Introduction
Medulloblastoma, an embryonic neuroepithelial tumor
of the cerebellum, is the most common type of malignant brain tumor
in children and among childhood central nervous system tumors,
medulloblastoma has high cancer-associated mortality (1). Medulloblastoma arises from mutated
remaining primitive neuroectoderm cells in the ventricle and grows
in the cerebellar vermis, regularly invading through the ependyma
and brainstem (2). Children with
medulloblastoma are generally treated with surgery followed by
chemotherapy and radiation, a combination of treatments with
five-year survival rates as high as 70–80% (3), but which also induces marked toxicity
and adverse cognitive effects that substantially impact the quality
of life of the patient.
Medulloblastoma consist of four predominant
subgroups with distinct molecular characteristics: Group 1 tumors
exhibit mutations in the sonic hedgehog (SHH) gene and its protein
receptors; group 2 tumors are induced by changes in WNT signaling,
generally through the main responsive gene, β-catenin; group 3
tumors occur due to alterations in transforming growth
factor-β-orthodenticle homeobox 2 signaling; and group 4 tumors
arise from MYC and MYCN mutations (4–7). For
example, SHH signaling pathway mutations are predominantly
associated with nodular/desmoplastic and anaplastic
medulloblastomas, which presumably arise from granule neuron
precursor cells (8). Tumors with
activating mutations in the WNT signaling pathway commonly present
as the classic medulloblastoma histological subtype, which
contributes to 7–15% of medulloblastoma cases (9). However, the majority of oncogenic
factors of this heterogeneous cancer remain unknown. Therapies that
target key oncogenic promoters, such as SHH, have not been
successful, underscoring the importance of identifying alternative
targets.
Small nuclear ribonucleoprotein-associated
polypeptide N (SNRPN), located on chromosome bands 15q11-13, is
imprinted and paternally expressed in somatic tissues and has
comparable expression levels in all regions of the brain (10). SNRPN, which is a member of the
small nuclear ribonucleic particle SMB/SMN family, is involved in
pre-mRNA processing, possibly through tissue-specific alternative
splicing events. The SNRPN expression pattern is to a certain
extent controlled by methylation in the 5′-untranslated region of
exon 1 on the maternally derived chromosome. SNRPN was the first
expressed gene identified in the Prader-Willi syndrome (PWS)
critically deleted region (10).
Previous studies have predominantly focused on SNRPN gene
methylation. For example, various studies have demonstrated that
SNRPN imprinting may be involved in the regulation of multiple
types of cancer, including germ cell tumors (GCTs) (11), acute myeloid leukemia (12) and human uterine leiomyomas
(13). SNRPN methylation patterns
in germ cell tumors have been reported to reflect primordial germ
cell development (11). The
variable methylation of SNRPN also supports the hypothesis that
intracranial GCTs are associated with neural stem cells (14). In addition, complete maintenance of
SNRPN imprinting has been observed in human uterine leiomyomas
(13). Analysis of the data from
eight papillary thyroid carcinoma (PTC) tumor samples revealed that
amplifications of SNRPN occurred solely in tumors with a wild type
B-type Raf kinase (BRAF) (15).
All the aforementioned results indicate that SNRPN may be critical
in tumorigenesis. In the present study, to analyze the role of
SNRPN in medulloblastoma, the effect of SNRPN on cell growth was
investigated in vitro using the Daoy human medulloblastoma
cell line.
Materials and methods
Cell culture
The Daoy and D283Med human medulloblastoma cell
lines were obtained from the Cell Bank of Chinese Academy of
Sciences (Shanghai, China). The two types of cells were maintained
in Eagle’s minimum essential medium (EMEM) (Sigma-Aldrich, St.
Louis, MO, USA) containing 1% L-Glu, supplemented with 10%
heat-inactivated fetal bovine serum (FBS) (Sigma-Aldrich) at 37°C
in a 5% CO2 humidified atmosphere.
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA of the cultured cells was extracted using
TRIzol® solution (Invitrogen, Carlsbad, CA, USA). RNA
quality was assessed with a Bioanalyzer instrument (Agilent
Technologies, Palo Alto, CA, USA). cDNA was immediately
reverse-transcribed from the isolated RNA using the SuperScript III
First-Strand Synthesis system (Invitrogen), and was subsequently
used to amplify SNRPN by qPCR using Ex-Taq DNA polymerase (Takara
Bio, Inc., Shiga, Japan). Subsequent qPCR amplification was
analyzed using the Bio-Rad Connect real-time PCR platform (Bio-Rad
Laboratories, Hercules, CA, USA) and was performed using 2 μg cDNA
with the following conditions: initial denaturation at 95°C for 1
min, followed by 40 cycles of denaturation at 95°C for 5 sec and
annealing extension at 60°C for 20 sec. The absorbance value was
read at the extension stage. β-actin served as the input reference.
The primers used were as follows: SNRPN forward,
5′-GTTTTGGGTCTGGTGTTGCT-3′ and reverse, 5′-TCATTACCTGCTGGGATGGT-3′;
β-actin, forward, 5′-GTGGACATCCGCAAAGAC-3′ and reverse,
5′-AAAGGGTGTAACGCAACTA-3′. The relative mRNA expression levels were
determined using the following formula: 2−ΔCT [cycle
threshold (CT)], where ΔCT = CT (target gene) − CT (β-actin).
Construction of SNRPN short hairpin
(sh)RNA-expressing lentivirus (Lv)
To produce the SNRPN shRNA-expressing cell lines, an
shRNA (5′-AATCTTCATTGGCACCTTTACTCGAGTAAAGGTGCCAATGAAGATTCTTTTT-3′)
targeting the human SNRPN gene (NCBI accession number, NM_003097)
was inserted into a pFH-L plasmid (Shanghai Hollybio, Shanghai,
China). A scrambled siRNA sequence (5′-TTCTCCGAACGTGTCACGT-3′) with
no homology to the mammalian genome served as a control (Con). The
Lv-based shRNA-expressing vectors were constructed, verified by DNA
sequencing, and were designated pFH-L-shSNRPN and pFH-L-shCon. For
the transfection, Daoy cells at a density of 5×104
cells/well were seeded in six-well plates and cultured for 72 h to
reach 90% confluence. At 2 h prior to transfection, the medium was
replaced with serum-free EMEM. The plasmid mixture that contained
pFH-L-shSNRPN (or pFH-L-shCon) and pVSVG-I/pCMVΔR8.92 packaging
vectors, as well as Lipofectamine 2000 (Invitrogen), was added to
the Daoy cells. After 5 h incubation, the medium was replaced with
EMEM containing 10% FBS. At 48 h after transfection, lentiviral
particles (Lv-shSNRPN or Lv-shCon) were harvested and purified by
ultra-centrifugation, according to methods described in previous
studies (16,17). At 72 h after infection, the viral
titer was determined by counting the number of green fluorescence
protein (GFP)-expressing cells under fluorescence microscopy, as
described in a previous study (18).
Western blot analysis
Daoy and D283Med cell lysates were prepared with 2X
sodium dodecyl sulfate (SDS) sample buffer containing 100 mM
Tris-HCl (pH 6.8), 10 mM ethylenediaminetetraacetic acid, 4% SDS
and 10% glycine. The homogenate was subsequently centrifuged at
12,000 × g for 15 min at 4°C and the supernatant was collected and
preserved at −80°C. A bicinchoninic acid kit (Pierce, Rockford, IL,
USA) was used to determine protein concentration. Protein lysates
were electrophoresed on 10% SDS-PAGE gels and then transferred to
nitrocellulose membranes using a semi-dry electrotransferring unit
(Bio-Rad). The membranes were incubated in 5% non-fat dry milk in
tris-buffered saline and Tween® (TBST)buffer containing
10 mM Tris-HCl, pH 8.0, 150 mM NaCl and 0.1% Tween-20 for 2 h at
room temperature. Subsequently, the membranes were blotted with the
following primary antibodies: Rabbit anti-SNRPN (1:1,000 dilution;
#HPA003482; Sigma-Aldrich) or mouse anti-GAPDH (1:1,000 dilution;
#sc-32233; Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA)
overnight at 4°C. Following three washes with TBST (5 min each),
the blots were incubated with the corresponding horseradish
peroxidase-conjugated secondary antibodies: Goat anti-rabbit
immunoglobulin G (IgG; 1:5,000 dilution; Santa Cruz Biotechnology,
Inc.) and goat anti-mouse IgG (1:5,000 dilution; Santa Cruz
Biotechnology, Inc.) for 2 h at room temperature. Immunoreactivity
was detected using enhanced chemoluminescent autoradiography
(Amersham, Piscataway, NJ, USA).
MTT assay
Cell proliferation was determined using a MTT
colorimetric assay. The MTT assay measures the conversion of MTT to
insoluble formazan by the dehydrogenase enzymes of the intact
mitochondria of living cells. Subsequent to infection for four
days, the Daoy cells were washed with phosphate-buffered saline
(PBS), trypsinized with EMEM containing 10% FBS, seeded into a
96-well plate at a concentration of 2.5×103 cells/well
and incubated at 37°C. The number of viable cells was measured at
daily intervals on days 1–5. At each time-point, 10 μl MTT reagent
(Sigma-Aldrich; 5 mg/ml in PBS) was added to the cultured cells and
the mixtures were incubated for 4 h at 37°C. At the end of the
incubation period, the medium was carefully removed and 100 μl
acidified isopropanol containing 10% SDS, 5% isopropanol and 0.01
mol/l HCl was added. The reaction product was quantified by
measuring the absorbance at 595 nm using an enzyme-linked
immunosorbent assay plate reader (Bio-Rad Laboratories). The
experiments were repeated three times.
Colony formation assay
In vitro tumorigenicity was determined on the
basis of cell growth in a plate colony formation assay. Daoy cells
(500 cells/well) receiving one of three treatments (Con, Lv-shCon
or Lv-shSNRPN) were seeded in six-well plates. After six days
incubation, the cells were washed with PBS buffer and stained with
0.5% crystal violet in 20% methanol for 20 min. Following crystal
purple staining, images of visible colonies were captured and the
number of colonies (>50 cells/colony) was counted using Colony
Counter software (Image-Pro® Plus version 6.0, Media
Cybernetics, Inc., Rockville, MD, USA). The morphology and size of
the colonies was examined under a microscope (Olympus Corporation,
Tokyo, Japan).
Fluorescence-activated cell sorting
analysis
The Daoy cells were collected five days after
infection with Lv-shSNRPN or Lv-shCon, and seeded in 6 cm dishes at
2×105 cells/dish. The cells were then cultured for a
further 40 h and subsequently collected. Following washes with
ice-cold PBS, the cells were suspended in ~0.5 ml 70% cold alcohol
and maintained at 4°C for 30 min. Subsequently, the cells were
treated with 100 mg/ml DNase-free RNase (Sigma-Aldrich) and
incubated for 30 min at 37°C. Propidium iodide (50 mg/ml;
Sigma-Aldrich) was added immediately to the cell suspension. Prior
to analysis, the suspension was filtered through a 50 mm nylon mesh
and a total of 1×104 stained cells were counted by a
flow cytometer (FACSCalibur; BD Biosciences, Franklin Lakes, NJ,
USA).
Statistical analysis
Data were analyzed using GraphPad Prism software
version 6.00 for Windows (GraphPad Software, Inc., La Jolla, CA,
USA). Values are expressed as the mean ± standard deviation of
three independent experiments. The statistical significance of the
differences between groups was determined by a repeated-measures
analysis of variance test. P<0.05 was considered to indicate a
statistically significant difference.
Results
SNRPN expression profile in
medulloblastoma cell lines
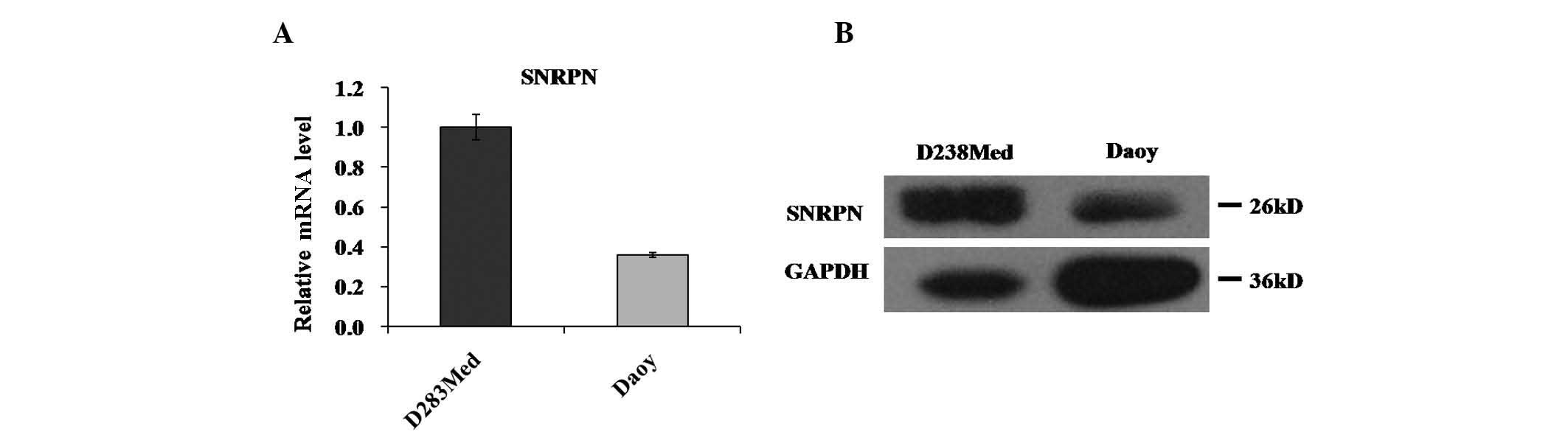
To investigate the role of SNRPN in human
medulloblastoma, the expression levels of SNRPN in human
medulloblastoma cell lines were assessed. The SNRPN mRNA and
protein levels in the two most widely used medulloblastoma cell
lines, Daoy and D283Med, were measured. SNRPN mRNA was observed to
be highly expressed in the two cell lines, as determined by RT-qPCR
(Fig. 1A). Western blot analysis
revealed similar results for SNRPN protein, which was highly
expressed in the Daoy and D283Med cells (Fig. 1B).
Knockdown of SNRPN by infection with
Lv-shSNRPN in medulloblastoma cells
In order to investigate the role of SNRPN in
medulloblastoma, Daoy cells were transfected with Lv that stably
expressed SNRPN-specific siRNA (Lv-shSNRPN) and reporter gene GFP.
To detect whether the recombinant Lvs successfully infected Daoy
cells, GFP signals were observed with a fluorescence microscope
(x10 objective lens). The rate of positive enhanced (e)GFP
expression in cells remained at >80% subsequent to Lv-shSNRPN
and Lv-shCon transfection, which indicated that this lentiviral
vector-based RNAi system was successfully established (Fig. 2A).
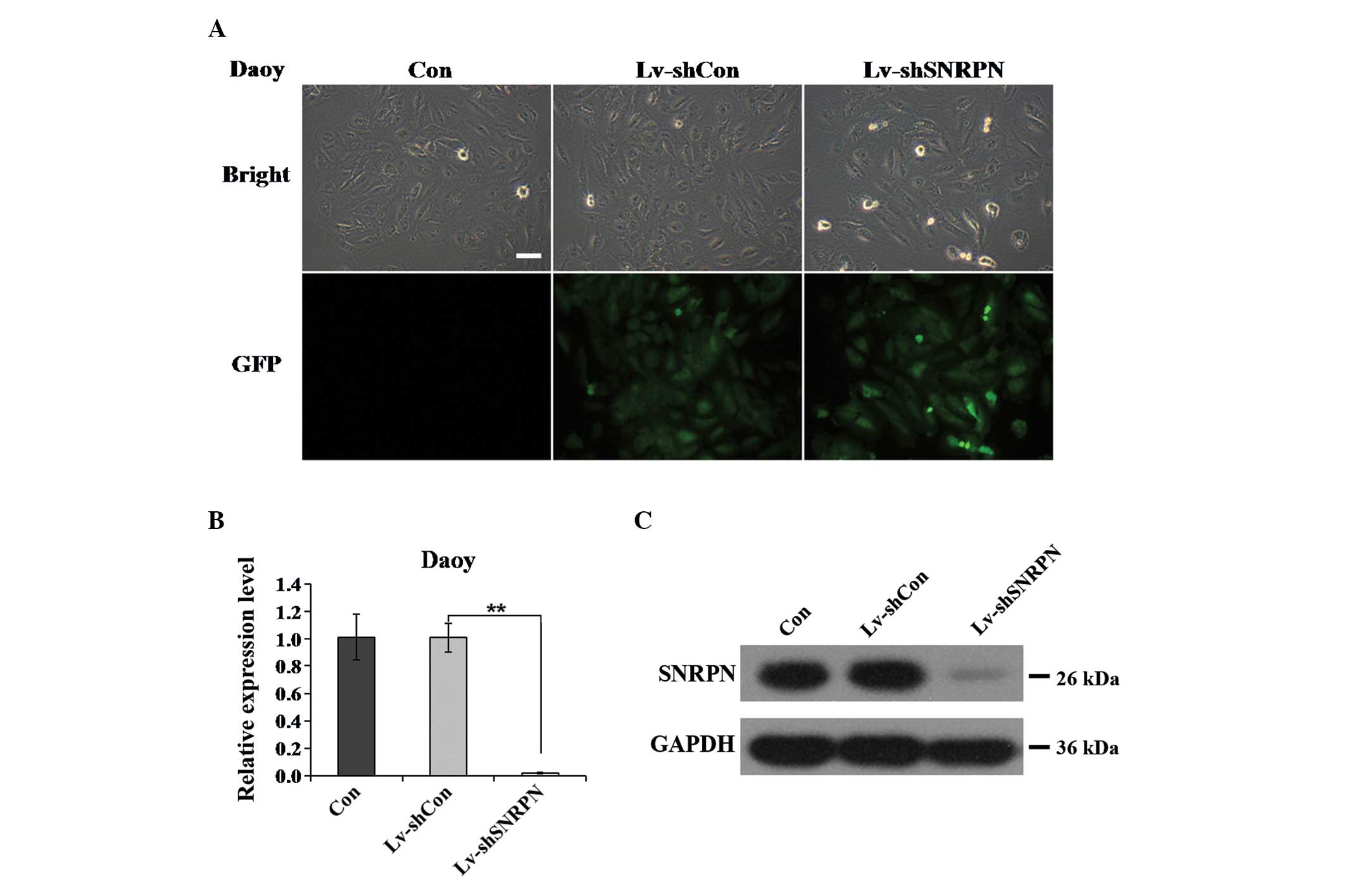 | Figure 2Lv-mediated silencing of SNRPN in Daoy
cells. (A) Determination of infection efficiency in Daoy cells.
Representative images of Daoy cells after five days of Lv infection
are shown (scale bar, 50 μm). (B and C) Expression analysis of
SNRPN mRNA and protein levels in Daoy cells following one of three
treatments (Con, Lv-shCon or Lv-shSNRPN) by (B) RT-qPCR and (C)
western blotting. β-actin gene and GAPDH protein served as internal
controls for RT-qPCR and western blotting, respectively.
(**P<0.01). Lv, lentivirus; SNRPN, small nuclear
ribonucleoprotein-associated polypeptide N; RT-qPCR, reverse
transcription-polymerase chain reaction; Con, control, sh, short
hairpin; GFP, green fluorescence protein. |
To further verify that the SNRPN gene was
successfully knocked down by Lv-shSNRPN, the expression levels of
SNRPN mRNA in the Daoy cells were assessed using RT-qPCR. After
three days lentiviral infection, the SNRPN mRNA expression levels
in the Daoy cells infected with Lv-shSNRPN were significantly
reduced, as compared with the uninfected and Lv-shCon-infected
cells (P<0.01; Fig. 2B). The
SNRPN knockdown efficiency was satisfactory, with a 98.1% reduction
in the Daoy cells infected with Lv-shSNRPN. To confirm the
silencing of SNRPN protein, western blotting was performed in the
Daoy cells using antibodies against SNRPN. Only a weak band was
observed in the Lv-shSNRPN group, although no significant
differences in the SNRPN expression levels between the Con group
and the Lv-shCon group were identified (Fig. 2C). Therefore, the constructed
Lv-shSNRPN effectively knocked down SNRPN expression in the Daoy
cells.
Effect of SNRPN knockdown on
medulloblastoma cell proliferation and colony formation
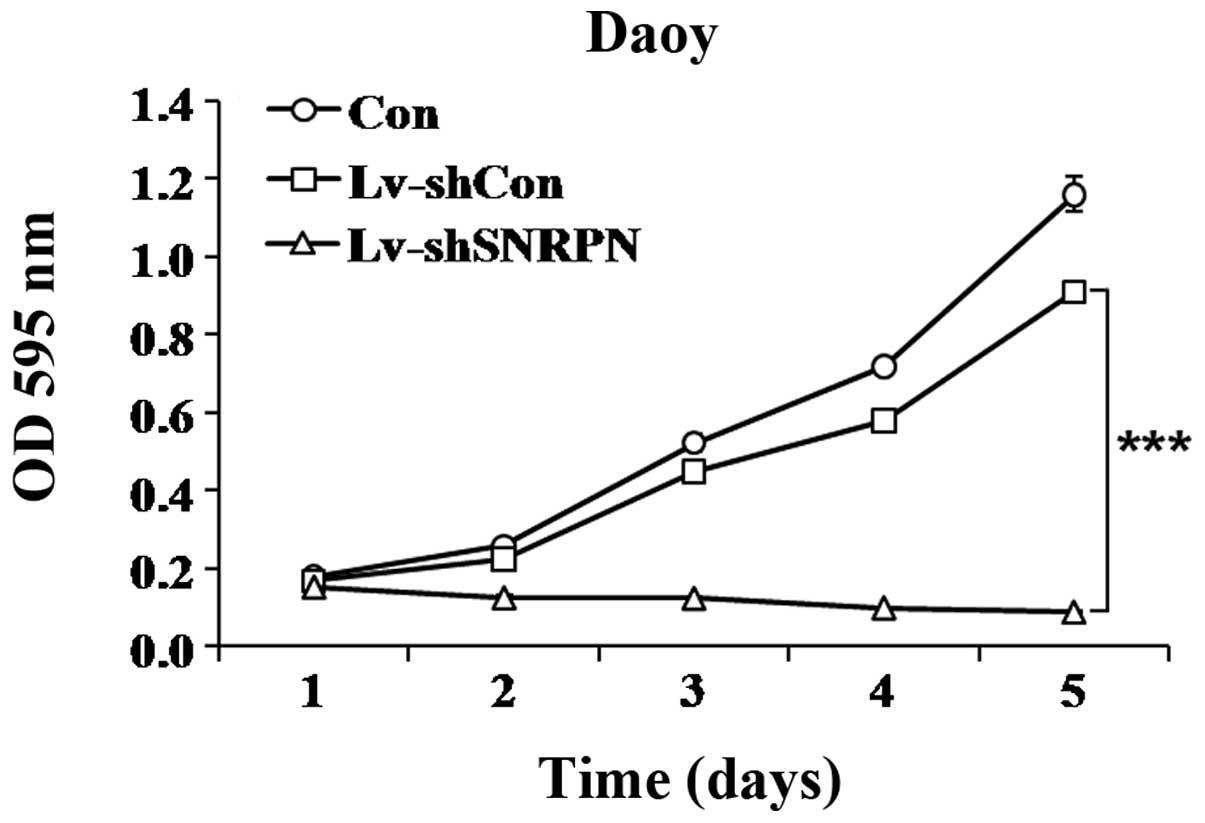
To investigate whether the alteration in the levels
of SNRPN affected the proliferation of Daoy cells, MTT cell
viability and colony formation assays were conducted. A growth
curve revealed that the number of Lv-shSNRPN infected cells
increased at a markedly slower rate than the uninfected cells or
the Lv-shCon infected cells (Fig.
3). On day 5 following Lv-shSNRPN infection, the proliferation
rate was markedly reduced, with a 90.2% reduction.
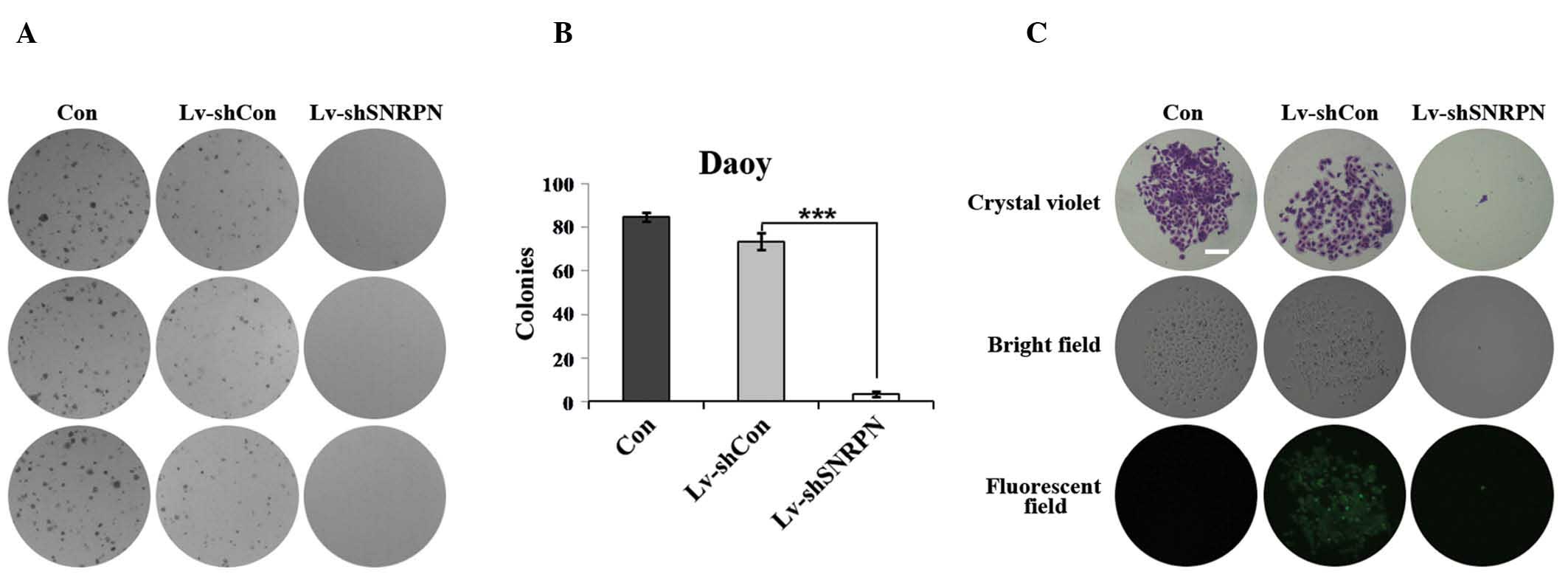
In addition, the Lv-shSNRPN-infected cells produced
fewer colonies than the uninfected cells or the Lv-shCon-infected
cells (Fig. 4A). Only three
SNRPN-depleted Daoy cell colonies were detected as compared with
~85 and 74 uninfected and Lv-shCon infected cell colonies,
respectively (Fig. 4B).
Furthermore, the size of each colony was also evidently smaller in
SNRPN-depleted cells than in the uninfected cells or
Lv-shCon-infected cells (Fig. 4C).
These data indicate that SNRPN functions as a positive regulator of
proliferation and colony formation in Daoy cells.
Effect of SNRPN knockdown on the cell
cycle distribution in the medulloblastoma cells
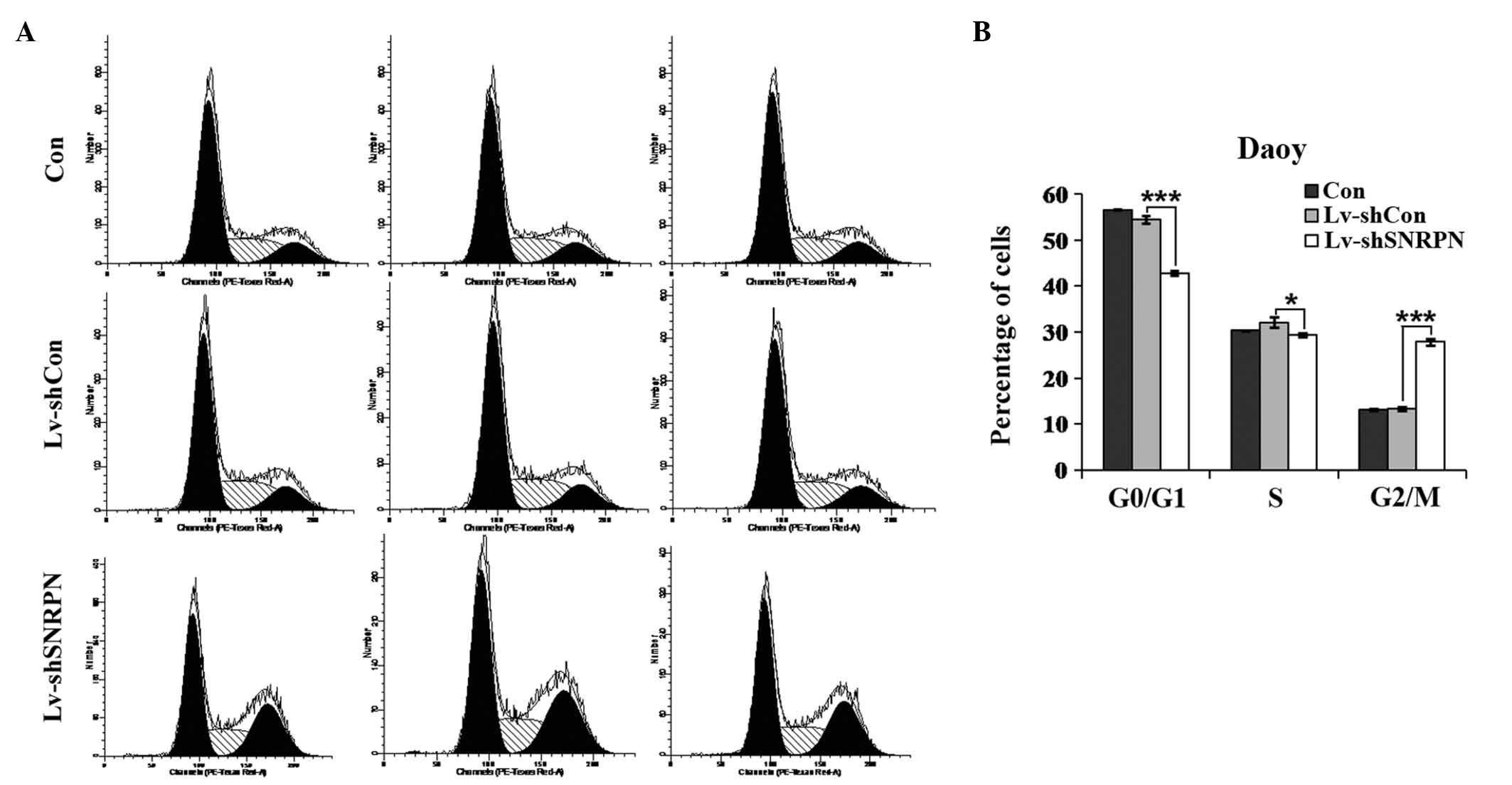
To determine the effects of SNRPN depletion on the
cell cycle distribution, flow cytometry was conducted on Daoy cells
infected with no Lv, Lv-shCon or Lv-shSNRPN, respectively. The
proportion of the cell population in G0/G1 phase in the
SNRPN-depleted Daoy cells (42.83±0.44%) was significantly lower
than those in uninfected cells (56.54±0.12%) and Lv-shCon infected
cells (54.47±0.81%), whereas the percentage of the cell population
in G2/M phase in the SNRPN-depleted Daoy cells (27.86±0.67%) was
markedly higher than those in the uninfected cells (13.12±0.15%)
and the Lv-shCon-infected cells (13.39±0.37%; Fig. 5). In addition, the proportion of
the cell population in S phase in the SNRPN-depleted Daoy cells
(29.31±0.40%) was marginally lower than those in uninfected cells
(30.34±0.10%) and Lv-shCon-infected cells (32.14±1.14%). These
results indicate that SNRPN may modulate the growth of
medulloblastoma cells through the regulation of cell cycle
progression.
Discussion
Medulloblastomas are aggressive primitive
neuroectodermal tumors that appear in the cerebellum,
Medulloblastomas develop from granule cell-like precursors that
have escaped from terminal differentiation. As a histological
class, medulloblastoma is the most common type of pediatric brain
tumor (19,20). Although there has been marked
progress in the understanding of the molecular mechanisms involved
in the etiology of medulloblastoma, and in the development of
combination treatment using surgery, chemotherapy and radiation,
the five-year survival rate for patients with medulloblastoma
remains at 70–80%, with the survival rate in young children and
infant patients even lower (3). In
addition, almost all survivors suffer from adverse, life-long
consequences from treatment. These undesirable effects are
attributable to the detrimental impacts of surgical procedures,
radiation and chemotherapy on the developing brain (21). Therefore, the identification of
novel therapeutics that improve the cure rate without harmful side
effects is imperative. A previous study demonstrated that
medulloblastoma may be divided into four molecular subtypes: WNT,
SHH, group 3 and group 4 (7).
These four subtypes exhibit notable differences in gene expression,
as determined by cDNA microarrays and immunohistochemistry
(22,23). Current therapeutic methods that
target key oncogenic promoters, such as WNT and SHH, have a number
of limitations.
In the present study, SNRPN, a polypeptide of a
small nuclear ribonucleoprotein complex, was observed to exert an
important role in cell growth in medulloblastoma cell lines.
Knockdown of SNRPN by infection with SNRPN-specific siRNA markedly
inhibited the proliferation of the medulloblastoma cells. In
addition, a colony formation assay was used to assess the
capability of cell growth in anchorage-independent conditions
comparable with the in vivo situation (24). Knockdown of SNRPN may also impair
the anchorage-independent growth of medulloblastoma cells.
Altogether, to the best of our knowledge, this is the first study
to report that SNRPN exerts a key role in medulloblastoma cell
growth.
To further investigate the mechanism of cell growth
inhibition, the cell cycle progression of the Daoy medulloblastoma
cells was determined by flow cytometric analysis. The results
revealed that the cell cycle distribution was altered following
Lv-shSNRPN infection. SNRPN knockdown significantly increased the
percentage of cells in G2/M phase, while simultaneously reducing
the proportion of cells in G0/G1 phase. The results raise the
question of the underlying mechanism of SNRPN regulation of the
medulloblastoma cell cycle. SNRPN is involved in alternative
splicing of pre-mRNA, possibly in a tissue-specific manner
(25). We hypothesize that SNRPN
recognizes specific nucleic acid sequences of specific genes
involved in the G2/M and the G1 cell cycle checkpoints, such as p53
and p21 (26,27) and regulates the expression of the
corresponding protein molecules. However, due to the limitation of
the present study, the specific underlying mechanism was not
further investigated.
SNRPN was the first protein in the PWS critically
deleted region to be identified. Numerous studies concerning the
methylation of the SNRPN gene in cancer have been published. For
example, SNRPN methylation patterns in germ cell tumors have been
reported to reflect primordial germ cell development (11). The variable methylation of SNRPN
also indicates an association between intracranial GCTs and neural
stem cells (14). In addition,
complete maintenance of SNRPN imprinting has been observed in human
uterine leiomyomas (13). Analysis
of the data from eight PTC tumor samples revealed that
amplifications of SNRPN occurred solely in tumors with a wild-type
BRAF (15). According to the
results of previous studies and those of the present study, SNRPN
regulates various tumor characteristics, including proliferation,
colony formation and the cell cycle; therefore SNRPN methylation in
medulloblastoma may also be altered and may be associated with a
specific subtype of medulloblastoma. The SNRPN methylation level in
medulloblastoma requires further analysis.
In conclusion, to the best of our knowledge, this is
the first study to define SNRPN as a functional mediator of
medulloblastoma cell growth. Knockdown of SNRPN was demonstrated to
significantly inhibit medulloblastoma cell growth and induce G2/M
phase arrest in vitro. Thus, the present study may provide a
novel therapeutic approach to treat patients with
medulloblastoma.
References
|
1
|
Polkinghorn WR and Tarbell NJ:
Medulloblastoma: tumorigenesis, current clinical paradigm, and
efforts to improve risk stratification. Nat Clin Pract Oncol.
4:295–304. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rossi A, Caracciolo V, Russo G, Reiss K
and Giordano A: Medulloblastoma: From molecular pathology to
therapy. Clin Cancer Res. 14:971–976. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gilbertson RJ: Medulloblastoma: signalling
a change in treatment. Lancet Oncol. 5:209–218. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cho YJ, Tsherniak A, Tamayo P, et al:
Integrative genomic analysis of medulloblastoma identifies a
molecular subgroup that drives poor clinical outcome. J Clin Oncol.
29:1424–1430. 2011. View Article : Google Scholar :
|
|
5
|
Kool M, Koster J, Bunt J, et al:
Integrated genomics identifies five medulloblastoma subtypes with
distinct genetic profiles, pathway signatures and
clinicopathological features. PLoS One. 3:e30882008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Thompson MC, Fuller C, Hogg TL, et al:
Genomics identifies medulloblastoma subgroups that are enriched for
specific genetic alterations. J Clin Oncol. 24:1924–1931. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zollo M: Genetics of recurrent
medulloblastoma. Lancet Oncol. 14:1147–1148. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gilbertson RJ and Ellison DW: The origins
of medulloblastoma subtypes. Annu Rev Pathol. 3:341–365. 2008.
View Article : Google Scholar
|
|
9
|
Eberhart CG: Molecular diagnostics in
embryonal brain tumors. Brain Pathol. 21:96–104. 2011. View Article : Google Scholar
|
|
10
|
Reed ML and Leff SE: Maternal imprinting
of human SNRPN, a gene deleted in Prader-Willi syndrome. Nat Genet.
6:163–167. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bussey KJ, Lawce HJ, Himoe E, et al: SNRPN
methylation patterns in germ cell tumors as a reflection of
primordial germ cell development. Genes Chromosomes Cancer.
32:342–352. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Benetatos L, Hatzimichael E, Dasoula A, et
al: CpG methylation analysis of the MEG3 and SNRPN imprinted genes
in acute myeloid leukemia and myelodysplastic syndromes. Leuk Res.
34:148–153. 2010. View Article : Google Scholar
|
|
13
|
Hashimoto K, Azuma C, Kamiura S, et al:
Maintenance of imprinting of the insulin-like growth factor II gene
(IGF2) and the small nuclear ribonucleoprotein polypeptide N gene
(SNRPN) in the human uterus and leiomyoma. Gynecol Obstet Invest.
41:50–54. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lee SH, Appleby V, Jeyapalan JN, et al:
Variable methylation of the imprinted gene, SNRPN, supports a
relationship between intracranial germ cell tumours and neural stem
cells. J Neurooncol. 101:419–428. 2011. View Article : Google Scholar
|
|
15
|
Finn S, Smyth P, O’Regan E, et al:
Low-level genomic instability is a feature of papillary thyroid
carcinoma: an array comparative genomic hybridization study of
laser capture microdissected papillary thyroid carcinoma tumors and
clonal cell lines. Arch Pathol Lab Med. 131:65–73. 2007.PubMed/NCBI
|
|
16
|
Sakoda T, Kasahara N, Hamamori Y and Kedes
L: A high-titer lentiviral production system mediates efficient
transduction of differentiated cells including beating cardiac
myocytes. J Mol Cell Cardiol. 31:2037–2047. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Soneoka Y, Cannon PM, Ramsdale EE, et al:
A transient three-plasmid expression system for the production of
high titer retroviral vectors. Nucleic Acids Res. 23:628–633. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tiscornia G, Singer O and Verma IM:
Production and purification of lentiviral vectors. Nat Protoc.
1:241–245. 2006. View Article : Google Scholar
|
|
19
|
Rubin JB and Rowitch DH: Medulloblastoma:
A problem of developmental biology. Cancer Cell. 2:7–8. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wechsler-Reya R and Scott MP: The
developmental biology of brain tumors. Annu Rev Neurosci.
24:385–428. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mabbott DJ, Spiegler BJ, Greenberg ML,
Rutka JT, Hyder DJ and Bouffet E: Serial evaluation of academic and
behavioral outcome after treatment with cranial radiation in
childhood. J Clin Oncol. 23:2256–2263. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ellison DW, Dalton J, Kocak M, et al:
Medulloblastoma: clinicopathological correlates of SHH, WNT, and
non-SHH/WNT molecular subgroups. Acta Neuropathol. 121:381–396.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Northcott PA, Hielscher T, Dubuc A, et al:
Pediatric and adult sonic hedgehog medulloblastomas are clinically
and molecularly distinct. Acta Neuropathol. 122:231–240. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang LH: Molecular signaling regulating
anchorage-independent growth of cancer cells. Mt Sinai J Med.
71:361–367. 2004.PubMed/NCBI
|
|
25
|
Ohosone Y, Mimori T, Griffith A, et al:
Molecular cloning of cDNA encoding Sm autoantigen: derivation of a
cDNA for a B polypeptide of the U series of small nuclear
ribonucleoprotein particles. Proc Natl Acad Sci USA. 86:4249–4253.
1989. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Agarwal ML, Agarwal A, Taylor WR and Stark
GR: p53 controls both the G2/M and the G1 cell cycle checkpoints
and mediates reversible growth arrest in human fibroblasts. Proc
Natl Acad Sci USA. 92:8493–8497. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Niculescu AB III, Chen X, Smeets M, Hengst
L, Prives C and Reed SI: Effects of p21(Cip1/Waf1) at both the G1/S
and the G2/M cell cycle transitions: pRb is a critical determinant
in blocking DNA replication and in preventing endoreduplication.
Mol Cell Biol. 18:629–643. 1998.PubMed/NCBI
|