Introduction
Post-traumatic stress disorder (PTSD) is an anxiety
disorder that develops following exposure to a life-threatening
traumatic experience and is characterized by symptoms that often
endure for years, including continuous re-experience of the
traumatic event, avoidance of stimuli associated with the trauma,
numbing of general responsiveness and increased arousal (1). The pathophysiology of PTSD has been
studied widely in neuroscience (2), however, the mechanism is not fully
understood.
It is well known that the dysfunction of the
hypothalamo-pituitary-adrenal (HPA) axis is one of the core
neuroendocrine abnormalities correlated with PTSD, characterized by
low levels of adrenocorticotropic hormone (ACTH), plasma and
urinary cortisol and enhanced suppression of cortisol in response
to a low-dose administration of dexamethasone (3–6).
Single-prolonged stress (SPS) is a reliable animal model of PTSD
based on the time-dependent dysregulation of the HPA axis and has
been developed and employed for PTSD research (7–9). A
number of studies have shown that the medial prefrontal cortex
(mPFC) is important in modulating the responses of the HPA axis to
emotional stress (10,11). Thus, the mPFC has been investigated
to reveal the mechanism(s) of PTSD.
There are low levels of plasma glucocorticoid (GC)
in PTSD (2). Negative feedback on
the HPA axis regulated by GC occurs via the high density of
corticosteroid receptors (CsRs) (12). CsRs in the brain can be divided
into two types, I and II. Type I has a high affinity for
corticosterone and is similar to the human mineralocorticoid
receptor (MR). Type II has a low affinity for corticosterone,
corresponding to the human glucocorticoid receptor (GR) (13). MR and GR are involved in the
physiological regulation of the HPA axis. MR is involved in the
negative feedback regulation of the HPA axis at the basic level,
which maintains basal activities of the HPA axis and is correlated
with the threshold and sensitivity of the central stress response
(14).
Little is currently known regarding MR in the brain,
particularly in the mPFC, due to the differences between the roles
of MR in the brain and other areas. In this study, MR expression in
the mPFC of PTSD rats was examined in order to provide experimental
evidence for further studies related to the pathogenesis of
HPA.
Materials and methods
Experimental animals
A total of 80 male Wistar rats aged 7 or 8 weeks at
the start of the study and weighing approximately 180 g were
provided by the Animal Experimental Center, China Medical
University (Shenyang, China). Rats were individually housed in an
air-conditioned room (22±1°C and 55±5% humidity) on a 12-h
light/dark cycle with free access to food and water.
The rats were raised in the laboratory for at least
7 days prior to conducting the experiment. Experiments were carried
out in accordance with the National Institute of Health Guide for
the care and use of laboratory animals. All efforts were made to
reduce the number of animals used and to minimize animal suffering
during the experiment.
Model establishment and grouping
Animals were divided randomly into 5 groups (n=18):
i) control; ii) SPS 1 day; iii) SPS 7 days; iv) SPS 14 days; v) SPS
28 days. Control animals remained in their home cages with no
handling for 7 days and were sacrificed at the same time as the SPS
groups. SPS-rats underwent the SPS procedure on the first day. The
SPS protocol was based on a combined plural stress paradigm
(7,15): immobilization (compression with
plastic bags) for 2 h, forced swimming for 20 min (24±1°C), rest
for 15 min, followed by ether anesthesia (until consciousness was
lost). The rats were returned to their home cage and left
undisturbed until they were sacrificed for experiments.
Perfusion-based sections
Rats from both the normal control and SPS groups
were prepared by left ventricle perfusion and fixation (16) and post-fixed in the same fixative
at 4°C for 6–10 h. The samples were then immersed in a 20% sucrose
solution in 0.01 M phosphate buffer (PB, pH 7.4) at 4°C. Samples
were fast frozen on liquid nitrogen and sectioned, 12-μm coronal
sections were prepared for morphological studies.
Immunohistochemical analysis of MR
The sections were treated with 5% bovine serum
albumin (BSA), and 0.3% Triton X-100 in PBS for 30 min at room
temperature (RT) to block non-specific staining. The sections were
then incubated with rabbit polyclonal antibody against MR (Santa
Cruz Biotechnology, Inc., Santa Cruz, CA, USA; 1:500) in 2% BSA-PBS
overnight at 4°C. After being washed with PBS, the sections were
incubated with goat anti-rabbit IgG (Boster Biological Technology,
Ltd., Wuhan, China; 1:100) for 2 h and then with
streptomycin-avidin-biotin-peroxidase complex (SABC) for 1 h. The
sections were washed three times with PBS after each incubation and
subsequently incubated with 3,3′-diaminobenzidine (DAB) and
H2O2. To assess non-specific staining, a few
sections in every experiment were incubated in buffer without
primary antibody.
Five slides were randomly selected from each group.
Each slide was randomly selected from five visual fields in the
mPFC (x400). The optical density (OD) of positive cells in each
field was recorded to evaluate the average. The OD of the
MR-immunopositive cells was analyzed using a MetaMorph/DPIO/BX41
morphology image analysis system.
Western blotting of MR
Rats of each group were rapidly decapitated and the
brains were removed and immediately placed in a dish standing on
crushed ice. The mPFC was then dissected according to the rat brain
atlas of Paxinos et al (17) using a stereomicroscope, and
snap-frozen in liquid nitrogen. Materials were obtained as above.
Samples of normal control and SPS rats were homogenized with a
sample buffer containing 200 mM TBS, pH 7.5, 4% SDS, 20% glycerol,
10% 2-mercaptoethanol and were denatured by boiling for 3 min. The
protein fraction (30 μg/lane) prepared from each sample was
separated by 12% (w/v) gradient sodium dodecyl sulfate
(SDS)-polyacrylamide gel electrophoresis (PAGE) and electroblotted
onto a PVDF membrane (Millipore, Bedford, MA, USA) from the gel by
a semi-dry blotting apparatus (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA).
The membrane was blocked with 5% dried skimmed milk,
0.05% Tween-20 in TBST at room temperature for 2 h and incubated
with rabbit anti-MR polyclonal antibody (1:1,000) overnight at
4°C.
The blots were washed three times with TBST and then
incubated with goat anti-rabbit IgG-HRP (Santa Cruz Biotechnology
Inc.; 1:5,000) for 2 h at room temperature and washed with TBST.
Following incubation, the PVDF membrane was washed three times with
TBST prior to visualization by enhanced chemiluminescence (ECL;
Amersham Pharmacia Biotech, Buckinghamshire, UK). To confirm equal
protein loading the same blots were reincubated with antibodies
specific for β-actin (Abcam, Cambridge, UK; 1:2,500). β-actin
immunoreactivity was detected using ECL. The OD was analyzed on the
Gel Image Analysis System (Tanon 2500R, Shanghai, China). Each MR
band was normalized with respect to its corresponding β-actin band
and the values were expressed as an intensity ratio.
Reverse transcription-polymerase chain
reaction (RT-PCR) was used to detect MR
Total mRNA of each group was extracted from the mPFC
according to the instructions of the TRIzol kit (Invitrogen,
Carlsbad, CA, USA) and 1 μg of total RNA was reverse transcribed
into cDNA. An RNA PCR kit (AM Ver. 3.0, Takara Bio, Inc., Otsu,
Japan) was used to amplify the cDNA. The primers were designed and
synthesized by Shenggong Biotech Company (Shanghai, China)
according to the serial number from Genbank (Table I). The reaction was started at 94°C
for 4 min and amplified to 36 cycles of 45 sec at 94°C, 40 sec at
60°C, 50 sec at 72°C and ended with a 10-min extension at 72°C.
β-actin mRNA used as an internal control was co-amplified with
MR-mRNA. The products were observed after electrophoresis on 1.2%
agarose gel and the OD of each band was analyzed on the Gel Image
Analysis System. The level of MR-mRNA was determined by calculating
the density ratio of MR mRNA/β-actin mRNA.
 | Table IMR and β-actin sequences. |
Table I
MR and β-actin sequences.
| Name | Upstream primer
5′-3′ | Downstream primer
5′-3′ | Product size
(bp) |
|---|
| MR |
agaagatgcatcagtctgcc |
gtgatgatctccaccagcat | 380 |
| β-actin |
atcacccacactgtgcccatc |
acagagtacttgcgctcagga | 542 |
Statistical analysis
Data were shown as the means ± standard error. Data
among groups were analyzed by one-way analysis of variance (ANOVA)
using SPSS 13.0 software (SPSS, Inc., Chicago, IL, USA). P<0.05
was considered to indicate a statistically significant
difference.
Results
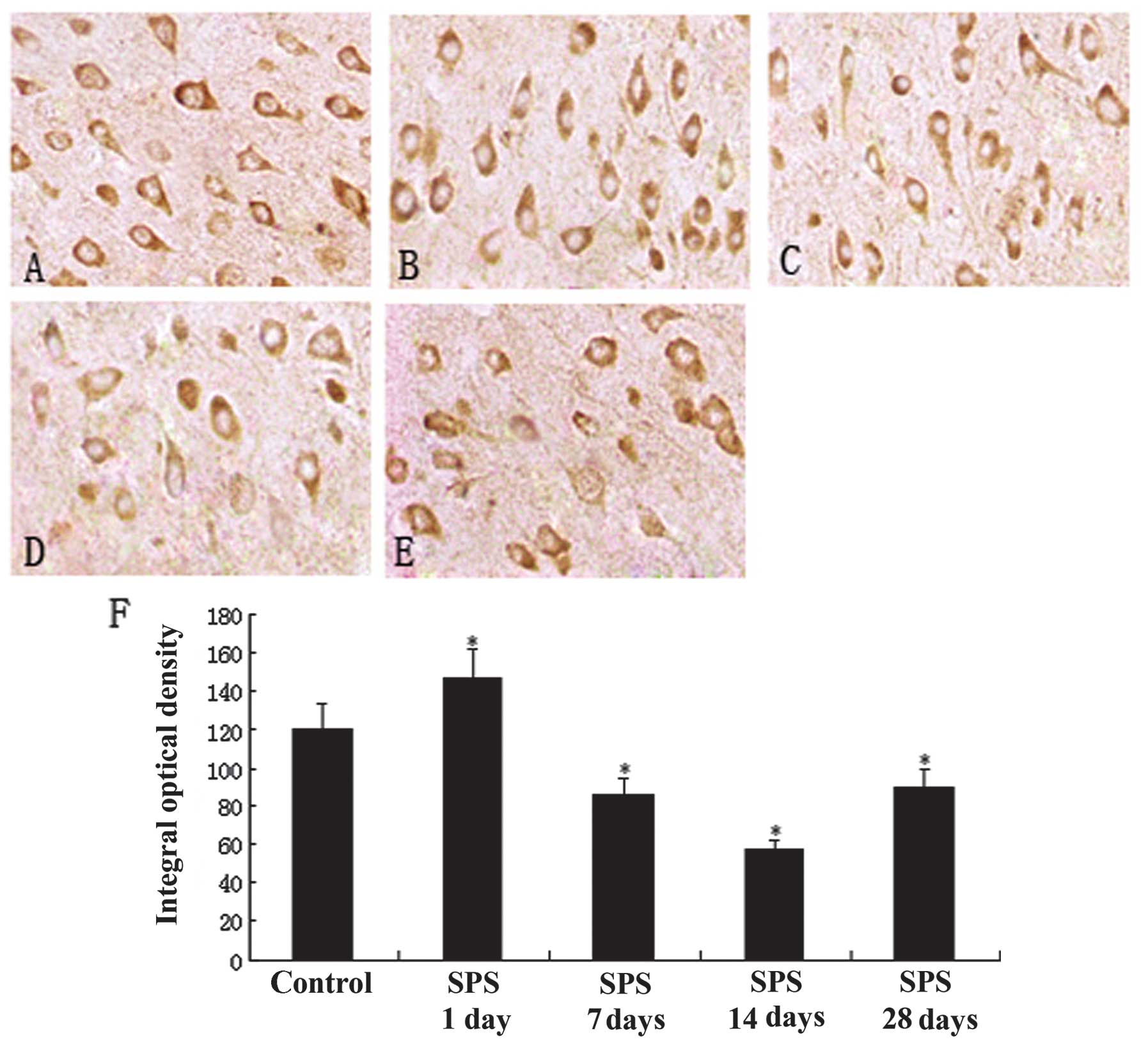
Immunohistochemical analysis results of
MR
MR was widely distributed throughout the mPFC
region, primarily in the cytoplasm, and appeared as puffy particles
(Fig. 1A-E). Evaluation of MR
content by immunohistochemistry indicated a significant change in
the SPS model groups compared with the normal control group. The
staining results showed that the number of MR-positive cells was
less in the normal control group (Fig.
1A) and that of SPS rats was significantly increased and peaked
at 7 days after exposure to SPS (P<0.01; Fig. 1C). Positive MR expression gradually
decreased in the SPS-14 day group (Fig. 1D). However, in the SPS-28 day group
positive expression increased again (Fig. 1E), but remained lower than that of
the control group. The analysis results are shown in Fig. 1F (P<0.05).
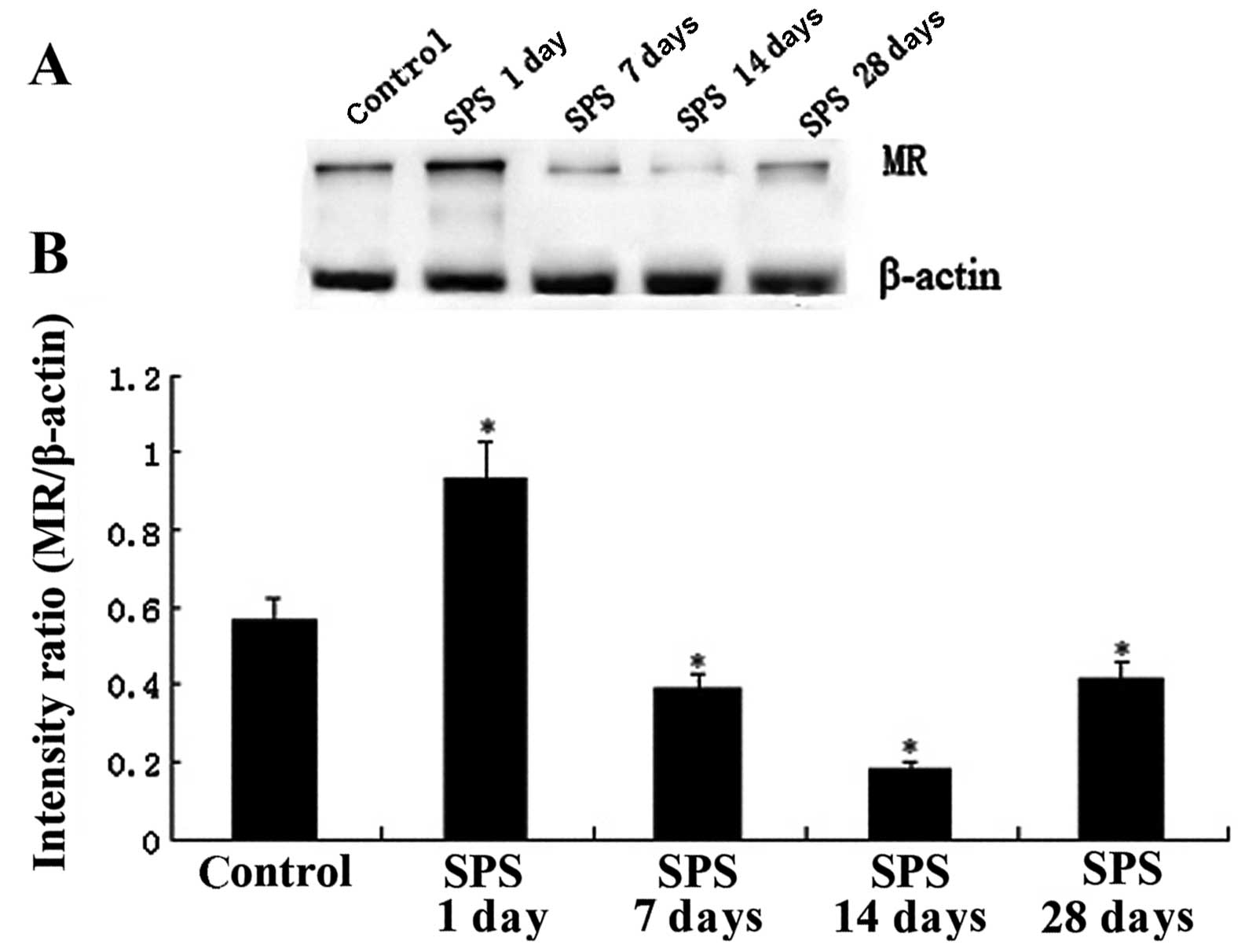
Western blotting analysis of MR
Immunoreactive signals for MR and β-actin appeared
on western blotting at 112 and 42 kDa, respectively (data not
shown), while the mean value of band densities of the control group
was set as 100%. Data were expressed as normalized OD. Changes of
MR expression in the mPFC region between the control and SPS groups
are shown in Fig. 2 (P<0.05).
The MR protein expression of mPFC showed a significant change
following SPS stimulation and the trend of change was consistent
with the results of the immunohistochemistry.
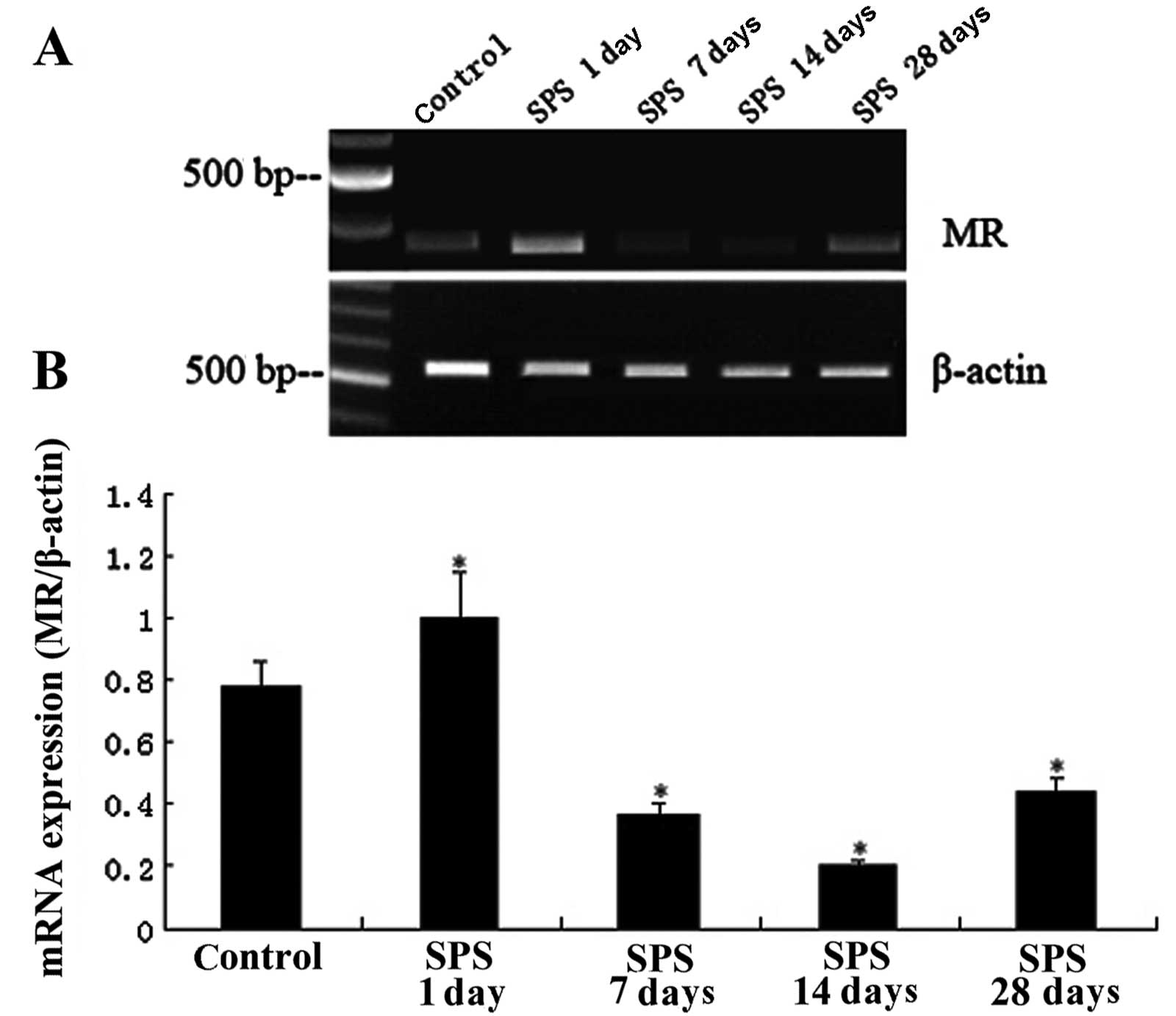
RT-PCR analysis of MR mRNA
The levels of MR mRNA were normalized with the
β-actin mRNA level. The results indicated a significant change
following SPS stimulation and the trend of the change was
consistent with the results of the immunohistochemical and western
blot analysis (P<0.05; Fig.
3).
Discussion
PTSD is thought to involve a dysfunction in the
response to fear-related stimuli. The four major characteristic
symptoms of PTSD are: re-experiencing, avoidance, numbing and
hyperarousal (1).
Previous studies have paid little attention to the
role of MR in the rat central nervous system compared with that of
GR and have mainly investigated an adjusting function of the
water-salt balance in epithelial cells. The present opinion is that
MR exists in the brain tissue, most substantially in the
hippocampus. GR and MR coexist in certain brain regions, including
the hippocampus CA1, CA2, CA3 and dentate gyrus sub-region
(18). The choice function of MR
on mineralocorticoid (aldosterone) in the hippocampus is different
from that in epithelial cells. GC combines with the two sets of
receptors, GR and MR, at the same time after stimulation. The
binding affinity of corticosterone to MR is approximately 10 times
higher than that to GR. Low levels of GC combine with MR first,
only high levels of GC can combine with GR after MR is saturated.
MR is a high-affinity and low-capacity receptor system for GC,
while GR is a low-affinity and high-capacity receptor system for GC
(19).
The effects of corticosterone are mediated by MR and
are also regulated by GR. GR is constantly occupied during the
physiological rhythm rising period following GC stress and GC
affects the paraventricular nucleus (PVN) through negative
feedback. The hippocampus maintains excitability when MR activity
dominates and inhibition projects to the PVN through
cross-synapses, inhibiting the basic activity of the HPA. Instead,
the activation of GR inhibits hippocampal signal output with the
increase of GC concentrations, which leads to inhibition of the PVN
neurons. The functions of the two types of receptor are
interrelated (20). MR defects
make corticosterone responses occur more quickly and produce a more
GR-mediated effect. The above information shows the importance of
MR- and GR-mediated effects in balancing HPA regulation. Therefore,
more and more researchers support the balance theory of the
receptor-mediated role, which assumes that MR primarily maintains
the function of GC and GR is mainly involved in a GC-negative
feedback role (21). Since the
affinity between MR and GC is higher than that between GR and GC,
in the early HPA axis activation or basic level, GC first combines
with the MRs of the hippocampus and maintains basal HPA activity,
known as proactive feedback. When the GC concentration continues to
rise, the high concentration GR continues to combine with GC,
inhibiting HPA axis activity via GR-negative feedback, known as
reactive feedback (22). GC allows
HPA activity to remain at the appropriate level through two types
of feedback modes, MR- or GR-mediated, imbalance of which leads to
disease. Long-term abnormal corticosteroid levels affect the
hippocampal cell response and endanger cell survival (23). Therefore, the hippocampus also
functions as a control site of the HPA axis.
One study has shown that MR also exists in the mPFC,
with the exception of the hippocampus (20). Selective knockout of MR from the
mouse forebrain damages spatial learning ability. By contrast,
selective expression of MR in the forebrain reduces anxious
behavior of the mouse after stress (20), indicating that MR also exists
outside the area of the hippocampus, but its mechanism is not
clear. In this experiment MR expression is mainly observed in the
cytoplasm of the mPFC, as the transcription role has not yet
occurred in the nucleus. MR expression was upregulated
significantly after SPS and then decreased 7 days after SPS
stimulation, indicating that there was not only a
feedback-regulated increase of GR in the hippocampus after stress,
but also a change of MR in the mPFC. Expression of MR gradually
decreased, but was not restored until 28 days later and differed
from that prior to the experiment. This finding indicates that
changes of MR in the mPFC were in accordance with HPA axis
regulation, as well as changing with time after SPS stimulation.
However, the change of MR in the mPFC may be the result of an HPA
axis change, or an MR change leading to an HPA axis change, as the
role of MR closely correlates with that of GR, which may play a
different role in the same cells. Further studies are needed to
confirm the mechanism by which changes in MR occur.
This study revealed the presence of mPFC-MR at the
protein level using immunohistochemistry and western blotting.
There were also quantitative changes in PTSD experimental animals,
which all provided an experimental basis for revealing the
pathogenetic mechanism of PTSD. Further studies are needed to
explore the relevant functional mechanisms and effective
interventions in order to find new methods of treatment. However,
this experiment is limited to the study of mPFC-MR receptors of
PTSD rats, thus, receptor changes of other parts in the brain and
its effects on the HPA-axis need further study.
At present, the pathogenesis of PTSD is not yet
entirely clear. PTSD may cause a series of biochemical
abnormalities and dysfunction of the mPFC, leading to dysfuction of
the brain. Thus, more studies should be conducted to determine the
pathogenesis of PTSD.
Acknowledgements
This research was supported by a grant from the
National Natural Science Foundation of China (No. 81171282). The
authors would like to thank the anonymous reviewers for their
valuable comments on how to improve the quality of the paper.
References
|
1
|
American Psychiatric Association.
Diagnostic and Statistical Manual of Mental Disorders. 4th edition.
DSM-IV. American Psychiatric Press; Washington, DC: 1994
|
|
2
|
de Kloet ER: Stress: a neurobiological
perspective. Tijdschr Psychiatr. 51:541–550. 2009.(In Dutch).
|
|
3
|
Yehuda R, Halligan SL, Grossman R, et al:
The cortisol and glucocorticoid receptor response to low dose
dexamethasone administration in aging combat veterans and holocaust
survivors with and without posttraumatic stress disorder. Biol
Psychiatry. 52:393–403. 2002. View Article : Google Scholar
|
|
4
|
Bachmann AW, Sedgley TL, Jackson RV,
Gibson JN, Young RM and Torpy DJ: Glucocorticoid receptor
polymorphisms and post-traumatic stress disorder.
Psychoneuroendocrinology. 30:297–306. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yehuda R, Golier JA, Halligan SL, Meaney M
and Bierer LM: The ACTH response to dexamethasone in PTSD. Am J
Psychiatry. 161:397–1403. 2004. View Article : Google Scholar
|
|
6
|
de Kloet CS, Vermetten E, Geuze E,
Kavelaars A, Heijnen CJ and Westenberg HG: Assessment of HPA-axis
function in posttraumatic stress disorder: pharmacological and
non-pharmacological challenge tests, a review. J Psychiatr Res.
40:550–567. 2006.PubMed/NCBI
|
|
7
|
Takahashi T, Morinobu S, Iwamoto Y and
Yamawaki S: Effect of paroxetine on enhanced contextual fear
induced by single prolonged stress in rats. Psychopharmacology
(Berl). 189:165–173. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Iwamoto Y, Morinobu S, Takahashi T and
Yamawaki S: Single prolonged stress increases contextual freezing
and the expression of glycine transporter 1 and vesicle-associated
membrane protein 2 mRNA in the hippocampus of rats. Prog
Neuropsychopharmacol Biol Psychiatry. 31:642–651. 2007. View Article : Google Scholar
|
|
9
|
Khan S and Liberzon I: Topiramate
attenuates exaggerated acoustic startle in an animal model of PTSD.
Psychopharmacology (Berl). 172:225–229. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Figueiredo HF, Bruestle A, Bodie B, Dolgas
CM and Herman JP: The medial prefrontal cortex differentially
regulates stress-induced c-fos expression in the forebrain
depending on type of stressor. Eur J Neurosci. 18:2357–2364. 2003.
View Article : Google Scholar
|
|
11
|
Radley JJ, Arias CM and Sawchenko PE:
Regional differentiation of the medial prefrontal cortex in
regulating adaptive responses to acute emotional stress. J
Neurosci. 26:12967–12976. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nagalski A and Kiersztan A: Physiology and
molecular mechanism of glucocorticoid action. Postepy Hig Med Dosw.
18:133–145. 2010.(In Polish).
|
|
13
|
Kessler RC: Posttraumatic stress disorder:
the burden to the individual and to society. J Clin Psychiatry.
61:4–12. 2000.PubMed/NCBI
|
|
14
|
Rogalska J: Mineralocorticoid and
glucocorticoid receptors in hippocampus: their impact on neurons
survival and behavioral impairment after neonatal brain injury.
Vitam Horm. 3:391–419. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kohda K, Harada K, Kato K, Hoshino A,
Motohashi J, Yamaji T, Morinobu S, Matsuoka N and Kato N:
Glucocorticoid receptor activation is involved in producing
abnormal phenotypes of single-prolonged stress rats: a putative
post-traumatic stress disorder model. Neuroscience. 148:22–33.
2007. View Article : Google Scholar
|
|
16
|
Liu HY: Technical operations and its
common problems of perfusion fixation in mice. Qiqihaer Yixueyuan
Xuebao. 27:13412006.
|
|
17
|
Paxinos G, Watson CR and Emson PC:
AChE-stained horizontal sections of the rat brain in stereotaxic
coordinates. J Neurosci Methods. 3:129–149. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhe D, Fang H and Yuxiu S: Expressions of
hippocampal mineralocorticoid receptor (MR) and glucocorticoid
receptor (GR) in the single-prolonged stress-rats. Acta Histochem
Cytochem. 41:89–95. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
De Kloet ER, Vreugdenhil E, Oitzl MS and
Joels M: Brain corticosteroid receptor balance in health and
disease. Endocr Rev. 19:269–330. 1998.
|
|
20
|
Kolber BJ, Wieczorek L and Muglia LJ:
Hypothalamic-pituitary-adrenal axis dysregulation and behavioral
analysis of mouse mutants with altered glucocorticoid or
mineralocorticoid receptor function. Stress. 11:321–338. 2008.
View Article : Google Scholar
|
|
21
|
Sousa N, Cerqueira JJ and Almeida OF:
Corticosteroid receptors and neuroplasticity. Brain Res Rev.
57:561–570. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nishi M and Kawata M: Dynamics of
glucocorticoid receptor and mineralocorticoid receptor:
implications from live cell imaging studies. Neuroendocrinology.
85:186–192. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kellner M, Baker DG, Yassouridis A,
Bettinger S, Otte C, Naber D and Wiedemann K: Mineralocorticoid
receptor function in patients with posttraumatic stress disorder.
Am J Psychiatry. 159:1938–1940. 2002. View Article : Google Scholar : PubMed/NCBI
|