Introduction
Ventricular arrhythmias (VAs) account for the
majority of cardiovascular-associated mortalities in patients with
ischemic heart disease. The mechanism of VAs is associated with gap
junction (GJ) remodeling involving connexin 43 (Cx43) reduction at
the intercalated disc (ID) and Cx43 lateralization (1). Previous studies have reported that
the dynamic movements of Cx43 are associated with microtubules and
their associated proteins (2,3).
Considering that microtubular disruption is an early cellular
reaction to hypoxia in cardiac myocytes (4,5), it
is presumed that preservation of the microtubules may affect GJ
functions and VAs during myocardial ischemia.
Taxol is a microtubular stabilizer that maintains
polymerized microtubular structure and ameliorates hypoxia-induced
myocyte injury (6).
Taxol-proliferated microtubules are resistant to cold,
Ca2+ and colchicines (7). Previous studies have revealed that
microtubule stabilizers preserve cardiac function and prevent VAs
in an isolated heart (8,9). However, the electrophysiological
consequences of in vivo microtubule preservation during
myocardial ischemia remains to be elucidated. In the present study,
an attempt was made to determine the electrophysiological effects
of taxol using an in vivo rat model with myocardial
ischemia-reperfusion (IR).
Materials and methods
Animal model and experimental
protocols
Experiments were approved by the Institutional
Animal Care and Use Committee of Wannan Medical College (Wuhu,
China). A total of 50 Sprague-Dawley rats (250 to 300 g) were
randomly assigned to five groups: Control group, IR group and three
taxol pretreatment groups. In the control and pretreatment groups,
normal saline and taxol (Sigma-Aldrich, St. Louis, MO, USA; 0.1,
0.3 and 0.9 μM·kg−1 in 0.5 ml saline) were injected
intraperitoneally (IP) 30 min prior to ischemia induction.
Each rat was anesthetized (pentobarbital sodium, 50
mg/kg IP; Sigma-Aldrich) and artificially ventilated. Once the
pericardium was opened, a 6.0 silk atraumatic suture (Jinghua Co.,
Shanghai, China) was passed around the left anterior descending
coronary artery at the level of the left atrial appendage. Ischemia
was induced for 20 min by ligating the suture, following which it
was confirmed by an elevated ST segment in the electrocardiogram
(ECG) and changes in myocardial color from red to pale. Reperfusion
was initiated by reopening the ligation for 20 min. The control
group was established by following the same procedure, but without
closing the suture.
ECG and monophasic action potentials
(MAPs)
ECG traces were recorded continuously, and signals
<10 Hz and >100 Hz were filtered out. Epicardial MAPs were
recorded at specific time points intermittently in normal sinus
rhythm (10). MAP signals were
amplified and recorded on a computer for subsequent analysis. A
commercially available physiological signals recording and analysis
system (RM6240; Chengdu Instrument Company, Chengdu, China) was
used to digitalize, store and analyze MAP signals. The software was
used to analyze action potential durations (APD90) at
90% of repolarization. Dispersion of action potential duration
(APDd) represented the absolute value of the time difference
between the maximum and minimum APD.
Analysis of VAs
VAs were analyzed according to Lambeth conventions
(11). The score system (9) used to quantify arrhythmias was as
follows: 0, <10 ventricular premature contractions (VPCs); 1,
≥10 VPCs; 2, 1–5 episodes of ventricular tachycardia (VT); 3, >5
episodes of VT or 1 episode of ventricular fibrillation (VF); 4,
2–5 episodes of VF; 5, >5 episodes of VF; 6, VT or VF, or the
two together with total combined duration ≤300 sec; 7, VT or VF, or
the two together with total combined duration >300 sec.
Tubulin polymerization assay
A tubulin polymerization assay was performed using a
previously described method (6).
Myocardial tissue from the ischemic area (1 g) was minced and
homogenized in different buffers. For the total tubulin fraction,
the myocardium was homogenized in 1% SDS buffer containing 10 mM
Tris·HCl (pH 7.4), 0.5 mM dithiothreitol and 1 mM
Na3VO4. The mixture was boiled for 5 min and
centrifuged at 16,000 × g at 4°C for 10 min and then the
supernatant was saved as the total protein fraction. For the free
tubulin heterodimer and polymerized tubulin (microtubule)
fractions, the myocardium was homogenized in a microtubule
stabilization buffer containing 50% glycerol, 5% dimethyl
sulfoxide, 10 mM Na2HPO4, 0.5 mM ethylene
glycol tetraacetic acid and 0.5 mM MgSO4 and then
centrifuged at 100,000 × g at 25°C for 20 min. The supernatant was
saved as the tubulin heterodimer fraction, whereas the pellet was
saved as the tubulin microtubule fraction. For immunoblotting,
rabbit polyconal α-tubulin (1:2,000; ab125267; Abcam, Cambridge,
MA, USA) antibody was used to detect the free tubulin heterodimer
and polymerized tubulin.
Western blot analysis
Protein (100 μg) was denatured by heating at 95°C
for 5 min prior to resolution by SDS-PAGE and transferring to a
polyvinylidene difluoride membrane (Shanghai Threebio Technology
Co., Ltd, Shanghai, China). The membrane was blocked in
phosphate-buffered saline (Shanghai Threebio Technology Co., Ltd)
containing 0.2% Tween-20 (Shanghai Threebio Technology Co., Ltd)
and 5% skimmed milk for 2 h at 37°C and incubated overnight at 4°C
with primary polyclonal antibodies (rabbit anti-Cx43; 1:2,000;
ab11370; Abcam). The housekeeping protein,
glyceraldehyde-3-phosphate dehydrogenase (rabbit anti-GAPDH
polyclonal antibody; ab9485; Abcam) was used as a loading control.
Antibody binding was detected using horseradish
peroxidase-conjugated secondary antibody (1:2,000; Sigma-Aldrich)
and visualized using an enhanced chemiluminescence kit (Chemicon,
Temecula, CA, USA).
Immunofluorescence staining
Frozen tissue sections fixed in cold acetone (−20°C
for 5 min) were used for immunofluorescence staining. Nonspecific
antibody binding sites were blocked with 5% fetal bovine serum for
60 min. Tissue sections were labeled for 16 h at 4°C with rabbit
polyclonal anti-Cx43 (Cx43; 1:200, Abcam). Anti-rabbit
immunoglobulin G conjugated with fluorescein isothiocyanate was
used for detection and images were observed using a Zeiss confocal
microscope (LSM-510; Carl Zeiss, Gottingen, Germany).
Statistical analysis
Statistical analysis was performed using SPSS 16.0
software (SPSS, Inc., Chicago, IL, USA). Data are expressed as the
mean ± standard error of the mean and were analyzed using analysis
of variance followed by Newman-Keul’s t-test. P<0.05 was
considered to indicate a statistically significant difference.
Results
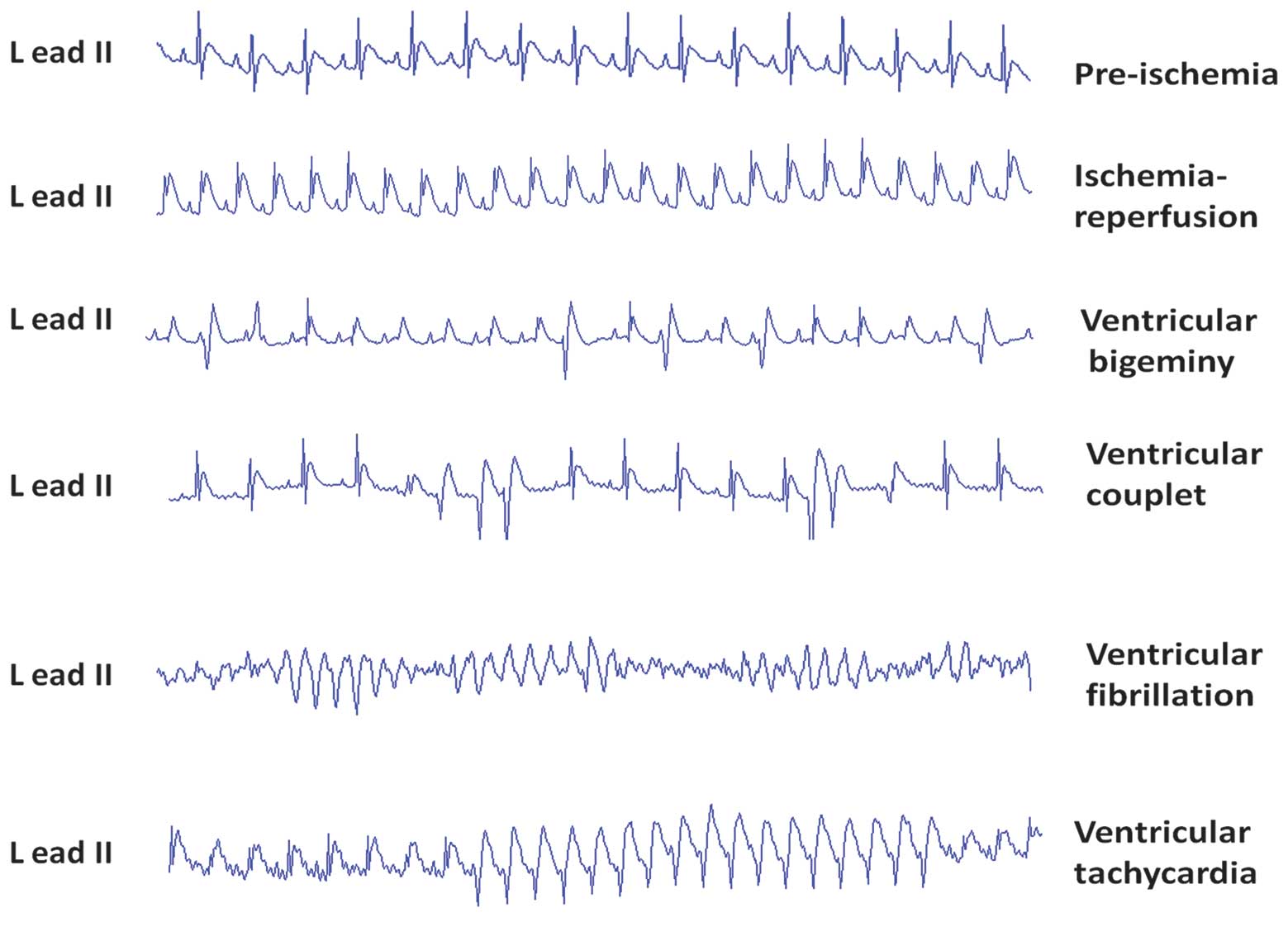
Effects of taxol on IR-induced VAs
In a preliminary experiment, it was observed that
>2 μM taxol may lead to lethal ventricular bradycardias. Thus,
the doses of taxol were reduced to 0.1, 0.3 and 0.9 μM. In the
control group, 6/10 rats had ≥10 VPCs and only 1 had an episode of
VT. In the IR group, all animals had >5 episodes of VT or
several episodes of VF. The spontaneous VTs/VFs appeared most
frequently (5 to 10 min) after ligation and then recovered
gradually. Within 2 min after reperfusion, VPCs/VTs/VFs occurred
transiently and automatically recovered rapidly. In three taxol
treatment groups, fewer animals had VTs and VFs than in the IR
group. However, no difference in VAs during reperfusion periods
between the IR group and taxol-treated groups was observed
(Fig. 1; Table I). The severity of VAs was
significantly decreased in taxol-treated groups in a dose-dependent
manner compared with the IR group based on the VA score system
(P<0.01; Fig. 1).
 | Table IEffects of taxol on
ischemia/reperfusion-induced ventricular arrhythmias. |
Table I
Effects of taxol on
ischemia/reperfusion-induced ventricular arrhythmias.
| Ischemia | Reperfusion | |
|---|
|
|
| |
|---|
| Group | VTs | VFs | VPCs | VTs | VFs | VPCs | VA Score |
|---|
| Control | 1/10 | 0 | 7/10 | - | - | - | 0.8±0.63 |
| IR | 10/10a | 6/10a | 10/10 | 8/10 | 3/10 | 10/10 | 3.3±0.95a |
| IR + 0.1 μM
taxol | 10/10a | 4/10a | 10/10 | 7/10 | 3/10 | 10/10 | 2.7±0.48ab |
| IR + 0.3 μM
taxol | 10/10a | 3/10ab | 10/10 | 6/10 | 2/10 | 10/10 | 2.5±0.53ab |
| IR + 0.9 μM
taxol | 7/10a | 0/10ab | 10/10 | 6/10 | 2/10 | 10/10 | 1.7±0.45ab |
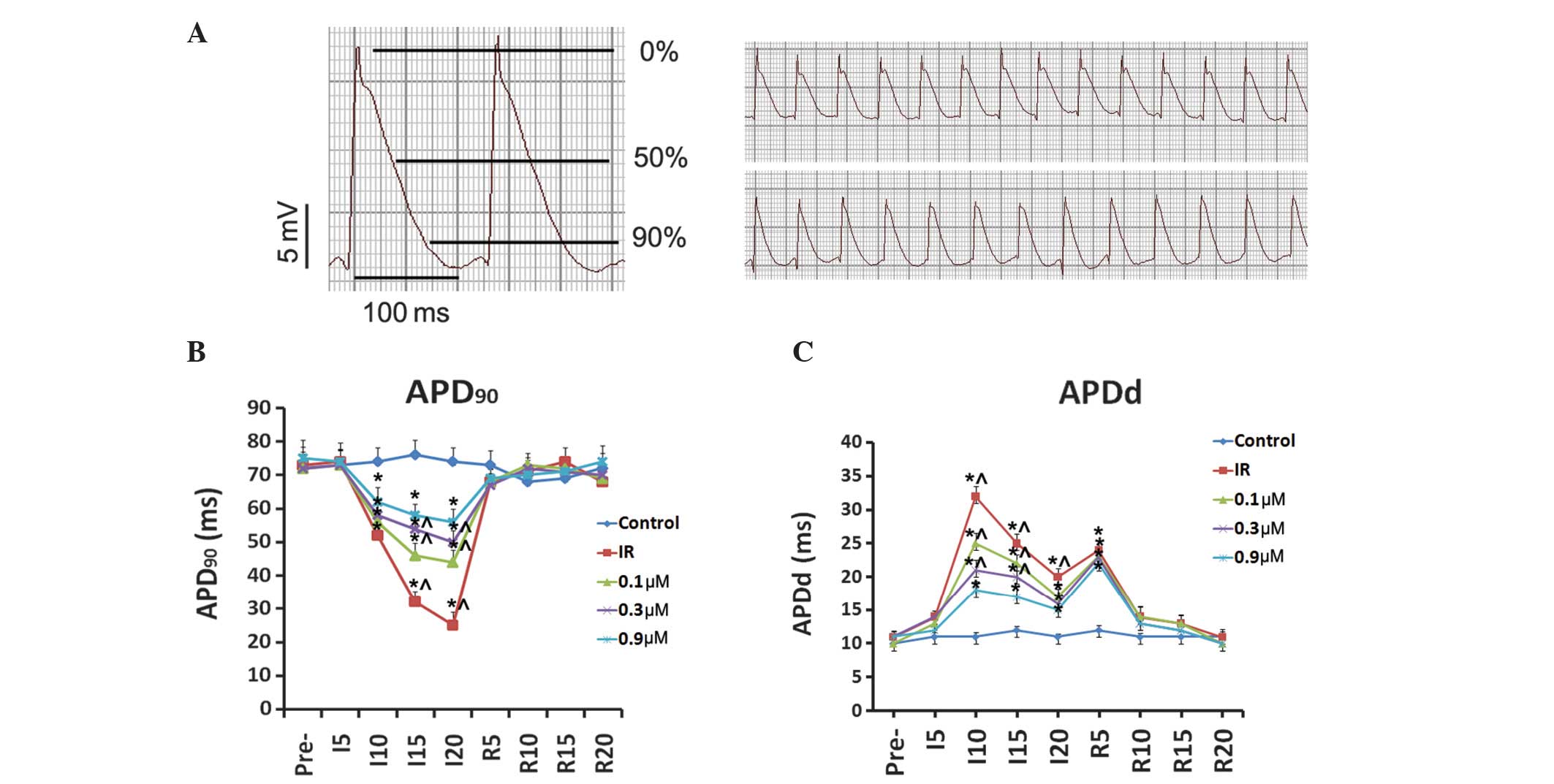
Effects of taxol on IR-induced MAP
changes
The MAP duration at APD90 of the ischemic
epicardium in the IR group was significantly shorter than that of
the control group (Fig. 2). When
perfusion was restored, the APD90 rapidly recovered
within 5 min and returned to baseline levels. The APDd was
significantly increased in the IR group compared with that in the
control group. Taxol treatment partially but significantly restored
APD90 and reduced APDd during ischemia in the
pretreatment groups. However, no significant difference in
APD90 was observed during the reperfusion periods
between the IR group and taxol-treated groups.
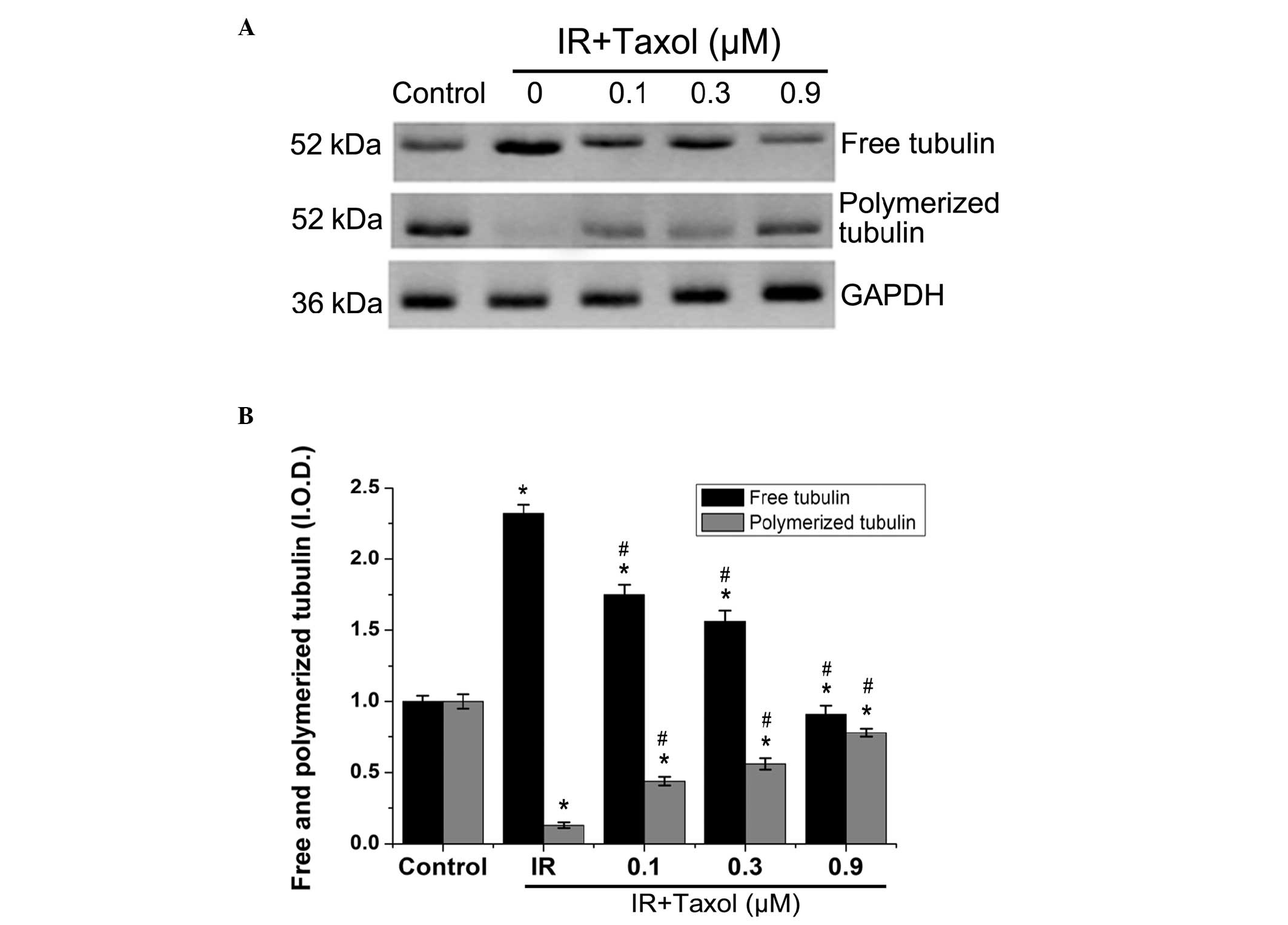
Effects of taxol on IR-induced
microtubular depolymerization
The relative levels of free tubulin in the IR group
increased significantly after the heart was exposed to IR compared
with the control group (Fig. 3).
In addition, polymerized tubulin in the IR group significantly
decreased. Taxol treatment maintained tubulin polymerization
significantly and reduced free tubulin in a dose-dependent manner
(P<0.01).
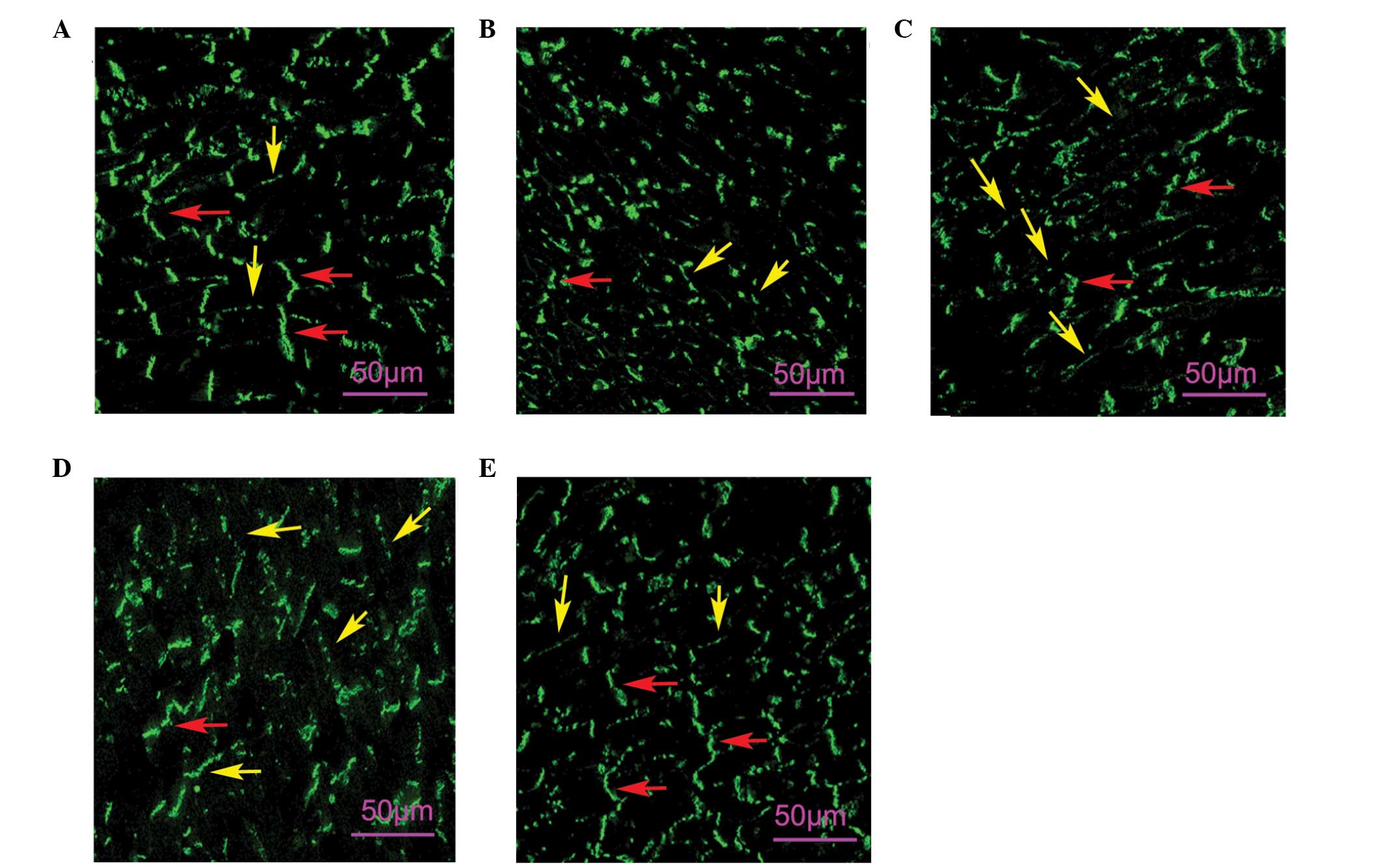
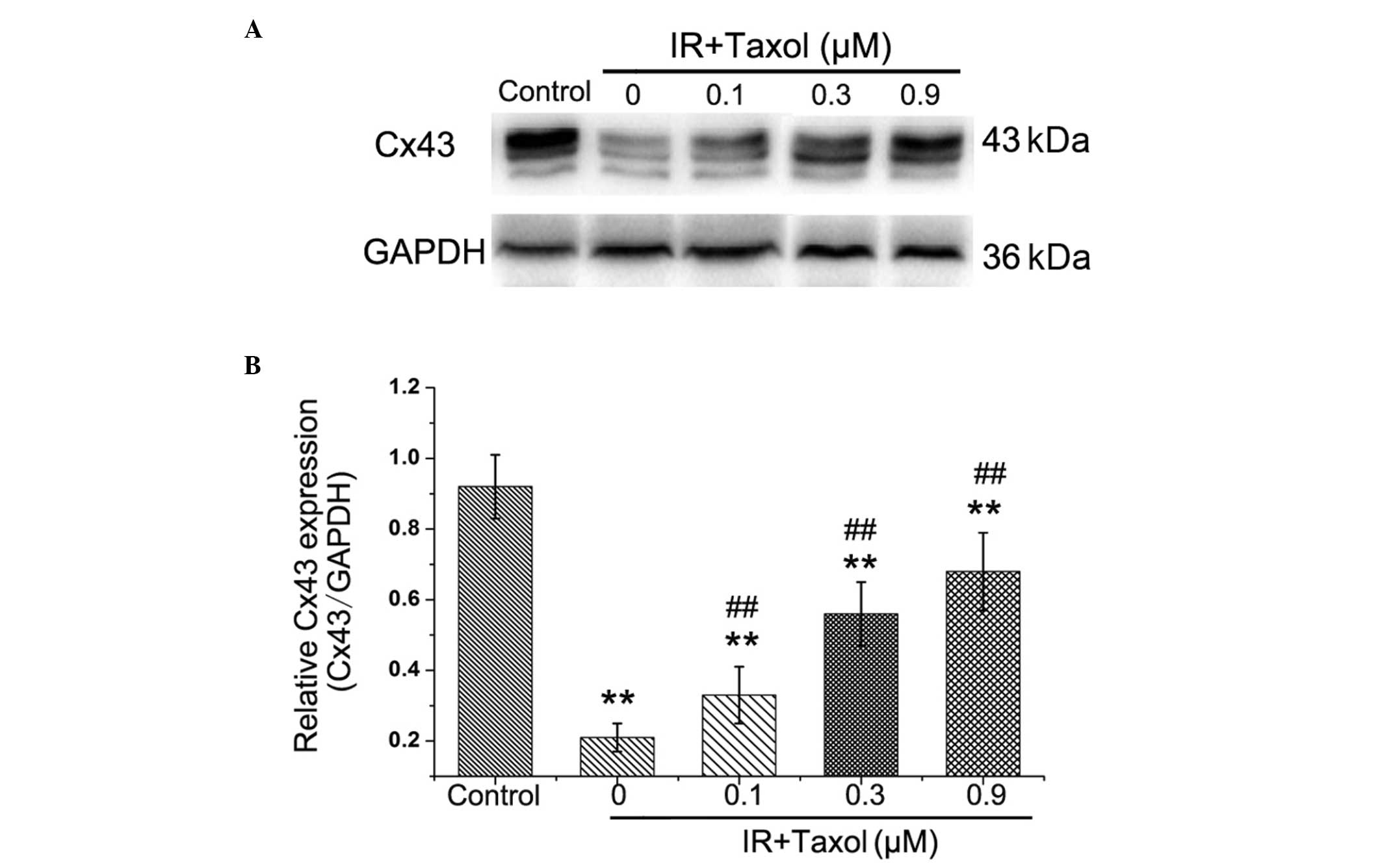
Effects of taxol on IR-induced Cx43
distribution in the heart
The Cx43 staining signal in the non-ischemic heart
was concentrated at the IDs (red arrows, Fig. 4A), whereas little Cx43 was located
in the lateral cardiomyocyte surfaces (yellow arrows, Fig. 4A). In the hearts of the IR group,
the Cx43 signal was markedly reduced in ID and redistributed to the
lateral cardiomyocyte surfaces (Fig.
4B). The hearts subjected to taxol pretreatment exhibited
significantly improved Cx43 distribution compared with the IR
groups (Fig. 4C, D and E).
Effects of taxol on IR-induced Cx43
expression
The relative levels of Cx43 protein expression in
myocardial tissue were reduced significantly in the IR group
compared with the control group (Fig.
5). Taxol treatment significantly improved Cx43 expression in a
dose-dependent manner (P<0.01).
Discussion
Microtubules are important components of the
cytoskeleton. They have important functions in protein synthesis,
intracellular trafficking and intracellular signaling. During these
processes, microtubules require a dynamic balance between free
tubulin and polymerized dimers in the cytoplasm (12,13).
Previous studies have demonstrated that microtubules are
responsible for regulating Ca2+ channels, APD and
membrane potential (14). In the
present study, it was observed that susceptibility to VAs increased
in an in vivo rat model with IR, however the susceptibility
was also partly but significantly reduced by taxol pretreatment in
a dose-dependent manner. The present results are in agreement with
previous ex vivo experiments (8,9),
indicating that stabilizing microtubules have significant
anti-arrhythmic effects during myocardial IR in vivo.
Previous studies have revealed that during
myocardial ischemia, either APD shortening or APDd enlargement
generates pro-arrhythmic substrates (15,16).
APD duration undergoes a temporary prolongation and then gradually
recovers following reperfusion (17). Considering that microtubules
regulate APD and membrane potential (14), taxol may also affect APD duration
and APDd during myocardial IR. In the present study,
APD90 shortened rapidly following coronary occlusion,
however was restored within 5 min after reperfusion. Taxol
treatment significantly ameliorated ischemia-induced APD shortening
and APDd, which can reduce ischemia-induced arrhythmias. Thus the
beneficial effects of taxol on ischemia-induced arrhythmias may be
due to the improvement in APD changes. In cultured myocytes,
microtubular depolymerization is an early cellular reaction to
hypoxia (4). Acute hypoxia leads
to the collapse of microtubular networks and reversible cellular
damage (18). The interruption of
microtubules occurs prior to mitochondrial damage and myocyte
injury (19,20). Taxol may stabilize polymerized
microtubules and ameliorate hypoxia-induced myocyte injury
(6). The present data demonstrated
that taxol treatment significantly preserved polymerized tubulin
and reduced free tubulin during IR.
GJ remodeling is associated with cardiac arrhythmias
during myocardial ischemia. Previous studies have revealed that
oxidative stress can affect microtubules and lead to subsequent
perturbation of Cx43 delivery to the plasma membrane in the human
and mouse myocardium (2,3). In the present study, it was observed
that in the normal ventricular tissue, Cx43 was localized at IDs.
It was also identified that taxol pretreatment significantly
improved Cx43 distribution and expression in the ischemic
myocardial tissue. The stabilization of Cx43 during ischemia may be
associated with reduction of VAs through taxol pretreatment.
The present study has several limitations. The
phosphorylation levels of Cx43 that may modulate GJ function during
ischemia were not detected (21).
In addition, the present study was not able to exclude the
possibility that GJ preservation of taxol was secondary to the
reduction of oxidative stress, which can reduce the levels of
connexin passing to the plasma membrane (22). Taxol may also be able to modulate
ion channels (14). Therefore,
further studies should focus on identifying the ion channels
preserved by taxol during ischemic injury.
The present study demonstrated that taxol
pretreatment may stabilize microtubules, preserve GJs and reduce
the severity of VAs during myocardial IR. These findings indicated
that microtubule stabilization may be of benefit in modifying the
VA susceptibility of ischemic hearts.
Acknowledgements
The present study was supported by grants from the
National Natural Science Foundation of Anhui Province (grant nos.
1208085QH156 and 1208085MH129) and the National Natural Science
Foundation of China (grant no. 81200142).
References
|
1
|
Kieken F, Mutsaers N, Dolmatova E, et al:
Structural and molecular mechanisms of gap junction remodeling in
epicardial border zone myocytes following myocardial infarction.
Circ Res. 104:1103–1112. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Shaw RM, Fay AJ, Puthenveedu MA, von
Zastrow M, Jan YN and Jan LY: Microtubule plus-end-tracking
proteins target gap junctions directly from the cell interior to
adherens junctions. Cell. 128:547–560. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Smyth JW, Hong TT, Gao D, et al: Limited
forward trafficking of connexin 43 reduces cell-cell coupling in
stressed human and mouse myocardium. J Clin Invest. 120:266–279.
2010. View
Article : Google Scholar :
|
|
4
|
Vandroux D, Schaeffer C, Tissier C, et al:
Microtubule alteration is an early cellular reaction to the
metabolic challenge in ischemic cardiomyocytes. Mol Cell Biochem.
258:99–108. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Devillard L, Vandroux D, Tissier C, et al:
Involvement of microtubules in the tolerance of cardiomyocytes to
cold ischemia-reperfusion. Mol Cell Biochem. 307:149–157. 2008.
View Article : Google Scholar
|
|
6
|
Teng M, Dang YM, Zhang JP, et al:
Microtubular stability affects cardiomyocyte glycolysis by
HIF-1alpha expression and endonuclear aggregation during early
stages of hypoxia. Am J Physiol Heart Circ Physiol.
298:H1919–H1931. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Howarth FC, Calaghan SC, Boyett MR and
White E: Effect of the microtubule polymerizing agent taxol on
contraction, Ca2+ transient and L-type Ca2+
current in rat ventricular myocytes. J Physiol. 516(Pt 2): 409–419.
1999. View Article : Google Scholar
|
|
8
|
Xiao J, Zhao H, Liang D, et al: Taxol, a
microtubule stabilizer, improves cardiac contractile function
during ischemia in vitro. Pharmacology. 85:301–310. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xiao J, Cao H, Liang D, et al: Taxol, a
microtubule stabilizer, prevents ischemic ventricular arrhythmias
in rats. J Cell Mol Med. 15:1166–1176. 2011. View Article : Google Scholar
|
|
10
|
Franz MR: Current status of monophasic
action potential recording: theories, measurements and
interpretations. Cardiovasc Res. 41:25–40. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Walker MJ, Curtis MJ, Hearse DJ, et al:
The Lambeth Conventions: guidelines for the study of arrhythmias in
ischaemia infarction, and reperfusion. Cardiovasc Res. 22:447–455.
1988. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kawakami K, Chiba T, Katagiri N, et al:
Paclitaxel increases high voltage-dependent calcium channel current
in dorsal root ganglion neurons of the rat. J Pharmacol Sci.
120:187–195. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
De Vuyst E, Boengler K, Antoons G, Sipido
KR, Schulz R and Leybaert L: Pharmacological modulation of
connexin-formed channels in cardiac pathophysiology. Br J
Pharmacol. 163:469–483. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Calaghan SC, Le Guennec JY and White E:
Cytoskeletal modulation of electrical and mechanical activity in
cardiac myocytes. Prog Biophys Mol Biol. 84:29–59. 2004. View Article : Google Scholar
|
|
15
|
Tsuburaya R, Yasuda S, Ito Y, et al:
Eicosapentaenoic acid reduces ischemic ventricular fibrillation via
altering monophasic action potential in pigs. J Mol Cell Cardiol.
51:329–336. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen Y, Zhang Q, Liao YH, et al: Effect of
tumor necrosis factor-alpha on neutralization of ventricular
fibrillation in rats with acute myocardial infarction. Mediators
Inflamm. 2011:5652382011. View Article : Google Scholar
|
|
17
|
Guo X, Gao X, Wang Y, Peng L, Zhu Y and
Wang S: IKs protects from ventricular arrhythmia during cardiac
ischemia and reperfusion in rabbits by preserving the
repolarization reserve. PLoS One. 7:e315452012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hori M, Sato H, Kitakaze M, et al:
Beta-adrenergic stimulation disassembles microtubules in neonatal
rat cultured cardiomyocytes through intracellular Ca2+
overload. Circ Res. 75:324–334. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Saetersdal T, Greve G and Dalen H:
Associations between beta-tubulin and mitochondria in adult
isolated heart myocytes as shown by immunofluorescence and
immunoelectron microscopy. Histochemistry. 95:1–10. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Devillard L, Vandroux D, Tissier C, et al:
Tubulin ligands suggest a microtubule-NADPH oxidase relationship in
postischemic cardiomyocytes. Eur J Pharmacol. 548:64–73. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Beardslee MA, Lerner DL, Tadros PN, et al:
Dephosphorylation and intracellular redistribution of ventricular
connexin43 during electrical uncoupling induced by ischemia. Circ
Res. 87:656–662. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Smyth JW, Hong TT, Gao D, et al: Limited
forward trafficking of connexin 43 reduces cell-cell coupling in
stressed human and mouse myocardium. J Clin Invest. 120:266–279.
2010. View
Article : Google Scholar :
|