Introduction
Cancer accounts for 1 in 8 deaths worldwide and is
rapidly becoming a global pandemic. Unfortunately, there are two
key obstacles to the effective treatment of cancer: drug toxicities
and multi-drug resistance (MDR). Current methods for treating
cancer with chemotherapy have severe toxicities that can include
myelosuppression, neurotoxicity, immune-suppression and mutagenic
and carcinogenic effects (1–3). In
addition, complete elimination of tumor cells post-treatment with
chemotherapy is hindered by the development of MDR. The main cause
for the development of MDR in cancer cells has been attributed to
the overexpression of ABC drug transporters, resulting in a
decrease in drug uptake or an increase in drug efflux from the
cancer cells.
Currently, there is increased interest in using and
developing natural products that overcome the problems of toxicity
and drug resistance of chemotherapy agents. Natural products from
several different sources have been identified as potential novel
therapeutic candidates for cancer treatment, including products
derived from plants, animals, vitamins, minerals and microorganisms
(4). These include vitamin E
(5), tetrandrine that is a
bisbenylisoquinoline alkaloid from the root of Stephenia
tetrandra (6), the natural
isoflavone compound genistein (7)
and green tea that contains flavonoids and catechins (8). In addition, we have developed several
natural biological response modifiers (BRMs) which exhibit
anticancer effects against several cancer types and have minimal,
if any, side effects. These include MGN-3/Biobran, an arabinoxylan
from rice bran (9–11); marina crystal minerals (MCM), a
crystallized mixture of minerals and trace elements from sea water
(12), Thymax, gross thymic extract
(13) and PFT, a novel kefir
product (14).
Several of the above natural dietary products that
have been shown to act as potent anticancer agents have
subsequently also been shown to possess effective chemosensitizing
properties. For example, MGN-3/Biobran has been shown to sensitize
cancer cells to several chemotherapeutic agents such as cisplatin,
doxorubicin, daunorubicin (DNR), adriamycin and paclitaxel in
vitro and in vivo (10,15–18),
and it has been shown to enhance the effects of interventional
therapies for the treatment of hepatocellular carcinoma (19). Curcumin can sensitize tumors to
different chemotherapeutic agents by mechanisms that include MDR
modulation (20,21). Restoration of drug sensitivity has
been achieved by several agents including vitamin E, which has been
shown to act as a P-gp inhibitor (22), tetrandrine (6,23,24)
and the flavonoid quercetin, which is an MDR modulator and thus a
potential chemosensitizer (25).
We also found that bakers and brewers yeast,
Saccharomyces cerevisiae, acts as an anticancer agent. This
natural product is a commercially available food supplement and a
necessary component for the production of fermented foods (such as
bread and beer). Its anticancer activity has been evidenced in
in vitro studies involving several human cancer cell lines
that undergo apoptosis upon the phagocytosis of killed S.
cerevisiae. These cancer cells include breast, tongue and colon
(26–29). In addition, S. cerevisiae can
induce apoptosis in nude mice bearing human breast cancer (30,31)
and in Swiss albino mice bearing Ehrlich carcinoma (32). Studies have shown that one of the
metabolites of yeast, trehalose, exerts anticancer effects when
combined with synthetic agents. Such yeast metabolites may
represent another mechanism by which yeast exhibits an anticancer
effect (33). Furthermore, we have
recently shown that S. cerevisiae sensitizes different
mammary cancer cells to chemotherapy in vitro, with the
IC50 value for paclitaxel being significantly reduced
against breast cancer cells in the presence of S. cerevisiae
(34).
In the present study, we evaluate the ability of
S. cerevisiae to sensitize cancer cells to chemotherapy,
paclitaxel, using mice bearing Ehrlich ascites carcinoma (EAC).
While our previous study demonstrated yeasts ability to sensitive
human and murine breast cancer cells to chemotherapeutic agents
in vitro (34), the present
study sought to explore whether this in vitro effect is
observable in vivo using an EAC mouse model. EAC is a
spontaneous murine mammary adenocarcinoma (35) that appeared first as a spontaneous
breast cancer in a female mouse and has been commonly used as an
experimental tumor by transplanting tumor tissues subcutaneously
from mouse to mouse (36). EAC is
an undifferentiated carcinoma, is originally hyperdiploid, and has
high transplantable capability, rapid proliferation, shorter life
span and 100% malignancy (37). EAC
models are frequently utilized by our group and others in the
development of anti-tumorigenic agents (34,38).
Results of the present study show that S. cerevisiae can
sensitize EAC cells in mice to paclitaxel by mechanisms involving
induction of apoptosis. In addition, S. cerevisiae combined
with paclitaxel at low dose has a more significant anticancer
effect than paclitaxel alone at high dose. The present study shows
that yeast is a potent chemosensitizer and it may have clinical
implications for the treatment of breast cancer.
Materials and methods
Paclitaxel (Taxol®)
Paclitaxel was purchased from Bristol-Myers Squibb
Inc. (Princeton, NJ, USA). It was supplied with an initial
concentration of 100 mg/16.7 ml. Each ml of sterile non-pyrogenic
solution contains 6 mg paclitaxel, 527 mg of purified
Cremophor® EL (polyoxyethylated castor oil) and 49.7%
(vol/vol) dehydrated alcohol, United States Pharmacopeia (USP).
Preparation of S. cerevisiae
Commercially available bakers and brewers yeast,
S. cerevisiae, was used in suspensions that were washed once
with phosphate-buffered saline (PBS). It was then incubated for 1 h
at 90°C to kill the yeast and washed three times with PBS.
Quantification was carried out using a hemocytometer, and cell
suspensions were adjusted to 1×107 cells/ml (34). The yeast was given locally to
mice-bearing EAC at 0.1 ml/mouse three times a week for 25 days
starting from day 8 of tumor cell inoculation.
Animals
Eighty female Swiss albino mice (2 months old)
weighing 19–21 g were used in the present study. The mice were
purchased from the National Cancer Institute, Cairo University,
(Cairo, Egypt) and were housed in our animal research facility at
constant temperature (24°C±2°C) 50°F at 10% relative humidity, and
alternating 12-h light/dark cycles. Mice were accommodated for 1
week prior to experiments. Animals were provided with standard cube
pellets and water ad libitum. The diet consists of casein
(12.5%), fats (1.0%), wheat flour (80%), bran (3.3%), olive oil
(2.3%), DL-methionine (0.5%), vitamins and salt mixture (0.2%) and
water (0.2%). The ratio of total calories was ~18% protein, 73%
carbohydrate and 9% fat. The pellets were purchased from Misr Oil
& Soap Co. (Cairo, Egypt). The actual food intake was
previously monitored and found to be from 4 to 5 g/day/animal
weighing 20±2 g. Animal protocols were in compliance with the Guide
for the Care and Use of Laboratory Animals at the University of
Mansoura, Egypt.
Preparation of Ehrlich ascites
carcinoma (EAC) cells and tumor transplantation
EAC cells were kindly supplied by the National
Cancer Institute, Cairo University, (Cairo, Egypt) and were
maintained by weekly intraperitoneal transplantation of
2.5×106
cells in female Swiss albino mice. In this experiment, solid tumors
were produced by intramuscular injection with 0.2 ml EAC cells
(2.5×106 cells) in the right thigh of the lower limb of
the mice. Viability of the tumor cells was assessed to be 95% by
trypan blue dye exclusion method.
Experimental design
At 8 days post-tumor cell inoculation, mice bearing
a solid EAC tumor mass of ~100 mm3 were used in the
study. Mice were divided into eight groups (n=10): i) normal
control untreated mice; ii) control mice treated with yeast alone;
iii) mice bearing tumors receiving intratumoral (i.t.) injections
of PBS (EAC); iv) mice bearing tumors receiving i.t. injections of
yeast (1×107 cells/ml); v) mice bearing tumors receiving
paclitaxel at a high dose (paclitaxel-H) (10 mg/kg); vi) mice
bearing tumors receiving paclitaxel at a low dose (paclitaxel-L) (2
mg/kg); vii) mice bearing tumors receiving yeast plus paclitaxel-H;
and viii) mice bearing tumor receiving yeast plus paclitaxel-L.
Mice received paclitaxel and/or yeast 3 days/week commencing on day
8 post EAC cell inoculation until the end of the experiment (day
30). On day 30, animals were anesthetized and sacrificed by
cervical dislocation. The tumors were excised and immediately
frozen or fixed in 10% neutral formalin for histopathological
examination or in 2.5 glutaraldehyde for electron microscopy.
Evaluation of body weight and tumor
weight
Body weight (BW)
Mice were examined for initial, final and net BWs at
day 30. The net final BW = final BW - tumor weight. BW gain was
determined as the difference between the initial and the net final
BW.
Tumor weight (TW)
On day 30, the mice were euthanized and the solid
tumors were excised for weight (TW/g) determination before
freezing.
Flow cytometric analysis
Cell preparation for flow
cytometry
Tumor tissues were excised from EAC-bearing mice
under different treatment conditions, cut into pieces, and gently
rubbed through fine nylon gauze (40–50 mesh count/cm; HD 140
Zuricher Buteltuch fabrik AG, Zürich, Switzerland). Samples were
then washed through the gauze with Tris-ethylenediaminetetraacetic
acid (Tris-EDTA) buffer at pH 7.5 [3.029 g of 0.1 M
Tris-(hydroxymethyl aminomethane), 1.022 g of 0.07 M HCl and 0.47 g
of 0.005 M Tris-EDTA]. Cells were suspended in PBS, centrifuged for
5 min at 200–300 × g, resuspended in sterile PBS (cell density
~1×106 cells/ml), and then fixed in 70% ice-cold ethanol
in PBS and stored at −20°C until used.
Analysis of apoptosis, DNA damage and
cell proliferation assay
Quantitative detection of apoptosis with cleaved
poly(ADP-ribose) polymerase (PARP), DNA damage using phosphorylated
H2AX (γH2AX form), and cell proliferation by bromodeoxyuridine
(BrdU) were simultaneously determined in vivo by multicolor
flow cytometric analysis using the Apoptosis, DNA Damage and Cell
Proliferation kit specific to incorporated BrdU, γH2AX and cleaved
PARP (BD Pharmingen; BD Biosciences, San Diego CA, USA) following
the manufacturers instructions.
Annexin-V/PI
Annexin V is a protein that binds to
phosphatidylserine (PS) residues which are exposed on cell surfaces
of apoptotic, but not normal cells. During apoptosis, the PS groups
are exposed to the exterior of the cell membrane. This binding of
PS with Annexin V is an established biochemical marker of
apoptosis. Induction of apoptosis caused by yeast and paclitaxel in
EAC-primary tumors was quantitatively determined through flow
cytometry using the Annexin V-conjugated Alexa Fluor 488 Apoptosis
Detection kit following the manufacturers instructions (BD
Biosciences).
Flow cytometric analysis of T helper
cells (CD4+) and T cytotoxic cells
(CD8+)
Flow cytometric analysis (FACS) of CD4+
and CD8+ T cells that infiltrated EAC tumors was
performed using mouse anti-CD4+ FITC (clone GK1.5) and
mouse anti-CD8+ FITC (clone 53–6.7) (BD Biosciences).
Tumor cell suspensions in PBS at a concentration of
1×106 cells/ml were prepared, centrifuged for 5 min at
200–300 × g, and the supernatant was discarded. Afterwards the cell
pellets were re-suspended in 500 µl PBS. Subsequently, 1 ml of
suspension was dispensed in a flow cytometric tube. Cells were
incubated with 25 µl of anti-CD4+ or
anti-CD8+ in the dark for 30 min at 4°C. The supernatant
was discarded and the cells were washed twice by PBS, pH (7.2).
Then, 200 µl paraformaldehyde solution was added to each tube,
mixed well and maintained in the dark at 4°C until FACS analysis
following the manufacturers instructions.
Immunostaining for proliferation
marker Ki-67
Ki-67 is a cell proliferation marker for tumor
progression commonly used as an early predictor in breast cancer.
Immunohistochemical staining was performed on formalin-fixed
paraffin-embedded tissues. Staining was performed using
streptoavidin-biotin method as previously described (39) by using the Histostain-Plus kit
(Santa Cruz Biotechnology, Santa Cruz, CA, USA) which contains 10%
non-immune serum, biotinylated secondary antibody and
streptoavidin-peroxidase. Immunohistochemical reactivities of Ki-67
expression in the different groups were quantified by image
analysis (Immuno Ratio-JPEG2000 virtual slide microscope; Institute
of Biomedical Technology, University of Tampere, Tampere,
Finland).
Histopathological studies
At 30 days post inoculation, the animals were
sacrificed, and the tumor specimens under different treatment
conditions were removed and fixed in 10% neutral buffered formalin
for at least 24 h. The tissues were embedded in paraffin using a
routine method and then cut into 3–4 µm thickness with a microtome
and stained with hematoxylin and eosin (H&E). The
H&E-stained slides were examined under a light microscope
(magnification, ×4-x40).
Electron microscopy (EM)
Small fragments (1 mm3 blocks) of tissue
were fixed in 2.5% glutaraldehyde, and then put into sodium
cacodylate buffer the night before processing and postfixed with
osmium tetroxide. The tissues were then embedded in resin.
Semi-thin sections were cut using a glass knife and stained with
toluidine blue. Semi-thin sections are required to confirm the
presence of desired cells and to select the best tissue block for
thin-sectioning. Finally, thin sections were cut on a diamond
knife. Routine staining was performed with uranyl acetate followed
by lead citrate (40), and examined
using JEOL Electron Microscope (JEOL Ltd., Tokyo, Japan) operating
at 60 kV.
Statistical analysis
Values are reported as mean ± standard error (SE)
and data were analyzed using one-way analysis of variance (ANOVA)
followed by least significant difference (LSD) post hoc test for
multiple comparisons. P<0.05 was considered to indicate a
statistically significant result.
Results
This study evaluated the
chemosensitizing effect of yeast in mice bearing tumor
Parameters under investigation included body weight,
tumor weight, cell proliferation, DNA damage, apoptosis in tumor
cells, CD4+ and CD8+ T cells infiltrated in
tumor tissue and Ki-67 expression. Histopathology and
ultra-structural examinations were also used. In addition, adverse
side-effects from the yeast treatment and/or paclitaxel were also
monitored.
Changes in the body weight (BW)
As shown in Fig. 1,
all animal groups showed an increase in BW after one month of
treatment, as compared to the initial BW. The percentage of weight
gain compared to initial body weight was as follows: i) control
untreated mice, 15.5%; ii) yeast-treated control mice, 15.4%; iii)
untreated EAC-bearing mice showed only 2.0%; iv) yeast-treated
EAC-bearing mice showed 9.3%; v and vi) paclitaxel-treated (high
and low doses) EAC-bearing mice showed 9.4 and 7.0%, respectively;
and vii and viii) paclitaxel (high and low) combined with yeast was
18 and 13.7%, respectively. The results demonstrated that the
combined treatment with yeast and paclitaxel-L showed an increase
of 13.7% in BW, while treatment with paclitaxel-H alone presented
an increase of 9.4%.
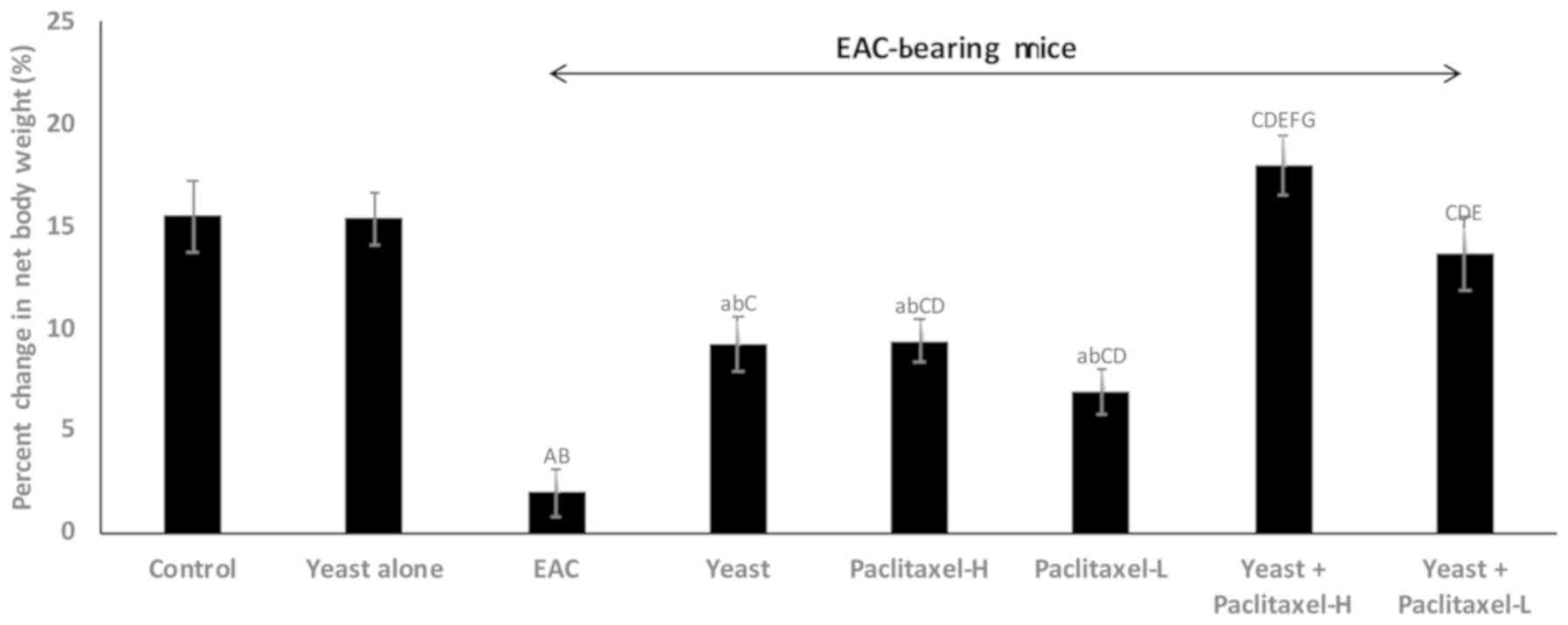 | Figure 1.Effect of yeast and/or paclitaxel on
body weight change (g). Data are expressed as mean ± SE of 10
mice/group). aP<0.05 and AP<0.01,
significantly different from the normal control untreated group.
bP<0.05 and BP<0.01, significantly
different from the control treated with yeast group.
CP<0.01, significantly different from the EAC group.
DP<0.01, significantly different from the
yeast-treated EAC group. EP<0.01, significantly
different from the paclitaxel-H-treated EAC group.
FP<0.01, significantly different from the
paclitaxel-L-treated EAC group. GP<0.01,
significantly different from the yeast + paclitaxel-H-treated EAC
group. Net final body weight = (final body weight - tumor weight).
Body weight change = (net final body weight - initial body weight).
EAC, Ehrlich ascites carcinoma. Groups: EAC, mice bearing tumors
receiving intratumoral (i.t.) injections of PBS; Yeast, mice
bearing tumors receiving i.t. injections of yeast (1×107
cells/ml); paclitaxel-H, mice bearing tumors receiving paclitaxel
at a high dose (10 mg/kg); paclitaxel-L, mice bearing tumors
receiving paclitaxel at a low dose (2 mg/kg); Yeast + Paclitaxel-H,
mice bearing tumors receiving yeast plus paclitaxel-H; Yeast +
Paclitaxel-L, mice bearing tumors receiving yeast plus
paclitaxel-L. |
Animals were monitored to observe potential toxic
side effects of yeast treatment. Injections (i.t.) of S.
cerevisiae alone or in the presence of paclitaxel showed no
adverse side-effects as indicated by normality of feeding/drinking
and life activity patterns for the entire treatment period.
Changes in tumor weight (TW)
The percent change in TW on day 30 in the different
groups, as compared to EAC-bearing mice (5.3±0.11 g) was examined.
Data depicted in Fig. 2 show
significant suppressive effects on TW by individual treatments,
including the following: yeast alone −46.8%, paclitaxel-H alone
−52.6% and paclitaxel-L alone −33.6%; and combined treatments with
yeast plus paclitaxel-H and yeast plus paclitaxel-L showed −75.1
and −63.7%, respectively. Results of this study revealed that
combined treatment with yeast and paclitaxel-L showed a more
significant decrease in TW (63.7%) than that caused by paclitaxel-H
alone (−52.6%). Similar trends were observed in the last tumor
volume (data not shown).
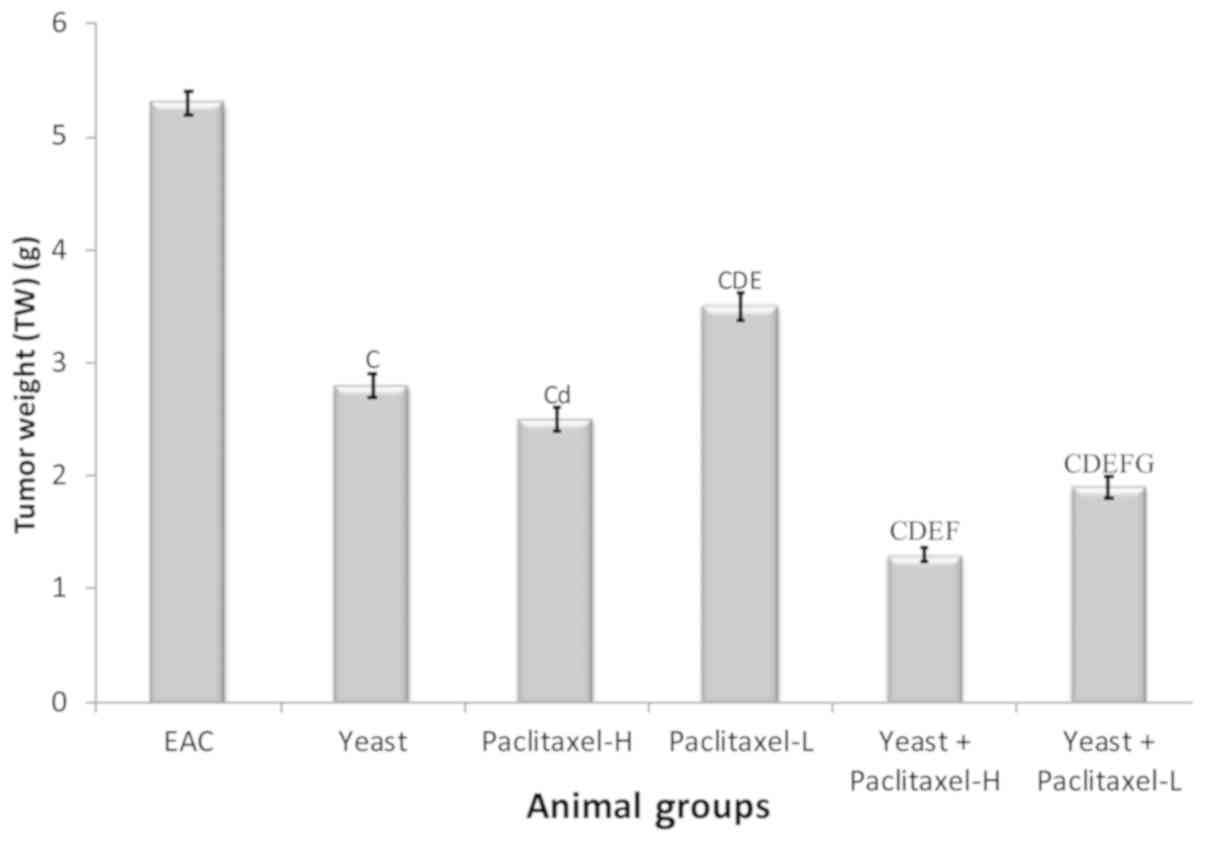 | Figure 2.Tumor weight of the EAC mouse-bearing
tumors treated with yeast in the presence or absence of paclitaxel.
Data are expressed as mean ± SE of 10 mice/group.
CP<0.01, significantly different from the EAC group.
dP<0.05 and DP<0.01, significantly
different from the yeast-treated EAC group. EP<0.01,
significantly different from the paclitaxel-H-treated EAC group.
FP<0.01, significantly different from the
paclitaxel-L-treated EAC group. GP<0.01,
significantly different from the yeast + paclitaxel-H-treated EAC
group. EAC, Ehrlich ascites carcinoma. Groups: EAC, mice bearing
tumors receiving intratumoral (i.t.) injections of PBS; Yeast, mice
bearing tumors receiving i.t. injections of yeast (1×107
cells/ml); paclitaxel-H, mice bearing tumors receiving paclitaxel
at a high dose (10 mg/kg); paclitaxel-L, mice bearing tumors
receiving paclitaxel at a low dose (2 mg/kg); Yeast + Paclitaxel-H,
mice bearing tumors receiving yeast plus paclitaxel-H; Yeast +
Paclitaxel-L, mice bearing tumors receiving yeast plus
paclitaxel-L. |
Flow cytometric analysis of cell
proliferation, DNA damage and apoptosis of EAC
Determination of cell
proliferation
Fig. 3 shows the
suppressive effect of yeast and/or paclitaxel on the percentage of
tumor cell proliferation as compared to EAC: treatment with yeast
alone showed −45% (P<0.01); treatment with paclitaxel-L alone
and paclitaxel-H alone was −35% (P<0.05) and −42% (P<0.01),
respectively; and combined treatments with yeast plus paclitaxel-L
and yeast plus paclitaxel-H showed −56 and −68% (P<0.01),
respectively. Thus, it appears that the combined treatment with
yeast and paclitaxel-L resulted in a larger decrease in cell
proliferation as compared to paclitaxel-H alone.
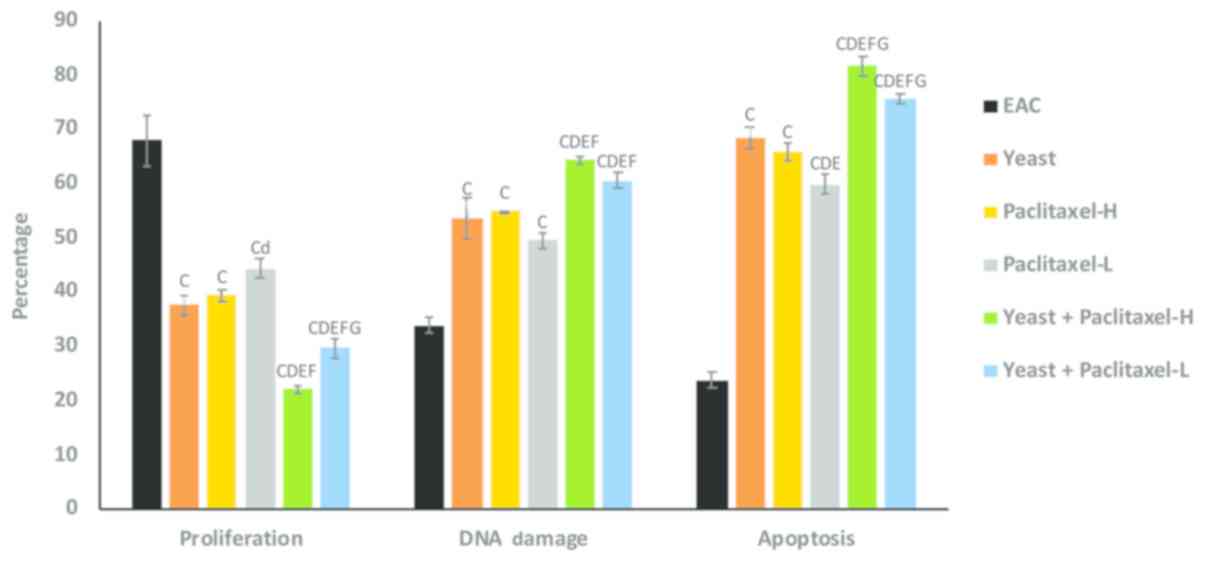 | Figure 3.Effect of yeast and/or paclitaxel on
the percentage of cell proliferation, DNA damage and apoptosis in
tumor tissues of the different groups. Data are expressed as mean ±
SE (5 mice/group). CP<0.01, significantly different
from the EAC group. dP<0.05 and
DP<0.01, significantly different from the
yeast-treated EAC group. EP<0.01, significantly
different from the paclitaxel-H-treated EAC group.
FP<0.01, significantly different from the
paclitaxel-- treated EAC group. GP<0.01,
significantly different from the yeast + paclitaxel-H-treated EAC
group. EAC, Ehrlich ascites carcinoma. Groups: EAC, mice bearing
tumors receiving intratumoral (i.t.) injections of PBS; Yeast, mice
bearing tumors receiving i.t. injections of yeast (1×107
cells/ml); paclitaxel-H, mice bearing tumors receiving paclitaxel
at a high dose (10 mg/kg); paclitaxel-L, mice bearing tumors
receiving paclitaxel at a low dose (2 mg/kg); Yeast + Paclitaxel-H,
mice bearing tumors receiving yeast plus paclitaxel-H; Yeast +
Paclitaxel-L, mice bearing tumors receiving yeast plus
paclitaxel-L. |
Determination of DNA damage
As shown in Fig. 3,
individual treatments of yeast, paclitaxel-L and paclitaxel-H
caused significant increase in the percent change in DNA damage. In
addition, combined treatments with yeast plus paclitaxel-L and
yeast plus paclitaxel-H showed even greater damage. Again, it
appears that the combined treatment with yeast and paclitaxel-L
resulted in a greater increase in DNA damage as compared to
paclitaxel-H alone.
Determination of apoptosis
Data in Fig. 3
showed that the individual treatments of yeast, paclitaxel-L and
paclitaxel-H caused a significant increase in tumor cell apoptosis.
In addition, combined treatments with yeast plus paclitaxel-L and
yeast plus paclitaxel-H showed higher increase in tumor cell
apoptosis. Results again showed that treatment with yeast plus
paclitaxel-L resulted in a larger increase in apoptosis as compared
to paclitaxel-H alone.
Quantitative determination of
apoptosis by Annexin V/PI double staining
A quantitative analysis of apoptosis in tumor cells
induced by yeast and/or paclitaxel was performed by flow cytometry
using Annexin V/PI double staining. Fig. 4 summarizes the results of early,
late and total apoptosis post-treatment with yeast in the presence
and absence of paclitaxel. The percent change in early apoptotic
cell count in the different groups, as compared to the control
untreated EAC-bearing mice, was as follows: treatment of yeast
alone showed 179%; treatment with paclitaxel-L alone and
paclitaxel-H alone was 60% (P<0.01) and 326% (P<0.01),
respectively; and treatment with yeast plus paclitaxel-L and yeast
plus paclitaxel-H showed 471 and 582% (P<0.01), respectively.
Combined treatment with yeast and paclitaxel-L resulted in an
increase of 471% in early apoptosis, as compared to the 326%
increase achieved by a paclitaxel-H alone. A similar trend of
results was noted in the percent change in late and total apoptotic
cell count in the different groups as compared to control untreated
EAC-bearing mice.
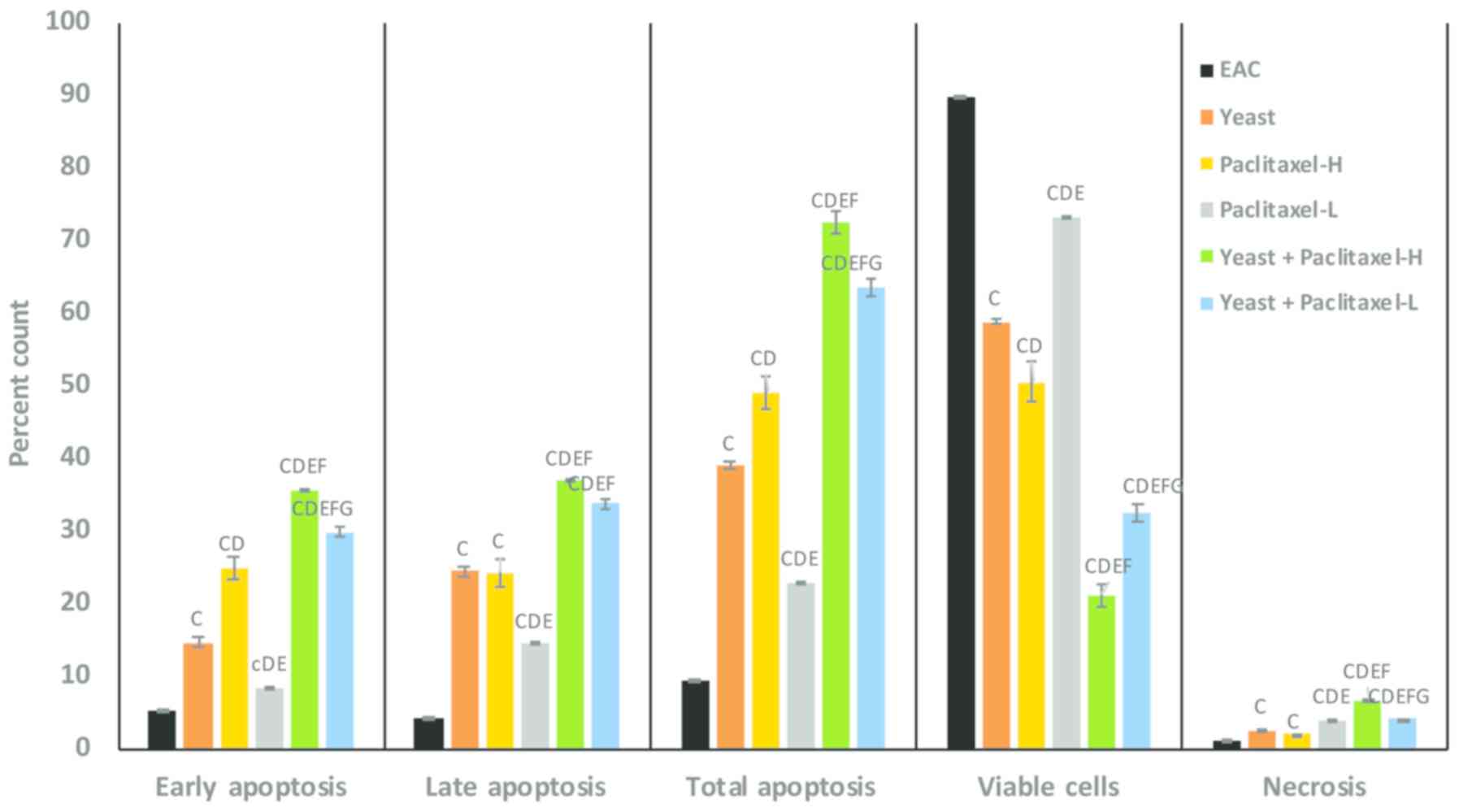 | Figure 4.Effect of paclitaxel and/or yeast
treatment on the Annexin V/PI labeling in tumor tissues of the
different groups. Data are expressed as mean ± SE (5 mice/group).
cP<0.05 and CP<0.01, significantly
different from the EAC group. DP<0.01, significantly
different from the yeast-treated EAC group. EP<0.01,
significantly different from the paclitaxel-H-treated EAC group.
FP<0.01, significantly different from the
paclitaxel-L-treated EAC group. GP<0.01,
significantly different from the yeast + paclitaxel-H-treated EAC
group. EAC, Ehrlich ascites carcinoma. Groups: EAC, mice bearing
tumors receiving intratumoral (i.t.) injections of PBS; Yeast, mice
bearing tumors receiving i.t. injections of yeast (1×107
cells/ml); paclitaxel-H, mice bearing tumors receiving paclitaxel
at a high dose (10 mg/kg); paclitaxel-L, mice bearing tumors
receiving paclitaxel at a low dose (2 mg/kg); Yeast + Paclitaxel-H,
mice bearing tumors receiving yeast plus paclitaxel-H; Yeast +
Paclitaxel-L, mice bearing tumors receiving yeast plus
paclitaxel-L. |
Furthermore, data in Fig. 4 shows the percentage of viable tumor
cells in mice under different treatment conditions as compared to
control untreated EAC-bearing mice. Mice treated with a combination
of yeast and paclitaxel-L resulted in a decrease in the percentage
of viable tumor cells by −64% (P<0.01) while paclitaxel-H alone
showed a decrease of −44% (P<0.01). In addition, the percent
change in necrotic cells showed that combined treatment with yeast
and paclitaxel-L resulted in an increase of 323% (P<0.01) in
tumor cell necrosis, as compared to a 110% (P<0.01) increase
achieved by paclitaxel-H alone.
Evaluation of Ki-67 cell proliferation
marker
Cell proliferation in EAC cells treated with yeast
with or without paclitaxel was determined by Ki-67 marker
immunohistochemical staining (Table
I). Control untreated EAC bearing mice showed an increase in
Ki-67-positive nuclei of the neoplastic cells. The percent decrease
in cell proliferation in the different groups, as compared to the
EAC-bearing mice, was as follows: treatment of yeast alone showed
50%, EAC-bearing animals treated with paclitaxel-H alone and
paclitaxel-L alone was 40% (P<0.01) and 36% (P<0.01),
respectively; and animals treated with yeast plus paclitaxel-H and
yeast plus paclitaxel-L showed 96 and 95% (P<0.01),
respectively.
 | Table I.Immunohistochemical examination of
the inhibitory effect of yeast and or paclitaxel on the percentage
of proliferation of EAC tumor cells in the different groups using
the Ki-67 marker. |
Table I.
Immunohistochemical examination of
the inhibitory effect of yeast and or paclitaxel on the percentage
of proliferation of EAC tumor cells in the different groups using
the Ki-67 marker.
| Groups | Mean ± SE | % of change from
EAC |
|---|
| EAC | 18.2±0.17 | – |
| Yeast |
9.1±0.31a | 50 |
| Paclitaxel-H |
10.9±0.09a | 40 |
| Paclitaxel-L |
11.6±0.26a | 36 |
| Yeast +
Paclitaxel-H |
0.7±0.058a | 96 |
| Yeast +
Paclitaxel-L |
0.9±0.053a | 95 |
Detection of CD4+ and
CD8+ T cells infiltrating the tumor by flow cytometric
analysis
Infiltrated CD4+ T
cells
Table II shows that
the percent increase in infiltrated CD4+ T cells in the
different groups, as compared to EAC-bearing mice, is as follows:
treatment of yeast alone 57%, paclitaxel-H alone 49% and
paclitaxel-L alone 38%. In addition animals treated with yeast plus
paclitaxel-H and yeast plus paclitaxel-L showed 72 and 54%,
respectively.
| Table II.Effect of yeast and/or paclitaxel
treatment on T-helper lymphocytes (CD4+) infiltrated and
cytotoxic lymphocytes (CD8+) infiltrated in tumor
tissue. |
Table II.
Effect of yeast and/or paclitaxel
treatment on T-helper lymphocytes (CD4+) infiltrated and
cytotoxic lymphocytes (CD8+) infiltrated in tumor
tissue.
|
| Parameters |
|---|
|
|
|
|---|
| Groups | % CD4+ T
cells | % change from EAC
group | % CD8+ T
cells | % change from EAC
group |
|---|
| EAC | 28.11±0.27 |
| 11.48±0.45 |
|
| Yeast |
44.10±1.4C | 56.9 |
20.05±0.29C | 74.6 |
| Paclitaxel-H |
41.76±0.45C | 48.6 |
16.17±0.66C,D | 40.8 |
| Paclitaxel-L |
38.76±1.1C,D,e | 37.9 |
13.18±0.39D,E | 14.8 |
| Yeast +
Paclitaxel-H |
48.35±0.76C,E,F | 72.0 |
22.20±0.36C,d,E,F | 93.3 |
| Yeast +
Paclitaxel-L |
43.16±0.97C,F,G | 53.6 |
18.55±0.92C,D,e,F,G | 61.5 |
Infiltrated CD8+ T
cells
Table II also shows
that the percent increase in infiltrated CD8+ T cells in
the different groups, as compared to EAC-bearing mice, is as
follows: treatment of yeast alone showed 75%, paclitaxel-H alone
41% and paclitaxel-L alone 15%. Data also showed that animals
treated with yeast plus paclitaxel-H and yeast plus paclitaxel-L
showed 93 and 62%, respectively.
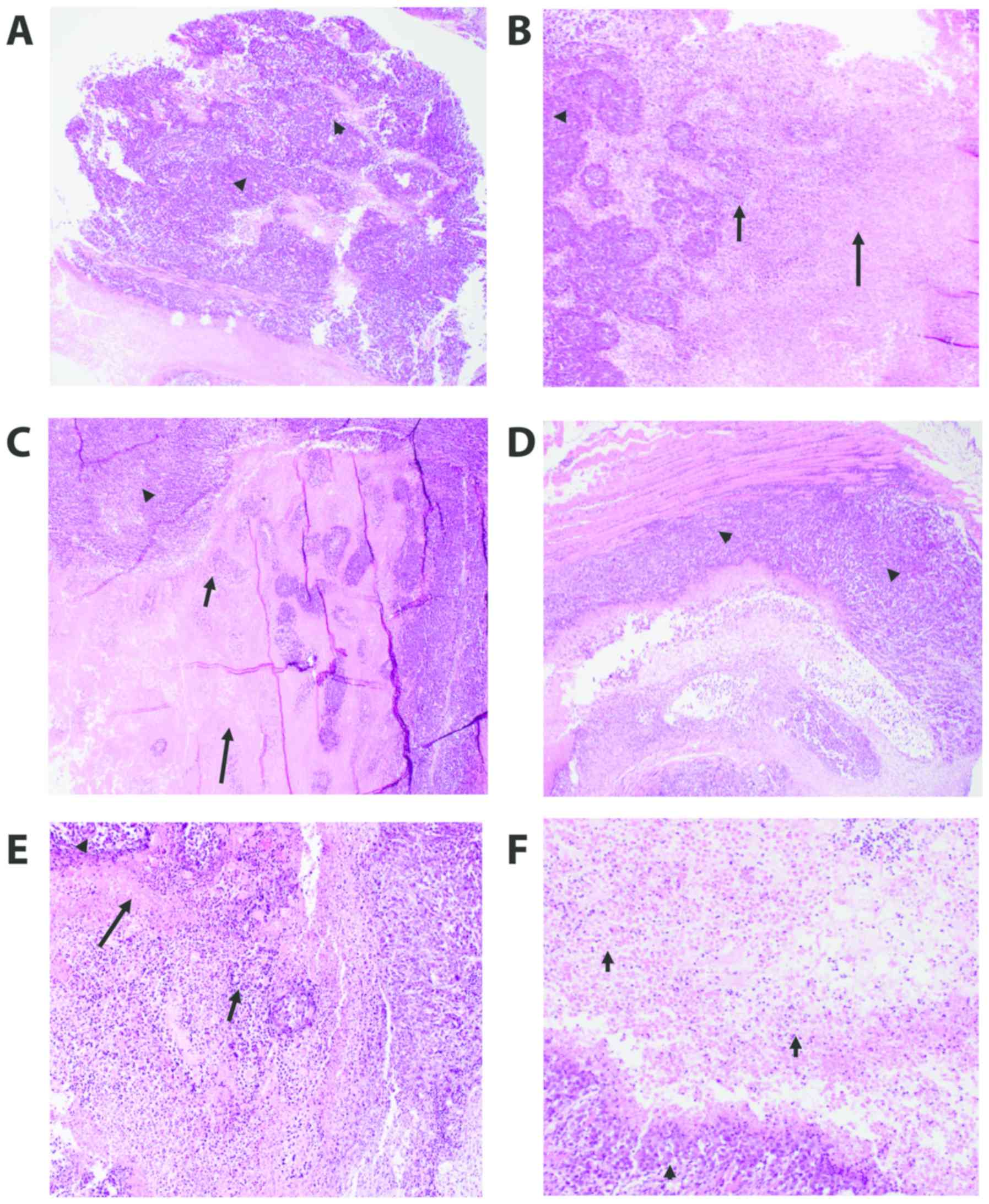
Histopathology of tumor tissues
We examined H&E staining of the solid tumors
removed from the EAC-bearing mice under different treatment
conditions using light microscopy. The different groups include the
control untreated EAC-bearing mice (A), EAC-bearing mice injected
with yeast (B), EAC-bearing mice treated with paclitaxel-H (C),
EAC-bearing mice treated with paclitaxel-L (D), EAC-bearing mice
treated with yeast plus paclitaxel-H (E), and EAC-bearing mice
treated with yeast plus paclitaxel-L (F). Results of the treatments
are illustrated in Table III and
Fig. 5A-F.
| Table III.Histopathological analysis of the
tumors from mice under the different treatment conditions. |
Table III.
Histopathological analysis of the
tumors from mice under the different treatment conditions.
|
| Treatments |
|---|
|
|
|
|---|
| Parameters | EAC (A) | Yeast (B) | Paclitaxel-H
(C) | Paclitaxel-L
(D) | Yeast +
Paclitaxel-H (E) | Yeast +
Paclitaxel-L (F) |
|---|
| Sheets of highly
undifferentiated pleomorphic cells, % | 65% | 50% | 30% | 60% | 30% | 30% |
| Extent of
necrosis | 35% | 50% | 70% | 40% | 70% | 70% |
| a. Extent of
coagulative necrosis, % | 30% | 40% | 60% | 20% | 40% | 50% |
| b. Extent of
colliquative necrosis, % | 5% | 10% | 10% | 20% | 30% | 20% |
| Total number of
bizarre/multinucleated cells among the total number of HPFs per
slide | 55/5=11 | 66/22=3 | 392/49=8 | 408/68=6 | 255/51=5 | 244/61=4 |
| Total number of
mitoses per slide in high power fields (HPF or ×40) | 52/14=3.7 | 51/29=1.8 | 152/49=3.1 | 244/74=3.3 | 169/51=3.3 | 108/61=1.8 |
| Average number of
mitoses per HPF | 4 | 2 | 3 | 3 | 3 | 2 |
H&E-stained sections of the tumor in all groups
(Fig. 5A-F) showed poorly
differentiated invasive carcinoma. The neoplastic cells were large,
had a high N:C ratio and exhibited markedly pleomorphic nuclei.
Many mitoses including abnormal forms were noted. All tissue
sections of the tumor in all groups displayed tumor necrosis that
mostly occurred at the center of the tumor nodule. The necrotic
tumor appeared acellular, stained homogenously with red eosin,
retained the general architectural pattern of the tissue and was
found to be characterized by abrupt transition from viable to
necrotic tissue (coagulative necrosis). This necrotic area was
marginated by degenerated tumor cells and inflammatory cells
(colliquative necrosis).
The effects of the different treatments were
evaluated via testing the following parameters: size of the tumor,
extent of necrosis, number of bizarre/multinucleated tumor cells,
and the degree of proliferative activity of the tumor (number of
mitoses). Group A, the control untreated tumor, was used to compare
all the other groups. Injecting yeast to the tumor (B) caused an
appreciable tumor necrosis as compared with group A. Tumor necrosis
was found to be highest and comparable between groups
paclitaxel-high (C), yeast + paclitaxel-high (E) and yeast +
paclitaxel-low (F). The amount of tumor necrosis in paclitaxel-low
(D) was almost comparable with the control group (A). The tumor
aggressiveness decreased in all treated groups (B-F) but was least
in groups B and F as evidenced by the lowest number of bizarre
tumor cells and the number of mitoses.
In summary, histopathological examination revealed
that the addition of yeast to low dose paclitaxel (group F) caused
the greatest tumor necrosis and exhibited the least aggressive
features (the lowest number of bizarre tumor cells and the lowest
proliferative activity).
Ultrastructural observations of tumor
tissues
The ultrastructural examination of the tumor samples
excised from EAC-bearing mice treated with yeast in the absence or
presence of paclitaxel is illustrated in Fig. 6A-H. (A) Control untreated
EAC-bearing mice showed tumor cells with large nucleus or
binucleated with scattered clumps of heterochromatin across the
nucleoplasm and condensed around the nuclear membrane. Cells also
displayed scant rough endoplasmic reticulum (RER) and mitochondria
(M). (B and C) Treatment with yeast alone showed neoplastic cells
engulfing countless numbers of yeast (B). This triggered apoptosis
of cancer cells, as indicated by fragmented and condensed chromatin
inside nuclei, extensive vacuolation (V), and marked loss of
mitochondria (M) and RER (C). (D) Mice treated with paclitaxel-H
alone showed highly degenerated apoptotic cancer cells with
chromatin condensation and aggregation around the nuclear envelope,
extensive vacuolation, destroyed RER, damaged mitochondria and
numerous cytoplasmic debris. (E) Tumor tissues treated with
paclitaxel-L alone showed apoptotic cancer cells with marginated or
degraded chromatin materials in the nucleus, and the cytoplasm
contained plenty of electron-dense grainy free ribosomes. (F) Mice
treated with yeast plus paclitaxel-H showed apoptotic cancer cells
with condensation of chromatin material that were marginated around
the nuclear membrane, membrane blebbing, yeast, extensive
vacuolation, an increase in the degenerative area with cellular
debris, fibrous tissue and marked increase in RER degeneration and
mitochondrial destruction. (G and H) Yeast plus paclitaxel-L group,
showing apoptotic cells with fragmented nuclei, several yeast
uptaken by the cell, extensive vacuolation, and necrosis. An
increase in cellular debris and fibers in the inter-neoplastic
stroma, phagosomes and mitochondria with destructed cristae and
electron lucent matrix were observed.
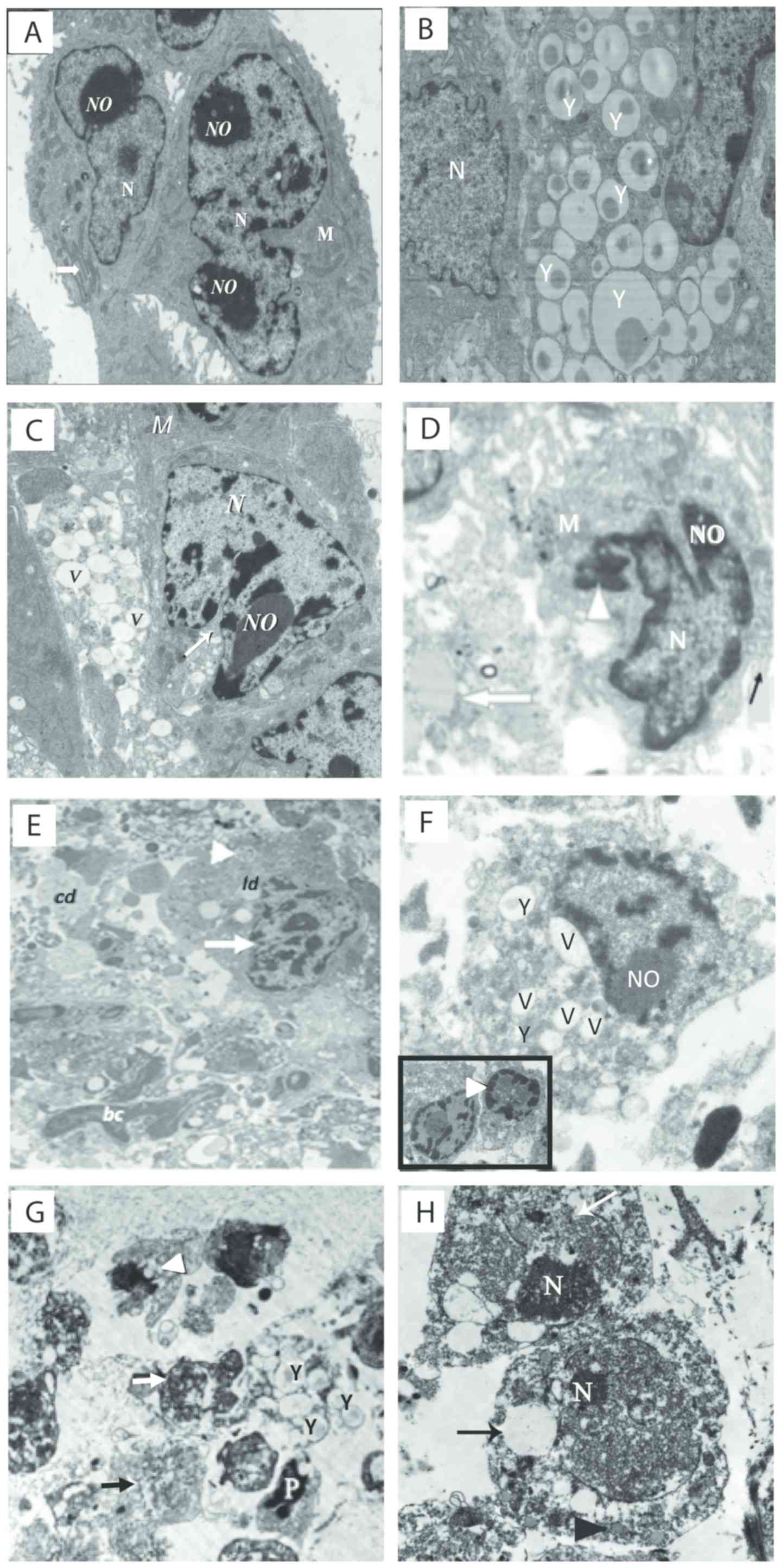 | Figure 6.(A-H) Electron micrographs of tumor
tissues from the Ehrlich ascites carcinoma (EAC)-bearing mice under
different treatments. (A) Untreated tumor tissues showing
neoplastic cells with large nucleus or binucleated (N) with defined
nucleolus (NO), mitochondria (M), and prominent RER (white arrow)
(×3,000). (B and C) Yeast-treated group. (B) Image showing
neoplastic cells engulfing massive amounts of yeast (Y). (C) Image
showing tumor cells with severe vacuolations (V), and segregation
and condensation of chromatin (×2,500). (D) Paclitaxel-H-treated
group showing condensation of chromatin (arrowhead) and around the
nuclear membrane, cell membrane blebbing (black arrow),
vacuolation, and muscle fiber (white arrow) (×4,000). (E)
Paclitaxel-L-treated group showing apoptotic cell (arrow head),
condensation of chromatin and identification of nucleus (white
arrow), cellular debris (cd), phagosome (white arrowhead), lipid
droplets (ld), and blood capillary (bc) (1,500). (F) Yeast +
paclitaxel-H-treated group showing neoplastic cells engulfing yeast
inside them (Y) and extensive vacuolation (V); the inset image
shows two apoptotic cells with fragmented nucleus and condensation
of chromatin material around the nuclear membrane (arrow head)
(×4,000). (G and H) Yeast + paclitaxel-L-treated group. (G) Image
showing apoptotic neoplastic cells (white arrow) with multiple
yeasts (Y), apoptotic cell (P), extensive cytoplasmic vacuolation
(black arrow) and marked loss of mitochondria (arrow head). (H)
Image showing two neoplastic cells with predominance of
heterochromatin in their nuclei (N), disruption in the nuclear
membrane (white arrow) and large electron lucent cytoplasmic
vacuoles (black arrow) (magnification, ×1,500 for G and ×2,000 for
H). Scale bar, 500 nm. Groups: EAC, mice bearing tumors receiving
intratumoral (i.t.) injections of PBS; Yeast, mice bearing tumors
receiving intratumoral (i.t.) injections of yeast (1×107
cells/ml); paclitaxel-H, mice bearing tumors receiving paclitaxel
at a high dose (10 mg/kg); paclitaxel-L, mice bearing tumors
receiving paclitaxel at a low dose (2 mg/kg); Yeast + Paclitaxel-H,
mice bearing tumors receiving yeast plus paclitaxel-H; Yeast +
Paclitaxel-L, mice bearing tumors receiving yeast plus
paclitaxel-L |
Discussion
Paclitaxel is a frequently used chemotherapeutic
agent that was originally isolated from the bark of the Pacific yew
tree and was approved in 1994 by the US Food and Drug
Administration (FDA) as Taxol® for the treatment of
patients with metastatic breast cancer. In addition, paclitaxel
exhibits a broad spectrum of activity against other human cancers,
such as ovarian, non-small cell lung cancers and malignant brain
tumors (41). Although paclitaxel
induces apoptotic effects on cancer cells at high concentrations
(42–45), its treatment is associated with
severe side-effects, including myelosuppression, peripheral
neuropathy, hypersensitivity reactions, alopecia and cardiac
disturbances and gastrointestinal toxicity (1–3,46).
Therefore, our research focused on finding dietary agents that may
have the ability to reduce the toxicity of chemotherapy by lowering
the concentration of drug administered to animals and patients
while maintaining potency against cancer cells. Our previous
research revealed that arabinoxylan rice bran, Biobran/MGN-3, has
the ability to sensitize cancer cells to paclitaxel in
animal-bearing tumors (47). In
addition, Biobran/MGN-3 was found to sensitize hepatocellular
carcinoma to chemotherapy and other interventional therapies in a
3-year randomized clinical trial (19). We recently investigated the ability
of another dietary agent, bakers and brewers yeast, S.
cerevisiae, to sensitize metastatic and non-metastatic breast
cancer cells to paclitaxel in vitro and showed the
IC50 value for paclitaxel was significantly reduced in
the presence of yeast (34).
In the present study, we evaluated the ability of
bakers yeast to sensitize cancer cells to paclitaxel in mice
bearing EAC. Here, we present evidence showing the ability of
bakers yeast to act as a chemosensitizer when used in combination
with paclitaxel. Co-treatment of yeast plus paclitaxel at a low
dose (2 mg/kg BW, paclitaxel-L) resulted in greater anticancer
effects than paclitaxel at a high dose (10 mg/kg BW, paclitaxel-H)
alone. This is clearly evidenced by a greater retardation of tumor
growth, greater increase in DNA damage and apoptosis and larger
decrease in cell proliferation as compared to paclitaxel-H alone.
In addition, histopathological examination confirmed that yeast
plus paclitaxel-L produced antitumor effects better than
paclitaxel-H alone. Thus, bakers yeast may be used in conjunction
with chemotherapy at low concentrations to achieve the same potency
as high-dose chemotherapy against cancer cells in mice-bearing
EAC.
Apoptosis is an important factor underlying the
anticancer effects by yeast and paclitaxel. Our previous studies
demonstrated that heat-killed bakers yeast is a potent inducer of
apoptosis. This is based on the phenomenon that phagocytosis of
yeast by cancer cells triggers apoptosis of cancer cells. Several
in vitro studies have demonstrated that human cancer cells
can phagocytize yeast, and subsequently these cancer cells showed
morphological signs of apoptosis such as nuclear fragmentation and
membrane blebbing, in addition, flow cytometric analysis showed
mitochondrial polarization and increased activation of caspases-8,
−9 and −3 in the BCC post culture with yeast (26,27,29).
Furthermore, the current ultra-structural analysis showed
neoplastic cells engulfing countless numbers of yeast, which
triggered the apoptosis of cancer cells as indicated by fragmented
and condensed chromatin inside nuclei and extensive vacuolized
cytoplasm. This yeast-induced apoptosis of tumor cells ultimately
resulted in significant retardation of tumor growth. These results
are in accordance with our previous studies that showed yeast i.t.
injection suppressed the growth of mice-bearing EAC tumors
(32) and nude mice-bearing human
MCF-7 tumor cells. In these studies, treatments with yeast showed
active involvement of lysosomes in the phagocytosis and digestion
of yeast and in cancer cell apoptosis (31). Further studies showed that
yeast-induced apoptosis of human MBC cells occurs by a mechanism
involving intracellular Ca2+ that may trigger apoptotic
signals directly via intrinsic pathway of apoptosis (48).
As a member of the taxane family, paclitaxel
suppresses spindle microtubule dynamics. Inhibition of microtubules
results in the blockage of metaphase-anaphase transitions,
ultimately suppressing cell proliferation by inhibition of mitosis
(49,50). In the present study, treatment with
paclitaxel inhibited cell proliferation and induced DNA damage in
mice-bearing EAC tumors in a dose-dependent manner. Our research
and that of others showed that paclitaxel induces apoptosis in a
variety of cancer cells including BCCs by both intrinsic and
extrinsic pathways of apoptosis, involving modification of
mitochondrial membrane potential (MMP) and activation of caspase-8
and caspase-3 (51–55). Ultra-structural examination showed
that treatment with paclitaxel induced cancer cell apoptosis that
was associated with chromatin condensation and aggregation around
the nuclear envelope and the cytoplasm contained numerous
electron-dense grainy free ribosomes. The apoptotic effect by
paclitaxel is attributed to the ability of this agent to arrest
mitosis through microtubule stabilization.
In the present study, the ability of yeast to
sensitize EAC to paclitaxel was well illustrated. Fig. 3 shows that cotreatment of yeast plus
paclitaxel-L caused a greater decrease in cell proliferation than
paclitaxel-H alone (−56 vs. −42%), as examined by BrdU staining and
weak expression of KI-67 (a marker of cell proliferation) (95 vs.
40%), elevated DNA damage (+79 vs. +62%), and an enhancement of the
apoptotic tumor cells (+217 vs. +177%). These results are in accord
with our recent in vitro studies that showed that bakers yeast in
the presence of paclitaxel increased the sensitivity of three human
and murine BCC lines to paclitaxel (34).
Several dietary products have shown the ability to
modulate multi-drug resistance (MDR) and thus have potential as
chemosensitizers. For example, the flavonoid quercetin is an MDR
modulator (25), curcumin
sensitizes cervical cancer cells to cisplatin by suppressing MRP1
and Pgp1 (56) and vitamin E acts
as a P-gp inhibitor (22). Dietary
fatty acids also act as chemosensitizers by increasing the
intracellular chemotherapy drug accumulation in cancer cells
(57,58), and tetrandrine is likewise able to
restore drug sensitivity (6,23,24).
Flow cytometry results from the present study showed yeast to be a
chemosensitizer that possesses potential for adjuvant therapy in
the treatment of cancer, as exemplified by the fact that yeast in
the presence of paclitaxel-L enhances apoptosis in EAC to a greater
extent than paclitaxel-H alone. The mechanism(s) underlying the
chemosensitizing effect of yeast is not known, but could be
attributed to the ability of yeast to suppress MDR proteins or
increase the intracellular chemotherapy drug accumulation in cancer
cells.
The immunomodulatory effect of S. cerevisiae
may represent another mechanism by which this agent suppresses
tumor growth in EAC-bearing mice. Results of this study showed the
ability of yeast to act as a potent immune modulator, as indicated
by enhancement of CD4+ and CD8+ T cell tumor
infiltration. Earlier research has shown that yeast treatment can
increase recruitment of leukocytes, including macrophages into the
tumors, and modulate cytokine response, as indicated by elevation
of TNF-α and IFN-γ plasma levels and decreased IL-10 levels
(32). This suggests that S.
cerevisiae exerts anticancer effects via both apoptotic and
immunomodulatory properties. The findings of this study also showed
that paclitaxel enhanced tumor infiltration of CD4+ and
CD8+ T cells but to a lesser extent than yeast.
The histopathological results of yeast treatment
alone are in agreement with our previous results in which injection
of S. cerevisiae into mice bearing EAC was associated with
histopathology changes including extensive tumor necrosis,
apoptosis and ischemic (coagulative) and liquefactive necrosis
(32). In the present study, yeast
in combination with paclitaxel-L resulted in increased tumor
necrosis, decreased tumor aggressiveness and decreased tumor
proliferative activity relative to treatment with either yeast or
paclitaxel-L alone. Yeast plus paclitaxel-L showed a similar
magnitude of tumor necrosis as compared to paclitaxel-H alone and
yeast plus paclitaxel-H. Yeast plus paclitaxel-L also demonstrated
fewer bizarre/multinucleated cells and mitoses than paclitaxel-H.
Thus, yeast plus paclitaxel-L is more effective against breast
cancer than paclitaxel-H.
S. cerevisiae is viewed as a safe, non-toxic,
non-life threatening agent by the human population. In our previous
in vitro study (28), we
showed that yeast specifically affects cancer cells and not normal
cells. When cultured with yeast, breast cancer cells (MCF-7)
phagocytized the yeast and subsequently underwent apoptosis, while
there was virtually no phagocytosis of yeast by the normal
non-tumorgenic breast epithelial cells (MCF-10A). The safety of
yeast treatment has been additionally examined using two in
vivo approaches. In animal studies, S. cerevisiae showed
no toxic characteristics as manifested by the ability of all
animals to survive for 45 days post-treatment with yeast with
normal feeding/drinking and life activity patterns (31). Mice treated with yeast survived a
16-week treatment period and histopathology of different organs and
biochemical analysis of liver and kidney function were within the
limits of normal healthy mice (59). The work of others have also shown
that animals can tolerate relatively high doses of yeast without
any pathology detected up to at least 21 days post treatment,
including rats (60), mice
(61) and monkeys (62). Our current data are in accordance
with the above mentioned studies, where mice survived a 30-day
treatment period. In the second in vivo approach, human
studies have shown that i.v. injections of yeast glucans have been
given to humans to boost the immune system of patients undergoing
major surgery (63–64), patients with
Paracoccidioidomycosis (PCM), an endemic disease in most
Latin American countries, and for elimination of P.
brasiliensis (65). Results of
these studies revealed no adverse side effects associated with
yeast infusion (63,64,66). A
skin cancer study showed that treatment with yeast counteracted the
significant decrease in body weight due to cancer (59). Weight loss is common among people
with cancer, and it has been also observed in animal studies that
adipose tissue wasting can occur as soon as the tumor is palpable
(67).
In conclusion, bakers yeast can enhance the
apoptotic effect of paclitaxel and may suggest the use of yeast as
an adjuvant treatment during anticancer chemotherapy. These results
may have clinical implications for the treatment of breast cancer.
Future research needs to be directed towards examining the
chemosensitizing effect of yeast in other models, such as the
xenograft model. In addition, the ability of yeast as a
chemosensitizer should be investigated in combination with other
chemotherapeutic agents.
Acknowledgements
Not applicable.
Funding
The present study was partially supported by the
National Institutes of Health-National Institute on Minority Health
and Health Disparities (grant nos. U54MD007598 and
S21MD000103).
Availability of data and materials
The datasets used and analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors contributions
MG and NKBED designed the experiments and analyzed
and interpreted data. AZM analyzed and interpreted the data. LT
carried out the histopathology analyses. DP contributed to the
statistical analysis of the data. TAH performed the experiments and
analyzed the data. All authors drafted, reviewed, edited, read and
approved the manuscript and agree to be accountable for all aspects
of the research in ensuring that the accuracy or integrity of any
part of the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
Animal protocols were in compliance with the Guide
for the Care and Use of Laboratory Animals at the University of
Mansoura, Egypt and the experiments were approved by the
University.
Patient consent for publication
Not applicable.
Competing interests
The authors declared that they have no competing
interests.
References
|
1
|
Gligorov J and Lotz JP: Preclinical
pharmacology of the taxanes: Implications of the differences.
Oncologist. 9 (Suppl 2):3–8. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Guastalla JP, Lhommé C, Dauplat J, Namer
M, Bonneterre J, Oberling F, Pouillart P, Fumoleau P, Kerbrat P,
Tubiana N, et al: Taxol (paclitaxel) safety in patients with
platinum pretreated ovarian carcinoma: An interim analysis of a
phase II multicenter study. Ann Oncol. 5 (Suppl 6):S33–S38.
1994.PubMed/NCBI
|
|
3
|
Ohtsu T, Sasaki Y, Tamura T, Miyata Y,
Nakanomyo H, Nishiwaki Y and Saijo N: Clinical pharmacokinetics and
pharmacodynamics of paclitaxel: A 3-hour infusion versus a 24-hour
infusion. Clin Cancer Res. 1:599–606. 1995.PubMed/NCBI
|
|
4
|
Newman DJ, Cragg GM and Snader KM: Natural
products as sources of new drugs over the period 1981–2002. J Nat
Prod. 66:1022–1037. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sylvester PW, Kaddoumi A, Nazzal S and El
Sayed KA: The value of tocotrienols in the prevention and treatment
of cancer. J Am Coll Nutr. 29 (Suppl):324S–333S. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen YJ: Potential role of tetrandrine in
cancer therapy. Acta Pharmacol Sin. 23:1102–1106. 2002.PubMed/NCBI
|
|
7
|
Farina HG, Pomies M, Alonso DF and Gomez
DE: Antitumor and antiangiogenic activity of soy isoflavone
genistein in mouse models of melanoma and breast cancer. Oncol Rep.
16:885–891. 2006.PubMed/NCBI
|
|
8
|
Rafieian-Kopaei M and Movahedi M: Breast
cancer chemopreventive and chemotherapeutic effects of Camellia
Sinensis (green tea): An updated review. Electron Physician.
9:3838–3844. 2017. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Badr El-Din NK, Noaman E and Ghoneum M: In
vivo tumor inhibitory effects of nutritional rice bran supplement
MGN-3/Biobran on Ehrlich carcinoma-bearing mice. Nutr Cancer.
60:235–244. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Badr El-Din NK, Abdel Fattah SM, Pan D,
Tolentino L and Ghoneum M: Chemopreventive activity of
MGN-3/Biobran against chemical induction of glandular stomach
carcinogenesis in rats and its apoptotic effect in gastric cancer
cells. Integr Cancer Ther. 15:NP26–NP34. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Pérez-Martínez A, Valentín J, Fernández L,
Hernández-Jiménez E, López-Collazo E, Zerbes P, Schwörer E, Nuñéz
F, Martín IG, Sallis H, et al: Arabinoxylan rice bran
(MGN-3/Biobran) enhances natural killer cell-mediated cytotoxicity
against neuroblastoma in vitro and in vivo. Cytotherapy.
17:601–612. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ghoneum M and Gollapudi S: Susceptibility
of the human LNCaP prostate cancer cells to the apoptotic effect of
marina crystal minerals (MCM) in vitro. Oncol Rep. 22:155–159.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ghoneum M, Seto Y, Sato S, Ghoneum A,
Braga M and Gollapudi S: Gross thymic extract, Thymax, induces
apoptosis in human breast cancer cells in vitro through the
mitochondrial pathway. Anticancer Res 28 (3A). 1603–1609. 2008.
|
|
14
|
Ghoneum M and Gimzewski J: Apoptotic
effect of a novel kefir product, PFT, on multidrug-resistant
myeloid leukemia cells via a hole-piercing mechanism. Int J Oncol.
44:830–837. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gollapudi S and Ghoneum M: MGN-3/Biobran,
modified arabinoxylan from rice bran, sensitizes human breast
cancer cells to chemotherapeutic agent, daunorubicin. Cancer Detect
Prev. 32:1–6. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ghoneum M: Wheat and rice in disease
prevention and health. In: Apoptosis and Arabinoxylan Rice Bran.
Watson RR, Preedy V and Zibadi S: Elsevier Science & Technology
Book; pp. 399–404. 2014
|
|
17
|
Endo Y and Kanbayashi H: Modified rice
bran beneficial for weight loss of mice as a major and acute
adverse effect of Cisplatin. Pharmacol Toxicol. 92:300–303. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jacoby HI, Wnorowski G, Sakata K and Maeda
H: The effect of MGN-3 on cisplatin and doxorubicin induced
toxicity in the rat. J. Nutraceuticals Funct Med Foods. 3:3–11.
2001. View Article : Google Scholar
|
|
19
|
Bang MH, Van Riep T, Thinh NT, Song H,
Dung TT, Van Truong L, Van Don L, Ky TD, Pan D, Shaheen M, et al:
Arabinoxylan rice bran (MGN-3) enhances the effects of
interventional therapies for the treatment of hepatocellular
carcinoma: A three-year randomized clinical trial. Anticancer Res.
30:5145–5151. 2010.PubMed/NCBI
|
|
20
|
Goel A and Aggarwal BB: Curcumin, the
golden spice from Indian saffron, is a chemosensitizer and
radiosensitizer for tumors and chemoprotector and radioprotector
for normal organs. Nutr Cancer. 62:919–930. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Limtrakul P: Curcumin as chemosensitizer.
Adv Exp Med Biol. 595:269–300. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rege BD, Kao JP and Polli JE: Effects of
nonionic surfactants on membrane transporters in Caco-2 cell
monolayers. Eur J Pharm Sci. 16:237–246. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tian H and Pan QC: A comparative study on
effect of two bisbenzylisoquinolines, tetrandrine and berbamine, on
reversal of multidrug resistance. Yao Xue Xue Bao. 32:245–250.
1997.(In Chinese). PubMed/NCBI
|
|
24
|
Choi SU, Park SH, Kim KH, Choi EJ, Kim S,
Park WK, Zhang YH, Kim HS, Jung NP and Lee CO: The
bisbenzylisoquinoline alkaloids, tetrandine and fangchinoline,
enhance the cytotoxicity of multidrug resistance-related drugs via
modulation of P-glycoprotein. Anticancer Drugs. 9:255–261. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen C, Zhou J and Ji C: Quercetin: A
potential drug to reverse multidrug resistance. Life Sci.
87:333–338. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ghoneum M and Gollapudi S: Induction of
apoptosis in breast cancer cells by Saccharomyces
cerevisiae, the bakers yeast, in vitro. Anticancer Res 24 (3a).
1455–1463. 2004.
|
|
27
|
Ghoneum M and Gollapudi S: Modified
arabinoxylan rice bran (MGN-3/Biobran) enhances yeast-induced
apoptosis in human breast cancer cells in vitro. Anticancer Res 25
(2A). 859–870. 2005.
|
|
28
|
Ghoneum M and Gollapudi S: Synergistic
role of arabinoxylan rice bran (MGN-3/Biobran) in S.
cerevisiae-induced apoptosis of monolayer breast cancer MCF-7
cells. Anticancer Res 25 (6B). 4187–4196. 2005.
|
|
29
|
Ghoneum M, Hamilton J, Brown J and
Gollapudi S: Human squamous cell carcinoma of the tongue and colon
undergoes apoptosis upon phagocytosis of Saccharomyces
cerevisiae, the bakers yeast, in vitro. Anticancer Res 25 (2A).
981–989. 2005.
|
|
30
|
Ghoneum M, Brown J and Gollapudi S: ‘Yeast
therapy for the treatment of cancer and its enhancement by
MGN-3/Biobran, an arabinoxylan rice bran’ in apoptosis review. Nova
Science Publishers Inc.; Hauppague, NY: pp. 185–200. 2007
|
|
31
|
Ghoneum M, Wang L, Agrawal S and Gollapudi
S: Yeast therapy for the treatment of breast cancer: A nude mice
model study. In Vivo. 21:251–258. 2007.PubMed/NCBI
|
|
32
|
Ghoneum M, Badr El-Din NK, Noaman E and
Tolentino L: Saccharomyces cerevisiae, the Bakers Yeast, suppresses
the growth of Ehrlich carcinoma-bearing mice. Cancer Immunol
Immunother. 57:581–592. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yarkoni E, Lederer E and Rapp HJ:
Immunotherapy of experimental cancer with a mixture of synthetic
muramyl dipeptide and trehalose dimycolate. Infect Immun.
32:273–276. 1981.PubMed/NCBI
|
|
34
|
Badr El-Din NK, Mahmoud AZ, Hassan TA and
Ghoneum M: Bakers yeast sensitizes metastatic breast cancer cells
to paclitaxel in vitro. Integr Cancer Ther. 17:542–550. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Calixto-Campos C, Zarpelon AC, Corrêa M,
Cardoso RD, Pinho-Ribeiro FA, Cecchini R, Moreira EG, Crespigio J,
Bernardy CC, Casagrande R, et al: The Ehrlich tumor induces
pain-like behavior in mice: A novel model of cancer pain for
pathophysiological studies and pharmacological screening. BioMed
Res Int. 2013:6248152013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ehrlich and Apolant. Beobachtungen Über
Maligne Mausentumoren, . Berlin: Klin Wochenschr; 28. pp. 871–874.
1905, (In German).
|
|
37
|
Kaleoğlu Ö and İşli N: Ehrlich -Lettre
Asit Tümörü. Tip Fak Mecm. 40:978–984. 1977.(In Turkish).
|
|
38
|
Mishra S, Tamta AK, Sarikhani M, Desingu
PA, Kizkekra SM, Pandit AS, Kumar S, Khan D, Raghavan SC and
Sundaresan NR: Subcutaneous Ehrlich ascites carcinoma mice model
for studying cancer-induced cardiomyopathy. Sci Rep. 8:55992018.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Cattoretti G, Becker MH, Key G, Duchrow M,
Schlüter C, Galle J and Gerdes J: Monoclonal antibodies against
recombinant parts of the Ki-67 antigen (MIB 1 and MIB 3) detect
proliferating cells in microwave-processed formalin-fixed paraffin
sections. J Pathol. 168:357–363. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Reynolds ES: The use of lead citrate at
high pH as an electron microscopy. J Cell Biol. 17:208–212. 1963.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Marupudi NI, Han JE, Li KW, Renard VM,
Tyler BM and Brem H: Paclitaxel: A review of adverse toxicities and
novel delivery strategies. Expert Opin Drug Saf. 6:609–621. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Gustafson DL, Long ME, Zirrolli JA, Duncan
MW, Holden SN, Pierson AS and Eckhardt SG: Analysis of docetaxel
pharmacokinetics in humans with the inclusion of later sampling
time-points afforded by the use of a sensitive tandem LCMS assay.
Cancer Chemother Pharmacol. 52:159–166. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Xu G, Pan J, Martin C and Yeung S-CJ:
Angiogenesis inhibition in the in vivo antineoplastic effect of
manumycin and paclitaxel against anaplastic thyroid carcinoma. J
Clin Endocrinol Metab. 86:1769–1777. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Stassi G, Todaro M, Zerilli M,
Ricci-Vitiani L, Di Liberto D, Patti M, Florena A, Di Gaudio F, Di
Gesù G and De Maria R: Thyroid cancer resistance to
chemotherapeutic drugs via autocrine production of interleukin-4
and interleukin-10. Cancer Res. 63:6784–6790. 2003.PubMed/NCBI
|
|
45
|
Piñeiro D, Martín ME, Guerra N, Salinas M
and González VM: Calpain inhibition stimulates caspase-dependent
apoptosis induced by taxol in NIH3T3 cells. Exp Cell Res.
313:369–379. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Potemski P and Płuzańska A:
Pharmacological action of paclitaxel. Pol Merkur Lekarski. 6:27–29.
1999.(In Polish). PubMed/NCBI
|
|
47
|
Badr El-Din NK, Ali DA, Alaa El-Dein M and
Ghoneum M: Enhancing the apoptotic effect of a low dose of
paclitaxel on tumor cells in mice by arabinoxylan rice bran
(MGN-3/Biobran). Nutr Cancer. 68:1010–1020. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Ghoneum M, Matsuura M, Braga M and
Gollapudi S: S. cerevisiae induces apoptosis in human
metastatic breast cancer cells by altering intracellular
Ca2+ and the ratio of Bax and Bcl-2. Int J Oncol.
33:533–539. 2008.PubMed/NCBI
|
|
49
|
Jordan MA and Wilson L: The use and action
of drugs in analyzing mitosis. Methods Cell Biol. 61:267–295. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Morse DL, Gray H, Payne CM and Gillies RJ:
Docetaxel induces cell death through mitotic catastrophe in human
breast cancer cells. Mol Cancer Ther. 4:1495–1504. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Ghoneum M, Badr El-Din NK, Ali DA and
El-Dein MA: Modified arabinoxylan from rice bran, MGN-3/biobran,
sensitizes metastatic breast cancer cells to paclitaxel in vitro.
Anticancer Res. 34:81–87. 2014.PubMed/NCBI
|
|
52
|
Yu XJ, Sun K, Tang XH, Zhou CJ, Sun H, Yan
Z, Fang L, Wu HW, Xie YK and Gu B: Harmine combined with paclitaxel
inhibits tumor proliferation and induces apoptosis through
down-regulation of cyclooxygenase-2 expression in gastric cancer.
Oncol Lett. 12:983–988. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Gonçalves A, Braguer D, Carles G, André N,
Prevôt C and Briand C: Caspase-8 activation independent of
CD95/CD95-L interaction during paclitaxel-induced apoptosis in
human colon cancer cells (HT29-D4). Biochem Pharmacol.
60:1579–1584. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Peng ZG, Liu DC, Yao YB, Feng XL, Huang X,
Tang YL, Yang J and Wang XX: Paclitaxel induces apoptosis in
leukemia cells through a JNK activation-dependent pathway. Genet
Mol Res. 15:150139042016. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Varbiro G, Veres B, Gallyas F Jr and
Sumegi B: Direct effect of Taxol on free radical formation and
mitochondrial permeability transition. Free Radic Biol Med.
31:548–558. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Roy M and Mukherjee S: Reversal of
resistance towards cisplatin by curcumin in cervical cancer cells.
Asian Pac J Cancer Prev. 15:1403–1410. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Abulrob AN, Mason M, Bryce R and Gumbleton
M: The effect of fatty acids and analogues upon intracellular
levels of doxorubicin in cells displaying P-glycoprotein mediated
multidrug resistance. J Drug Target. 8:247–256. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Gelsomino G, Corsetto PA, Campia I,
Montorfano G, Kopecka J, Castella B, Gazzano E, Ghigo D, Rizzo AM
and Riganti C: Omega 3 fatty acids chemosensitize multidrug
resistant colon cancer cells by down-regulating cholesterol
synthesis and altering detergent resistant membranes composition.
Mol Cancer. 12:1372013. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Elwakkad A, Ghoneum M, El-sawi M, Mohamed
SI, Gamal el Din AA, Pan D and Elqattan GM: Bakers yeast induces
apoptotic effects and histopathological changes on skin tumors in
mice. Cogent Med. 5:14376732018. View Article : Google Scholar
|
|
60
|
Fahrig R: Development of host-mediated
mutagenicity tests-yeast systems. II. Recovery of yeast cells out
of testes, liver, lung, and peritoneum of rats. Mutat Res.
31:381–394. 1975. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Frezza D, Zeiger E and Gupta BN: The
intrasanguineous host-mediated assay procedure distribution and
retention of yeast in the mouse. Mutat Res. 64:295–305. 1979.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Maejima K, Shimoda K, Morita C, Fujiwara T
and Kitamura T: Colonization and pathogenicity of Saccharomyces
cerevisiae, MC16, in mice and cynomolgus monkeys after oral and
intravenous administration. Jpn J Med Sci Biol. 33:271–276. 1980.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Babineau TJ, Marcello P, Swails W, Kenler
A, Bistrian B and Forse RA: Randomized phase I/II trial of a
macrophage-specific immunomodulator (PGG-glucan) in high-risk
surgical patients. Ann Surg. 220:601–609. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Babineau TJ, Hackford A, Kenler A,
Bistrian B, Forse RA, Fairchild PG, Heard S, Keroack M, Caushaj P
and Benotti P: A phase II multicenter, double-blind, randomized,
placebo-controlled study of three dosages of an immunomodulator
(PGG-glucan) in high-risk surgical patients. Arch Surg.
129:1204–1210. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Wanke B and Londero AT: Epidemiology and
paracoccidioidomycosis infection. Paracoccidioidomycosis. Franco M,
Lacaz CS, Restrepo-Moreno A and Del Negro G: CRC Press; Boca Raton:
pp. 109–120. 1994
|
|
66
|
Meira DA, Pereira PC, Marcondes-Machado J,
Mendes RP, Barraviera B, Pellegrino Júnior J, Rezkallah-Iwasso MT,
Peracoli MT, Castilho LM, Thomazini I, et al: The use of glucan as
immunostimulant in the treatment of paracoccidioidomycosis. Am J
Trop Med Hyg. 55:496–503. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Costa G and Holland JF: Effects of Krebs-2
carcinoma on the lipide metabolism of male Swiss mice. Cancer Res.
22:1081–1083. 1962.PubMed/NCBI
|