Introduction
Epithelial ovarian cancer is classified into four
major subgroups: serous, clear cell, endometrioid and mucinous
(1). Ovarian clear cell carcinoma
(OCCC) has phenotypes distinct from those of the other subgroups,
such as resistance to chemotherapy, poor prognosis, an association
with endometriosis, a higher incidence of thrombosis as a
complication and a higher incidence among Japanese individuals
(2–4). OCCC contains mutations in various
tumor suppressors and oncogenes, including AT-rich interactive
domain-containing protein 1A (ARID1A),
phosphatidylinositol-3-kinase catalytic subunit (PIK3CA), KRAS and
TP53 (5). Furthermore, a gain in
DNA copy number is frequently found in chromosome 20, which
contains the potential oncogene zinc finger protein 217 (ZNF217),
while losses in DNA copy number are observed at the CDKN2A/2B and
LZTS1 loci (6,7).
We recently analyzed somatic copy number alterations
in OCCC using comparative genomic hybridization (CGH) (8). In agreement with previous reports
(7), we found recurrent
amplification of the 20q13 locus containing ZNF217 and the 17q23.2
locus harboring PPM1D. Furthermore, we found recurrent
amplification of the entire chromosome 8q, the EGFR and HER2/ERBB2
gene loci, and losses of chromosomes 9q, 13q and 17q. In the
present study, we reanalyzed this CGH data and further identified
amplification in the chromosomal regions 8p11.21–22 and 12p13.31 as
well as 20q13.2, and losses of small segments in the chromosomal
regions 3q26.1, 4q13.2 and 22q11.23 in OCCC. Moreover, we showed
that these DNA copy number alterations are associated with poor
patient prognosis.
Materials and methods
Specimens
Tumor specimens were surgically obtained from 110
OCCC patients (95 Japanese and 15 Korean patients). All the
patients provided written informed consent. This study was
performed in accordance with the Helsinki Declaration and was
approved by the Ethics Committee of the Jikei University School of
Medicine (8). The characteristics
of the patients are summarized in Table
I. Our study sample was considered to be representative of the
OCCC patient population. Japanese and Korean individuals are
genetically very similar (9). The
response to chemotherapy was defined as: i) platinum sensitive, if
no relapse or progression was noted within 6 months after the last
cycle of chemotherapy; ⅱ) platinum resistant, if the patients did
not respond to therapy or responded initially but relapsed or
progressed within 6 months after the last cycle of
chemotherapy.
 | Table I.Characteristics of the OCCC
patients. |
Table I.
Characteristics of the OCCC
patients.
| Characteristics | Data |
|---|
| Total patients,
n | 110 |
| Age (years), median
(range) | 53 (30–86) |
| Stage, n |
|
| I | 61 |
| II | 14 |
| III | 32 |
| IV | 3 |
| Endometriosis, n
(%) | 54 (49) |
| Thrombosis, n
(%) | 12 (11) |
| Sensitivity to
chemotherapy, n (%) |
|
|
Sensitive | 60 (55) |
|
Resistant | 26 (24) |
| Disease duration
(months), median (IQR) | 17.8 (10–36) |
| Progression, n
(%) | 33 (30) |
| Deceased, n (%) | 21 (19) |
| Amplification, n
(%) |
|
|
8p11.22 | 20 (18) |
|
8p11.21 | 14 (13) |
|
12p13.31 | 13 (12) |
|
20q13.2 | 14 (13) |
| Deletion |
|
|
3q26.1 | 36 (33) |
|
4q13.2 | 21 (19) |
|
22q11.23 | 4 (4) |
CGH analysis
CGH analysis was performed according to the
manufacturer's instructions (Agilent Technologies, Inc., Santa
Clara, CA, USA). DNA was extracted from tumor and normal tissues
using the AllPrep DNA/RNA Mini kit and the QIAamp DNA Mini kit,
respectively (Qiagen, Dusseldorf, Germany). Tumor and normal DNA
digested with Rsa1 and Alu1 was labeled with Cy5 and
Cy3, respectively, using the Agilent Genomic DNA Enzymatic Labeling
kit (Agilent Technologies, Inc.). The labeled DNA was hybridized to
Human Genome CGH Microarray kit 244A (G4411B; Agilent Technologies,
Inc.), which contains ~236,381 probes annotated against the
National Center for Biotechnology Information (NCBI) Build 36. The
array was scanned with an Agilent G2565BA Microarray Scanner, and
the fluorescent signals were analyzed using Feature Extraction
software, version 10.7.3.1 (Agilent Technologies, Inc.). DNA copy
number aberrations were identified using the ADM-2 algorithm in
CytoGenomics Software (Agilent Technologies, Inc.). Regions with a
copy number gain were defined as those with an average
log2 ratio ≥0.9 across at least three consecutive
probes. Regions with a copy number loss were defined as those with
an average log2 ratio ≤-1.5 across at least three
consecutive probes. All genomic positions were defined according to
the University of California Santa Cruz (UCSC) Human version hg19.
The P-value corresponding to each interval was calculated using the
normal probability distribution function and the score of that
interval. The abnormal chromosomal regions among the OCCC
population were defined by applying T-Test Common Aberration
algorithm (Genomic Workbench; Agilent Technologies, Inc.).
Droplet digital PCR (ddPCR)
DNA was extracted from tumor tissues and buffy coat
samples using the AllPrep DNA/RNA Mini kit and the QIAamp DNA Mini
kit, respectively (Qiagen). Probes were from the Universal Probe
Library (UPL) (Roche Diagnostics, Basel, Switzerland). A minor
groove-binding (MGB) probe labeled with 6-carboxyfluorescein (FAM)
was used for ddPCR to assess the copy number of ANK1, MYST3, IKBKB,
POLB and ZNF217. A MGB probe labeled with VIC (Thermo Fisher
Scientific, Inc., Waltham, MA, USA) was used to quantitate
endogenous control EFTUD2. Primers of ~20 bases were designed using
Primer3Plus and purchased from Invitrogen (Thermo Fisher
Scientific, Inc.). Primer sequences are listed in Table II. Reaction mixtures (20 µl)
containing 5 or 10 ng of digested sample DNA, ddPCR supermix for
probes (Bio-Rad Laboratories, Inc., Hercules, CA, USA), 1,000 nM of
each primer and 250 nM of each probe were loaded into the QX100
Droplet Generator (10). The
samples were amplified on the conventional Bio-Rad T100 Thermal
Cycler (95°C for 10 min, followed by 40 cycles of 95°C for 30 sec
and 60°C for 60 sec, with a final elongation step of 98°C for 10
min). The plate, containing the droplet amplicons, was subsequently
loaded into the QX100 Droplet Reader (Bio-Rad Laboratories, Inc.).
The number of gene copies was calculated using QuantaSoft software
(Bio-Rad Laboratories, Inc.). The average number of droplets
generated was 12,136. Data were analyzed using the Wilcoxon rank
sum test.
| Table II.Primer/probe sequences. |
Table II.
Primer/probe sequences.
| Gene | Sequences |
|---|
| ANK1 |
|
|
Forward |
5′-TCCCCTGAATTAAGCCTTCAG-3′ |
|
Reverse |
5′-CCACCCCAGGGACTCTTTAC-3′ |
|
Probe | ROCHE UPL #55 |
| MYST3 |
|
|
Forward |
5′-CATGTTGTTTTCCCCTTTCAA-3′ |
|
Reverse |
5′-ACGAAAAAGAAATTCCAACTGTG-3′ |
|
Probe | ROCHE UPL #69 |
| IKBKB |
|
|
Forward |
5′-ATCATTGTGGGCTGCAGATT-3′ |
|
Reverse |
5′-TGGGGAATCCTCTCTCCACT-3′ |
|
Probe | ROCHE UPL #19 |
| POLB |
|
|
Forward |
5′-TTGAACCATCATCAGCGAAT-3′ |
|
Reverse |
5′-CTCAAGTGTCAAAAGAAAATCTGC-3′ |
|
Probe | ROCHE UPL #41 |
| ZNF217 |
|
|
Forward |
5′-CACGATTGATTGGACTCTTCC-3′ |
|
Reverse |
5′-CACGTCAATCACATGATCAGAA-3′ |
|
Probe | ROCHE UPL #41 |
| ADAM5 |
|
|
Forward |
5′-CCAGTGCCTAGAAGAGTGTCTG-3′ |
|
Reverse |
5′-CAGACTCTGTGACTCCTTTGTATCA-3′ |
|
Probe | ROCHE UPL #5 |
| Chr12
ENST00000518709 |
|
|
Forward |
5′-CTGTAGCCATCTGTCCAAGTGT-3′ |
|
Reverse |
5′-AGGCAAGCAGAGGAAATCTG-3′ |
|
Probe | ROCHE UPL #19 |
Exome capture library and whole-exome
sequencing
DNA libraries were generated from genomic DNA
derived from tumor and/or normal tissues of the OCCC patients, and
subsequently sequenced. From this, exome-captured sequencing
libraries were produced using Agilent SureSelect XT Human All Exon
v5 (Agilent Technologies, Inc.). The captured DNA was sequenced
using the Illumina HiSeq 2500 platform (Illumina, Inc., San Diego,
CA, USA) with paired-end reads of 100 bp for insert libraries
according to the manufacturer's instructions. We deposited all DNA
sequence data used in this study to the National Bioscience
Database Center (NBDC) Human Database (http://humandbs.biosciencedbc.jp/).
Exome sequence data analysis
Low quality (q<15) and adaptor regions were
trimmed using in-house software beforehand. Read sequences were
mapped using Bowtie 2 (11) to the
human reference genome (B37). After filtering by pair mapping
distance, mapping uniqueness and pair orientation, the mapping
result files were converted into pileup format using SAMtools
(12). Copy number changes, or
amplifications and deletions were detected from coverage
comparisons between tumor and normal data sets. Focal gains and
losses were defined as occurring on gene segments.
Kaplan Meier analysis
Progression-free survival (PFS) rates and overall
survival (OS) rates were calculated using the Kaplan-Meier method
and evaluated using the log-rank test. Confidence intervals (CIs)
were assessed by univariate and multivariate Cox proportional
hazards models. Multivariate Cox models were used to determine
whether the increased hazard rates for PFS and OS attributed to
gene amplification in the univariate model were still present after
accounting for patient-, disease-, and prior treatment-related
factors. P<0.05 was considered to be statistically
significant.
Results
The characteristics of the 110 patients (95 Japanese
and 15 Korean patients) studied are summarized in Table I. With a median follow-up of 17.8
months (range, 10–36 months), 33 (30%) patients relapsed and 21
(19%) patients died. The mean age was 53 years (range, 30–86
years). FIGO stages were: stage I, 61 patients; stage II, 14
patients; stage III, 32 patients; stage IV, 3 patients.
Endometriosis was diagnosed in 54 (49%) patients. Thrombosis was
observed in 12 (11%) patients. The response to chemotherapy was:
platinum sensitive, 60 (55%) patients; platinum resistant, 26 (24%)
patients.
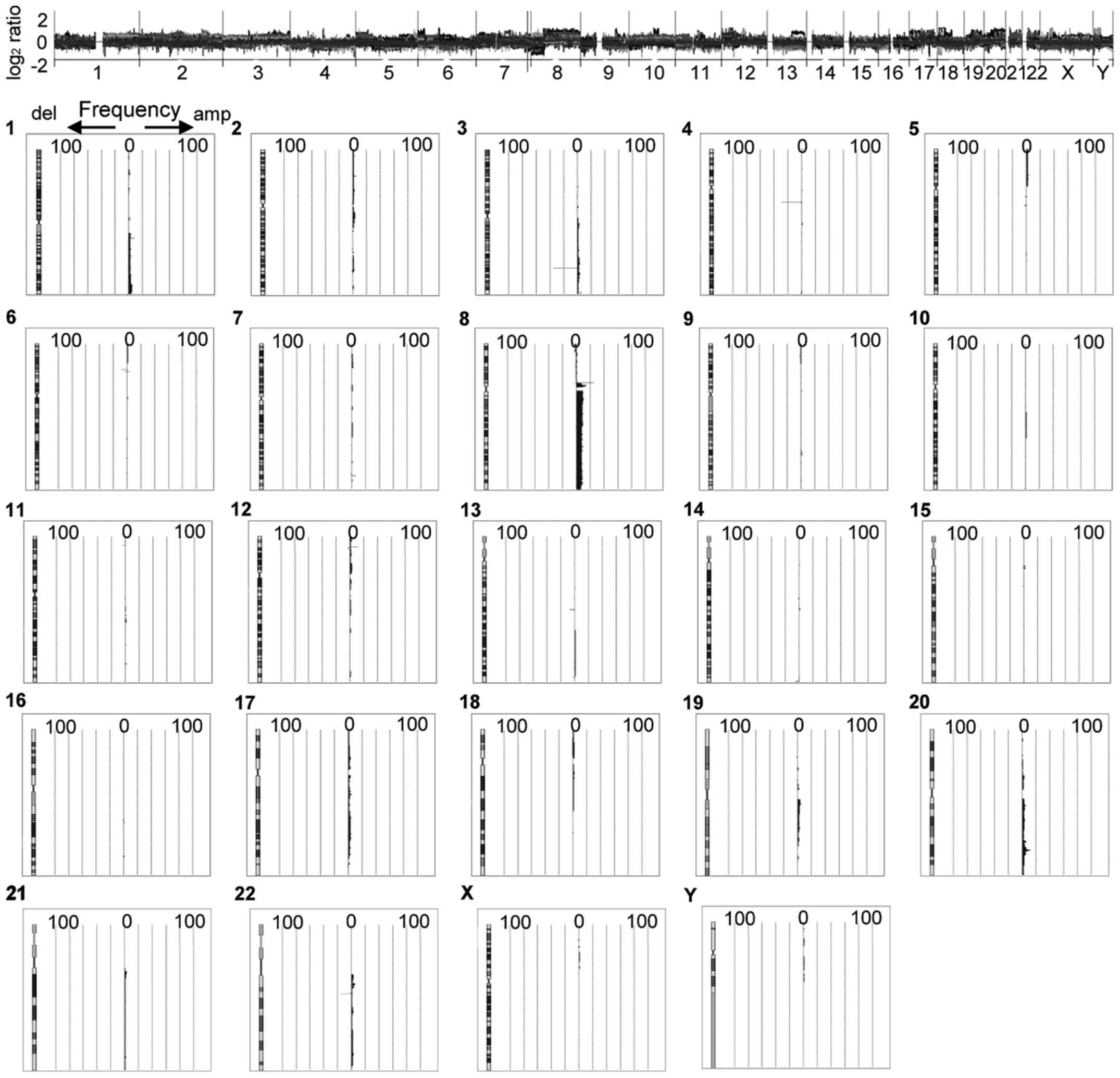
In our previous CGH analysis of somatic copy number
alterations in OCCC (8), we
selected regions with a minimum average log2 ratio ±0.25
(8). In the present study, we
reanalyzed the CGH data from 110 OCCC patients: regions with an
average log2 ratio ≥0.9 or ≤-1.5 were defined as those
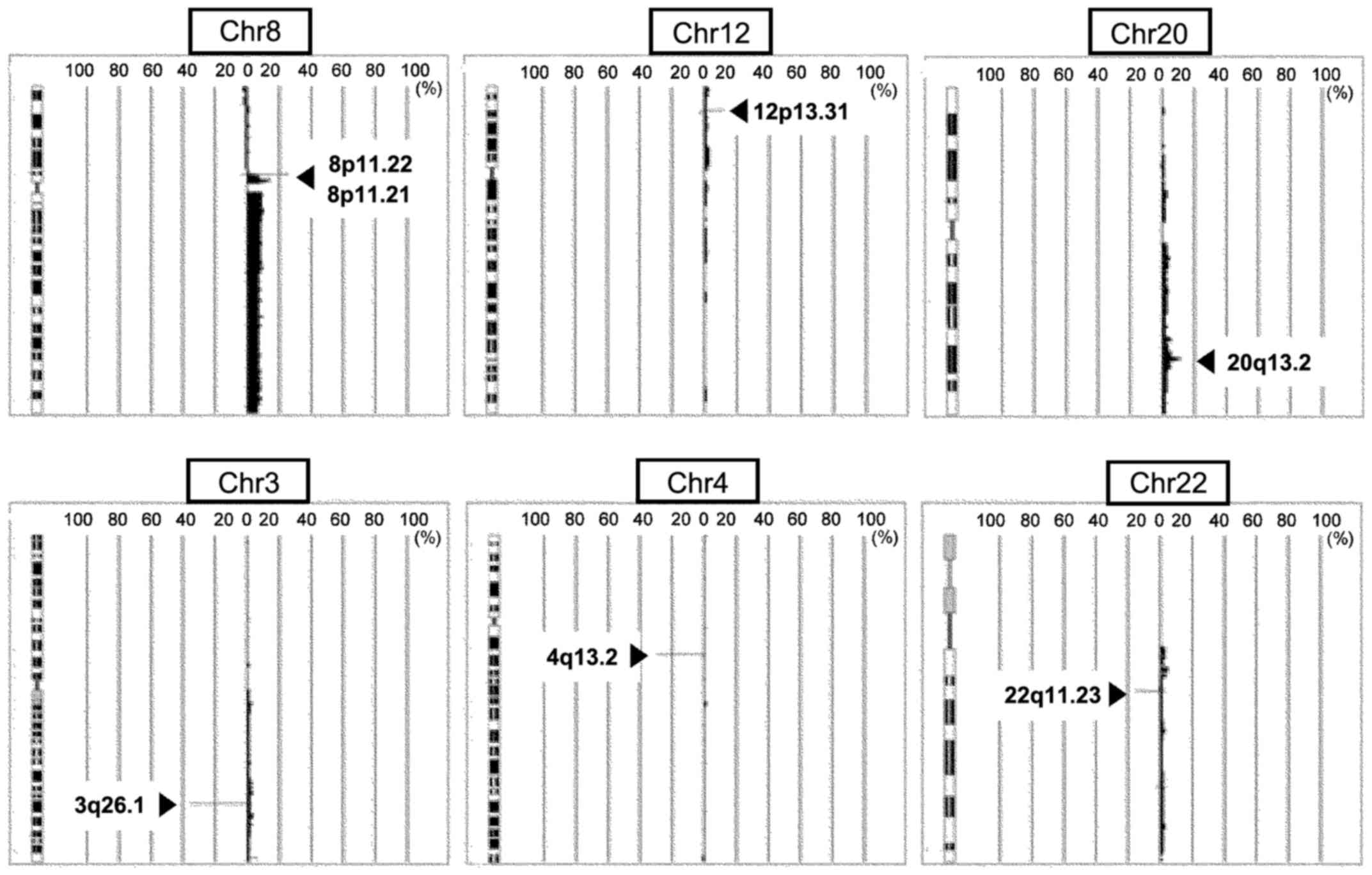
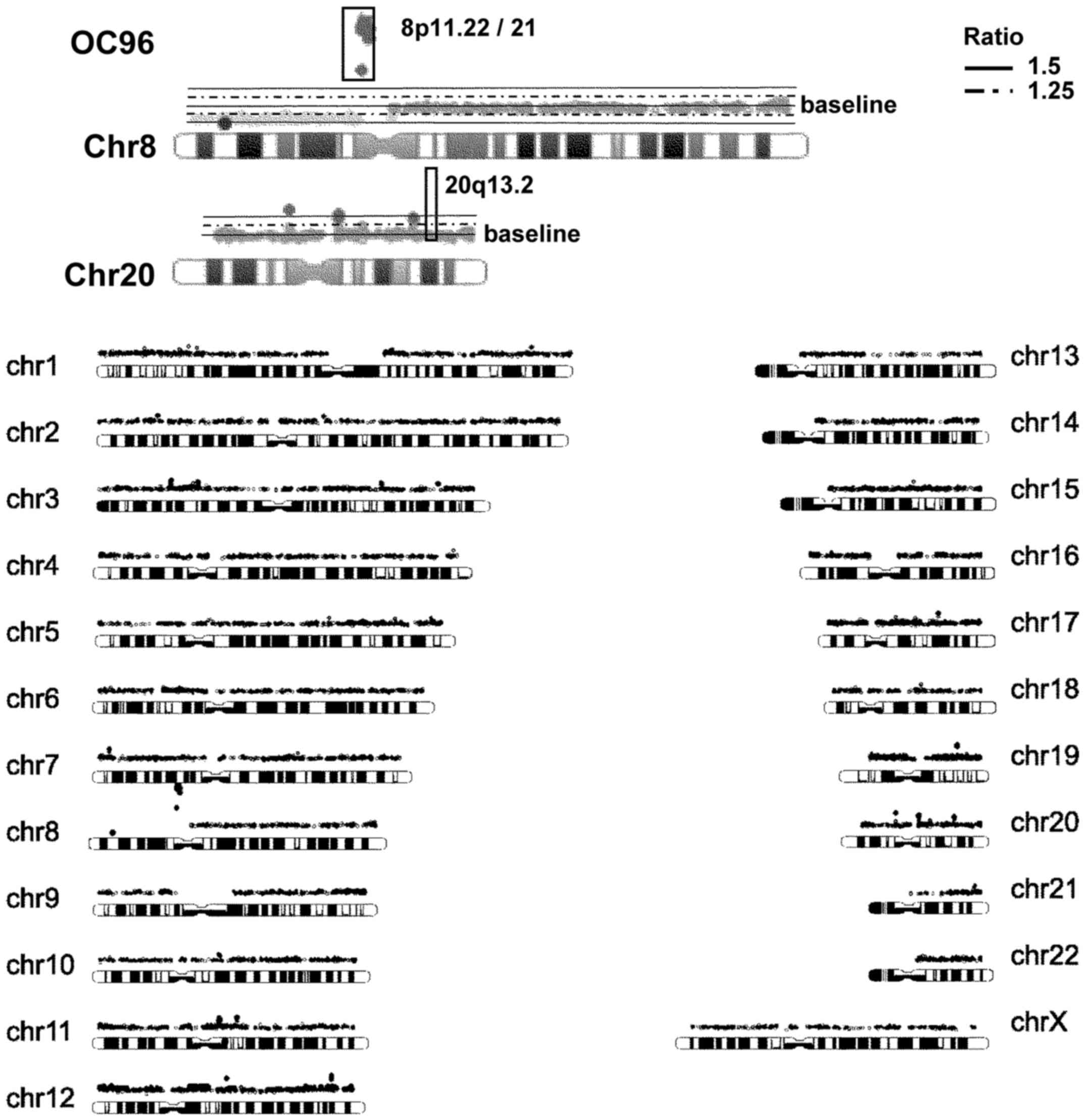
with a copy number gain or loss, respectively (Figs. 1 and 2). We identified increases in DNA copy
number of the chromosomal region 8p11.22, which harbors ADAM5p and
ADAM3A, in tumor tissues from ~25% of patients; 8p11.21, harboring
a number of genes, including ANK1, MYST3/KAT6A, IKBKB, POLB and
DKK4, in ~14–16% of patients; 12p13.31, harboring the long
non-coding RNAs (lncRNAs) ENST00000518709 and ENST00000545710, in
~13% of patients; and 20q13.2, harboring the candidate oncogene
ZNF217, in 15% of patients (Fig. 1
and Table III). In addition, we
identified losses of small segments (regions with an average signal
ratio ≤-0.5) in the chromosomal regions 8p11.22 and 12p13.31 in
normal samples from 5 and 17% of OCCC patients, respectively.
| Table III.Copy number aberrations detected by
CGH analysis in 110 OCCC patient tissues. |
Table III.
Copy number aberrations detected by
CGH analysis in 110 OCCC patient tissues.
|
|
| Region |
|
|
|
|---|
|
|
|
|
|
|
|
|---|
| Chr no. | Cytoband | Start | End | Genes | No. of patients, n
(%) | P-value |
|---|
| Amplification
log2≥0.9 |
| Chr8 | 8p11.22 | 39237438 | 39386158 | ADAM5p/ADAM3A | 28 (25) | 1.928E-25 |
|
| 8p11.21 | 41640598 | 41889042 | ANK1/MYST3 | 16 (15) | 1.647E-17 |
|
|
| 42033118 | 42543909 |
PLAT/IKBKB/POLB/DKK4/VDAC/SLC20A2/SMIM19 | 14 (13) |
| Chr12 | 12p13.31 | 9637323 | 9693948 |
ENST00000518709/ENST00000545710 | 13 (12) | 5.38E-19 |
| Chr20 | 20q13.2 | 52163628 | 52215265 | ZNF217 | 14 (13) | 6.349E-26 |
| Deletion
log2≤1.5 |
| Chr3 | 3q26.1 | 162514534 | 162619141 | BC073807 | 36 (33) | 2.253E-34 |
| Chr4 | 4q13.2 | 69375140 | 69483277 | UGT2B17 | 21 (19) | 5.293E-27 |
| Chr22 | 22q11.23 | 24371205 | 24390254 |
GSTT1/LOC391322 | 4 (4) | 3.94E-15 |
We also identified losses of small segments in the
chromosomal region 3q26.1, which harbors the lncRNA BC073807;
4q13.2, harboring UDP-glucuronosyltransferase 2B17 (UGT2B17); and
22q11.23, harboring glutathione S-transferase θ 1 (GSTT1) and
D-dopachrome tautomerase-like (LOC391322), in tumor tissues from
33, 19 and 4% patients, respectively (Fig. 1 and Table III). In addition, we identified
losses of small segments in the chromosomal regions 3q26.1, 4q13.2
and 22q11.23 in normal samples from 45, 36 and 24% patients,
respectively.
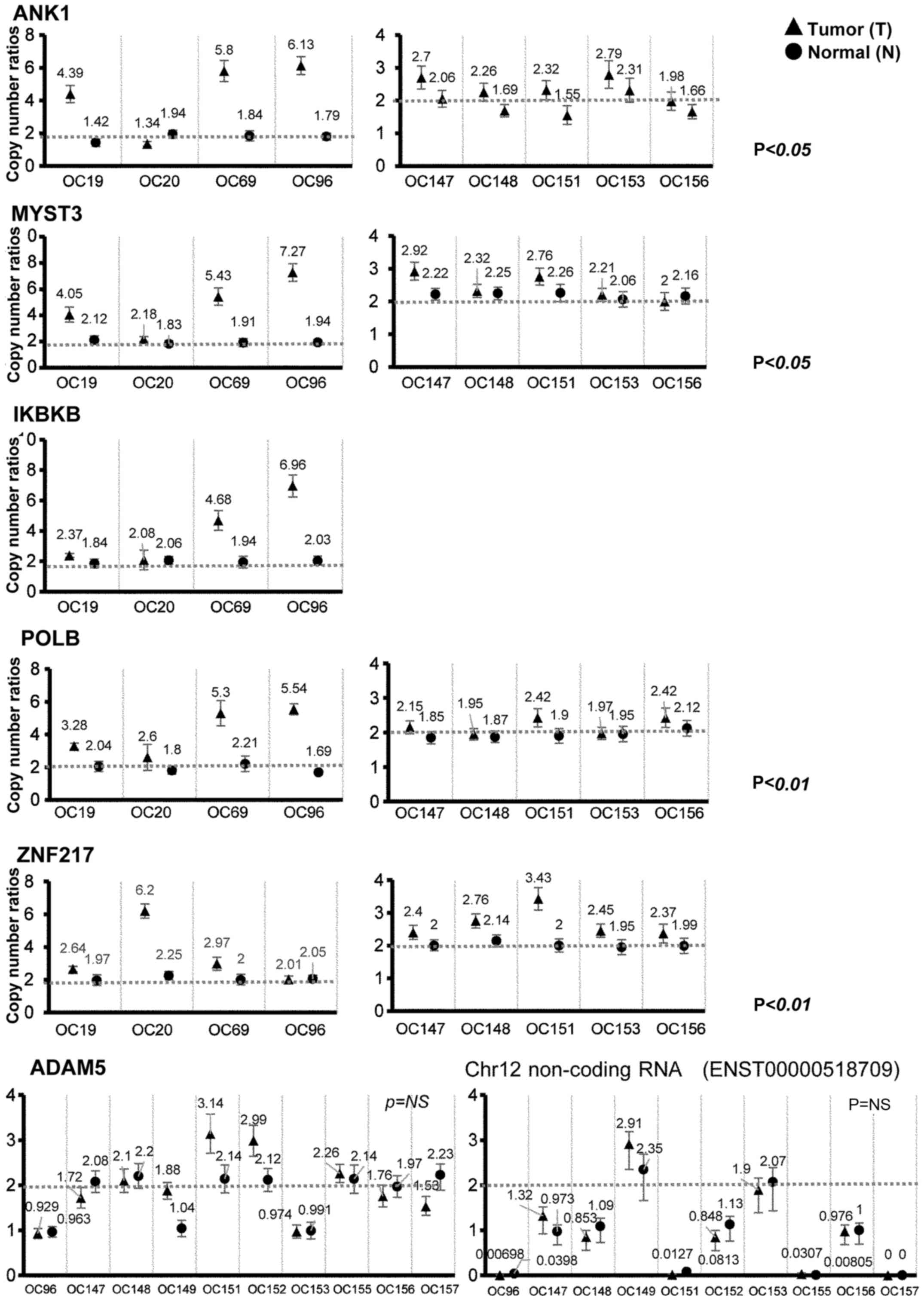
To validate these results, we performed ddPCR
analysis of copy number for ANK1, MYST3, IKBKB and POLB in four
OCCC patients (Fig. 4, left
panels). Consistent with the results of the CGH analysis, these
genes were amplified in three OCCC patients with 8p11.21
amplification, OC19, OC69 and OC96, but not in a patient without
8p11.21 amplification, OC20 (Fig.
3). We also confirmed that ZNF217 was amplified in a patient
with 20q13.2 amplification, OC20. Analysis of 9 patients confirmed
the amplification of these genes (Fig.
4; 4 patients in the left panels and 5 patients in the right
panels). ADAM5p and the lncRNA ENST00000518709 appeared to be
amplified, but no statistical significance was found (Fig. 4). In addition, consistent with the
results of CGH analysis, we observed losses of ADAM5p and the
lncRNA ENST00000518709 in normal samples in 3 out of 10 and 7 out
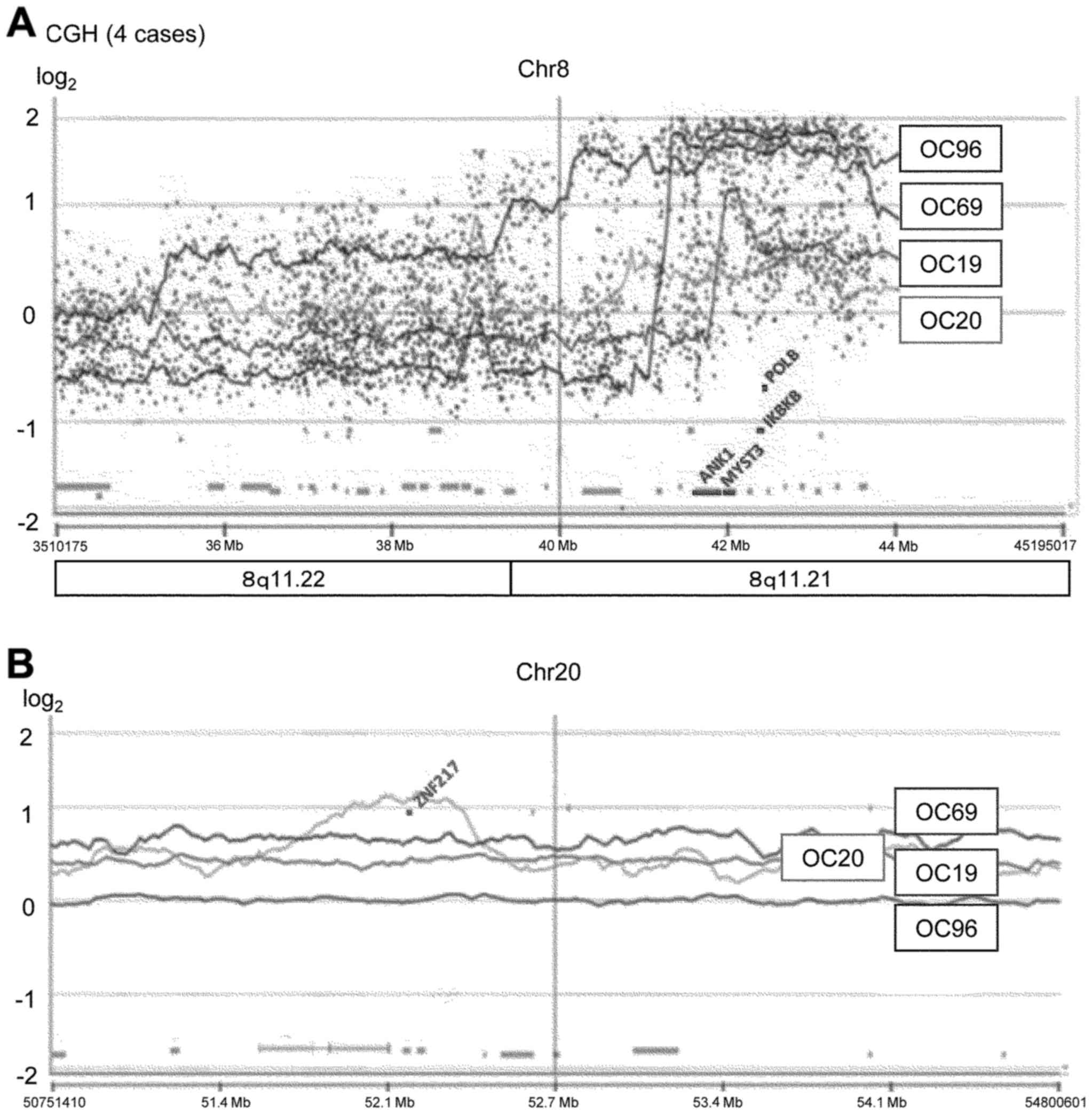
of 9 OCCC patients, respectively. Furthermore, we performed exome
sequence analysis of 3 patients and confirmed that 8p11.21, but not
20q13.2, was amplified in OC96 (Fig.
5 and Table IV).
| Table IV.Analyses of copy number aberrations
by CGH array, ddPCR and Exome-seq. |
Table IV.
Analyses of copy number aberrations
by CGH array, ddPCR and Exome-seq.
|
| Tumor/normal
ratio |
|---|
|
|
|
|---|
| Gene/method | OC19 | OC20 | OC69 | OC96 | OC147 | OC148 |
|---|
| ANK1 |
| CGH
array | 1.51 | 1.22 | 2.43 | 3.20 |
|
|
|
ddPCR | 3.09 | 0.69 | 3.15 | 3.42 | 1.31 | 1.33 |
|
Exome-seq |
|
|
| 3.66 | 0.83 | 1.20 |
| MYST3 |
| CGH
array | 2.14 | 1.22 | 2.42 | 3.20 |
|
|
|
ddPCR | 1.91 | 1.19 | 2.84 | 3.75 | 1.31 | 1.01 |
|
Exome-seq |
|
|
| 3.5 | 1.22 | 1.18 |
| IKBKB |
| CGH
array | 1.40 | 1.22 | 2.17 | 3.20 |
|
|
|
ddPCR | 1.28 | 1.00 | 2.41 | 3.74 |
|
|
|
Exome-seq |
|
|
| 3.55 |
|
|
| POLB |
| CGH
array | 1.41 | 1.22 | 2.18 | 3.20 |
|
|
|
ddPCR | 1.60 | 1.44 | 2.39 | 3.28 | 1.16 | 1.04 |
|
Exome-seq |
|
|
| 3.54 | 1.19 | 1.19 |
| ZNF217 |
| CGH
array | 1.26 | 2.02 | 1.27 | 1.03 |
|
|
|
ddPCR | 1.34 | 2.75 | 1.45 | 0.98 | 1.2 | 1.31 |
|
Exome-seq |
|
|
| 0.97 | 1.19 | 1.36 |
We finally investigated the relationship between the
amplification or loss of these genes and survival, sensitivity to
chemotherapy, thrombosis and endometriosis. The Kaplan-Meier
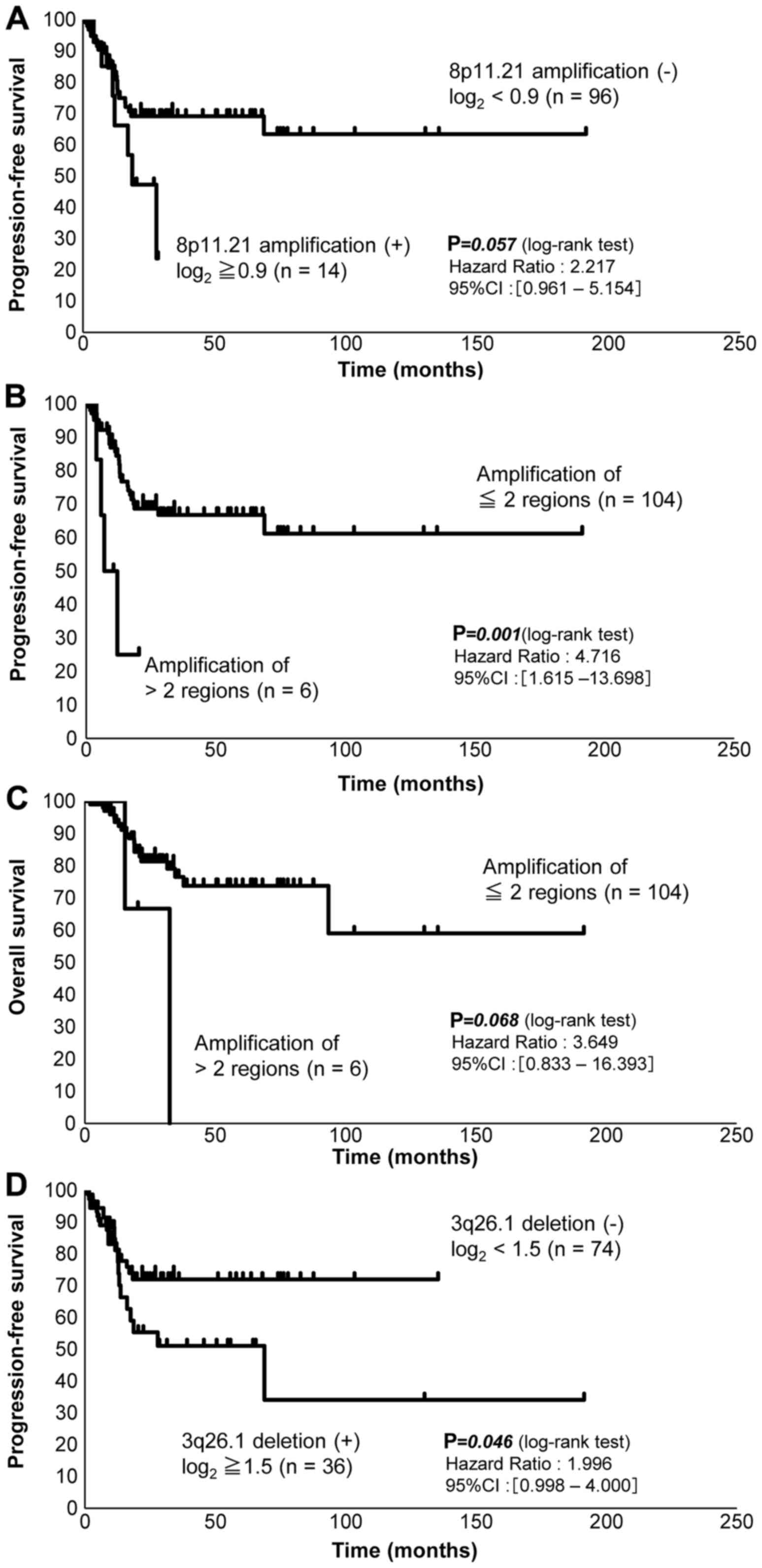
survival analysis showed that patients with amplification of
chromosome 8p11.21 had shorter PFS than those without such
amplification (P=0.057, 95% CI: 0.961–5.154) (Fig. 6A). Patients with amplification in
three of the four chromosomal regions 8p11.21, 8p11.22, 12p13.31
and 20q13.2 had shorter OS (P=0.068, 95% CI: 0.833–16.393) as well
as shorter PFS (P=0.001, 95% CI: 1.615–13.698) (Fig. 6B and C, Table V). Patients with a deletion in the
chromosomal region 3q26.1 had shorter PFS (P=0.046, 95% CI:
0.998–4.000) (Fig. 6D). Analysis of
the correlation between copy number alterations and sensitivity to
chemotherapy revealed that amplification of 12p13.3 or three of the
four chromosomal regions 8p11.21, 8p11.22, 12p13.31 and 20q13.2 was
associated with resistance to platinum-based chemotherapy (P=0.011
or 0.012, respectively) (Table
VI). We also found that a deletion in the chromosomal region
3q26.1 was associated with chemotherapy resistance (P=0.008)
(Table VI). We did not observe any
association between copy number alterations and sensitivity to
chemotherapy, FIGO stage, thrombosis or endometriosis.
| Table V.Multivariate analysis of prognostic
factors for 110 OCCC patients (n=110). |
Table V.
Multivariate analysis of prognostic
factors for 110 OCCC patients (n=110).
|
|
| Progression-free
survival | Overall
survival |
|---|
|
|
|
|
|
|---|
| Factors | Category | HR | 95% CI | P-value | HR | 95% CI | P-value |
|---|
| Age (years) | <60/≥60 | 1.125 | 0.462–2.740 | 0.795 | 0.797 | 0.288–2.212 | 0.664 |
| FIGO stage | III–IV/I–II | 6.211 | 2.949–12.987 | <0.001 | 3.937 | 1.612–9.615 | 0.003 |
| Race |
Japanese/Korean | 1.204 | 0.408–3.558 | 0.735 | 1.128 | 0.245–5.181 | 0.644 |
| Endometriosis
Amplification |
Absence/presence | 1.949 | 0.906–4.195 | 0.088 | 2.122 | 0.760–5.931 | 0.151 |
| >2 regions | Yes/no | 8.771 | 2.673–28.571 | <0.001 | 4.219 | 0.849–20.8333 | 0.078 |
| Table VI.Copy number alterations and
chemosensitivity of the OCCC cases. |
Table VI.
Copy number alterations and
chemosensitivity of the OCCC cases.
|
| 8p11.22 | 8p11.21 | 12p13.31 | 20q13.2 |
|
|
|---|
|
|
|
|
|
|
|
|
|---|
| Amplification | Yes | No | Yes | No | Yes | No | Yes | No | >2 regions | ≤2 regions |
|---|
| Sensitive | 15 | 46 | 6 | 55 | 3 | 58 | 8 | 53 | 1 | 60 |
| Resistant | 8 | 18 | 4 | 22 | 6 | 20 | 5 | 21 | 4 | 22 |
| Total | 23 | 64 | 10 | 77 | 9 | 78 | 13 | 74 | 5 | 82 |
| P-value | 0.550 | 0.458 | 0.011 | 0.464 | 0.012 |
|
| 3q26.1 | 4q13.2 |
22q11.23 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Deletion | Yes | No | Yes | No | Yes | No |
|
|
|
|
| Sensitive | 13 | 48 | 11 | 50 | 1 | 60 |
|
|
|
|
| Resistant | 13 | 13 | 5 | 21 | 2 | 24 |
|
|
|
|
| Total | 26 | 61 | 16 | 71 | 3 | 84 |
|
|
|
|
| P-value | 0.008 | 0.895 | 0.157 |
|
|
|
|
Discussion
In the present study, we showed that the chromosomal
regions 8p11.21, 8p11.22 and 12p13.31 are amplified and the
chromosomal regions 3q26.1, 4q13.2 and 22q11.23 are deleted in
OCCC. Kaplan-Meier survival analyses revealed that patients with
amplification within 8p11.21 or a deletion within 3q26.1 had a
shorter PFS than those without such copy number alterations.
Furthermore, patients with amplification in three of the four
chromosomal regions 8p11.21, 8p11.22, 12p13.31 and 20q13.2 had
shorter OS. Consistent with these results, we found that
amplification of 12p13.3 or three of the four chromosomal regions
8p11.21, 8p11.22, 12p13.31 and 20q13.2, or a deletion in the
chromosomal region 3q26.1 is associated with chemotherapy
resistance (Table VI). Thus,
amplification in 8p11.21, 8p11.22, 12p13.31 and 20q13.2 and a
deletion in the chromosomal region 3q26.1 may be critical for the
survival of OCCC patients.
The chromosomal region 8p11.21 encodes a number of
genes, including ANK1, MYST3, IKBKB and POLB, which have been
reported to play important roles in the development of a number of
cancers (13,14). It is therefore possible that
amplification of the genes encoded in this region contribute to the
development of OCCC.
ADAM5p and ADAM3A encoded in the chromosomal region
8p11.2 are pseudogenes of the ADAM metallopeptidases (15). It has recently been reported that
pseudogenes can function as lncRNAs, which can play important roles
in tumorigenesis. For example, PTENP1 functions as a competing
endogenous RNA to suppress tumor progression (16). UPAT, which is encoded by the
pseudogene of the amine oxidase copper containing-3 (AOC3) gene,
interacts with and stabilizes the epigenetic factor UHRF1 by
interfering with its β-TrCP-mediated ubiquitination, thereby
promoting the tumorigenicity of colorectal cancer cells (17). In addition, it has been reported
that the chromosomal region 8p11.22 is amplified and ADAM5p and
ADAM3A are overexpressed in conjunctival squamous cell carcinoma
and glioma (18,19). We also found that the lncRNA
ENST00000518709 is amplified in OCCC patients. Thus, it is possible
that amplification of these lncRNAs is important for the
development and progression of OCCC. In addition, we observed
losses of 8p11.22 and/or 12p13.31 in normal samples from 5 and 17%
of OCCC patients, respectively. The significance of these losses in
normal tissues remains to be investigated.
While we also found that the chromosomal regions
3q26.1, 4q13.2 and 22q11.23 were deleted in 30, 19 and 4% of OCCC,
respectively, we also identified losses of small segments in these
chromosomal regions in normal samples from 45, 36 and 24% of
patients, respectively. CGH analysis did not discriminate between
the segments deleted in normal samples and those deleted only in
tumor tissues. These loci encode the lncRNA BC073807, UGT2B17 (a
member of the uridine diphosphoglucuronosyltransferase protein
family), and GSTT1 and LOC391322, respectively. It remains to be
investigated whether deletion of these genes contributes to the
development of OCCC. It is also possible that deletion of these
regions may disrupt the three-dimensional networks of chromosomal
interactions and thereby lead to altered gene expression (20–22).
Elucidation of the functional significance of these deletions in
the development of OCCC is underway in our laboratories.
In conclusion, we identified somatic copy number
alterations that have prognostic value in patients with OCCC. We
also identified copy number alterations associated with
chemotherapy resistance. However, it remains to be confirmed
whether amplification of these genes indeed results in changes in
the expression of encoded proteins. Furthermore, it remains to be
seen whether amplification or deletion of these genes contributes
to the development, progression and/or chemosensitivity of OCCC.
The identification and characterization of these genes could
provide novel insights into the mechanisms of OCCC development and
chemosensitivity. We speculate that these genes could be promising
molecular targets and/or markers for OCCC therapy.
Acknowledgements
Not applicable.
Funding
The present study was supported by Grants-in-Aid for
Scientific Research on Innovative Areas (Integrative Analysis and
Regulation of Cellular Diversity, no. 17H06325) from MEXT, Japan,
and Project for Cancer Research and Therapeutic Evolution
(P-CREATE, no. 17cm0106103h0002) from the Japan Agency for Medical
Research and Development.
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
AM, TH, AO and TA conceived and designed the
experiments. AM, MK, YK, TI and KS performed the experiments. AM,
TH and TA analyzed the data. MU and AO provided
reagents/materials/analysis tools. AM, TH and TA wrote the report.
All authors read and approved the manuscript and agree to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the study are appropriately
investigated and resolved.
Ethics approval and consent to
participate
All the OCCC patients provided written informed
consent. This study was performed in accordance with the Helsinki
Declaration and was approved by the Ethics Committee of the Jikei
University School of Medicine
Consent for publication
Not applicable.
Competing interests
The authors state that they have no competing
interests.
References
|
1
|
Gilks CB, Ionescu DN, Kalloger SE, Köbel
M, Irving J, Clarke B, Santos J, Le N, Moravan V and Swenerton K:
Cheryl Brown Ovarian Cancer Outcomes Unit of the British Columbia
Cancer Agency: Tumor cell type can be reproducibly diagnosed and is
of independent prognostic significance in patients with maximally
debulked ovarian carcinoma. Hum Pathol. 39:1239–1251. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Matsuda T, Marugame T, Kamo K, Katanoda K,
Ajiki W and Sobue T: Japan Cancer Surveillance Research Group:
Cancer incidence and incidence rates in Japan in 2006; Based on
data from 15 population-based cancer registries in the monitoring
of cancer incidence in Japan (MCIJ) project. Jpn J Clin Oncol.
42:139–147. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mackay HJ, Brady MF, Oza AM, Reuss A,
Pujade-Lauraine E, Swart AM, Siddiqui N, Colombo N, Bookman MA,
Pfisterer J, et al: Gynecologic Cancer InterGroup: Prognostic
relevance of uncommon ovarian histology in women with stage III/IV
epithelial ovarian cancer. Int J Gynecol Cancer. 20:945–952. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Duska LR, Garrett L, Henretta M, Ferriss
JS, Lee L and Horowitz N: When ‘never-events’ occur despite
adherence to clinical guidelines: The case of venous
thromboembolism in clear cell cancer of the ovary compared with
other epithelial histologic subtypes. Gynecol Oncol. 116:374–377.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kuo KT, Mao TL, Jones S, Veras E, Ayhan A,
Wang TL, Glas R, Slamon D, Velculescu VE, Kuman RJ, et al: Frequent
activating mutations of PIK3CA in ovarian clear cell carcinoma. Am
J Pathol. 174:1597–1601. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kuo KT, Mao TL, Chen X, Feng Y, Nakayama
K, Wang Y, Glas R, Ma MJ, Kurman RJ, Shih IeM, et al: DNA copy
numbers profiles in affinity-purified ovarian clear cell carcinoma.
Clin Cancer Res. 16:1997–2008. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tan DS, Iravani M, McCluggage WG, Lambros
MB, Milanezi F, Mackay A, Gourley C, Geyer FC, Vatcheva R, Millar
J, et al: Genomic analysis reveals the molecular heterogeneity of
ovarian clear cell carcinomas. Clin Cancer Res. 17:1521–1534. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Okamoto A, Sehouli J, Yanaihara N, Hirata
Y, Braicu I, Kim BG, Takakura S, Saito M, Yanagida S, Takenaka M,
et al: Somatic copy number alterations associated with Japanese or
endometriosis in ovarian clear cell adenocarcinoma. PLoS One.
10:e01169772015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tian C, Kosoy R, Lee A, Ransom M, Belmont
JW, Gregersen PK and Seldin MF: Analysis of East Asia genetic
substructure using genome-wide SNP arrays. PLoS One. 3:e38622008.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Miotke L, Lau BT, Rumma RT and Ji HP: High
sensitivity detection and quantitation of DNA copy number and
single nucleotide variants with single color droplet digital PCR.
Anal Chem. 86:2618–2624. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Langmead B and Salzberg SL: Fast
gapped-read alignment with Bowtie 2. Nat Methods. 9:357–359. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li H, Handsaker B, Wysoker A, Fennell T,
Ruan J, Homer N, Marth G, Abecasis G and Durbin R: 1000 Genome
Project Data Processing Subgroup: The sequence alignment/map format
and SAMtools. Bioinformatics. 25:2078–2079. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Omura N, Mizuma M, MacGregor A, Hong SM,
Ayars M, Almario JA, Borges M, Kanda M, Li A, Vincent A, et al:
Overexpression of ankyrin1 promotes pancreatic cancer cell growth.
Oncotarget. 7:34977–34987. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Guo JP, Shu SK, He L, Lee YC, Kruk PA,
Grenman S, Nicosia SV, Mor G, Schell MJ, Coppola D, et al:
Deregulation of IKBKE is associated with tumor progression, poor
prognosis, and cisplatin resistance in ovarian cancer. Am J Pathol.
175:324–333. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Brocker CN, Vasiliou V and Nebert DW:
Evolutionary divergence and functions of the ADAM and ADAMTS gene
families. Hum Genomics. 4:43–55. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Marsit CJ, Zheng S, Aldape K, Hinds PW,
Nelson HH, Wiencke JK and Kelsey KT: PTEN expression in
non-small-cell lung cancer: Evaluating its relation to tumor
characteristics, allelic loss, and epigenetic alteration. Hum
Pathol. 36:768–776. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Taniue K, Kurimoto A, Sugimasa H, Nasu E,
Takeda Y, Iwasaki K, Nagashima T, Okada-Hatakeyama M, Oyama M,
Kozuka-Hata H, et al: Long noncoding RNA UPAT promotes colon
tumorigenesis by inhibiting degradation of UHRF1. Proc Natl Acad
Sci USA. 113:1273–1278. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Barrow J, Adamowicz-Brice M, Cartmill M,
MacArthur D, Lowe J, Robson K, Brundler MA, Walker DA, Coyle B and
Grundy R: Homozygous loss of ADAM3A revealed by genome-wide
analysis of pediatric high-grade glioma and diffuse intrinsic
pontine gliomas. Neuro-oncol. 13:212–222. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Asnaghi L, Alkatan H, Mahale A, Othman M,
Alwadani S, Al-Hussain H, Jastaneiah S, Yu W, Maktabi A, Edward DP,
et al: Identification of multiple DNA copy number alterations
including frequent 8p11.22 amplification in conjunctival squamous
cell carcinoma. Invest Ophthalmol Vis Sci. 55:8604–8613. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dekker J: Gene regulation in the third
dimension. Science. 319:1793–1794. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Andrey G, Montavon T, Mascrez B, Gonzalez
F, Noordermeer D, Leleu M, Trono D, Spitz F and Duboule D: A switch
between topological domains underlies HoxD genes collinearity in
mouse limbs. Science. 340:12341672013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nora EP, Lajoie BR, Schulz EG, Giorgetti
L, Okamoto I, Servant N, Piolot T, van Berkum NL, Meisig J, Sedat
J, et al: Spatial partitioning of the regulatory landscape of the
X-inactivation centre. Nature. 485:381–385. 2012. View Article : Google Scholar : PubMed/NCBI
|