Introduction
Dust mites are the predominant source of indoor
allergens worldwide, and can exacerbate allergic diseases including
asthma, rhinitis and dermatitis (1–3).
Studies regarding the biological, chemical and structural
properties of dust mite allergens have been critical for advancing
the diagnosis and treatment of allergic disease (4,5). In
particular, allergens of the commonly found house dust mite
Dermatophagoides farinae (Der f, Acari: Pyroglyphidae) have
been extensively investigated. In crude extracts of D.
farinae >30 components have been identified that can induce
human immunoglobulin (Ig)E antibodies (4,6–7).
A total of 24 groups of dust mite allergens are
currently listed in the International Union of Immunological
Societies (IUIS) nomenclature dataset (http://www.allergen.org/). These allergens belong to
various protein families, and three groups have been identified to
possess trypsin activity: Group 3 comprises trypsins; group 6,
chymotrypsins; and group 9, collagenolytic serine proteases
(8). Previous studies have
suggested that Der f 3, Der f 6 and Der f 9 are capable of
activating the kallikrein-kinin system, which is involved in
inflammation in normal human plasma (8–10).
In addition, Der f 3 has been shown to activate the complement
system, resulting in the production of anaphylatoxins (11). Therefore, these allergen groups are
likely important contributors to the hypersensitivity response
produced by exposure to house dust mites.
Current research in immunotherapy for allergic
disease is aimed at producing novel treatments that relieve the
symptoms caused by hypersensitive responses (12). Allergen-specific immunotherapy
(SIT) works by inducing the production of blocking antibodies,
creating a shift toward a T-helper 1 response and inducing
tolerance (13–14). SIT is able to induce IgG
antibodies, which compete with IgE for allergen binding, thus
inhibiting activation of IgE-dependent mast cells and basophils,
and reducing IgE-mediated allergic inflammation (15–16).
Conventional immunotherapies rely on crude dust mite extracts,
however SIT is more successful with recombinant mite allergens
(17). The use of recombinant
allergens requires a good understanding of their structures and
functions, in order to determine how they bind IgE and elicit an
allergic response.
To improve understanding regarding the contributions
of Der f 3, Der f 6, and Der f 9 to the allergenicity of D.
farinae, the present study used a bioinformatics approach to
compare amino acid sequences and predict their physiochemical
characterizations, secondary structures, tertiary structures and
B-cell epitopes. These analyses may aid the future development of
SIT for Der f allergens.
Materials and methods
Sequence retrieval
The amino acid sequences of Der f 3, Der f 6 and Der
f 9 were retrieved from the International Union of Immunological
Societies (IUIS) nomenclature database (18) and the National Center for
Biotechnology Information protein sequence database (accession nos.
BAA09920.1, AAF28423.1 and AAP35067.1, respectively). Signal
peptide sequences were removed from the present analysis.
Physiochemical characterization
Physiochemical characteristics, including
theoretical isoelectric point (pI), molecular weight, number of
positively and negatively charged residues, extinction
coefficients, instability index, aliphatic index and grand average
of hydropathicity (GRAVY), were predicted using ExPASy ProtParam
(19).
Primary and secondary structure
analysis
DNAMAN version 6.0 software (Lynnon, Corp.,
Pointe-Claire, QC, Canada) was used to align the protein sequences.
Prediction of transmembrane helices within the proteins was
performed using the TMHMM 2.0 server (20). Predictions of the secondary
structure (α-helices, β-sheets and random coils) were made using
the Jpred 3 secondary structure prediction server (21). The active site and functional
domains of the proteins were predicted using ExPASy PROSITE
(19).
Three-dimensional (3D) model building and
evaluation
Homology modeling and molecular dynamics were used
to construct 3D structures of Der f 3, Der f 6 and Der f 9.
Selecting an appropriate template is a critical stage to certify
the quality of the final 3D structure. A Protein Basic Local
Alignment Search Tool (BLASTP) online server (22) search, with default parameters, was
performed against the Protein Data Bank (PDB), in order to identify
suitable templates of Der f 3, Der f 6, and Der f 9 for homology
modeling. Following the BLASTP search, appropriate templates for
generating 3D models of the proteins were selected, based on a high
score, lower e-value and maximum sequence identity. BLAST results
that indicated potential templates for modeling were PDB ID: 1A0J_A
for Der f 3, which displayed 32% amino acid sequence identity; and
PDB ID: 1DST_A and PDB ID: 4D8N_A for Der f 6 and Der f 9
respectively, which displayed 32 and 34% amino acid sequence
identities. Prediction of the 3D structure of Der f 3, Der f 6 and
Der f 9 was conducted using the homology-modeling program MODELLER
version 9.12, as previously described (23,24).
Following alignment of the query and template proteins, and removal
of potential errors, loop refinement was performed using the loop
optimization method of MODELLER 9.12. The predicted structures were
saved in PDB format, and energy minimization was performed using
the GROMOS96 force field in Swiss-PdbViewer, in order to rectify
unfavorable clashes and improve the stereochemical quality
(25).
Model evaluation is important to verify the quality
of the 3D model. Therefore, PROCHECK (26) was used to evaluate the quality of
the predicted models by Ramachandran plot assessment. Furthermore,
model verification was performed using the overall quality factor
(ERRAT) (27) and Verify 3D
(28) programs. Superimposition of
the query and template structures, and visualization of the
generated 3D models was performed using UCSF Chimera 1.8 (29).
B-cell antibody epitope prediction
Amino acid sequences of Der f 3, Der f 6 and Der f 9
were submitted to Immune Epitope Database and Analysis Resource
(IEDB) (30), ABCpred (31) and BepiPred (32) servers, in order to predict the
possible linear B-cell epitopes with the default threshold. IEDB
uses a collection of methods to predict linear B-cell epitopes,
based on sequence characteristics of the antigen using amino acid
scales and hidden Markov models (HMM). BepiPred predicts the
location of linear B-cell epitopes, with a core epitope threshold
set at 0.35, using a combination of HMM and a propensity scale
method. ABCpred predicts B-cell epitopes in an antigen sequence,
and was the first server developed based on the recurrent neural
network (machine-based technique), using fixed-length patterns with
a default threshold of 0.51. The predicted epitope regions were
mapped to the predicted secondary and 3D protein structures, and
DNAMAN 6.0 was used to generate a multiple-sequence alignment to
identify the epitope regions of Der f 3, Der f 6 and Der f 9.
Results
Physiochemical characterization of Der f
3, Der f 6 and Der f 9
A comparison between the physiochemical
characteristics of Der f 3, Der f 6 and Der f 9, as predicted using
ProtParam, is presented in Table
I. Der f 3 and Der f 6 were shown to possess similar
characteristics; whereas fewer similarities were observed with Der
f 9. The pI values indicated that Der f 9 is an alkaline protein,
and Der f 3 is more acidic, as compared with Der f 6. Extinction
coefficients of Der f 3, Der f 6 and Der f 9 depended on the molar
extinction coefficients of Tyr, Trp and Cys residues. Stability of
Der f 3, Der f 6 and Der f 9 was investigated by analyzing the
instability, aliphatic and GRAVY indices of these three proteins.
The instability index value was lowest for Der f 6, therefore Der f
6 is likely to be the most stable of the three allergens. The
aliphatic index is defined as the relative volume occupied by
aliphatic side chains (alanine, valine, isoleucine and leucine);
and all three proteins were predicted to be fat-soluble. The GRAVY
index indicated that all three proteins were also hydrophilic.
 | Table IComparison of physiochemical
characteristics of Der f 3, Der f 6 and Der f 9 allergens of
Dermatophagoides farinae. |
Table I
Comparison of physiochemical
characteristics of Der f 3, Der f 6 and Der f 9 allergens of
Dermatophagoides farinae.
| Parameter | Der f 3 | Der f 6 | Der f 9 |
|---|
| Accession no. | BAA09920.1 | AAF28423.1 | AAP35067.1 |
| Number of amino
acids | 232 | 230 | 224 |
| Total number of
atoms | 3439 | 3491 | 3334 |
| Formula |
C1087H1710N302O349S10 |
C1094H1738N298O349S12 |
C1053H1672N290O329S9 |
| Molecular weight
(u) | 24913.9 | 25034.3 | 23947.1 |
| Arg+Lys | 21 | 22 | 17 |
| Asp+Glu | 26 | 25 | 13 |
| Theoretical pI | 5.36 | 5.81 | 8.68 |
| Extinction
coefficients (M−1cm−1 at 260 nm) | 38765–38390 | 35785–35410 | 35785–35410 |
| Instability
index | 30.14 | 29.24 | 32.69 |
| Aliphatic
index | 82.28 | 80.04 | 88.84 |
| GRAVY | −0.284 | −0.369 | −0.016 |
Protein sequence alignment and active
site prediction
Protein sequence alignment was performed using the
default parameters of DNAMAN 6.0. Der f 3, Der f 6 and Der f 9
exhibited a 45.61% sequence identity. Prosite prediction
demonstrated that Der f 3, Der f 6 and Der f 9 each contain an
active site which is formed of a catalytic triad of histidine,
aspartate and either serine or cysteine (Fig. 1). In addition, two
sequence-specific trypsin functional domains of Der f 3, Der f 6
and Der f 9 (XTAAHC and XXCXGDS(C)GGPXV; bold
indicates active site amino acid residues), which surround the
histidine and serine/cysteine active sites, were shown to be highly
conserved in the proteins (Fig. 1,
domains I and II).
Secondary structure prediction
The amino acid sequences of Der f 3, Der f 6 and Der
f 9 were submitted to TMHMM 2.0, which is capable of predicting
transmembrane helices. The three proteins were not predicted to
contain any transmembrane helices, and localization was predicted
to be outside of the membrane. Secondary structure prediction,
using Jpred 3, demonstrated that the three proteins contain
α-helices, β-sheets and random coils (Fig. 2). The largest number of residues
was observed in the random coils, followed by the β-sheets and the
α-helices. More residues were detected in the α-helices of Der f 3,
as compared with Der f 6 and Der f 9. In addition, more residues
were detected in the β-sheets of Der f 6, as compared with Der f 3
and Der f 9 (Table II).
| Table IISecondary structure elements for Der
f 3, Der f 6 and Der f 9 allergens of Dermatophagoides
farinae. |
Table II
Secondary structure elements for Der
f 3, Der f 6 and Der f 9 allergens of Dermatophagoides
farinae.
| Allergen | α-helices (%) | β-sheets (%) | Random coils
(%) |
|---|
| Der f 3 | 5.60 (2
domains) | 29.74 (13
domains) | 64.66 |
| Der f 6 | 5.22 (2
domains) | 32.17 (14
domains) | 62.61 |
| Der f 9 | 4.91 (2
domains) | 30.36 (14
domains) | 64.73 |
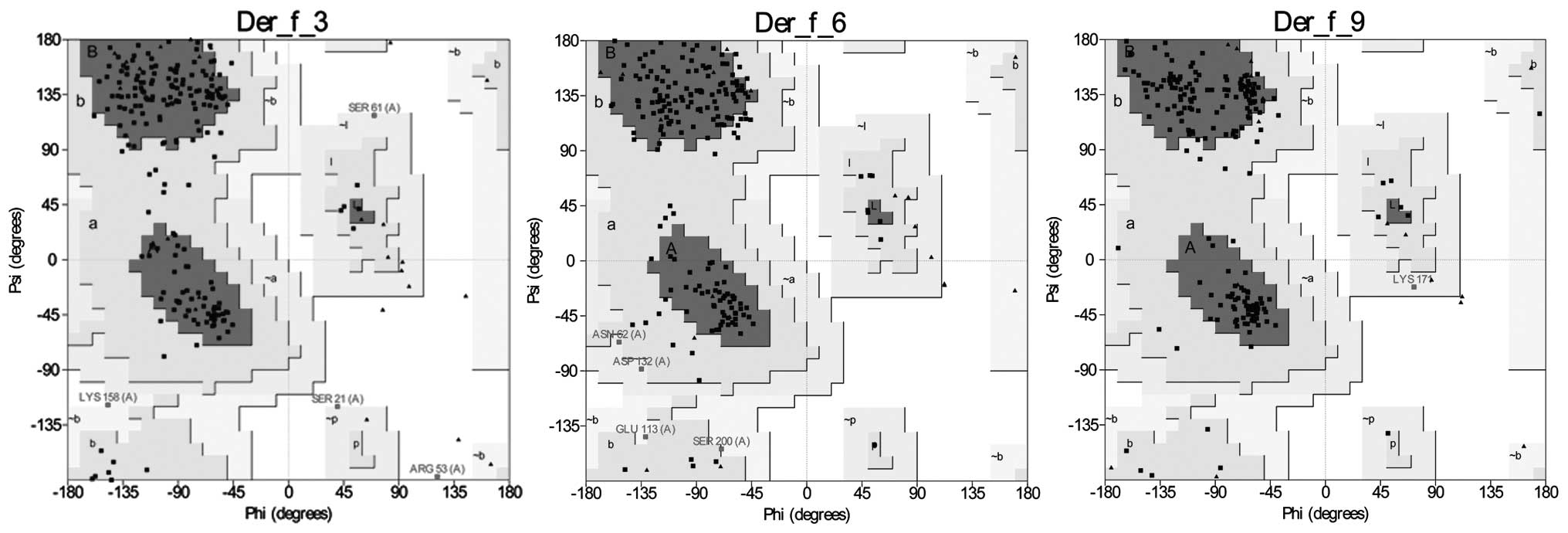
3D model building, refinement, and
evaluation
3D structures of Der f 3, Der f 6 and Der f 9 were
generated using MODELLER 9.12 with the following templates: PDB
ID:1A0J_A, 1DST_A and 4D8N_A, respectively. The three models were
viewed using Chimera 1.8, and a Ramachandran plot analysis of the
models was conducted using PROCHECK. Analysis of the models
indicated a high probability of confirmation, with 99.5, 100 and
100% of residues of Der f 3, Der f 6 and Der f 9, respectively, in
the allowed region of the Ramachandran plot (Fig. 3). It is generally accepted that if
90% of residues are present in the allowed region of a Ramachandran
plot, the model is reliable. Verification of the 3D analysis
demonstrated that 94.85, 84.42 and 89.78% of the Der f 3, Der f 6
and Der f 9 residues, respectively, had an average 3D-1D score of
<0.2, thus indicating that the three models were compatible with
their sequences. Furthermore, the ERRAT of the models of Der f 3,
Der f 6 and Der f 9 were 81.860, 82.297 and 84.722, respectively.
These results suggest that all computationally-generated models in
the present study are considered to be of good quality and suitable
for structural analysis.
The predicted 3D structures of Der f 3, Der f 6 and
Der f 9, like the secondary structures, were shown to contain
α-helices, β-sheets and random coils; however, the number of amino
acids of these three structure elements differed slightly between
the 3D and secondary structures. The percentage of overall amino
acids located in α-helices was 6.90 (two domains), 12.17 (four
domains) and 10.27% (three domains) in Der f 3, Der f 6 and Der f
9, respectively; whereas the percentage of amino acids in β-sheets
was 27.59 (13 domains), 26.96 (13 domains) and 30.80% (12 domains),
respectively (Fig. 4). 3D
structure overlap of the Der f 3, Der f 6 and Der f 9 proteins
identified more overlapping in the α-helix and β-sheet domains, as
compared with the random coil domains. The three active sites of
Der f 3, Der f 6 and Der f 9 were shown to completely overlap
within the 3D structure, and fold onto each other to constitute the
active site of the enzyme (Fig.
4).
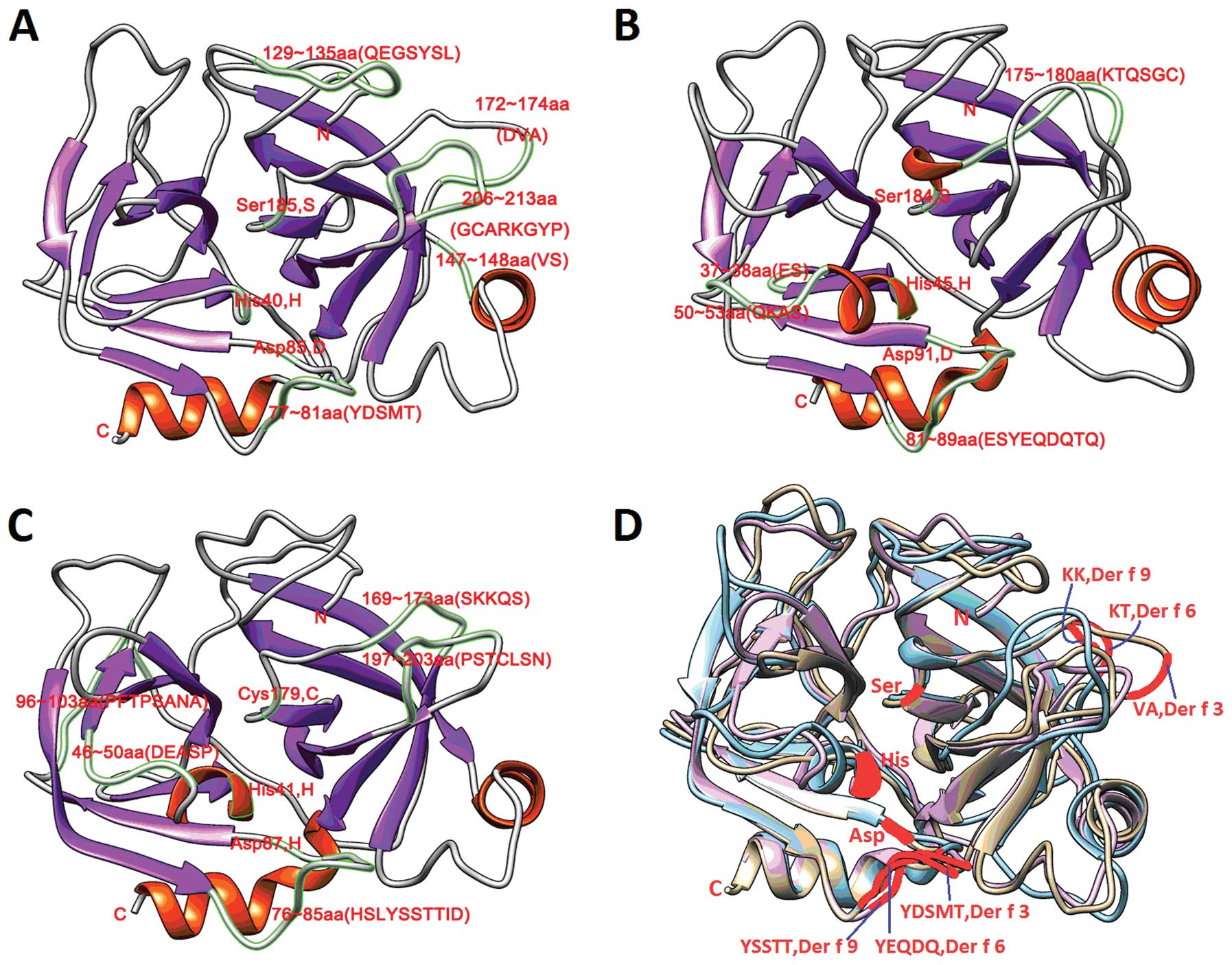 | Figure 4Homology-modeled structures of Der f
3, Der f 6, and Der f 9 allergens of Dermatophagoides
farinae. (A) Secondary structure of Der f 3 contains two
α-helices and 13 β-sheets. Histidine, aspartate and serine formed
into catalytic triad of active sites in Der f 3, which fold onto
each other in the 3D model. (B) Secondary structure of Der f 6
contains four α-helices and 13 β-sheets. Histidine, aspartate and
serine formed into catalytic triad of active sites in Der f 6,
which fold onto each other in the 3D model. (C) Secondary structure
of Der f 9 contains three α-helices and 12 β-sheets. Histidine,
aspartate and cysteine formed into catalytic triad of active sites
in Der f 9, which fold onto each other in the 3D model. α-helices
are shown in orange, β-sheets in purple and random coils in gray.
(D) 3D structural overlap of Der f 3, Der f 6, and Der f 9 revealed
more overlapping in α-helix and β-sheet domains, whereas there was
less overlapping in β-turn and random coil domains. Active site
residues of Der f 3, Der f 6, and Der f 9 overlapped completely in
the 3D structure and folded onto each other to constitute catalytic
triad of active sites. Der f 3 is shown in light brown, Der f 6 in
light blue and Der f 9 in purple. The active sites and overlapping
epitopes are shown in red. |
The predicted secondary and 3D structures of Der f
3, Der f 6 and Der f 9 had slightly different compositions.
Peptides 103–105 coiled into a β-sheet in the predicted secondary
structure of Der f 3; however, this was not observed in the 3D
structure prediction. In Der f 6, peptides 109–110 coiled into
β-sheets only in the predicted secondary structure; whereas
peptides 45–49 and 181–183 coiled into α-helices and 187–190 into a
β-sheet only in the predicted 3D structure. In Der f 9, peptides
104–106 coiled into a β-sheet only in the secondary structure;
whereas peptides 40–43 coiled into an α-helix and 164–168 into a
β-sheet only in its 3D structure.
Predicted B-cell epitopes
B-cell epitopes of Der f 3, Der f 6, and Der f 9
were predicted using IEDB Analysis Resource, ABCpred and BepiPred,
and were verified in the predicted secondary and 3D structures. Der
f 3, Der f 6 and Der f 9 were predicted to contain 5, 4 and 5 main
potential epitopes, respectively, all located in random coils
(Table III). Amino acid sequence
alignment and overlapping 3D structures demonstrated an overlap in
the epitopes of Der f 3, Der f 6 and Der f 9 (Fig. 1; domains A and B, peptides 83–87
and 179–180; Fig. 4D), however the
residues in two overlapping epitope sequences were not
identical.
| Table IIIB-cell epitopes for Der f 3, Der f 6
and Der f 9 allergens of Dermatophagoides farinae. |
Table III
B-cell epitopes for Der f 3, Der f 6
and Der f 9 allergens of Dermatophagoides farinae.
| Allergen | Predicted epitope
sequences | Peptide
numbers |
|---|
| Der f 3 | YDSMT | 77–81 |
| QEGSYSL | 129–135 |
| VS | 147–148 |
| DVA | 172–174 |
| GCARKGYP | 206–213 |
| Der f 6 | ES | 37–38 |
| QKAS | 50–53 |
| ESYEQDQTQ | 81–89 |
| KTQSGC | 175–180 |
| Der f 9 | DEASP | 46–50 |
| HSLYSSTTID | 76–85 |
| PFTPSANA | 96–103 |
| SKKQS | 169–173 |
| PSTCLSN | 197–203 |
Discussion
Allergic diseases are one of the most common types
of human disease. Besides avoiding allergen exposure,
allergen-specific immunotherapy (SIT) is the only curative
treatment option (33). Patients
respond to symptomatic pharmacotherapy, such as antihistamines and
corticosteroids, however the majority of patients report inadequate
symptom relief (34). SIT provides
an alternative disease-specific treatment, which induces tolerance
to the allergen (35). SIT can
result in long-term remission of allergic symptoms and may reduce
the chances of developing novel sensitization to other allergens
(17).
Recently, the increased prevalence of allergic
diseases has placed greater demand on diagnostic and therapeutic
products; however, crude mite allergen extracts that are applied
for diagnostic and therapeutic purposes do not meet the demand
(36). Crude extracts include
non-allergenic antigens and ineffective concentrations of important
allergens, resulting in numerous side effects (37). The use of recombinant allergens
offers a better prospect for the rational and accurate diagnosis
and treatment of allergies. Pure and standardized recombinant
allergens contain the majority of the IgE-binding epitopes of an
allergen source, and can be formulated to replace natural extracts
(38). The use of recombinant
allergens in diagnosis allows the exact identification of the
molecules that are provoking the allergic reaction (37). Der f 3, Der f 6 and Der f 9
allergens are currently listed in the IUIS nomenclature dataset,
and developing recombinant allergens of these three proteins may
contribute to the diagnosis and treatment of mite-induced allergic
diseases.
The results of the present study indicated that Der
f 9 is alkaline, whereas Der f 3 and Der f 6 are acidic. Der f 3
and Der f 6 were shown to possess more similar characteristics to
one another, as compared with Der f 9. These three proteins are
45.61% identical in amino acid sequence and contain three active
sites: His, Asp and Ser/Cys, which are common residues constituting
the active site of a protease. A 3D analysis indicated that these
sites are likely to fold onto each other, constituting the active
site of the enzyme. Furthermore, two trypsin functional domains
were identified in Der f 3, Der f 6 and Der f 9 (XTAAHC and
XXCXGDS(C)GGPXV), which have high amino acid sequence identity and
were shown to surround the His and Ser/Cys active sites.
Secondary and tertiary protein structures are
important for the prediction of epitopes. α-Helices and β-sheets
are two common secondary structures of protein, whose conformations
are maintained by hydrogen bonds, rendering it unlikely for epitope
sequences to be located within them. Conversely, random coils are
located in the surface-exposed region proteins, and often contain
epitope sequences (39). In the
present study, the secondary and tertiary structures of Der f 3,
Der f 6 and Der f 9 were predicted to contain α-helices, β-sheets
and random coils. Epitope prediction identified 4–5 potential
epitopes located in the random coils of each of the three
allergens. Furthermore, the epitope sequences of Der f 3, Der f 6
and Der f 9 were shown to overlap in two domains. However, the
amino acids in these two domains were not 100% identical, this is
likely a reflection of the <50% identity between Der f 3, Der f
6 and Der f 9 sequences.
In conclusion, the present study conducted a
biochemical and genetic analysis of the Der f 3, Der f 6 and Der f
9 allergens. The results may provide a basis for further studies of
the allergenicity and function of these three allergens, and may
contribute to the development of a vaccine for allergen-specific
immunotherapy.
Acknowledgements
The present study was supported by the National
Sciences Foundation of China (grant nos. NSFC31272369 and
NSFC81001330) and the Jiangsu Provincial Health Department (grant
no. Z200914).
Abbreviations:
|
3D
|
three-dimensional
|
|
IUIS
|
International Union of Immunological
Societies
|
|
NCBI
|
National Center for Biotechnology
Information
|
|
PDB
|
Protein Data Bank
|
|
IEDB
|
Immune Epitope Database and Analysis
Resource
|
References
|
1
|
Milián E and Díaz AM: Allergy to house
dust mites and asthma. P R Health Sci J. 23:47–57. 2004.PubMed/NCBI
|
|
2
|
Nadchatram M: House dust mites, our
intimate associates. Trop Biomed. 22:23–37. 2005.
|
|
3
|
Bunnag C, Jareoncharsri P, Tantilipikorn
P, Vichyanond P and Pawankar R: Epidemiology and current status of
allergic rhinitis and asthma in Thailand - ARIA Asia-Pacific
Workshop report. Asian Pac J Allergy Immunol. 27:79–86.
2009.PubMed/NCBI
|
|
4
|
Cui Y: Structural biology of mite
allergens. Mol Biol Rep. 40:681–686. 2013. View Article : Google Scholar
|
|
5
|
Morales M, Iraola V, Leonor JR and Carnés
J: Enzymatic activity of allergenic house dust and storage mite
extracts. J Med Entomol. 50:147–54. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Thomas WR, Smith WA, Hales BJ, Mills KL
and O’Brien RM: Characterization and immunobiology of house dust
mite allergens. Int Arch Allergy Immunol. 129:1–18. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Thomas WR, Smith WA and Hales BJ: The
allergenic specificities of the house dust mite. Chang Gung Med J.
27:563–569. 2004.PubMed/NCBI
|
|
8
|
Cui Y, Zhou Y, Shi W, et al: Molecular
cloning, expression, sequence analyses of dust mite allergen Der f
6 and its IgE-binding reactivity with mite allergic asthma patients
in southeast China. Mol Biol Rep. 39:961–968. 2012. View Article : Google Scholar
|
|
9
|
Takahashi K, Aoki T, Kohmoto S, et al:
Activation of kallikrein-kinin system in human plasma with purified
serine protease from Dermatophagoides farinae. Int Arch Allergy
Appl Immunol. 91:80–85. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Polosa R: Role of the kinin-kallikrein
pathway in allergic diseases. Allergy. 48:217–225. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Maruo K, Akaike T, Ono T, Okamoto T and
Maeda H: Generation of anaphylatoxins through proteolytic
processing of C3 and C5 by house dust mite protease. J Allergy Clin
Immunol. 100:253–260. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Schei MA, Hessen JO and Lund E: House-dust
mites and mattresses. Allergy. 57:538–542. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Durham SR and Till SJ: Immunologic changes
associated with allergen immunotherapy. J Allergy Clin Immunol.
102:157–164. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Valenta R: The future of antigen-specific
immunotherapy of allergy. Nat Rev Immunol. 2:446–453.
2002.PubMed/NCBI
|
|
15
|
Larché M, Akdis CA and Valenta R:
Immunological mechanisms of allergen-specific immunotherapy. Nat
Rev Immunol. 6:761–771. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kowalski ML and Jutel M: Mechanisms of
specific immunotherapy of allergic diseases. Allergy. 53:485–492.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cappella A and Durham SR: Allergen
immunotherapy for allergic respiratory diseases. Hum Vaccin
Immunother. 8:1499–1512. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Larsen JN and Løwenstein H: Allergen
nomenclature. J Allergy Clin Immunol. 97:577–578. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wilkins MR, Gasteiger E, Bairoch A, et al:
Protein identification and analysis tools in the ExPASy server.
Methods Mol Biol. 112:531–552. 1999.PubMed/NCBI
|
|
20
|
Krogh A, Larsson B, von Heijne G and
Sonnhammer EL: Predicting transmembrane protein topology with a
hidden Markov model: application to complete genomes. J Mol Biol.
305:567–580. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cole C, Barber JD and Barton GJ: The Jpred
3 secondary structure prediction server. Nucleic Acids Res.
36:W197–W201. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Altschul SF, Madden TL, Schäffer AA, et
al: Gapped BLAST and PSI-BLAST: a new generation of protein
database search programs. Nucleic Acids Res. 25:3389–3402. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sali A and Blundell TL: Comparative
protein modeling by satisfaction of spatial restraints. J Mol Biol.
234:779–815. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Eswar N, Webb B, Marti-Renom MA, et al:
Comparative protein structure modeling using Modeller. Curr Protoc
Bioinformatics. Chapter 5(Unit 5.6) View Article : Google Scholar : 2006.
|
|
25
|
Guex N and Peitsch MC: SWISS-MODEL and the
Swiss-PdbViewer: an environment for comparative protein modeling.
Electrophoresis. 18:2714–2723. 1997. View Article : Google Scholar
|
|
26
|
Laskowski RA, MacArthur MW, Moss DS and
Thornton JM: PROCHECK: A program to check the stereochemical
quality of protein structures. J Appl Cryst. 26:283–291. 1993.
View Article : Google Scholar
|
|
27
|
Colovos C and Yeates TO: Verification of
protein structures: patterns of non-bonded atomic interactions.
Protein Sci. 2:1511–1519. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lüthy R, Bowie JU and Eisenberg D:
Assessment of protein models with three-dimensional profiles.
Nature. 356:83–85. 1992. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Pettersen EF, Goddard TD, Huang CC, et al:
UCSF Chimera - a visualization system for exploratory research and
analysis. J Comput Chem. 25:1605–1612. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Vita R, Peters B and Sette A: The curation
guidelines of the immune epitope database and analysis resource.
Cytometry A. 73:1066–1070. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Saha S and Raghava GP: Prediction of
continuous B-cell epitopes in an antigen using recurrent neural
network. Proteins. 65:40–48. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Larsen JE, Lund O and Nielsen M: Improved
method for predicting linear B-cell epitopes. Immunome Res.
2:22006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Klimek L and Pfaar O: A comparison of
immunotherapy delivery methods for allergen immunotherapy. Expert
Rev Clin Immunol. 9:465–474. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wu AY: Immunotherapy - Vaccines for
allergic diseases. J Thorac Dis. 4:198–202. 2012.PubMed/NCBI
|
|
35
|
Frew AJ: Allergen immunotherapy. J Allergy
Clin Immunol. 125:S306–S313. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Cui Y, Zhou Y, Ma G, et al: Cloning,
bioinformatics analysis, and expression of the dust mite allergen
Der f 5 of Dermatophagoides farinae. Braz J Med Biol Res.
45:746–752. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Schmidt G, Gadermaier G, Pertl H, et al:
Production of recombinant allergens in plants. Phytochem Rev.
7:539–552. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Larché M: Peptide and recombinant
immunotherapy. Immunol Allergy Clin North Am. 31:377–389. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Sikic K, Tomic S and Carugo O: Systematic
comparison of crystal and NMR protein structures deposited in the
protein data bank. Open Biochem J. 4:83–95. 2010. View Article : Google Scholar
|