Introduction
Various cell types have been used as donors in
somatic cell nuclear transfer (SCNT) technology to produce cloned
animals from various species (1–4).
While previous studies have indicated that the origin of the donor
cell used in the cloning procedure is an important determinant of
the developmental competence of SCNT embryos (5,6),
there is limited information available with regard to the donor
cell effect on porcine cloning efficiency.
In addition, the efficiency of producing transgenic
cloned animals depends on improving the combination of technologies
involved, with the preparation and induction of competent
transfected donor cells being a crucial upstream step. However, the
genetic modification of donor cells requires long-term culture for
adequate cell line establishment, followed by the time required for
the transfection procedure. Therefore, the effects of apoptosis and
senescence stemming from long-term donor cell culture on SCNT
efficiency remain controversial (7,8) and
require further evaluation.
In order to address these issues, the present study
used four cell lines, two transgenic cell lines and the
corresponding non-transfected controls.
Following SCNT, donor cells transfected with a
tetracycline on/off responsive gene provide a tool to control the
expression of a specific gene at any time during development
(9). Therefore, in the present
study fibroblasts originating from crossbred Landrace x Yorkshire x
Duroc (LYD) pig fibroblasts were transfected with a tetracycline
responsive gene. In addition, since triple cell lines appear to be
resistant to hyperacute rejection during xenotransplantation
procedures (10), an additional
fibroblast cell line was used originating from Yucatan mini-pigs
and co-transfected with three genes, namely the complement
regulatory factors, human decay accelerating factor (hDAF) and
human (h) CD59, as well as H-transferase.
Therefore, the objective of the current study was to
understand the contribution of transgenic donor cell features to
variations in SCNT efficiency. For this purpose, various parameters
associated with viability and senescence were compared among four
fetal fibroblast cell lines, including morphology, growth rate,
mRNA expression and reactive oxygen species (ROS) production.
Materials and methods
Cell lines and culture procedure
Two fetal fibroblast cell lines were used as
controls in the study, originating from LYD or Yucatan mini pigs.
LYD cells were transfected with a tetracycline-responsive gene
(tetracycline on/off cells). Yucatan cells were transfected with
the complement regulatory factors, hDAF and hCD59, as well as
H-transferase (triple-transfected cells). Thus, each transfected
line was compared with its originating control (LYD and Yucatan).
Cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM;
Gibco-BRL, Grand Island, NY, USA) containing 25 μM glucose (high
glucose medium) plus 10% fetal bovine serum (FBS; Gibco-BRL). A 130
μg/ml hygromycin B solution from Streptomyces hygroscopious
(Sigma-Aldrich, St. Louis, MO, USA) was used for culturing the
triple transgenic cell line. Cell cultures were maintained in a
6-well dish and subcultured at a 1:4 ratio once ~90% confluence was
observed. The culture medium was replaced every two days. Assays
were always performed in cells from the 9th population doubling
(PD) passage.
Determination of PD times and cell
counts
Following the initial seeding, counting was
performed once cells reached 95% confluence. For this purpose,
cells were trypsinized, harvested and centrifuged (Hanshin Medical
Co., Seoul, Republic of Korea) at 2,000 rpm for 3 min. The pellet
was then suspended in DMEM plus 10% FBS and an aliquot was used for
counting with a hemocytometer (Superior, Marienfeld, Germany).
Cells were seeded again and the entire aforementioned procedure was
repeated twice. Thus, PD times were calculated by averaging the
time required for the cell number to double during the exponential
growth phase, that is, during the three cell subculture from
passage 7 to passage 9.
Morphological analysis
Phase contrast microscopy (Leica Microsystems,
Wetzlar, Germany) was used to assess cell morphology. Examinations
were performed on specimens fixed for 5 min in 4% paraformaldehyde
at room temperature.
Quantitative polymerase chain reaction
(qPCR)
Total RNA was extracted from the various cell lines
using TRIzol reagent (Invitrogen Life Technologies, Carlsbad, CA,
USA), according to the manufacturer’s instructions. Complementary
DNA (cDNA) was prepared by the reverse transcription of 1 μg total
RNA using Moloney murine leukemia virus reverse transcriptase
(Invitrogen Life Technologies) and random primers (9-mers; Takara
Bio, Inc., Shiga, Japan). qPCR (Mx3000P qPCR System; Agilent
Technologies, Inc., Santa Clara, USA) was then performed with 1 μl
cDNA template added to 10 μl 2X SYBR Premix Ex Taq (Takara
Bio, Inc.) containing specific primers. Amplification was performed
for 40 cycles with cycling parameters as follows: Denaturation at
95°C for 30 sec, annealing at 55°C for 30 sec and extension at 72°C
for 30 sec. All primer sequences are presented in Table I. The expression of each target
gene was quantified relative to that of the internal control gene,
GAPDH. The relative quantification was based upon the
comparison of the threshold cycle (Ct) at a constant fluorescence
intensity. Relative mRNA expression (R) was calculated using the
following equation: R = 2−[ΔCtsample − ΔCtcontrol]. To
determine the normalized arbitrary value for each gene, values were
normalized against GAPDH.
 | Table IPrimers used for gene expression
analysis. |
Table I
Primers used for gene expression
analysis.
| Gene | Primer sequences
(5′-3′) | Product size
(bp) | GenBank accession
number |
|---|
| Bax | F:
TGCCTCAGGATGCATCTACC
R: AAGTAGAAAAGCGCGACCAC | 199 | XM_003127290 |
| Bcl-2 | F:
AGGGCATTCAGTGACCTGAC
R: CGATCCGACTCACCAATACC | 193 | NM_214285 |
| p53 | F:
CTTTGAGGTGCGTGTTTGTG
R: CGGATCTGGAGGGTGAAATA | 192 | NM_213824 |
| GAPDH | F:
GTCGGTTGTGGATCTGACCT
R: TTGACGAAGTGGTCGTTGAG | 207 | NM_001206359 |
Measurement of ROS
To determine the intracellular content of ROS, cells
were incubated with 5 μg/ml 2′,7′-dichlorodihydrofluorescein
diacetate (H2DCF-DA; Invitrogen Life Technologies) for
30 min at 37°C. The acetates of H2DCF-DA were cleaved by
intracellular esterases, releasing the reduced fluorescein, which
was then converted to dichlorofluorescein in the presence of ROS
and detected by fluorescence activated cell sorting (FACS; Becton
Dickinson Immunocytometry Systems, San Jose, CA, USA). Dye-loaded
cells were trypsinized for 2 min and the reaction was stopped with
phosphate-buffered saline. Living cells were pelleted by
centrifugation at 357 × g for 3 min. Following washing with PBS,
cells were resuspended in PBS and FACS analysis was performed using
emission and excitation wavelengths of 530 and 480 nm,
respectively. The detected levels of DCF fluorescence correlated
with the ROS levels. Relative levels were compared between the
transfected cell lines and their control counterparts.
Statistical analysis
Quantitative measurements were performed at least
three times, with data expressed as the mean ± standard error of
the mean. Comparisons of mean values between various groups were
performed by one-way analysis of variance. P<0.05 was considered
to indicate a statistically significant difference.
Results
Cell proliferation rate
For cells in culture, the mean PD time was
correlated with the proliferation rate. Mean PD times were 26, 40,
26 and 66 h for the LYD, Yucatan, tetracycline on/off and triple
transfected cell lines, respectively. Thus, PD times were longer
for LYD than for Yucatan originated fibroblasts. Transgenic
tetracycline on/off cell lines had a similar mean PD time to
non-transfected LYD fibroblasts, whereas triple transfected cell
lines required longer (P<0.05) PD times than non-transfected
Yucatan controls.
Morphological analysis
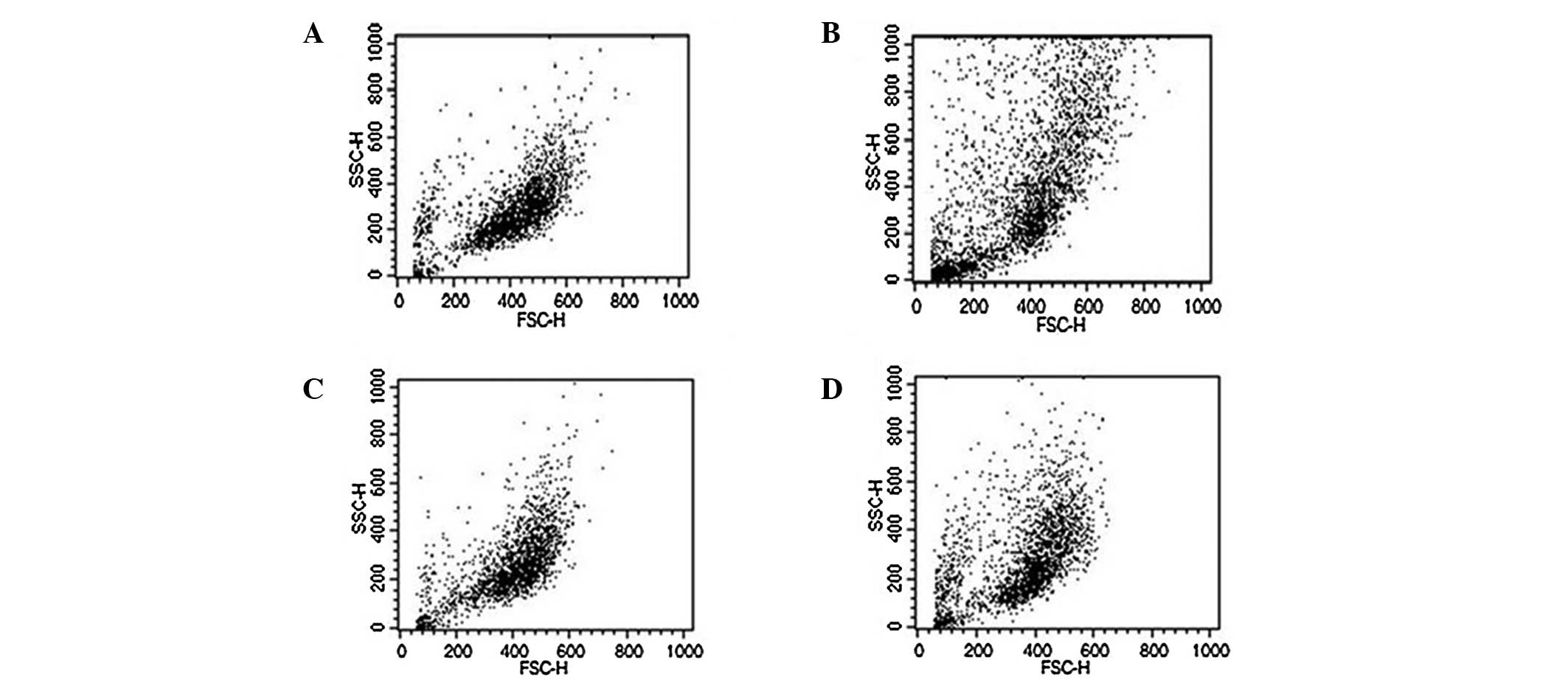
Based on the phase contrast microscopy observations,
fibroblasts from Yucatan mini-pigs were larger than those from LYD
crossbred pigs, which was consistent with the FACS analysis results
(Fig. 1). Conversely, there were
no morphological differences between the transfected and
corresponding non-transfected controls, with the exception of cell
density, as shown by the PD time calculation 3 days following
thawing (Figs. 1 and 2).
Gene-expression analysis
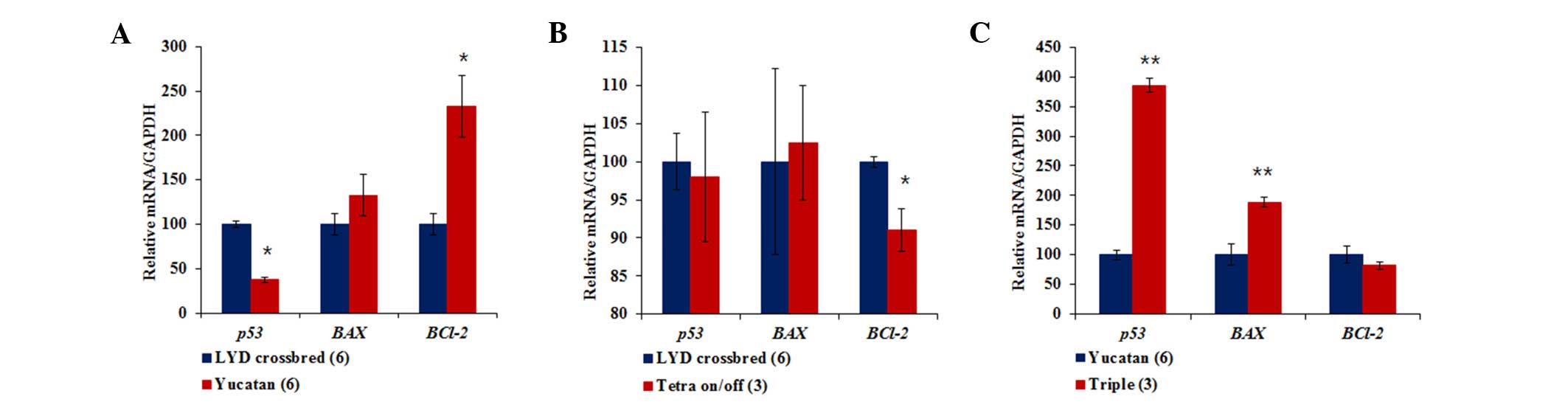
To assess cellular aging and apoptosis at the gene
level, the mRNA expression levels of TP53, Bax and
Bcl-2 were examined in the cell lines. Initially, Yucatan
fibroblasts had significantly lower levels of TP53 mRNA and
higher levels of Bcl-2 mRNA compared with LYD fibroblasts
(Fig. 3A). When comparing
tetracycline on/off transgenic cell lines with the original LYD
fibroblasts, the former expressed lower levels of Bcl-2
mRNA, with no differences in TP53 or Bax mRNA
expression levels (Fig. 3B). In
addition, triple transgenic cell lines exhibited a threefold higher
TP53 mRNA expression level and a twofold higher Bax
mRNA expression level than the non-transfected Yucatan fibroblasts
(Fig. 3C).
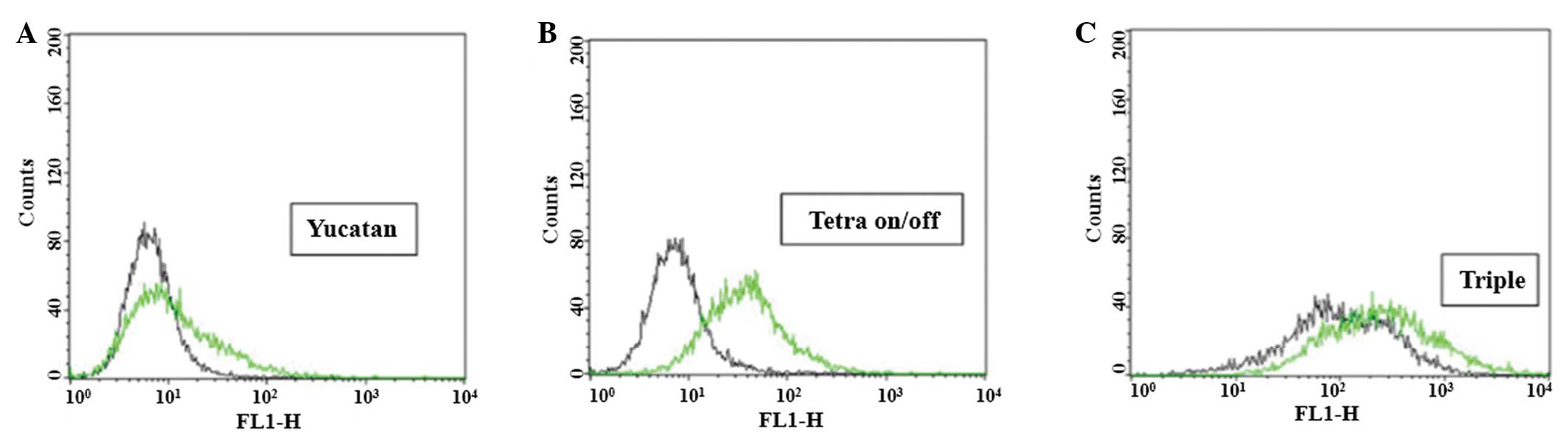
ROS production analysis
It has been reported that overproduction of ROS
initiates apoptosis (11).
Therefore, intracellular ROS levels were evaluated using a DCFH-DA
probe (Fig. 4) and FACS analysis
(Fig. 5). ROS levels were
significantly different depending on cell type and transfection
status. The histograms shown correspond to the representative
result from three independent experiments (Fig. 5). The rightward shift observed for
Yucatan fibroblasts is consistent with more extensive dye
oxidation, yielding a relative increase in fluorescence (Fig. 5A). In general, results were
consistent with the qualitative differences among cell lines and
hence indicated that the two transgenic cell lines exhibited higher
intracellular ROS levels when compared with their originating
non-transfected cell lines (Fig. 5B
and C).
Discussion
Survival rates of SCNT embryos are reportedly lower
in swine than in other domestic species (12). While a number of factors are likely
to contribute to the lower success rates, the main reasons for
consistent inefficiency of pig cloning remain unclear. A previous
study indicated that incomplete remodeling of the donor somatic
cell nucleus was a key factor contributing to the low success of
SCNT in swines (13). This was in
agreement with studies showing that the age, size, gender and type
of donor cell may affect the efficiency of SCNT (4,14–16).
Moreover, culturing donor cells for long time periods resulted in
decreased rates of in vitro embryos and to term development
in cattle (17). Altogether, these
studies indicate that nuclear donor cell preparation is a crucial
step affecting the developmental potential of reconstructed embryos
and the successful production of transgenic animals by SCNT. Thus,
the present study used four potential donor cell types, based on
the breed of origin and transgenic status, and analyzed parameters
associated with senescence and apoptosis.
The Bcl-2 family of proteins comprises
members with dual functions with regard to positively or negatively
regulating apoptosis. For example, Bcl-2 inhibits apoptosis
and proliferation, while Bax is pro-apoptotic and
proliferative (18). In addition,
p53, a protein involved in the regulation of cellular aging, is
postulated to inhibit the mammalian target of rapamycin and
suppress p21-induced senescence, thus causing quiescence (19). Thus, in the present study, relative
expression levels were measured for the aforementioned genes in
order to assess cell senescence and/or apoptosis, which may
negatively affect the potential of donor cells for SCNT purposes.
Notably, the Yucatan cell line exhibited lower expression levels of
TP53 mRNA and higher expression levels of Bcl-2 when
compared with other cell lines, which correlates with the PD times
observed. When compared with LYD fibroblasts, Yucatan cells
exhibited a 1.5-fold longer PD time. As for transgenic cells,
tetracycline on/off transfected cells exhibited a lower relative
expression of antiapoptotic Bcl-2 when compared with the
parental LYD cells, indicating that the transfected cells had a
diminished ability to avoid an apoptotic event. By comparison,
triple-transfected cell lines exhibited significantly higher
expression levels of TP53 and Bax when compared with
the parental Yucatan fibroblasts. It was therefore hypothesized
that this may enhance the probability of these cells undergoing
p53-dependent apoptosis, resulting in Bax-driven induction
of mitochondrial permeabilization (20). These observations indicate that
donor cell transfection may weaken the defense mechanisms against
apoptosis or even enforce pathways leading to programmed cell death
while in culture. For example, in a study by Li and Gao, CD59
transfected cells underwent caspase-3 activation and apoptosis
induction (21). While in this
study there was no effect on Bax expression levels, an
additional study showed that overexpression of CD59 inhibited
antiapoptotic Bcl-2 expression (22). These discrepancies are hypothesized
to be due to the differences in cell types; HeLa cells (cervical
cancer cells) and breast cancer cells were used in these two
studies.
At the cellular level, it has been reported that ROS
accelerates telomere shortening and induces senescence (23). Notably, ROS levels increase as
cells progress towards senescence through various pathways
(24,25). In the present study, a rightward
shift was observed in the FACS histogram for Yucatan fibroblasts
when compared with LYD cells, which is consistent with ROS
accumulation potentially due to aging. Thus, decreased antioxidant
capacity may account for a decrease in ROS scavenging, contributing
to aging-associated cell oxidative damage (26).
The p53 transcription factor is key in the signal
networks of senescence and ROS-induced apoptosis (27,28).
In the present study, increased ROS production was observed in
transfected cells, indicating that this procedure enhanced aging
and apoptosis. In addition, it was reported that p53-mediated
senescence (29) and apoptosis
(30) are enhanced in modified
cells. Thus, the significantly high production of ROS paired with
high expression levels of p53 and Bax in triple-transfected
cells markedly indicate that ROS-mediated activation of apoptosis
may potentially occur via a p53-driven pathway in this cell
line. Therefore, ROS may be acting as upstream factors activating
TP53, as well as downstream factors targeting p53-mediated
Bax activation. The interpretation of the correlation
between apoptosis and aging remains controversial. While several
studies are consistent with apoptosis being upregulated during
senescence in diverse cells, including cardiomyocytes (31), hepatocytes (32) and fibroblasts (33), other studies indicate that
oxidative stress does not induce apoptosis in senescent cells
(34) and that senescence may
attenuate apoptosis (35). The
different interpretations may have resulted from the variations in
species, strains or cell types used in the respective studies.
Questions remain with regard to assessing the
viability of transfected cell lines for use in SCNT. The effect of
transfected donor cells upon reconstructed SCNT embryos have been
previously reported (36). In the
present study, the average number of blastocyst cells was
significantly lower for the embryos reconstructed using transfected
fibroblasts.
In conclusion, the characteristics of individual
transgenic cell types and their associations with senescence and
apoptosis following transfection may be important determinants of
the developmental ability of SCNT embryos. However, further studies
are required to clarify the mechanisms involved.
Acknowledgements
This study was supported by the Basic Science
Research Program through the National Research Foundation of Korea,
funded by the Ministry of Education, Science and Technology (grant
no. 2012004885). The authors would like to thank the Sooam Biotech
Research Foundation for the donation of the cell lines (Yucatan
fibroblasts and triple-transfected cells).
Reference
|
1
|
Kato Y, Tani T, Sotomaru Y, Kurokawa K,
Kato J, Doguchi H, et al: Eight calves cloned from somatic cells of
a single adult. Science. 282:2095–2098. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lee BC, Kim MK, Jang G, Oh HJ, Yuda F, Kim
HJ, et al: Dogs cloned from adult somatic cells. Nature.
436:6412005. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Galli C, Lagutina I, Crotti G, Colleoni S,
Turini P, Ponderato N, et al: Pregnancy: a cloned horse born to its
dam twin. Nature. 424:6352003. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wakayama T and Yanagimachi R: Mouse
cloning with nucleus donor cells of different age and type. Mol
Reprod Dev. 58:376–383. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Oback B and Wells D: Practical aspects of
donor cell selection for nuclear cloning. Cloning Stem Cells.
4:169–174. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Batchelder CA, Hoffert KA, Bertolini M,
Moyer AL, Mason JB, Petkov SG, et al: Effect of the nuclear-donor
cell lineage, type, and cell donor on development of somatic cell
nuclear transfer embryos in cattle. Cloning Stem Cells. 7:238–254.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kubota C, Yamakuchi H, Todoroki J,
Mizoshita K, Tabara N, Barber M and Yang X: Six cloned calves
produced from adult fibroblast cells after long-term culture. Proc
Natl Acad Sci USA. 97:990–995. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Roh S, Shim H, Hwang WS and Yoon JT: In
vitro development of green fluorescent protein (GFP) transgenic
bovine embryos after nuclear transfer using different cell cycles
and passages of fetal fibroblasts. Reprod Fertil Dev. 12:1–6. 2000.
View Article : Google Scholar
|
|
9
|
Zhu Z, Zheng T, Lee CG, Homer RJ and Elias
JA: Tetracycline-controlled transcriptional regulation systems:
advances and application in transgenic animal modeling. Semin Cell
Dev Biol. 13:121–128. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ramírez P, Montoya MJ, Ríos A, García
Palenciano C, Majado M, Chávez R, et al: Prevention of hyperacute
rejection in a model of orthotopic liver xenotransplantation from
pig to baboon using polytransgenic pig livers (CD55, CD59, and
H-transferase). Transplant Proc. 37:4103–4106. 2005.PubMed/NCBI
|
|
11
|
Circu ML and Aw TY: Reactive oxygen
species, cellular redox systems, and apoptosis. Free Radic Biol
Med. 48:749–762. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pratt SL, Sherrer ES, Reeves DE and Stice
SL: Factors influencing the commercialisation of cloning in the
pork industry. Soc Reprod Fertil Suppl. 62:303–315. 2006.PubMed/NCBI
|
|
13
|
Zhao J, Whyte J and Prather RS: Effect of
epigenetic regulation during swine embryogenesis and on cloning by
nuclear transfer. Cell Tissue Res. 341:13–21. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yang F, Hao R, Kessler B, Brem G, Wolf E
and Zakhartchenko V: Rabbit somatic cell cloning: effects of donor
cell type, histone acetylation status and chimeric embryo
complementation. Reproduction. 133:219–230. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hosseini SM, Moulavi F, Foruzanfar M,
Hajian M, Abedi P, Rezazade-Valojerdi M, et al: Effect of donor
cell type and gender on the efficiency of in vitro sheep somatic
cell cloning. Small Rumin Res. 78:162–168. 2008. View Article : Google Scholar
|
|
16
|
Zhang YH, Song ES, Kim ES, Cong PQ, Lee
SH, Lee JW, et al: Effects of donor cell passage, size and type on
development of porcine embryos derived from somatic cell nuclear
transfer. Asian-Aust J Anim Sci. 22:194–200. 2009. View Article : Google Scholar
|
|
17
|
Hill JR, Winger QA, Long CR, Looney CR,
Thompson JA and Westhusin ME: Development rates of male bovine
nuclear transfer embryos derived from adult and fetal cells. Biol
Reprod. 62:1135–1140. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zinkel S, Gross A and Yang E: BCL2 family
in DNA damage and cell cycle control. Cell Death Differ.
13:1351–1359. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Leontieva OV and Blagosklonny MV: DNA
damaging agents and p53 do not cause senescence in quiescent cells,
while consecutive re-activation of mTOR is associated with
conversion to senescence. Aging (Albany NY). 2:924–935.
2010.PubMed/NCBI
|
|
20
|
Basu A and Haldar S: The relationship
between BcI2, Bax and p53: consequences for cell cycle progression
and cell death. Mol Hum Reprod. 4:1099–1109. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li XP and Gao MH: Effect of peptide seals
specific to CD59 on the expression of apoptosis-related genes in
HeLa cells. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi. 24:20–22. 2008.(In
Chinese).
|
|
22
|
Li B, Chu X, Gao M and Xu Y: The effects
of CD59 gene as a target gene on breast cancer cells. Cell Immunol.
272:61–70. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
von Zglinicki T: Oxidative stress shortens
telomeres. Trends Biochem Sci. 27:339–344. 2002.PubMed/NCBI
|
|
24
|
Passos JF and Von Zglinicki T: Oxygen free
radicals in cell senescence: are they signal transducers? Free
Radic Res. 40:1277–1283. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Terada LS: Specificity in reactive oxidant
signaling: think globally, act locally. J Cell Biol. 174:615–623.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lu CY, Lee HC, Fahn HJ and Wei YH:
Oxidative damage elicited by imbalance of free radical scavenging
enzymes is associated with large-scale mtDNA deletions in aging
human skin. Mutat Res. 423:11–21. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Vigneron A and Vousden KH: p53, ROS and
senescence in the control of aging. Aging (Albany NY). 2:471–474.
2010.PubMed/NCBI
|
|
28
|
Li PF, Dietz R and von Harsdorf R: p53
regulates mitochondrial membrane potential through reactive oxygen
species and induces cytochrome c-independent apoptosis blocked by
Bcl-2. EMBO J. 18:6027–6036. 1999. View Article : Google Scholar
|
|
29
|
Maier B, Gluba W, Bernier B, Turner T,
Mohammad K, Guise T, et al: Modulation of mammalian life span by
the short isoform of p53. Genes Dev. 18:306–319. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tyner SD, Venkatachalam S, Choi J, Jones
S, Ghebranious N, Igelmann H, et al: p53 mutant mice that display
early ageing-associated phenotypes. Nature. 415:45–53. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kajstura J, Cheng W, Sarangarajan R, Li P,
Li B, Nitahara JA, et al: Necrotic and apoptotic myocyte cell death
in the aging heart of Fischer 344 rats. Am J Physiol.
271:H1215–H1228. 1996.PubMed/NCBI
|
|
32
|
Muskhelishvili L, Hart RW, Turturro A and
James SJ: Age-related changes in the intrinsic rate of apoptosis in
livers of diet-restricted and ad libitum-fed B6C3F1 mice. Am J
Pathol. 147:20–24. 1995.PubMed/NCBI
|
|
33
|
Kujoth GC, Hiona A, Pugh TD, Someya S,
Panzer K, Wohlgemuth SE, et al: Mitochondrial DNA mutations,
oxidative stress, and apoptosis in mammalian aging. Science.
309:481–484. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gansauge S, Gansauge F, Gause H, Poch B,
Schoenberg MH and Beger HG: The induction of apoptosis in
proliferating human fibroblasts by oxygen radicals is associated
with a p53- and p21WAF1CIP1 induction. FEBS Lett. 404:6–10. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xiao ZQ, Moragoda L, Jaszewski R, Hatfield
JA, Fligiel SE and Majumdar AP: Aging is associated with increased
proliferation and decreased apoptosis in the colonic mucosa. Mech
Ageing Dev. 122:1849–1864. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kurome M, Ishikawa T, Tomii R, Ueno S,
Shimada A, Yazawa H and Nagashima H: Production of transgenic and
non-transgenic clones in miniature pigs by somatic cell nuclear
transfer. J Reprod Dev. 54:156–63. 2008. View Article : Google Scholar : PubMed/NCBI
|