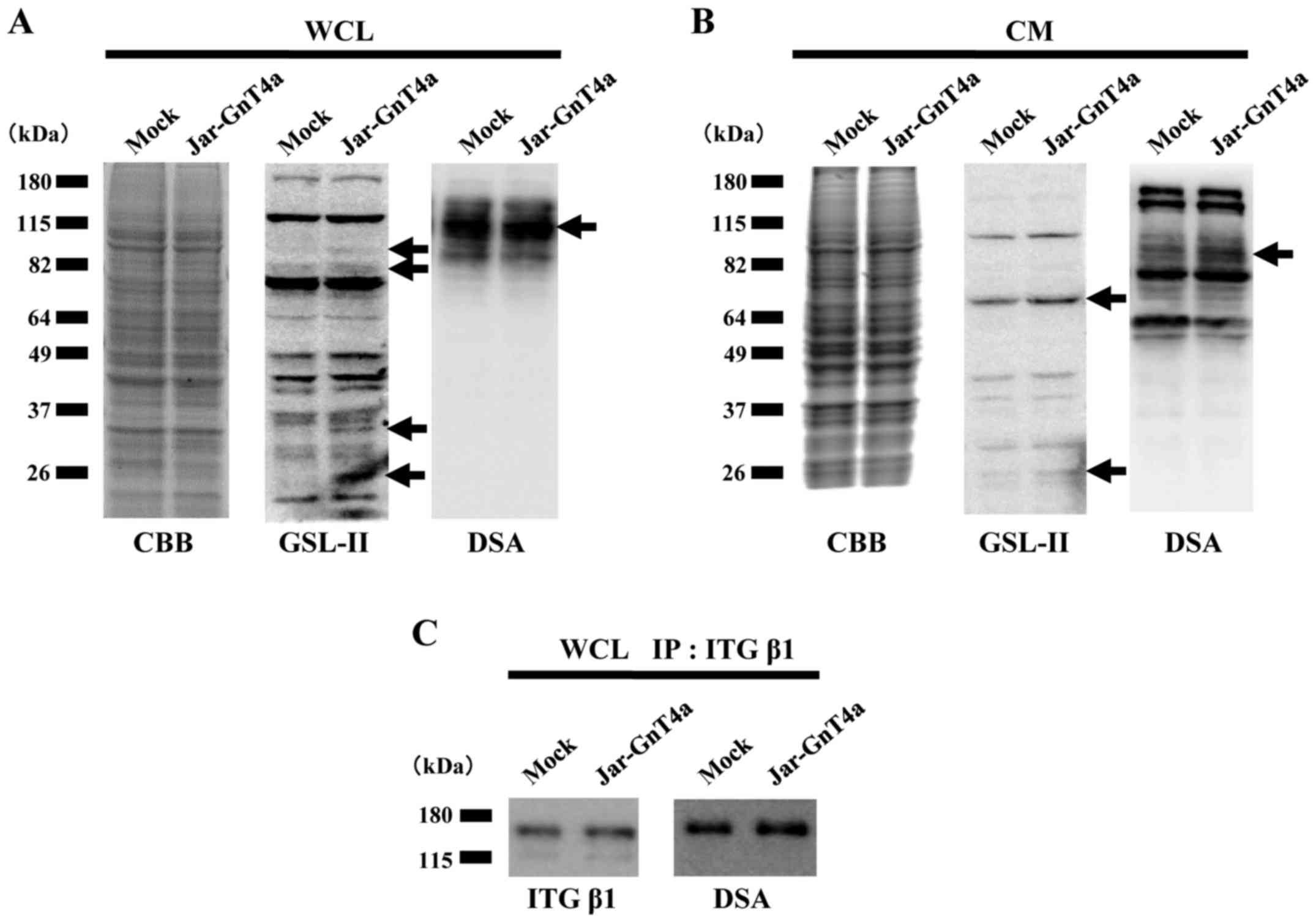
malignant potential of choriocarcinoma, we compared the protein
expression patterns and the level of β1,4GlcNAc branches of
N-glycans between Jar-GnT4a cells and mock cells. There was
no significant difference in the protein levels of whole cell
lysate and conditioned media between Jar-GnT4a and mock cells (left
panels, Fig. 3A and B). However,
lectin blot analyses with GSL-II and DSA which specifically
recognize β1,4GlcNAc branches of N-glycans, showed that the
overexpression of GnT-IVa mediated the addition of N-glycans
to several proteins in whole cell lysate or conditioned media
(arrows in centre and right panels, Fig. 3A and B). Because GSL-II has high
specificity for agalactosylated β1,4GlcNAc branches of
N-glycans whereas DSA has high specificity for
galactosylated β1, 4GlcNAc branches of N-glycans, we used
both lectins in these experiments.
Significantly greater adherence to ECM and higher
rate of tumour formation in mice injected with Jar-GnT4a cells
compared to those injected with mock cells suggested the
possibility that an increase in highly branched N-glycans on
integrin β1 might result from GnT-IVa overexpression. As expected,
β1,4GlcNAc branches of N-glycans on integrin β1 in whole
cell lysate were increased, as illustrated by DSA blot analysis
(Fig. 3C). These results suggest
that GnT-IVa may contribute to the malignant behaviour of
choriocarcinoma through decorating integrin β1 with
N-glycans.
LAMP-2 was one of the target proteins
for N-glycan modification by GnT-IVa
To gain insight into the functional role of GnT-IVa
in choriocarcinoma, we tried to find target proteins other than
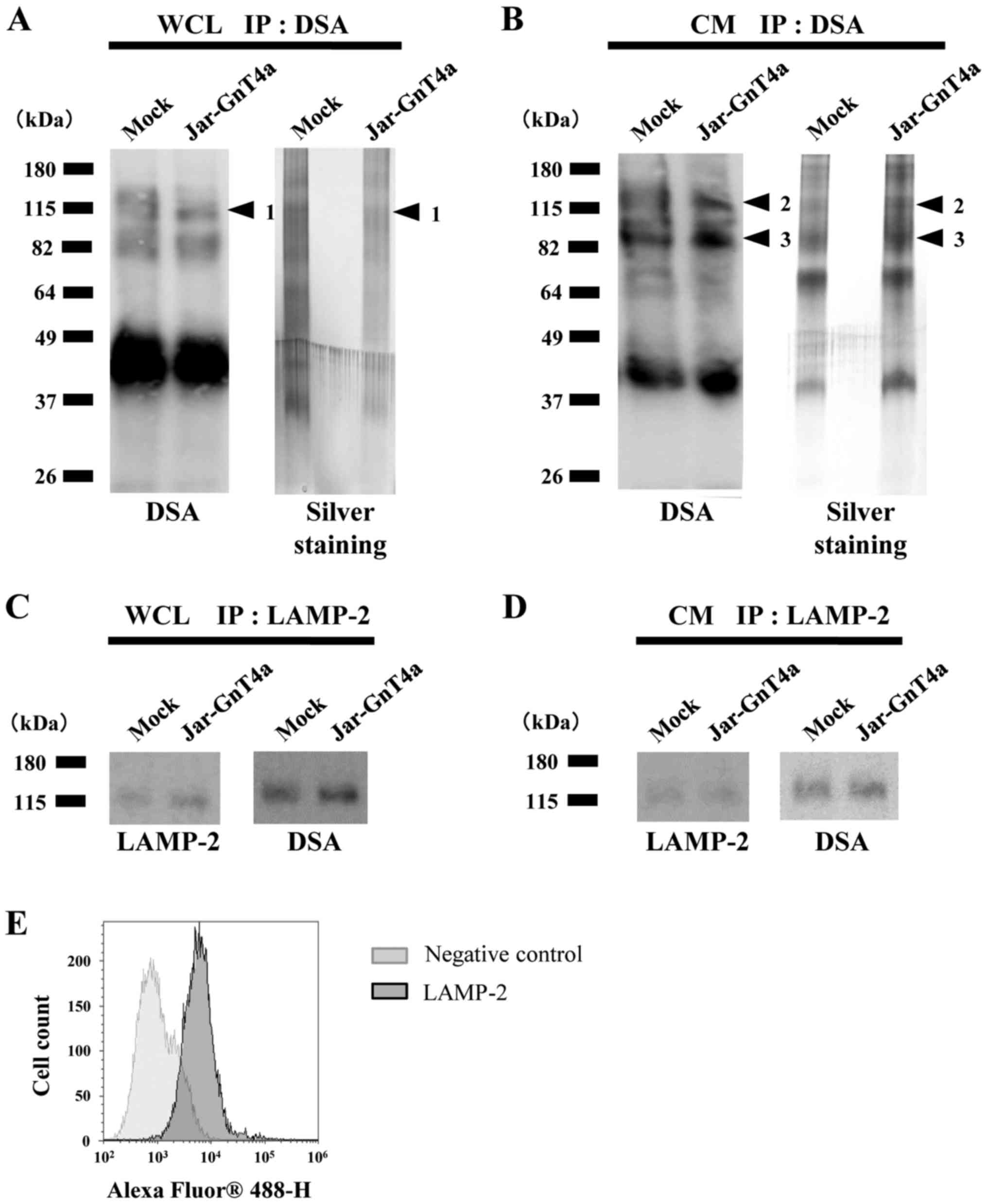
integrin β1 for GnT-IVa glycosylation. We concentrated the target
proteins in whole cell lysate and secreted proteins by lectin
immunoprecipitation with DSA, because DSA blot analysis showed
greater changes between Jar-GnT4a and mock cells than those by
GSL-II blot analysis (Fig. 3A and
B). DSA blot analyses and silver staining using
immunoprecipitated proteins revealed that some proteins reacted
more strongly in Jar-GnT4a cells than in mock cells (Fig. 4A and B). One and two protein bands
containing separated whole cell lysate and conditioned media,
respectively, were excised from the gels (arrowheads 1–3, Fig. 4A and B). Nano-LC/MS/MS analysis
identified the proteins contained in the three bands (Table I). We focused on lysosome-associated
membrane glycoprotein 2 (LAMP-2), because it was present in both
whole cell lysate and conditioned media.
 | Table I.Target proteins of GnT-IVa
glycosylation in choriocarcinoma shown by Nano-LC/MS/MS
analysis. |
Table I.
Target proteins of GnT-IVa
glycosylation in choriocarcinoma shown by Nano-LC/MS/MS
analysis.
| Band no. | Protein | Peptide
matching | Protein coverage
(%) |
|---|
| 1 | Lysosome-associated
membrane glycoprotein 2 | 4 | 11 |
| 2 | α actin | 8 | 11 |
| 2 | Lysosome-associated
membrane glycoprotein 2 | 3 | 9 |
| 3 | Phospholipid
transfer protein | 1 | 3 |
| 3 | Transferrin
receptor protein 1 | 4 | 5 |
We examined whether highly branched N-glycans
on LAMP-2 were increased by GnT-IVa overexpression. β1,4GlcNAc
branches of N-glycans on LAMP-2 in whole cell lysate and
conditioned medium were increased as illustrated by DSA blot
analyses (Fig. 4C and D),
indicating that LAMP-2 is one of the target proteins for GnT-IVa
glycosylation and that it might contribute to the malignant
potential of choriocarcinoma cells through GnT-IVa glycosylation.
The differences in the reactivity of immunoprecipitated LAMP-2
between Jar-GnT4a and mock cells were relatively modest in DSA blot
analyses in both whole cell lysate and conditioned media. This
might be due to considerable expression of GnT-IVa in mock cells
and LAMP-2 which carried β1,4GlcNAc branches of N-glycans
may be substantially present in mock cells. The amount of LAMP-2 in
whole cell lysate and conditioned media was confirmed by
immunoprecipitation using an anti-LAMP-2 antibody. Next, we
performed flow cytometric analysis to examine whether LAMP-2 was
expressed on the cell surface. Whole cell lysate contains
cytoplasmic proteins as well as cell surface proteins. Our results
showed that GnT-IVa overexpression changed the abilities of cell
adhesion to ECM. Flow cytometric analysis revealed that LAMP-2 was
present on the cell surface of Jar (Fig. 4E). Taken together, these results
suggest that GnT-IVa may contribute to the malignant behaviour of
choriocarcinoma through modifying N-glycans on integrin β1
and LAMP-2 which is expressed on the cell surface.
Discussion
In the present study, we showed that GnT-IVa
overexpression enhanced the malignant potential of choriocarcinoma
in vitro and in vivo, by increasing cellular adhesion
to ECM, cellular migration and invasion. These results complement
the results of a previous study which showed that GnT-IVa knockdown
in choriocarcinoma cells suppressed migration and invasion and
decreased cellular adhesion to ECM (20). Immunohistochemistry using GSL-II
demonstrated that highly branched N-glycans resulting from
the action of GnT-IVa were strongly detected in invasive mole and
choriocarcinoma, in proportion to the GnT-IVa protein expression.
Therefore, GnT-IVa may play an important role in accelerating the
malignancy of choriocarcinoma through addition of β1,4GlcNAc
branches to the N-glycans on some proteins. We tried to find
the target proteins for GnT-IVa glycosylation.
Immunoprecipitation showed that integrin β1 was a
target protein of GnT-IVa glycosylation in choriocarcinoma.
Integrins are transmembrane cell adhesion molecules which function
as cellular receptors of ECM ligands such as collagens, fibronectin
and laminins (27). It has been
reported that the N-glycan structure on integrin α or β
chains, resulting from the action of GnTs, affects
integrin-mediated cell migration. Zhao et al (28) demonstrated that α3β1
integrin-mediated cell migration on laminin 5 was competitively
regulated by GnT-III which catalyses the addition of bisected
GlcNAc on integrin and suppresses cell migration. GnT-V which forms
β1,6 GlcNAc branched N-glycans enhances cell migration. In
the present study, the results of cell adhesion assay and tumour
formation in mice suggested that GnT-IVa may increase cell adhesion
to ECM in choriocarcinoma, in vitro and in vivo.
Niimi et al (20) reported
that knocking down GnT-IVa in choriocarcinoma cells decreased
β1,4GlcNAc branched N-glycans on integrin β1 and suppressed
integrin-mediated cell migration. These results support that
GnT-IVa plays a role in the invasion of choriocarcinoma by
increasing the β1,4GlcNAc branched N-glycans on integrin
β1.
Lectin immunoprecipitation and nano-LC/MS/MS
revealed that LAMP-2 is another target protein of GnT-IVa
glycosylation in choriocarcinoma. Some glycoproteins, such as
integrin β1, CD147, carcinoembryonic antigens, gamma-glutamyl
transpeptidases and glucose transporter-2 have been reported as
target proteins for glycosylation by GnT-IVa in various carcinomas
(11,29–31).
LAMP-2 (CD107b) is a single-pass type 1 transmembrane protein which
is normally present in the lysosomal membranes and occasionally on
the surface of cell membranes in special situations including
cancers (32–35). LAMP-2 is heavily glycosylated, to
the extent that its carbohydrate side chains account for half of
the molecular weight (33,34). LAMP-2 has 16 N-glycans which
play crucial roles in cell adhesion to ECM and metastasis of cancer
cells because N-glycans configure cell surface ligands of
cancer cells for galectin-1 (36),
galectin-3 (35,37) and selectins (38) on ECM or the membranes of the counter
cell. Saitoh et al (38)
reported that a colon carcinoma cell line with high metastatic
potential expressed LAMP-2 at higher levels than that with low
metastatic potential. In the present study, we demonstrated that
LAMP-2 was expressed on the surface of choriocarcinoma cells and
that the reactivity of DSA to N-glycans of LAMP-2 was
increased by GnT-IVa overexpression. These results suggest that
GnT-IVa may contribute to choriocarcinoma invasion by increasing
β1,4GlcNAc branching on N-glycans of LAMP-2 as well as
integrin β1.
This study did not reveal the reason why LAMP-2 was
detected in secreted proteins of Jar cells in this study, although
it is very interesting that a transmembrane protein was present in
conditioned media. We performed nano-LC/MS/MS on secreted proteins
because the function of secreted proteins can be changed by
glycosylation and affect cell invasion. It might be that a part of
LAMP-2 on the cell surface was cleaved or included in exosomes and
released into conditioned media which resulted in LAMP-2 detection
in secreted proteins of Jar cells in this study. Previous studies
reported that exosomal biomolecules (proteins, lipids, RNA and DNA)
of cancer cells are associated with metastases to specific organs
by preparing a pre-metastatic niche (39) and that N-glycosylation
stabilizes LAMP-2 of exosomes and protects it from proteolysis for
delivery of exosomes to target cells (40). LAMP-2 was detected in exosomes from
HEK293FT cells which were transfected with tagged Lamp-2
introduction (40). It is suggested
that cell membrane is included in the process of generating
exosomes (41). It is possible that
choriocarcinoma-specific tumor exosomes have LAMP-2 and promote
organ-specific metastasis. However, more investigation is needed to
explore the functional roles and localization of LAMP-2 in
choriocarcinoma.
In conclusion, highly branched N-glycans
resulting from the action of GnT-IVa were present in invasive mole
and choriocarcinoma in proportion to GnT-IVa expression. GnT-IVa
may facilitate invasion of choriocarcinoma by promoting cell
adhesion, migration and invasion through β1,4GlcNAc glycosylation
of integrin β1 and LAMP-2.
Acknowledgements
We appreciate Dr Suguru Oguri (Tokyo University of
Agriculture) and Dr Aruto Yoshida (Kirin Brewery Co.) for providing
GnT-IVa/pSVL vector. We would like to thank Editage (www.editage.jp) for English language editing. The
present study was supported by grant-in-aid no. 23592445 (to E.Y.)
from the Japanese Ministry of Education, Culture, Sports, Science
and Technology.
References
|
1
|
Berkowitz RS and Goldstein DP: Chorionic
tumors. N Engl J Med. 335:1740–1748. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lurain JR: Gestational trophoblastic
disease I: Epidemiology, pathology, clinical presentation and
diagnosis of gestational trophoblastic disease, and management of
hydatidiform mole. Am J Obstet Gynecol. 203:531–539. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lurain JR: Gestational trophoblastic
disease II: Classification and management of gestational
trophoblastic neoplasia. Am J Obstet Gynecol. 204:11–18. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Berkowitz RS and Goldstein DP: Current
advances in the management of gestational trophoblastic disease.
Gynecol Oncol. 128:3–5. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Varki A: Biological roles of
oligosaccharides: All of the theories are correct. Glycobiology.
3:97–130. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dennis JW, Laferté S, Waghorne C, Breitman
ML and Kerbel RS: Beta 1–6 branching of Asn-linked oligosaccharides
is directly associated with metastasis. Science. 236:582–585. 1987.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pinho SS, Seruca R, Gärtner F, Yamaguchi
Y, Gu J, Taniguchi N and Reis CA: Modulation of E-cadherin function
and dysfunction by N-glycosylation. Cell Mol Life Sci.
68:1011–1020. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Oguri S, Minowa MT, Ihara Y, Taniguchi N,
Ikenaga H and Takeuchi M: Purification and characterization of
UDP-N-acetylglucosamine: α1,3-D-mannoside
β1,4-N-acetylglucosaminyltransferase
(N-acetylglucosaminyltransferase-IV) from bovine small intestine. J
Biol Chem. 272:22721–22727. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Minowa MT, Oguri S, Yoshida A, Hara T,
Iwamatsu A, Ikenaga H and Takeuchi M: cDNA cloning and expression
of bovine UDP-N-acetylglucosamine: alpha1, 3-D-mannoside
beta1,4-N-acetylglucosaminyltransferase IV. J Biol Chem.
273:11556–11562. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ide Y, Miyoshi E, Nakagawa T, Gu J,
Tanemura M, Nishida T, Ito T, Yamamoto H, Kozutsumi Y and Taniguchi
N: Aberrant expression of N-acetylglucosaminyltransferase-IVa and
IVb (GnT-IVa and b) in pancreatic cancer. Biochem Biophys Res
Commun. 341:478–482. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fan J, Wang S, Yu S, He J, Zheng W and
Zhang J: N-acetylglucosaminyltransferase IVa regulates metastatic
potential of mouse hepatocarcinoma cells through glycosylation of
CD147. Glycoconj J. 29:323–334. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhu TY, Chen HL, Gu JX, Zhang YF, Zhang YK
and Zhang RA: Changes in Nacetylglucosaminyltransferase III IV and
V in renal cell carcinoma. J Cancer Res Clin Oncol. 123:296–299.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
DArrigo A, Belluco C, Ambrosi A, Digito M,
Esposito G, Bertola A, Fabris M, Nofrate V, Mammano E, Leon A, et
al: Metastatic transcriptional pattern revealed by gene expression
profiling in primary colorectal carcinoma. Int J Cancer.
115:256–262. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kobata A and Takeuchi M: Structure,
pathology and function of the N-linked sugar chains of human
chorionic gonadotropin. Biochim Biophys Acta. 1455:315–326. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Endo Y, Yamashita K, Tachibana Y, Tojo S
and Kobata A: Structures of the asparagine-linked sugar chains of
human chorionic gonadotropin. J Biochem. 85:669–679.
1979.PubMed/NCBI
|
|
16
|
Kessler MJ, Reddy MS, Shah RH and Bahl OP:
Structures of N-glycosidic carbohydrate units of human chorionic
gonadotropin. J Biol Chem. 254:7901–7908. 1979.PubMed/NCBI
|
|
17
|
Mizuochi T, Nishimura R, Derappe C,
Taniguchi T, Hamamoto T, Mochizuki M and Kobata A: Structures of
the asparagine-linked sugar chains of human chorionic gonadotropin
produced in choriocarcinoma. Appearance of triantennary sugar
chains and unique biantennary sugar chains. J Biol Chem.
258:14126–14129. 1983.PubMed/NCBI
|
|
18
|
Mizuochi T, Nishimura R, Taniguchi T,
Utsunomiya T, Mochizuki M, Derappe C and Kobata A: Comparison of
carbohydrate structure between human chorionic gonadotropin present
in urine of patients with trophoblastic diseases and healthy
individuals. Jpn J Cancer Res. 76:752–759. 1985.PubMed/NCBI
|
|
19
|
Takamatsu S, Oguri S, Minowa MT, Yoshida
A, Nakamura K, Takeuchi M and Kobata A: Unusually high expression
of N-acetylglucosaminyltransferase-IVa in human choriocarcinoma
cell lines: A possible enzymatic basis of the formation of abnormal
biantennary sugar chain. Cancer Res. 59:3949–3953. 1999.PubMed/NCBI
|
|
20
|
Niimi K, Yamamoto E, Fujiwara S, Shinjo K,
Kotani T, Umezu T, Kajiyama H, Shibata K, Ino K and Kikkawa F: High
expression of N-acetylglucosaminyltransferase IVa promotes invasion
of choriocarcinoma. Br J Cancer. 107:1969–1977. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yamamoto E, Ito T, Abe A, Sido F, Ino K,
Itakura A, Mizutani S, Dovat S, Nomura S and Kikkawa F: Ikaros is
expressed in human extravillous trophoblasts and involved in their
migration and invasion. Mol Hum Reprod. 11:825–831. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Suzuki O, Nozawa Y, Kawaguchi T and Abe M:
Phaseolus vulgaris leukoagglutinating lectin-binding reactivity in
human diffuse large B-cell lymphoma and its relevance to the
patients clinical outcome: Lectin histochemistry and lectin blot
analysis. Pathol Int. 49:874–880. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yamashita Y, Tsurumi T, Mori N and Kiyono
T: Immortalization of Epstein-Barr virus-negative human B
lymphocytes with minimal chromosomal instability. Pathol Int.
56:659–667. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Oguri S, Yoshida A, Minowa MT and Takeuchi
M: Kinetic properties and substrate specificities of two
recombinant human N-acetylglucosaminyltransferase-IV isozymes.
Glycoconj J. 23:473–480. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yamamoto E, Ino K, Miyoshi E, Inamori K,
Abe A, Sumigama S, Iwase A, Kajiyama H, Shibata K, Nawa A, et al:
N-acetylglucosaminyltransferase V regulates extravillous
trophoblast invasion through glycosylation of alpha5beta1 integrin.
Endocrinology. 150:990–999. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Nakamura-Tsuruta S, Kominami J, Kamei M,
Koyama Y, Suzuki T, Isemura M and Hirabayashi J: Comparative
analysis by frontal affinity chromatography of oligosaccharide
specificity of GlcNAc-binding lectins, Griffonia simplicifolia
lectin-II (GSL-II) and Boletopsis leucomelas lectin (BLL). J
Biochem. 140:285–291. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Desgrosellier JS and Cheresh DA: Integrins
in cancer: Biological implications and therapeutic opportunities.
Nat Rev Cancer. 10:9–22. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhao Y, Nakagawa T, Itoh S, Inamori K,
Isaji T, Kariya Y, Kondo A, Miyoshi E, Miyazaki K, Kawasaki N, et
al: N-acetylglucosaminyltransferase III antagonizes the effect of
N-acetylglucosaminyltransferase V on alpha3beta1 integrin-mediated
cell migration. J Biol Chem. 281:32122–32130. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yamashita K, Totani K, Kuroki M, Matsuoka
Y, Ueda I and Kobata A: Structural studies of the carbohydrate
moieties of carcinoembryonic antigens. Cancer Res. 47:3451–3459.
1987.PubMed/NCBI
|
|
30
|
Yamashita K, Totani K, Iwaki Y, Takamisawa
I, Tateishi N, Higashi T, Sakamoto Y and Kobata A: Comparative
study of the sugar chains of gamma-glutamyltranspeptidases purified
from human hepatocellular carcinoma and from human liver. J
Biochem. 105:728–735. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ohtsubo K: Targeted genetic inactivation
of N-acetylglucosaminyltransferase-IVa impairs insulin secretion
from pancreatic beta cells and evokes type 2 diabetes. Methods
Enzymol. 479:205–222. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen JW, Murphy TL, Willingham MC, Pastan
I and August JT: Identification of two lysosomal membrane
glycoproteins. J Cell Biol. 101:85–95. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Carlsson SR, Roth J, Piller F and Fukuda
M: Isolation and characterization of human lysosomal membrane
glycoproteins, h-lamp-1 and h-lamp-2. Major sialoglycoproteins
carrying polylactosaminoglycan. J Biol Chem. 263:18911–18919.
1988.PubMed/NCBI
|
|
34
|
Fukuda M, Viitala J, Matteson J and
Carlsson SR: Cloning of cDNAs encoding human lysosomal membrane
glycoproteins, h-lamp-1 and h-lamp-2. Comparison of their deduced
amino acid sequences. J Biol Chem. 263:18920–18928. 1988.PubMed/NCBI
|
|
35
|
Sarafian V, Jadot M, Foidart JM, Letesson
JJ, Van den Brûle F, Castronovo V, Wattiaux R and Coninck SW:
Expression of Lamp-1 and Lamp-2 and their interactions with
galectin-3 in human tumor cells. Int J Cancer. 75:105–111. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Skrincosky DM, Allen HJ and Bernacki RJ:
Galaptin-mediated adhesion of human ovarian carcinoma A121 cells
and detection of cellular galaptin-binding glycoproteins. Cancer
Res. 53:2667–2675. 1993.PubMed/NCBI
|
|
37
|
Inohara H and Raz A: Identification of
human melanoma cellular and secreted ligands for galectin-3.
Biochem Biophys Res Commun. 201:1366–1375. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Saitoh O, Wang WC, Lotan R and Fukuda M:
Differential glycosylation and cell surface expression of lysosomal
membrane glycoproteins in sublines of a human colon cancer
exhibiting distinct metastatic potentials. J Biol Chem.
267:5700–5711. 1992.PubMed/NCBI
|
|
39
|
Hoshino A, Costa-Silva B, Shen TL,
Rodrigues G, Hashimoto A, Mark M Tesic, Molina H, Kohsaka S, Di
Giannatale A, Ceder S, et al: Tumour exosome integrins determine
organotropic metastasis. Nature. 527:329–335. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hung ME and Leonard JN: Stabilization of
exosome-targeting peptides via engineered glycosylation. J Biol
Chem. 290:8166–8172. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Vanni I, Alama A, Grossi F, Dal Bello MG
and Coco S: Exosomes: A new horizon in lung cancer. Drug Discov
Today. Mar 10–2017.Epub ahead of print). doi:
10.1016/j.drudis.2017.03.004. View Article : Google Scholar
|