Introduction
Colorectal cancer (CRC) is the third most common
type of malignancy in the developed western world and the third
leading cause of mortality from cancer in the United States
(1). Although the incidence of CRC
varies by ~20-fold worldwide, in Jordan, CRC is the most common
type of cancer in males and the second following breast cancer in
females (2). Although the survival
rate following excision of the primary tumor is high, when the
disease spreads to distant sites, it is usually incurable by
current systemic treatment options, including chemotherapy,
radiotherapy and immunotherapy. This is hypothesized to be largely
due to resistance of cancer cells to induction of programmed cell
death or apoptosis. Understanding and overcoming resistance
mechanism(s) of cancer cells to apoptosis may simplify the
identification of novel therapeutic targets and development of new
treatment options.
Taxanes, including paclitaxel and docetaxel are an
essential class of chemotherapeutic agents that exert anticancer
effects in vitro and in vivo against a number of
types of cancer, including lung, ovary, breast, and head and neck
cancer (3). The two taxanes were
isolated from the bark of the American Yew (Taxus
brevifolia) and later semi-synthesized from the foliage and
seeds of European Yew (Taxus baccata). Initially, taxanes
were described as antimitotic agents that bind to β-tubulin,
stabilize the microtubule polymer and protect it from disassembly,
which differs from other tubulin-targeted agents, including
colchicine that inhibit microtubule assembly resulting in blockage
of the cell cycle at the G2/M phase and cell apoptosis (4,5).
A number of mechanisms were proposed to explain
cancer cell resistance to taxanes, one of which is the
over-expression of P-glycoprotein which induces efflux of the drug
(6,7). Another reported mechanism that leads
to cancer cell resistance is the ability of taxanes to activate
numerous prosurvival signaling pathways, including the
Ras-Raf-MEK-ERK pathway (8–10).
The mitogen-activated protein kinases (MAPKs) are a
family of intracellular enzymes that comprise essential mediators
of signal transduction pathways and organize the cell response to
different extracellular stimuli (11,12).
The MAPK family comprises three major MAP kinase groups:
extracellular signal-regulated kinase (Erk1 and 2); c-Jun
N-terminal kinases (JNK 1, 2 and 3) and p38 MAPK (p38α, β, δ and γ
(13). Phosphorylation of MAPK by
MAPK kinase leads to its activation, each MAPK is activated by
specific MEK-protein kinase and individual MAPKs regulate distinct
cell processes involved in cell division, differentiation and
survival. The Ras-MEK-ERK pathway is associated with suppression of
apoptosis, while stress-activated kinases are associated with the
induction of apoptosis. ERK1/2 is located downstream of a group of
kinases, including protein kinase C (PKC), MEK1 and Raf-1. When
stimulated by extracellular signals, the kinases are
phosphorylated, leading to their activation. Ras/Raf/MEK/ERK
signaling activation is a hallmark of various types of cancer,
including breast, lung and colorectal cancer as well as melanoma
(14). Moreover, previous studies
have reported that variation in the composition and β-tubulin
isotype mutations, may lead to cancer cell resistance to taxanes
(15–17). Previously, it was shown that the
unfolded protein response (UPR), evident by the expression of
endoplasmic reticulum (ER) chaperone glucose-regulated protein 78
(GRP78), contributes to the antiapoptotic effects and resistance to
chemotherapy in a number of types of cancer (18,19).
Caffeine (1,3,7-trimethylxanthine) is a natural
stimulatory substance that is commonly present in a number of
sources, including leaves of tea, coffee beans, cocoa beverages,
soft drinks and even in chocolate products (20). Caffeine is also available in a
number of over-the-counter (OTC) remedies, including some pain
killers (21). Due to its frequent
and common consumption, caffeine may be considered the most
frequently ingested neuroactive compound (22,23).
Previous studies have reported several effects of caffeine,
including the suppression of cell proliferation (24), enhancement of radiation and
anticancer agents toxicity, as well as the elimination of chemical-
or radiation-induced delays in cell cycle progression (25,26).
Caffeine is potentially capable of inhibiting ultraviolet B-induced
skin cancer in mice (27,28) and a study by Nomura et
al(29) showed that caffeine
(0.5 mM) may inhibit malignant cell transformation induced by
epidermal growth factor. Caffeine has been the subject of intensive
studies due to various effects reported, including the effect of
caffeine on cell cycle function, the ability to induce apoptosis
and affecting specific regulatory proteins, including p53 (30–32).
In the current study, the effect of caffeine on paclitaxel-induced
cytotoxicity was investigated in CRC cell lines. In addition,
possible molecular targets for this effect were evaluated.
Materials and methods
Cell lines, antibodies and other
reagents
The human colon cancer cell line Colo205, was
cultured in DMEM containing 10% FCS (BioWhittaker, Verviers,
Belgium).
Paclitaxel was purchased from Sigma-Aldrich (St.
Louis, MO, USA) and stored as a 20 mM solution in dimethyl
sulfoxide (DMSO) with a final concentration of 0.1% (v/v) at −80°C
and diluted with DMEM medium prior to use. Propidium iodide (PI)
was purchased from Sigma-Aldrich. The rabbit polyclonal antibody
against ERK1/2 was purchased from Cell Signalling Technology
(Beverly, MA, USA). The ERK1/2 inhibitor (MEK Inhibitor, U0126) was
purchased from Calbiochem (La Jolla, CA, USA). Phosphorylated
ERK1/2, the rabbit MAbs against GRP78/Bip and Mcl-1 antibodies were
all purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA,
USA).
Apoptosis
Quantification of apoptotic cells by measurement of
sub-G1 DNA content using the PI method was carried out as described
in a previous study (17).
Western blot and protein expression
analysis
Cells were cultured to 80% confluency. The cells
were trypsinised and washed with medium twice followed by one cold
PBS wash. The cells were lysed with a Triton X-100 based lysis
buffer [10% Triton X-100, 10% glycerol, 150 mM NaCl, 20 mM Tris
(pH=7.5), 2 mM ethylene diamine tetraacetic acid (EDTA), 1 mM
phenylmethylsulphonyl fluoride (PMSF), 10 × 10−6 g
aprotinin and 10 × 10−6 g leupeptin]. Cell lysates were
transferred into microcentrifuge tubes and following 1 h on ice
were centrifuged at 4,500 × g for 30 min at 40°C. The protein
content of cell extracts was determined by the Bradford assay
(Bio-Rad, Hercules, CA, USA). A total of 20–30 μg of protein was
electrophoresed on 10–15% SDS-PAGE gels and transferred to
nitrocellulose membranes. The membranes were blocked, incubated
with primary Abs at the appropriate concentration and subsequently
incubated with horseradish peroxidase-conjugated goat anti-rabbit
IgG or goat anti-mouse IgG (1:3,000 dilutions; Bio-Rad). Labeled
bands were detected by an Immun-Star™ HRP Chemiluminescent kit and
images were captured. The intensity of the bands was quantified
with the Bio-Rad VersaDoc™ image system. The relative expression of
specific proteins was determined by dividing the densitometric
value of the test protein by that of the control (GAPDH).
Statistical analysis
Data are expressed as mean ± SE. The statistical
significance of intergroup differences in normally distributed
continuous variables was determined using a Student's t-test.
P≤0.05 was considered to indicate a statistically significant
difference.
Results
Caffeine does not induce apoptosis of CRC
cells and antagonizes paclitaxel-induced apoptosis
To study the effect of caffeine on
paclitaxel-induced apoptosis of CRC cells, Colo205 cells were first
treated with different doses of caffeine. Dose titration of
caffeine (0–20 μM) was performed and results indicated that
caffeine did not induce apoptosis of Colo205 cells (data not
shown). The combination effect of caffeine and paclitaxel against
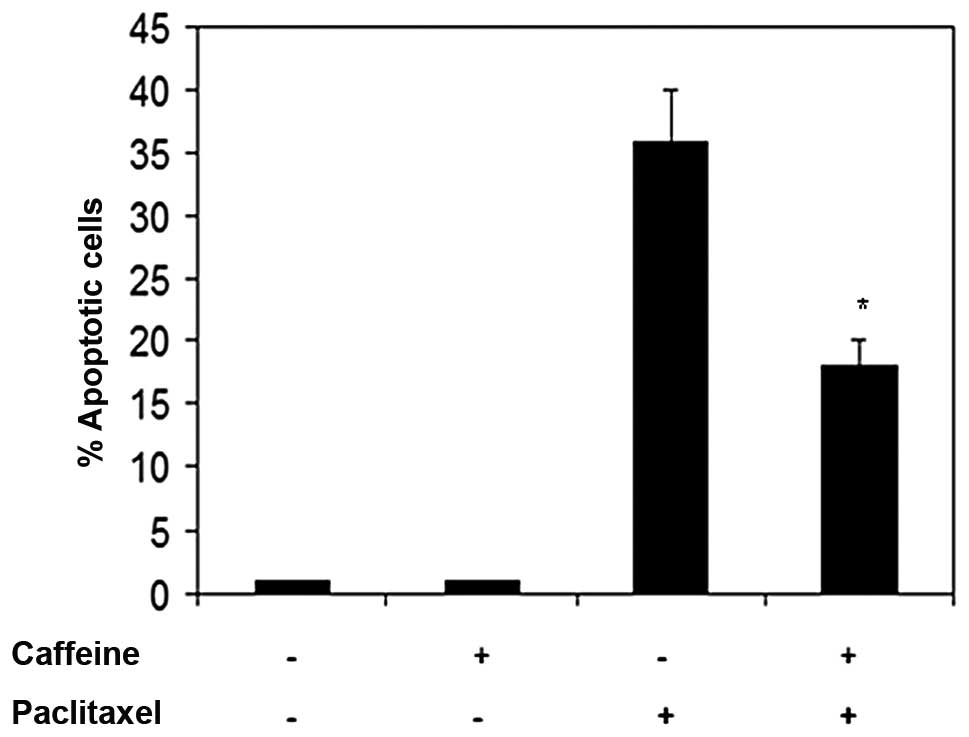
Colo205 cells was evaluated (Fig.
1). Results showed that pretreatment of Colo205 cells with
caffeine at 20 μM, prior to the addition of paclitaxel at 80 μM,
significantly inhibited paclitaxel-induced apoptosis.
Caffeine induces the MEK/ERK survival
pathway
A previous study showed that paclitaxel induces
activation of the MEK/ERK survival pathway (17). To gain a better understanding of
whether pretreatment with caffeine was responsible for antagonism
of paclitaxel-induced apoptosis by further increasing activation of
ERK1/2 pathway, Colo205 cells were pretreated with the chemical
inhibitor of MEK, U0126, prior to the addition of caffeine and
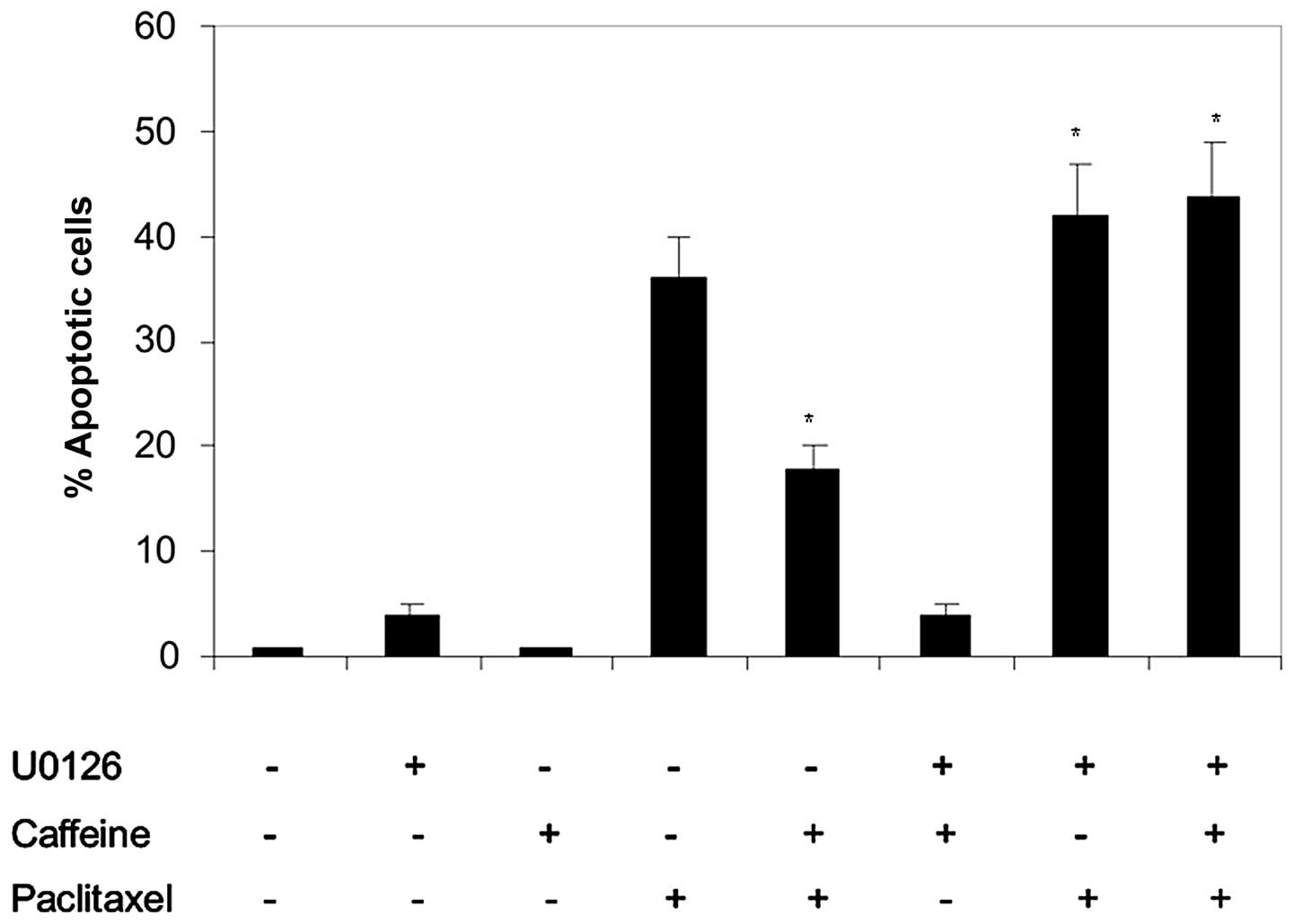
paclitaxel. Results in Fig. 2 show
that when the MEK-ERK pathway is inhibited, caffeine does not
reverse paclitaxel-induced cytotoxicity, indicating that such
antagonism may be mediated by activation of the MEK-ERK survival
pathway.
Caffeine induces Mcl-1 and GRP78 protein
levels but not survivin
To study whether caffeine-paclitaxel
combination-induced activation of MEK-ERK signaling may be
influencing other cell survival mediators, alterations in the
protein levels of Mcl-1, GRP78 and the inhibitor of apoptosis
protein, survivin, were evaluated in Colo205 cells following
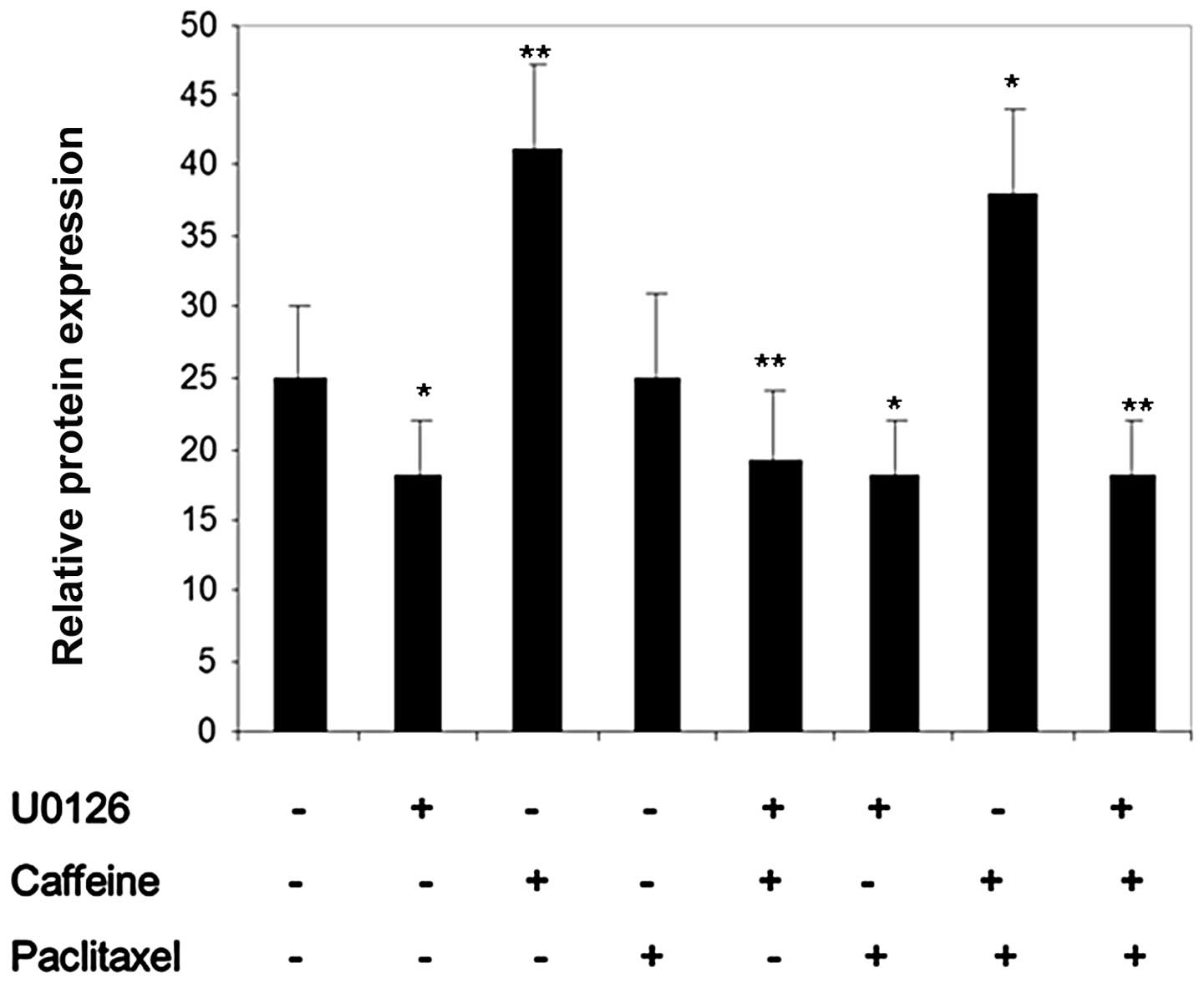
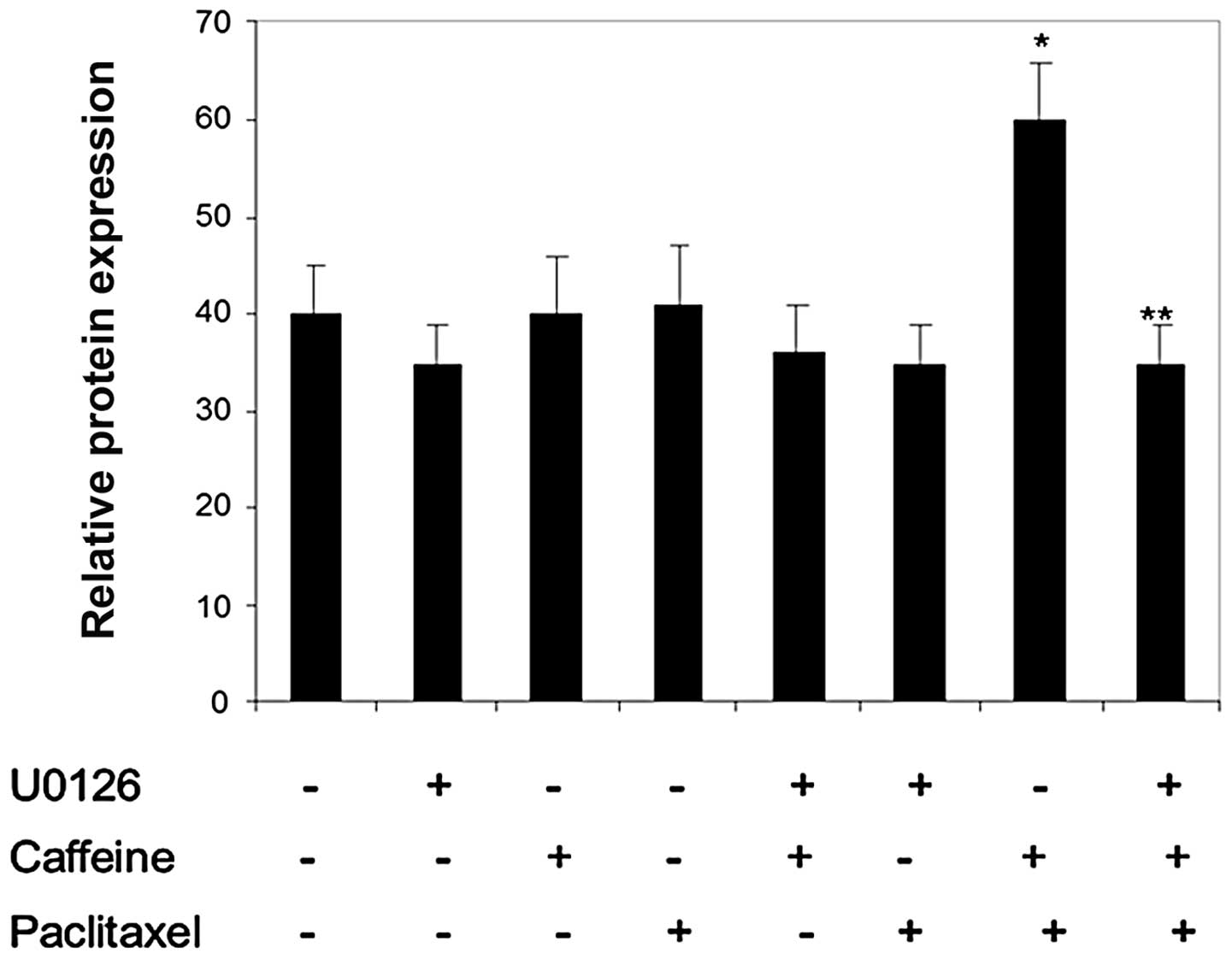
various treatments. Results in Figs.
3–5 showed that treatment of
Colo205 cells with caffeine induced levels of Mcl-1 and GRP78,
respectively, but not survivin. Treatment with paclitaxel induced
protein expression of GRP78 and surviving, however, this was not
observed to be significant. Levels of Mcl-1 were not affected by
paclitaxel treatment. Pretreatment of cells with caffeine followed
by paclitaxel resulted in a significant increase in all proteins,
particularly GRP78 and Mcl-1. This increase was almost completely
abrogated by pretreatment with the chemical inhibitor, U0126,
indicating that MEK-ERK signaling occurs upstream of caffeine and
paclitaxel-induced GRP78 and Mcl-1 upregulation.
Discussion
The aim of the present study was to evaluate the
effect of caffeine on the cytotoxicity induced by paclitaxel on CRC
cells. The aforementioned results reveal that sensitivity of CRC
cells to paclitaxel may be reversed with pretreatment of caffeine.
This is largely mediated by upregulation of the antiapoptotic
mediators Mcl-1 and GRP78, but not survivin.
It has been previously shown that paclitaxel induces
UPR in CRC as observed by upregulation of the UPR master marker
GRP78 and phosphorylation of eukaryotic initiation factor 2 α
subunit (33). The UPR is
essential for eukaryotic cell survival under ER stress and normal
growth conditions. To ensure that the ER protein-folding capacity
is not overwhelmed, the UPR is activated to remove the stress
condition forced on the ER and is organized by transcriptional
activation of a number of genes mediated by IRE1 and ATF6, a
general reduction in translation initiation and selective
translation of specific mRNAs mediated by PERK (34). However, if ER stress remains
unresolved, UPR may induce apoptotic cell death by activation of
various molecules, including caspases-4 and −12 and Bax/Bak
(35,36). Thus, the mechanism of
chemoresistance of CRC cells to paclitaxel-induced apoptosis may be
the activation of UPR.
Caffeine is known to differentially modify the
cytotoxicity of specific chemotherapeutic agents. This is
hypothesized to be due to DNA intercalation (37), inhibition of poly (ADP ribose)
polymerase (38) and cAMP
phosphodiesterase (39). Caffeine
was shown to increase the cytotoxic effect of cisplatin, a DNA
damaging agent, against lymphoma cells (40) and that of etoposide on HeLa
cervical carcinoma cells (41).
These effects were associated with the potential of caffeine to
induce cell cycle arrest, particularly at the G2/M phase, which was
followed by apoptosis (25,40,41).
In addition, caffeine has been shown to enhance paclitaxel-induced
apoptosis in human breast cancer MCF-7 cells (25). By contrast, caffeine inhibited
paclitaxel-induced apoptosis of lung cancer cells by suppression of
the effect of paclitaxel on cell cycle distribution (42). In agreement with this, the results
of the current study showed that caffeine antagonizes
paclitaxel-induced cytotoxicity in Colo205.
To elucidate the mechanism by which caffeine
prevents CRC cell sensitivity to paclitaxel, protein levels of
GRP78 were evaluated. Results indicated that caffeine induces
GRP78. These findings were consistent with previous studies,
indicating that treatment of condrocytes with caffeine induces ER
stress and increase intracellular calcium levels (43,44)
thus disturbing ER function and inducing UPR.
Furthermore, apoptosis mediated through the
mitochondrial pathway and ER has been shown to be regulated by the
Bcl-2 family proteins (45,46).
Antiapoptotic family members, including Bcl-XL, Bcl-2 and Mcl-1
protect mitochondrial integrity, while the proapoptotic proteins in
the family encourage the release of apoptogenic proteins, including
cytochrome c, Smac/DIABLO and AIF from the mitochondria and
calcium from the ER (45,46). Among the resistance mechanisms to
paclitaxel in several types of cancer, including melanoma, breast
and prostate cancers, is the upregulation of the antiapoptotic
Bcl-2 family protein Mcl-1 (47,48).
Results of this study have demonstrated that the treatment of
Colo205 cells with caffeine increases the protein levels of Mcl-1
indicating that caffeine contributes to paclitaxel-induced
apoptosis, at least in part, by induction of the Mcl-1 levels.
Another important class of proteins that is involved
in the regulation of apoptosis are the inhibitor of apoptosis
proteins (IAPs). This family includes IAP 1 and 2, XIAP, ML-IAP and
survivin. In general, they bind to caspases and inhibit their
activation (caspase-9) or prevent their effector action (caspases 3
and 7). Results of previous studies have shown that paclitaxel does
not induce the expression of survivin, however, the inhibition of
survivin may sensitize cancer cells to paclitaxel-induced
cytotoxicity (49). In agreement
with this, the present results have demonstrated that treatment of
Colo205 cells with either paclitaxel or caffeine did not induce
protein levels of survivin but treatment with a combination of
paclitaxel and caffeine has significantly increased survivin
levels. The cause of this result remains to be elucidated in future
studies.
Findings of previous studies have demonstrated that
members of Bcl-2 proteins and IAP may be transcriptional targets of
the MEK-ERK pathway (50,51). In the current study, caffeine
treatment resulted in the upregulation of Mcl-1, but not survivin.
Inhibition of the MEK-ERK pathway decreased the caffeine-induced
levels of Mcl-1. This finding may contribute to the role of
caffeine in protecting Colo205 cells from paclitaxel-induced
killing by stabilizing the mitochondrial and ER membranes via the
upregulation of Mcl-1 expression, preventing the release of
apoptotic molecules. This is hypothesized to occur by promoting the
binding of Mcl-1 to proapoptotic BH3-only members, including Bid,
Bim and Noxa (52–56).
In summary, the current observations indicated that
caffeine may prevent paclitaxel-induced apoptosis, most likely by
activation of the MEK-ERK signaling pathway. This activation has
led to an increase in levels of Mcl-1 and GRP78 that were capable
of protecting against the paclitaxel cytotoxic effect by
stabilizing mitochondria and ER membranes, thus inhibiting release
of apoptogenic molecules.
Acknowledgements
The authors would like to acknowledge the Jordan
University of Science and Technology, Irbid, Jordan, for financial
support (grant no. 230-2010).
References
|
1
|
Jemal A, Siegel R, Ward E, et al: Cancer
statistics, 2008. CA Cancer J Clin. 58:71–96. 2008. View Article : Google Scholar
|
|
2
|
Al-Tarawneh M, Khatib S and Arqub K:
Cancer incidence in Jordan, 1996–2005. East Mediterr Health J.
16:837–845. 2010.
|
|
3
|
Rowinsky EK and Donehower RC: Paclitaxel
(taxol). N Engl J Med. 332:1004–1014. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gonçalves A, Braguer D, Carles G, André N,
Prevôt C and Briand C: Caspase-8 activation independent of
CD95/CD95-L interaction during paclitaxel-induced apoptosis in
human colon cancer cells (HT29-D4). Biochem Pharmacol.
60:1579–1584. 2000.PubMed/NCBI
|
|
5
|
Bhalla KN: Microtubule-targeted anticancer
agents and apoptosis. Oncogene. 22:9075–9086. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Childs S, Yeh RL, Hui D and Ling V: Taxol
resistance mediated by transfection of the liver-specific sister
gene of P-glycoprotein. Cancer Res. 58:4160–4167. 1998.PubMed/NCBI
|
|
7
|
Lecureur V, Sun D, Hargrove P, et al:
Cloning and expression of murine sister of P-glycoprotein reveals a
more discriminating transporter than MDR1/P-glycoprotein. Mol
Pharmacol. 57:24–35. 2000.PubMed/NCBI
|
|
8
|
Subbaramaiah K, Hart JC, Norton L and
Dannenberg AJ: Microtubule-interfering agents stimulate the
transcription of cyclooxygenase-2. Evidence for involvement of
ERK1/2 and p38 mitogen-activated protein kinase pathways. J Biol
Chem. 275:14838–14845. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
McDaid HM and Horwitz SB: Selective
potentiation of paclitaxel (taxol)-induced cell death by
mitogen-activated protein kinase kinase inhibition in human cancer
cell lines. Mol Pharmacol. 60:290–301. 2001.PubMed/NCBI
|
|
10
|
Mhaidat NM, Thorne RF, Zhang XD and Hersey
P: Regulation of docetaxel-induced apoptosis of human melanoma
cells by different isoforms of protein kinase C. Mol Cancer Res.
5:1073–1081. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Troppmair J, Bruder JT, Munoz H, et al:
Mitogen-activated protein kinase/extracellular signal-regulated
protein kinase activation by oncogenes, serum, and
12-O-tetradecanoylphorbol-13-acetate requires Raf and is necessary
for transformation. J Biol Chem. 269:7030–7035. 1994.
|
|
12
|
Hommes DW, Peppelenbosch MP and van
Deventer SJ: Mitogen activated protein (MAP) kinase signal
transduction pathways and novel anti-inflammatory targets. Gut.
52:144–151. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Davis RJ: Signal transduction by the JNK
group of MAP kinases. Cell. 103:239–252. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Alessi DR, Saito Y, Campbell DG, et al:
Identification of the sites in MAP kinase kinase-1 phosphorylated
by p74raf-1. EMBO J. 13:1610–1619. 1994.PubMed/NCBI
|
|
15
|
Kavallaris M, Kuo DY, Burkhart CA, et al:
Taxol-resistant epithelial ovarian tumors are associated with
altered expression of specific beta-tubulin isotypes. J Clin
Invest. 100:1282–1293. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ranganathan S, Benetatos CA, Colarusso PJ,
Dexter DW and Hudes GR: Altered beta-tubulin isotype expression in
paclitaxel-resistant human prostate carcinoma cells. Br J Cancer.
77:562–566. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mhaidat NM, Thorne R, Zhang XD and Hersey
P: Involvement of endoplasmic reticulum stress in Docetaxel-induced
JNK-dependent apoptosis of human melanoma. Apoptosis. 13:1505–1512.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lee AS: GRP78 induction in cancer:
therapeutic and prognostic implications. Cancer Res. 67:3496–3499.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ma Y and Hendershot LM: ER chaperone
functions during normal and stress conditions. J Chem Neuroanat.
28:51–65. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wierzejska R: Caffeine - common ingredient
in a diet and its influence on human health. Rocz Panstw Zakl Hig.
63:141–147. 2012.(In Polish).
|
|
21
|
Tavares C and Sakata RK: Caffeine in the
treatment of pain. Rev Bras Anestesiol. 62:387–401. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Roehrs T and Roth T: Caffeine: sleep and
daytime sleepiness. Sleep Med Rev. 12:153–162. 2008. View Article : Google Scholar
|
|
23
|
Glade MJ: Caffeine-not just a stimulant.
Nutrition. 26:932–938. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Okano J, Nagahara T, Matsumoto K and
Murawaki Y: Caffeine inhibits the proliferation of liver cancer
cells and activates the MEK/ERK/EGFR signalling pathway. Basic Clin
Pharmacol Toxicol. 102:543–551. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Saunders DE, Lawrence WD, Christensen C,
Wappler NL, Ruan H and Deppe G: Paclitaxel-induced apoptosis in
MCF-7 breast-cancer cells. Int J Cancer. 70:214–220. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ribeiro JC, Barnetson AR, Jackson P, Ow K,
Links M and Russell PJ: Caffeine-increased radiosensitivity is not
dependent on a loss of G2/M arrest or apoptosis in bladder cancer
cell lines. Int J Radiat Biol. 75:481–492. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lu YP, Lou YR, Lin Y, et al: Inhibitory
effects of orally administered green tea, black tea, and caffeine
on skin carcinogenesis in mice previously treated with ultraviolet
B light (high-risk mice): relationship to decreased tissue fat.
Cancer Res. 61:5002–5009. 2001.PubMed/NCBI
|
|
28
|
Lu YP, Lou YR, Peng QY, Xie JG and Conney
AH: Stimulatory effect of topical application of caffeine on
UVB-induced apoptosis in the epidermis of p53 and Bax knockout
mice. Cancer Res. 64:5020–5027. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nomura M, Ichimatsu D, Moritani S, et al:
Inhibition of epidermal growth factor-induced cell transformation
and Akt activation by caffeine. Mol Carcinog. 44:67–76. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
He Z, Ma WY, Hashimoto T, Bode AM, Yang CS
and Dong Z: Induction of apoptosis by caffeine is mediated by the
p53, Bax, and caspase 3 pathways. Cancer Res. 63:4396–4401.
2003.PubMed/NCBI
|
|
31
|
Hashimoto T, He Z, Ma WY, et al: Caffeine
inhibits cell proliferation by G0/G1 phase arrest in JB6 cells.
Cancer Res. 64:3344–3349. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bode AM and Dong Z: The enigmatic effects
of caffeine in cell cycle and cancer. Cancer Lett. 247:26–39. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mhaidat NM, Alali FQ, Matalqah SM, et al:
Inhibition of MEK sensitizes paclitaxel-induced apoptosis of human
colorectal cancer cells by downregulation of GRP78. Anticancer
Drugs. 20:601–606. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang K and Kaufman RJ: Protein folding in
the endoplasmic reticulum and the unfolded protein response. Handb
Exp Pharmacol. 69–91. 2006. View Article : Google Scholar
|
|
35
|
Szegezdi E, Logue SE, Gorman AM and Samali
A: Mediators of endoplasmic reticulum stress-induced apoptosis.
EMBO Rep. 7:880–885. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tabas I and Ron D: Integrating the
mechanisms of apoptosis induced by endoplasmic reticulum stress.
Nat Cell Biol. 13:184–190. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lyles MB and Cameron IL: Caffeine and
other xanthines as cytochemical blockers and removers of
heterocyclic DNA intercalators from chromatin. Cell Biol Int.
26:145–154. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Geraets L, Moonen HJ, Wouters EF, Bast A
and Hageman GJ: Caffeine metabolites are inhibitors of the nuclear
enzyme poly(ADP-ribose)polymerase-1 at physiological
concentrations. Biochem Pharmacol. 72:902–910. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Howell LL: Comparative effects of caffeine
and selective phosphodiesterase inhibitors on respiration and
behavior in rhesus monkeys. J Pharmacol Exp Ther. 266:894–903.
1993.PubMed/NCBI
|
|
40
|
Shinomiya N, Shinomiya M, Wakiyama H,
Katsura Y and Rokutanda M: Enhancement of CDDP cytotoxicity by
caffeine is characterized by apoptotic cell death. Exp Cell Res.
210:236–242. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lock RB, Galperina OV, Feldhoff RC and
Rhodes LJ: Concentration-dependent differences in the mechanisms by
which caffeine potentiates etoposide cytotoxicity in HeLa cells.
Cancer Res. 54:4933–4939. 1994.PubMed/NCBI
|
|
42
|
Kitamoto Y, Sakurai H, Mitsuhashi N,
Akimoto T and Nakano T: Caffeine diminishes cytotoxic effects of
paclitaxel on a human lung adenocarcinoma cell line. Cancer Lett.
191:101–107. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhang PL, Lun M, Teng J, et al: Preinduced
molecular chaperones in the endoplasmic reticulum protect
cardiomyocytes from lethal injury. Ann Clin Lab Sci. 34:449–457.
2004.PubMed/NCBI
|
|
44
|
Ostergaard L, Simonsen U,
Eskildsen-Helmond Y, et al: Proteomics reveals lowering oxygen
alters cytoskeletal and endoplasmatic stress proteins in human
endothelial cells. Proteomics. 9:4457–4467. 2009. View Article : Google Scholar
|
|
45
|
Martinou JC and Green DR: Breaking the
mitochondrial barrier. Nat Rev Mol Cell Biol. 2:63–67. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Adams JM and Cory S: The Bcl-2 protein
family: arbiters of cell survival. Science. 281:1322–1326. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Ding Q, Huo L, Yang JY, et al:
Down-regulation of myeloid cell leukemia-1 through inhibiting
Erk/Pin 1 pathway by sorafenib facilitates chemosensitization in
breast cancer. Cancer Res. 68:6109–6117. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Wesarg E, Hoffarth S, Wiewrodt R, et al:
Targeting BCL-2 family proteins to overcome drug resistance in
non-small cell lung cancer. Int J Cancer. 121:2387–2394. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wang Z, Xie Y and Wang H: Changes in
survivin messenger RNA level during chemotherapy treatment in
ovarian cancer cells. Cancer Biol Ther. 4:716–719. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Boucher MJ, Morisset J, Vachon PH, Reed
JC, Lainé J and Rivard N: MEK/ERK signaling pathway regulates the
expression of Bcl-2, Bcl-X(L), and Mcl-1 and promotes survival of
human pancreatic cancer cells. J Cell Biochem. 79:355–369. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Hu P, Han Z, Couvillon AD and Exton JH:
Critical role of endogenous Akt/IAPs and MEK1/ERK pathways in
counteracting endoplasmic reticulum stress-induced cell death. J
Biol Chem. 279:49420–49429. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Wang YF, Jiang CC, Kiejda KA, Gillespie S,
Zhang XD and Hersey P: Apoptosis induction in human melanoma cells
by inhibition of MEK is caspase-independent and mediated by the
Bcl-2 family members PUMA, Bim, and Mcl-1. Clin Cancer Res.
13:4934–4942. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
O'Connor L, Strasser A, O'Reilly LA, et
al: Bim: a novel member of the Bcl-2 family that promotes
apoptosis. EMBO J. 17:384–395. 1998. View Article : Google Scholar
|
|
54
|
Cheng EH, Wei MC, Weiler S, et al: BCL-2,
BCL-X(L) sequester BH3 domain-only molecules preventing BAX- and
BAK-mediated mitochondrial apoptosis. Mol Cell. 8:705–711. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Korsmeyer SJ, Wei MC, Saito M, Weiler S,
Oh KJ and Schlesinger PH: Pro-apoptotic cascade activates BID,
which oligomerizes BAK or BAX into pores that result in the release
of cytochrome c. Cell Death Differ. 7:1166–1173. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Bouillet P, Purton JF, Godfrey DI, et al:
BH3-only Bcl-2 family member Bim is required for apoptosis of
autoreactive thymocytes. Nature. 415:922–926. 2002. View Article : Google Scholar : PubMed/NCBI
|