Introduction
Bone defects and non-union caused by various
diseases, such as trauma and tumors, are common orthopedic
disorders in the clinic. Treatment is limited by the finite nature
of autogenous bone materials; however, with the rise of tissue
engineering, this problem may be solved using new methods. Current
research (1–3) shows that bone morphogenetic protein
(BMP) belongs to the TGF-β superfamily, which plays a notable role
in inducing bone differentiation. Among its family members, BMP2 is
one of the strongest factors and the only growth factor that is
capable of singly inducing bone formation. HIF1 expression was
increased in low oxygen environments, and it could be combined with
the hypoxia response element of target genes, regulate gene
transcription of vascular endothelial growth factor (VEGF), and
play a crucial role in tissue angiogenesis (4–7). The
current study will build a recombinant adenovirus vector containing
a BMP2 and three mutant HIF1α (HIF1αmu)
(Ad-BMP2-IRES-HIF1αmu), to be transfected into rabbit
bone marrow mesenchymal stem cells (MSCs), and observe the mRNA and
protein expression of the two genes in the MSCs which will provide
support for gene therapy of clinical bone defects and bone
non-union.
Materials and methods
Primers
Primer sequences were designed according to the
sequence information of the target gene human BMP2 and the
HIF1αmu gene and the enzyme digestion site information
of the pIRES2-EGFP vector. NheI and BamHI sites were
respectively added at the 5′ end and the 3′ end of BMP2 primer, and
a 6xHIS tag was added at the C-cleavage site of the BMP2 gene.
BstXI and XbaI sites were respectively added at the
5′ end and 3′ end of the HIF1αmu primer, and a 3xFlag
tag was added at the C-cleavage site. The study was approved by the
ethics committee of Liaoning Medical University, Liaoning,
China.
Vector construction
The HIF1αmu fragment was amplified by PCR
using pShuttle-CMV-HIF1αmu-IRES-hrGFP-1 plasmid as the
template. The target fragment was recovered and double enzyme
digested with BstXI and XbaI. The vector pIRES2-EGFP
was double enzyme digested with BstXI and XbaI and
recovered by agarose gel electrophoresis. The BMP2 fragment was
amplified by PCR with pShuttle-CMV-BMP2 plasmid as the template.
The target fragment was double enzyme digested with NheI and
BamHI and recovered by agarose gel electrophoresis. The
correct pIRES2-HIF11αmu-flag vector was double enzyme
digested with NheI and BamHI and the larger fragment
was recovered. The previously mentioned enzyme was digested and the
recovered target gene was ligated with the vector fragment for 2 h
at 22°C, and then transformed into competent E. coli
DH5α.
Screening and identifying of positive
clones
The positive clones identified by colony PCR were
transferred into Kan-LB and agitated overnight at 37°C, and then
the plasmid was extracted the next day. The positive clones
screened by colony PCR and enzyme digestion were sequenced. The
confirmed clones were stored.
Recombination of adenovirus expression
vector
BMP2-IRES-HIF1αmu recombinant fragment
was amplified with BMP2-pIRES2-HIF1αmu vector as the
template and then recovered. The target fragment
BMP2-IRES-HIF1αmu was recombined to the pDONR221 vector
using the BP recombinant system of Invitrogen (Carlsbad, CA, USA).
The target sequence BMP2-IRES-HIF1αmu was recombined to
adenovirus vector pAd-BMP2-IRES-HIF1αmu using the LP
recombinant system of Invitrogen. The recombinant plasmids were
sequenced, and stored at −20°C until further use.
Titration of virus liquid
The target gene was transferred into HEK293 cells in
the exponential phase and the transfer process was observed under a
fluorescence microscope. An end-point dilution assay was used to
determine the titer of virus liquid.
Recombinant adenovirus transfection of
rabbit MSCs
Group A (experimental group): transfection with
Ad-BMP2-IRES-HIF1αmu; group B (positive control group
1): transfection with Ad-HIF1αmu-IRES-hrGFP-1; group C
(positive control group 2): transfection with Ad-BMP2-IRES-hrGFP-1;
group D (negative control group): transfection with
Ad-IRES-hrGFP-1; group E (blank group): without transfection
virus.
Rabbit MSCs within three generations were digested
by trypsin and mixed, then transferred into 6-well plates at a
density of 5×105/well, with routine culture for 24 h in
a cell culture box. The supernatant was discarded following firm
adherence.
Multiplicity of infection (MOI) = (pfu/ml value) ×
(virus liquid volume)/(cell number for transfection). Virus liquids
at MOI of 50, 100, 150 and 200 were respectively transferred into
MSCs, leaving two wells as the negative controls. Following
incubation for 1 h at 37°C, the cells were routinely cultured for
48–72 h with complete medium and then observed under the inversion
fluorescence microscope.
Every virus liquid in each group was respectively
transferred into rabbit MSCs at its best MOI value. The largest MOI
not causing a marked cytopathic effect was considered as the best
MOI. In this experiment, the best MOI=100. Fresh serum medium (2
ml) was respectively added to each flask of cells, and cells were
cultured for 3 h in a constant temperature cell culture box, then
an additional 2 ml medium was added to each flask of cells, with
continued culture for 24–72 h. The transfer effect was observed
under the inversion fluorescence microscope.
RT-PCR
Total RNA was extracted according to a kit that
contained 2 μl PrimeScript 1 Step Enzyme Mix and 25 μl 2X 1 Step
Buffer (Dye Plus). Then 2 μl upstream primer and 2 μl downstream
primer, 1 μl total RNA and 18 μl RNase-free ddH2O were
added to make up the 50 μl RT-PCR reaction solution. The RT-PCR
reaction was as follows: a cycle of 50°C for 30 min; a cycle of
94°C for 2 min; 30 cycles of 94°C for 30 sec, 60°C for 30 sec and
72°C for 1 min. After the reaction, 5–8 μl reaction solutions
underwent 1% agarose gel electrophoresis for 40 min. The
electrophoresis results were observed and the OD value of strips
was detected using a gel imaging system. This was repeated for
three times, and the relative OD values were separately
calculated.
Western blot analysis
Cellular total protein in each group was extracted
using cellular protein lysis buffer, and the protein concentration
in each group was detected by BCA; 5% spacer gel and 8% separation
gel were prepared for polyacrylamide gel electrophoresis at 60 V
for 30 min, and then 150 V for 1 h. Following electrophoresis, the
separation gel was washed three times and then placed in a transfer
box at 100 mA for 30 min. The film was then incubated with the
primary antibodies (1:1500) overnight at 4°C. The film was then
washed and then incubated with the secondary antibodies and
developer solution for 30 min at room temperature, avoiding light.
The film was then washed with eluent three times to terminate the
chromogenic reaction. The OD value of target strips and internal
reference on the film were analyzed using a gel imaging system. The
experiment was repeated three times, and the relative OD values
were calculated.
Statistical analysis
The experimental data were statistically analyzed
using SPSS17.0 software. Measurement data were expressed as the
means ± standard deviation (±s) and analyzed by one-way analysis of
variance. P<0.05 was considered to indicate a statistically
significant difference.
Results
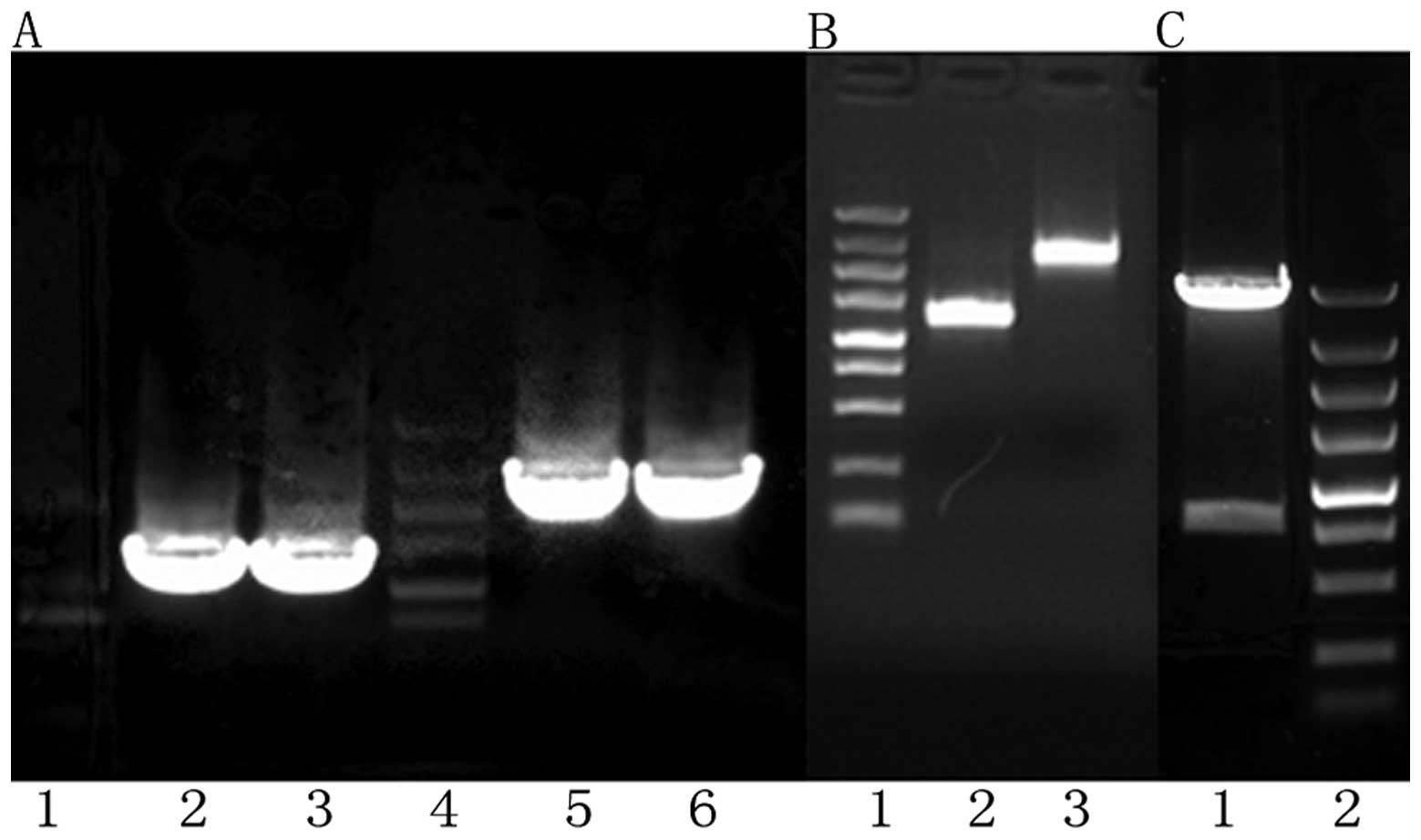
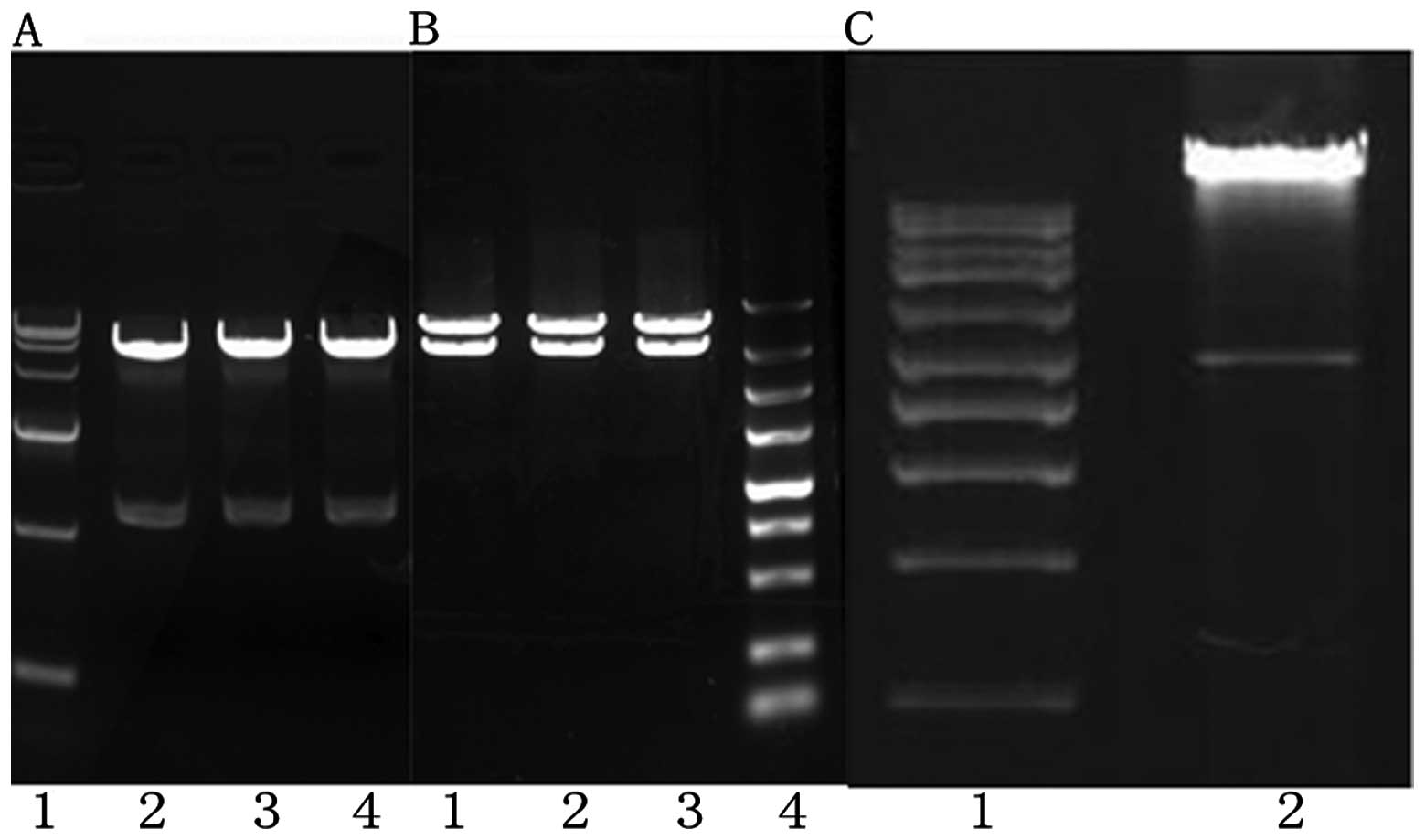
Recombinant identification
A 9789 bp recombinant plasmid strip, a 1191 bp BMP2
strip and a 2481 bp HIF1α strip were observed after agarose gel
electrophoresis, and the zymogram analysis of recombinant plasmid
was the same as expected. The BMP2 fragment and the HIF1α fragment
in the recombinant plasmid completely coincided with the BMP2
(NM001200) CDS area and the HIF1α (NM001530) CDS area in Genebank,
respectively, with insert fragments in the correct direction
(Figs. 1 and 2A and B).
Adv-BMP-2-IRES-HIF1αmu
identification
There were two strips at 3000 and 10000 bp as shown
by agarose gel electrophoresis following enzyme digestion, which
indicated that the shuttle plasmid had been successfully ligated to
the adenovirus genome and Adv-BMP-2-IRES-HIF1αmu had
been successfully constructed. The target fragment BMP2 was
detected by RT-PCR and agarose gel electrophoresis, and there was a
strip at 1191 bp, which is the same as expected. The target
fragment HIF1α was detected by RT-PCR and agarose gel
electrophoresis, and there was a strip at 2481 bp, which was the
same as expected (Fig. 2C).
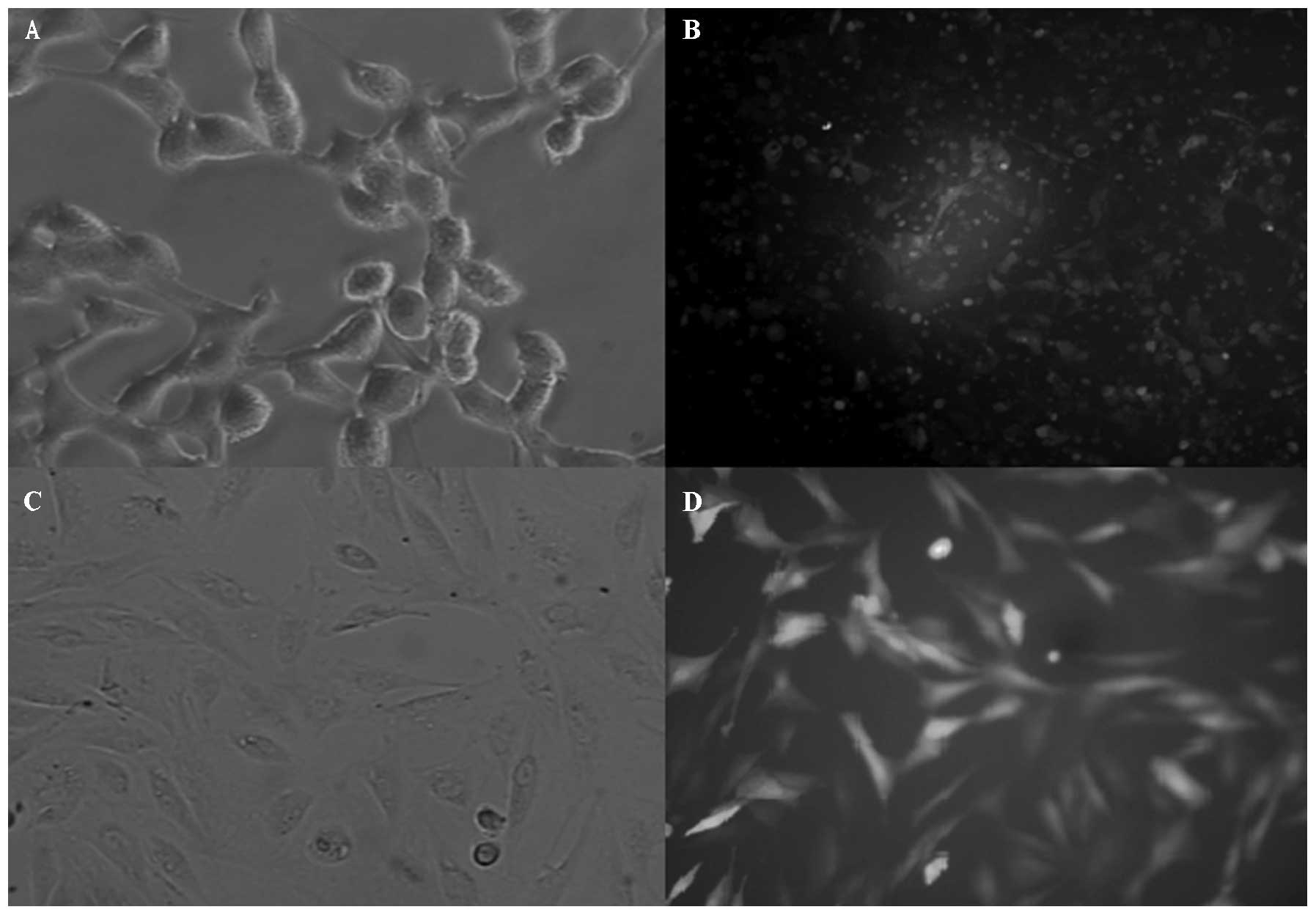
Transfer and titration of virus
liquid
Under inverted fluorescence microscopy, the majority
of cells appeared green and cell fragments had become detached and
showed a cytopathic effect. The titer of virus liquid was
3.16×108 according to the end-point dilution assay
(Fig. 3A and B).
Cell culture
After the original cultivation of rabbit bone marrow
stromal cells for one week, there was cell adherence to the wall
when the culture solution was changed. Cells were passaged when
they reached 80% confluence. The third generation had a higher
purity and a good adhesion (Fig. 3C
and D).
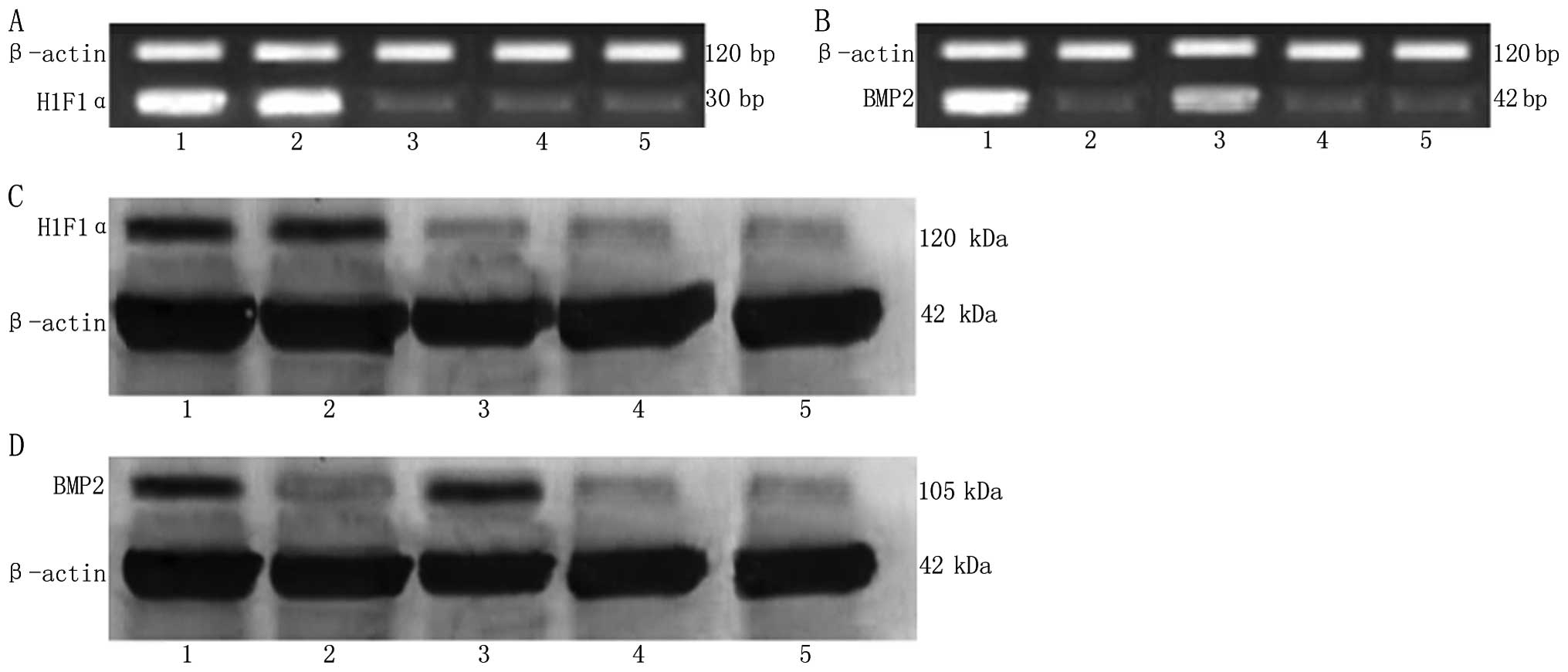
RT-PCR
There was no marked difference in the HIF1α mRNA
expression level between group A and B (P>0.05), with the same
result among groups C, D and E (P>0.05). The HIF1α mRNA
expression level in group A and B was markedly higher than that in
groups C, D and E, with a significant difference (P<0.01). There
was a significant difference in BMP2 mRNA expression level between
group A and C (P>0.05), but no marked difference among groups B,
D and E (P>0.05). The BMP2 mRNA expression level in group A and
C were markedly higher than that in groups B, D and E, with a
significant difference (P<0.01) (Fig. 4A and B, Table I).
 | Table IOD value of β-actin, BMP2 and HIF1α
detected by RT-PCR (mean ± SD). |
Table I
OD value of β-actin, BMP2 and HIF1α
detected by RT-PCR (mean ± SD).
| Group | Group A | Group B | Group C | Group D | Group E |
|---|
| β-actin | 7.73±0.09 | 7.75±0.09 | 7.52±0.30 | 7.39±0.29 | 7.48±0.06 |
| HIF1α | 7.10±0.10 | 7.18±0.23 | 2.08±0.18 | 2.25±0.23 | 2.34±0.29 |
| BMP2 | 8.13±0.08 | 2.18±0.15 | 6.20±0.05 | 2.18±0.18 | 2.14±0.09 |
| HIF1α relative OD
value | 0.92±0.02 | 0.93±0.04 | 0.28±0.03 | 0.30±0.02 | 0.31±0.04 |
| BMP2 relative OD
value | 1.05±0.01 | 0.28±0.02 | 0.82±0.03 | 0.29±0.04 | 0.29±0.03 |
Western blot analysis
The HIF1α protein expression level in group A and B
was significantly higher than that in the other three groups, with
a significant difference (P<0.01). However, there was no
significant difference among groups C, D and E (P>0.1). BMP2
protein expression level in group A and C was significantly higher
than that in the other three groups, with a significant difference
(P<0.01). However, there was no significant difference among
groups B, D and E (P>0.1). There was also a significant
difference in the BMP2 protein expression level between group A and
C (P<0.05) (Fig. 4C and D,
Table II).
| Table IIOD value of β-actin, BMP2 and HIF1α
detected by western blot analysis (mean ± SD). |
Table II
OD value of β-actin, BMP2 and HIF1α
detected by western blot analysis (mean ± SD).
| Group | Group A | Group B | Group C | Group D | Group E |
|---|
| β-actin | 2.93±0.09 | 2.98±0.11 | 2.99±0.21 | 2.95±0.12 | 2.90±0.07 |
| HIF1α | 1.02±0.14 | 1.12±0.18 | 0.18±0.08 | 0.18±0.12 | 0.17±0.13 |
| BMP2 | 1.51±0.02 | 0.18±0.14 | 1.02±0.23 | 0.18±0.14 | 0.17±0.14 |
| HIF1α relative OD
value | 0.35±0.03 | 0.38±0.02 | 0.06±0.03 | 0.06±0.04 | 0.06±0.04 |
| BMP2 relative OD
value | 0.52±0.02 | 0.06±0.05 | 0.34±0.03 | 0.06±0.02 | 0.06±0.04 |
Discussion
Previous studies reported that the incidence rate of
delayed union after fracture and non-union was approximately 5–10%,
and treatment of this remains a big problem in the field of
orthopedics (8–10). In the BMP family, BMP2 is the main
signaling protein regulating bone formation (11–13).
BMP2 could induce differentiation of pluripotent mesenchymal
progenitor cells into osteoblast cells, promoting bone formation of
osteoblast cells. There were also studies confirming that BMP2 was
capable of inducing pluripotent mesenchymal progenitor cells to
differentiate into osteoblast cells in vitro or in
vivo(14–15). During the process of bone defect
and fracture healing, BMP2 promoted the osteogenetic effect at the
fracture site, but following osteoblast induction, the lack of
blood supply at the fracture site and difficult angiogenesis
affected the repair process of the fracture.
It has been demonstrated that the activating
transcription factor HIF1α found in recent years is capable of
activating low oxygen metabolism and regulating the transcription
of angiogenesis-promoting genes. At the same time it also
participates in the regulation of VEGF and SDF21 expression
(16). Therefore, HIF1α has been
considered as one of the most promising genes in the clinical
application of promoting angiogenesis. However, HIF1α loses its
transcription activity due to its easy degradation under usual
oxygen conditions (17–19). Liu et al(20) successfully constructed recombinant
adenovirus-mediated mutations of hypoxia inducible factor
expression vector (HIF1αmu), which could be expressed
under usual oxygen conditions and rapidly accumulate in cells and
promote angiogenesis. The double gene co-expression of the
adenovirus vector with BMP2 and HIF1αmu constructed in
this experiment was expected to form a cooperative effect due to
the expression of both genes (7),
accelerating the repair process of fractures and bone defects. The
past use of a single gene vector had a poor treatment effect, and
conjunction with many genes and vectors often increased dosages of
adenovirus and virus infection efficiency (20). The advantage of two genes with a
single vector constructed in this experiment was the reduction of
the dosage of adenovirus required during bone defect treatment.
According to the results of RT-PCR and western blot
analysis, the expression level of HIF1α mRNA was significantly
increased following transfection into MSCs in group B. The
expression level of BMP2 mRNA was significantly increased following
transfection into MSCs in group C. The expression level of BMP2
mRNA and HIF1α mRNA were both not increased following transfection
into MSCs in group D and E, demonstrating that it was not
adenovirus expression vector itself or hrGFP-1 that caused the
increased expression level of BMP2 and HIF1α mRNA. The expression
levels of HIF1α mRNA and BMP2 mRNA were significantly increased at
the same time after transfection into MSCs in group A, and the
expression of BMP2 was higher than that in group C. There was no
significant difference in HIF1α expression between group A and B,
indicating that mutant HIF1α was likely to promote the osteogenetic
function of BMP2, which would provide a new direction for
angiogenic treatment of bone defect diseases. This point was
consistent with the experimental results we had expected.
The angiogenesis process in the osteogenesis system
is an extremely complex physical process. Importing mutations of
the HIF1α gene was an abnormal way to normalize tissue, and
promoted the formation of a new vascular network, but with an
unknown effect on other factors themselves participating in the
process of angiogenesis. The effect of mutant genes on HIF1α itself
will also require further research.
Acknowledgements
This study was supported by the Key talent project
of Liaoning Provincial Health Bureau, China (No. 2010921045).
References
|
1
|
Jorgensen NR, Henriksen Z, Sorensen OH and
Civitelli R: Dexamethasone, BMP-2, and 1, 25-dihydroxyvitamin D
enhance a more differentiated osteoblast phenotype: validation of
an in vitro model for human bone marrow-derived primary
osteoblasts. Steroids. 69:219–226. 2004. View Article : Google Scholar
|
|
2
|
Shin JH, Kim KH, Kim SH, et al: Ex vivo
bone morphogenetic protein-2 gene delivery using gingival
fibroblasts promotes bone regeneration in rats. J Clin Periodontol.
37:305–311. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cheng H, Jiang W, Phillips FM, et al:
Osteogenic activity of the fourteen types of human bone
morphogenetic proteins (BMPs). Bone Joint Surg Am. 85:1544–1552.
2003.PubMed/NCBI
|
|
4
|
Wan C, Gilbert SR, Wang Y, et al:
Activation of the hypoxia-inducible factor-1-alpha pathway
accelerates. Proc Nat Acad Sci U S A. 105:686–691. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hua Z, Nicola JM, Margaret TM and Simons
J: Nuclear expression of hypoxia-inducible factor 1alpha protein is
heterogeneous in human malignantcells under normoxic conditions.
Cancer Lett. 181:2332002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang Y, Wan C, Deng L, et al: The
hypoxia-inducible factor alpha pathway couples angiogenesis to
osteogenesis during skeletal development. J Clin Invest.
117:1616–1626. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Giaccia A, Siim BG and Johnson RS: HIF-1
as a target for drug development. Nat Rev Drug Discov. 2:803–811.
2003. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Diab T, Pritchard EM, Uhrig BA, Boerckel
JD, Kaplan DL and Guldberg RE: A silk hydrogel-based delivery
system of bone morphogenetic protein for the treatment of large
bone defects. J Mech Behav Biomed Mater. 11:123–131. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wigner NA, Luderer HF, Cox MK, Sooy K,
Gerstenfeld LC and Demay MB: Acute phosphate restriction leads to
impaired fracture healing and resistance to BMP-2. J Bone Miner
Res. 25:724–733. 2010.PubMed/NCBI
|
|
10
|
Kim TH, Oh SH, Na SY, Chun SY and Lee JH:
Effect of biological/physical stimulation on guided bone
regeneration through asymmetrically porous membrane. J Biomed Mater
Res A. 100:1512–1520. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chang SC, Lin TM, Chung HY, et al:
Large-scale bicortical skull bone regeneration using ex vivo
replication-defective adenoviral-mediated bone morphogenetic
protein-2 gene-transferred bone marrow stromal cells and composite
biomaterials. Neurosurgery. 65:75–81. 2009. View Article : Google Scholar
|
|
12
|
Liu Y, Lu Y, Tian X, et al: Fragmental
bone regeneration using an rhBMP-2-loaded
gelatin/nanohydroxyapatite/fibrin scaffold in a rabbit model.
Biomaterials. 30:6276–6285. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Baltzer AW, Ostapczuk MS, Stosch D and
Granrath M: The use of recombinant human bone morphogenetic
protein-2 for the treatment of a delayed union following femoral
neck open-wedge osteotomy. Orthop Rev (Pavia). 4:e42012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kanczler JM, Ginty PJ, White L, et al: The
effect of the delivery of vascular endothelial growth factor and
bone morphogenic protein-2 to osteoprogenitor cell populations on
bone formation. Biomaterials. 31:1242–1250. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kang SW, Bae JH, Park SA, et al:
Combination therapy with BMP-2 and BMSCs enhances bone healing
efficacy of PCL scaffold fabricated using the 3D plotting system in
a large fragmental defect model. Biotechnol Lett. 34:1375–1384.
2012. View Article : Google Scholar
|
|
16
|
Simon F, Bockhorn M, Praha C, et al:
Deregulation of HIF1-alpha and hypoxia- regulated pathways in
hepatocellular carcinoma and corresponding non-malignant liver
tissue--influence of a modulated host stroma on the prognosis of
HCC. Langenbecks Arch Surg. 395:395–405. 2010. View Article : Google Scholar
|
|
17
|
Lando D, Peet DJ, Whelan DA, Gorman JJ and
Whitelaw ML: Asparagine hydroxylation of the HIF transactivation
domain a hypoxic switch. Science. 295:858–861. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lim W, Cho J, Kwon HY, Park Y, Rhyu MR and
Lee Y: Hypoxia-inducible factor 1 alpha activates and is inhibited
by unoccupied estrogen receptor beta. FEBS Lett. 583:1314–1318.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Elson DA, Thurston G, Huang LE, et al:
Induction of hypervascularity without leakage or inflammation in
transgenic mice overexpressing hypoxia-inducible factor-1alpha.
Genes Dev. 15:2520–2532. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu DP, Wang GX, Hu L and Li C:
Construction of adenovirus-mediated eukaryotic expression vector
co-expressing mutant hypoxia-inducible factor-1 alpha target
protein and humanized Renilla reniformis green fluorescent protein
reporter molecule under normoxic conditions. J Clin Rehab Tiss Eng
Res. 14:3787–3792. 2010.
|