Introduction
Breast cancer is a notable cause of morbidity and
mortality in females, and is associated with a high incidence of
recurrence and treatment failure (1). Growing evidence suggests that in
patients with breast cancer, tumor metastasis and poor clinical
outcome may be due to the evasion of the effects of systemic
therapies by a small subset of stem-like cells, termed breast
cancer stem cells (BCSCs) (2). In
severe combined immunodeficiency (SCID) model mice, BCSCs are
identified by the presence of a combination of
CD44+/CD24−, aldehyde dehydrogenase 1
activity, mammosphere formation and tumorigenicity (3,4). The
BCSC population varies widely among cancerous tissues and cell
lines and is often associated with aggressive types of breast
cancer (5). Alterations in
critical signaling pathways such as Notch, Wnt/β-catenin and
Hedgehog allow stem cells to undergo uncontrolled proliferation
(6,7). Notably, previous studies have
suggested that the activation of Notch promotes the expansion of
BCSCs. In cells with enriched BCSC markers, Notch-1 signaling
activity was observed to increase 4-fold while Notch-4 signaling
activity increased 8-fold (8,9). In
addition, BCSCs are able to undergo epithelial-mesenchymal
transition (EMT), a process that is involved in the facilitation of
breast cancer progression (10–12).
It has been suggested that Snail1 activity is required for EMT
initiation, whereas Twist1 serves a role in the maintenance of EMT
in mammary epithelial and breast cancer cells (10,13).
Previous studies have suggested that special AT-rich
sequence-binding protein-1 (SATB1), which functions as a genome
organizer, is crucial in the progression of breast cancer towards
metastasis. Deregulation of SATB1 in malignant cells alone, in lieu
of multiple successive genomic aberrations, is sufficient to alter
the expression of a large number of genes required for the
progression of cancer to metastasis (14,15).
In the current study, the effects of SATB1 overexpression or
knockdown were investigated on the stem cell populations in the
breast cancer cell lines MCF-7 or BT-549, respectively, by
assessment of in vitro mammosphere formation and
CD44+/CD21− expression, and observation of
tumor formation in SCID mice. A previous study demonstrated that
the number of mammospheres generated was an indirect measure of
mammary stem cell self-renewal; mammosphere size was representative
of progenitor cell proliferation; and that the
CD44+/CD24− population of breast cancer cells
display characteristics of stem cells (3). Thus, these factors were investigated
in the present study. In order to determine a possible mechanism of
SATB1 in maintaining the BCSC population, the expression levels of
Notch1, Notch4, Hes1, Snail1 and Twist1 were examined in the MCF-7
and BT-549 cell lines.
Materials and methods
Lentiviral construction and cell
transfection
All lentiviral constructs were prepared by Shanghai
GeneChem Co., Ltd. (Shanghai, China). Lentivirus GV287-SATB1 and
Lentivirus GV115-SATB1-shRNA transfection was conducted in
accordance with the manufacturer’s instructions (GeneChem, Co.,
Ltd). The human SATB1 cDNA was subcloned into the GV287 lentiviral
vector (http://www.genechem.com.cn/Zaiti.aspx?zt=GV287) and
the human SATB1-small hairpin (sh)RNA targeted to SATB1 or negative
control (NC)shRNA (GeneChem, Co., Ltd) were subcloned into the
GV115 lentiviral vector (http://www.genechem.com.cn/Zaiti.aspx?zt=GV115).
Subsequently, the lentivirus vector and packaging plasmid mixes
were transfected into HEK293T cells (American Type Culture
Collection, Manassas, VA, USA) using Lipofectamine® 2000
(Invitrogen Life Technologies, Carlsbad, CA, USA). Following 48-h
transfection, the Dulbecco’s modified Eagle’s medium (DMEM; Gibco,
Life Technologies, Grand Island, NY, USA) was harvested and
filtered. Subsequent to confirmation via restriction digestion with
AgeI restriction enzyme (New England Biolabs, Beverly, MA, USA) and
DNA sequencing performed by GeneChem using a pyrosequencing method,
large-scale GV287-SATB1, GV115-SATB1-shRNA and GV115-NC-shRNA
viruses were produced and used for the transfection into the breast
cancer cell lines.
Cell lines culture and mammosphere
assay
The human breast cancer cell lines MCF-7 and BT-549
were obtained from the American Type Culture Collection (Manassas,
VA, USA) and maintained in high-glucose DMEM (GE Healthcare Life
Sciences, Logan, UT, USA) supplemented with 10% fetal bovine serum
(FBS; GE Healthcare Life Sciences) at 37°C in 5% CO2.
For transfection, the MCF-7 cells were infected with the
GV287-SATB1 or the control lentivirus GV287, whereas BT-549 cells
were infected with GV115-SATB1-shRNA or lentivirus GV115-NC-shRNA.
Following 12~16 h incubation, the viruses were removed and replaced
with fresh DMEM. For the mammosphere experiments, single-cell
suspensions of the breast cancer cells were plated on ultra-low
attachment plates (Corning Inc., Corning, NY, USA) at a density of
1×104 cells/well in DMEM supplemented with 2% (v/v) B-27
(Invitrogen Life Technologies) and 20 ng/ml EGF and bFGF
(Peprotech, Inc., Rocky Hill, NJ, USA). Fresh medium was added to
the culture every 48 h and images of the resultant non-adherent
mammospheres were captured in triplicate using a digital camera
(Coolpix 990; Nikon Corp., Tokyo, Japan) on day 10. The diameters
of the spheres were measured using Photoshop CS5 (Adobe Systems,
Inc., San Jose, CA, USA) and the average sphere sizes in each of
the 10 fields were calculated. Quantification of the efficiency of
sphere formation involved counting the mammospheres under a CK40
light microscope (Olympus Corp., Tokyo, Japan) at a magnification
of ×10 and recording the number of mammospheres/spheres formed in
the 96 wells divided by the original number of single cells seeded,
expressed as a percentage.
Flow cytometric analysis
Adherent cells were lifted using 0.25% (v/v) trypsin
and washed with phosphate-buffered saline (PBS) (Spectrum Chemical
(Shanghai) Co., Ltd, Shanghai, China), while mammosphere cells were
collected via centrifugation for 5 min at 300 × g with a XKA-2200
centrifuge (Xiangyi Group, Changsha, China), dissociated using
trypsin and washed with PBS. The dissociated cells were resuspended
to a final concentration of 5×106 cells/ml in PBS and
5×105 cells (100 μl) were incubated with 0.5~1.0 μg
phycoerythrin (PE)-conjugated mouse anti-human/mouse CD44 (12-0441)
and allophycocyanin (APC)-conjugated mouse anti-human CD24
antibodies (17-0247) or isotype matched control antibodies (1:200;
BD Biosciences, Heidelberg, Germany) at 4°C for 60 min. The cells
were then washed twice with ice-cold PBS and collected by
centrifugation for 5 min at 300 × g for flow cytometric analysis
using the FACSAria II (BD Biosciences) with FACSDiva software (BD
Biosciences) indicating APC (CD24) fluorescence on the x-axis and
PE (CD44) fluorescence on the y-axis.
Invasion assays
A Transwell system (24 wells; 8 μm pore size; BD
Biosciences) coated with 2 mg/ml basement membrane Matrigel (BD
Biosciences) was used for the in vitro invasion assays. A
total of 1×105 cells were suspended in serum-free DMEM
(Gibco) in the upper chamber of each well, while the lower chamber
of each well was filled with 750 μl DMEM supplemented with 10% FBS.
Subsequent to suspension for 24 h, the filters were fixed with
methanol and stained with 0.1% crystal violet (Spectrum Chemical
(Shanghai) Co., Ltd). The number of cells in at least five randomly
selected microscope fields (the filter was divided into 16
microscopic fields and five were selected using a random number
method) were then counted and underwent statistical analysis.
Implantation of cells in SCID model
mice
MCF-7 cells infected with GV287 or GV287-SATB1
lentivirus, or BT-549 cells infected with GV115-SATB1-shRNA or
GV115-NC-shRNA lentivirus, were washed in PBS and then injected
into the mammary fat pad of 5-week-old female SCID mice
anesthetized with pentobarbital sodium (Sigma-Aldrich) administered
via intraperitoneal injection (40 mg/kg). To test the success rates
of engraftment, 10 mice of each group were respectively injected
with 103, 104 and 105 cells, and
to test engraftment size, 6 mice of each group were injected with
106 cells. The mice (20–30 g), obtained from SLRC
Laboratory Animals (Shanghai, China) were maintained in laminar
flow rooms under constant temperature and humidity and received
estradiol supplementation (0.4 mg/kg; Novo Nordisk, Copenhagen,
Denmark) every week subsequent to cell injection. Mice were
inspected for tumor appearance daily by observation and palpation
for 12 weeks subsequent to cell injection. Tumor volumes were
calculated using the formula V=L(W2)/2, in which L
indicates length and W indicates tumor width. At the conclusion of
the experiments, mice were sacrificed by cervical dislocation and
the presence of each tumor nodule was confirmed by necropsy.
Experimental protocols were approved by the Ethics Committee for
Animal Experimentation of Jiangxi Cancer Hospital (Nanchang,
China).
Immunoblotting
Cells and tumor tissues were lysed in a laemmli
buffer (Bio Rad Laboratories, Inc., Hercules, CA, USA), boiled and
loaded onto SDS (Thermo Fisher Scientific, Rockford, IL,
USA)-polyacrylamide gels (Energy Chemical, Shanghai, China).
Following electrophoresis (Bio Rad Laboratories, Inc.), proteins
were transferred onto nitrocellulose membranes (GenScript USA Inc.,
Piscataway, NJ, USA) using Trans-Blot SD Semi-Dry Electrophoretic
Transfer Cell (Bio-Rad Laboratories, Inc.). Blots were incubated in
Tris-buffered saline (TBS) (Spectrum Chemical (Shanghai) Co., Ltd)
blocking buffer containing 2% milk for 1~2 h at room temperature
and then with the mouse monoclonal anti-human antibodies against
Notch1 (sc-373891), Hes1 (sc-166410), Snail1 (sc-271977) and Twist1
(sc-81417), as well as rabbit anti-human polyclonal antibodies
against SATB1 (sc-28676) and Notch4 (sc-5594) (Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA) diluted 1:200 in TBS with
Tween (TBST; containing 0.1% Tween-20 and 2% bovine serum albumin;
Spectrum Chemical (Shanghai) Co., Ltd) overnight at 4°C.
Subsequently, blots were washed and incubated with the appropriate
secondary antibodies (goat polyclonal anti-mouse IgG, sc-2005, and
goat polyclonal anti-rabbit IgG, sc-2004; Santa Cruz Biotechnology
Inc.) in TBST at a 1:100–1:200 dilution ratio and detected using
the BeyoECL Plus Western Blotting Detection System (Beyotime,
Haimen, China), in accordance with the manufacturer’s
instructions.
Histology and immunohistochemistry
Tumor tissues were fixed in 10% neutral buffered
formalin (pH 7.4; Spectrum Chemical (Shanghai) Co., Ltd), embedded
in paraffin (ApexBio Technology LLC, Houston, TX, USA), cut into
5-μm sections using a CUT6062 automatic paraffin slicing machine
(SLEE Medical GmbH, Mainz, Germany) and stained with hematoxylin
and eosin (Boster Biological Engineering Co., Ltd, Wuhan, China).
For immunostaining, the sections were deparaffinized, rehydrated
and stained with the anti-Ki67 antibody (Santa Cruz Biotechnology,
Inc.) and VECTASTAIN Elite ABC Kit (Vector Laboratories,
Burlingame, CA, USA) according to the manufacturer’s instructions.
For each slide examined, 1,000 cells were counted from 6 fields
(randomly selected from 16 microscopic fields) at ×20
magnification, and the percentage of Ki67-positive cells was
calculated from the number of total cells. The number of
Ki67-positive tumor cells in 100 tumor cells determined the Ki67
proliferation index.
Statistical analysis
Statistical analysis was performed using SPSS,
version 19.0 (IBM SPSS, Armonk, NY, USA). Fisher’s exact test was
used to determine associations between SATB1 expression and the
success rates of engraftment. The differences in the means of the
groups were analyzed with one-way analysis of variance or
independent-samples t-test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Knockdown or overexpression of SATB1
reduces or increases the capacity for mammosphere formation in
breast cancer cells, respectively
To examine the role of SATB1 in maintaining the BCSC
population, the lentivirus GV287-SATB1 was selected to overexpress
SATB1 in MCF-7 breast cancer cells, while the shRNA-lentivirus
GV115-SATB1-shRNA was selected to knockdown SATB1 in BT-549 cells.
SATB1 overexpression or knockdown was confirmed by western blotting
(Fig. 1A and B). When the MCF-7
cells transfected with GV287-SATB1 were cultured, the size and
number of mammospheres significantly increased compared with the
controls (Fig. 1C–E). In BT-549
cells transfected with GV115-SATB1-shRNA, the size and number of
mammospheres was significantly reduced compared with the controls
(Fig. 1F–H).
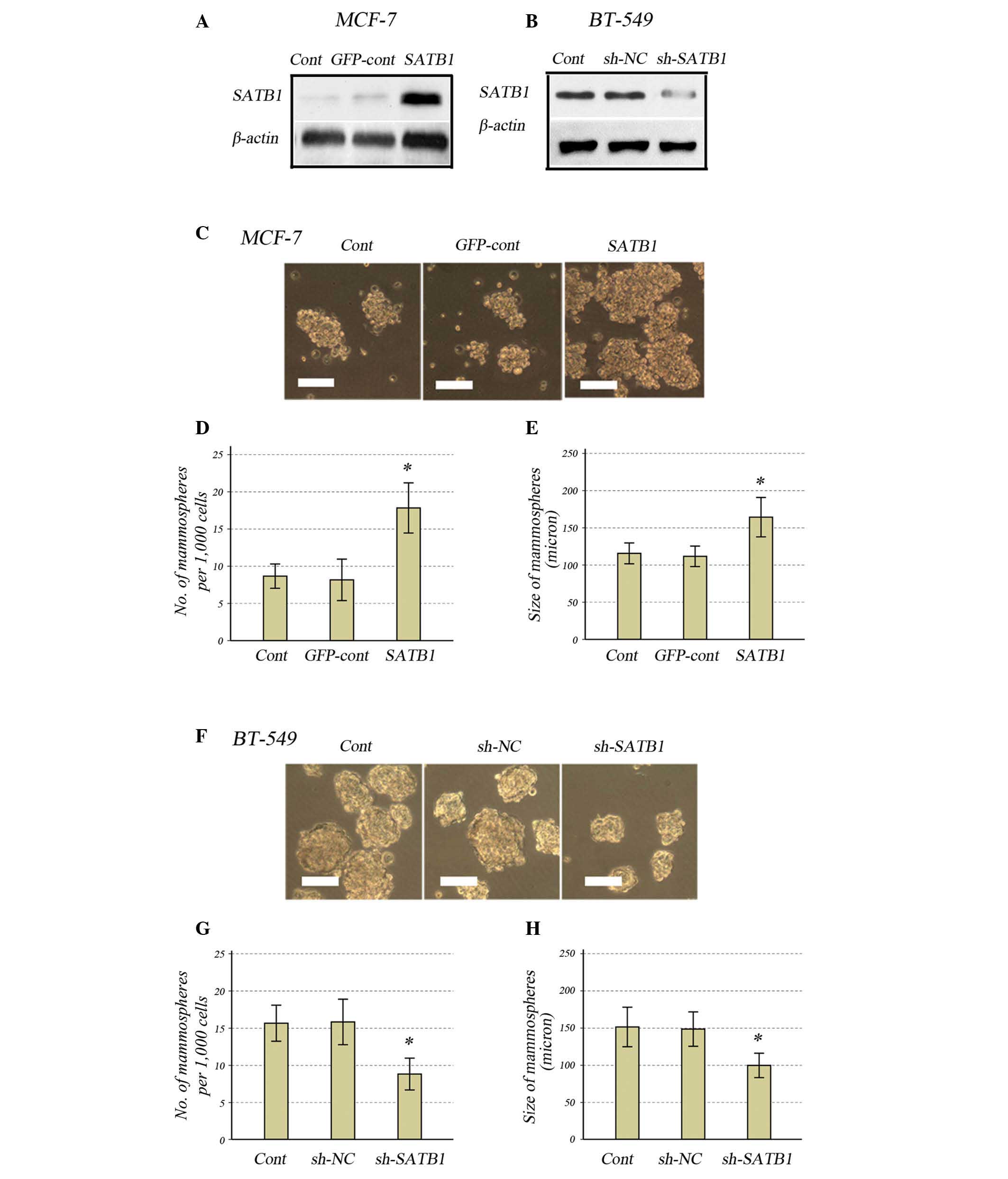 | Figure 1Effects of SATB1 expression on
mammosphere formation in human breast cancer cell lines. Western
blot analysis for SATB1 expression in (A) MCF-7 and (B) BT-549
cells. (C) Light microscope images of the MCF-7 cells
(magnification, ×200; scale bar, 10 μm). (D) Number of and (E) size
of mammospheres in SATB1-expressing MCF-7 cells compared with the
controls. (F) Light microscope images of the BT-549 cells
(magnification, ×200; scale bar, 10 μm). (G) Number of and (H) size
of mammospheres in SATB1-knockdown BT-549 cells compared with the
controls. All data are presented as the mean ± standard deviation
of two independent experiments in triplicate, *P<0.01
vs. controls. SATB1, special AT-rich sequence-binding protein-1;
cont, control; GFP, green fluorescent protein; sh, small hairpin;
NC, negative control. |
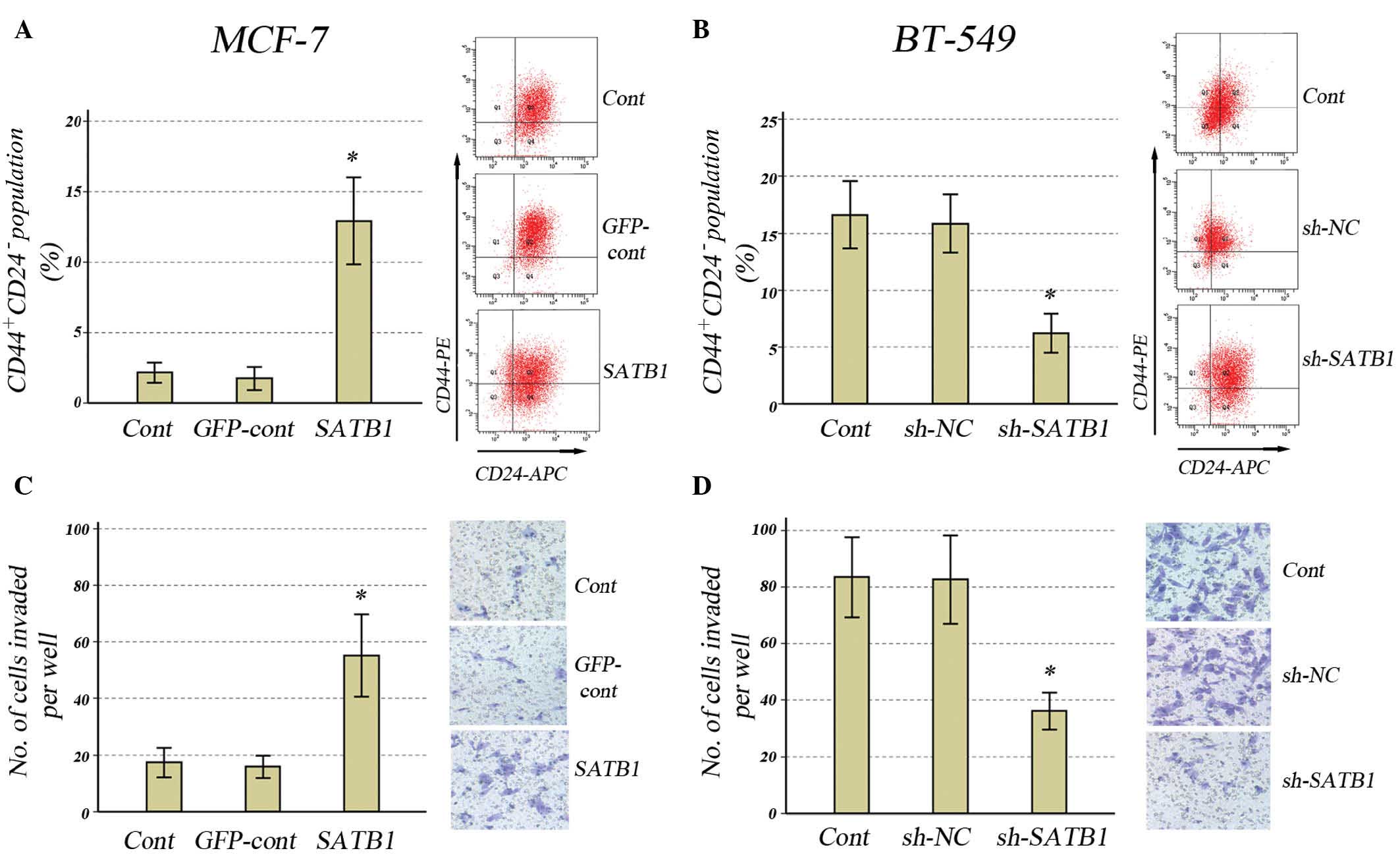
Effect of SATB1 expression on the
CD44+/CD24− population and tumor invasiveness
in breast cancer cells
In order to further determine the effects of SATB1
expression on the cancer stem cell population, the effects of SATB1
overexpression in MCF-7 cells and SATB1 knockdown in BT-549 cells
on CD44+/CD24− populations were investigated.
In MCF-7 cells transfected with GV287-SATB1, SATB1 overexpression
resulted in a significant increase in the
CD44+/CD24− population (Fig. 2A). In BT-549 cells transfected with
GV115-SATB1-shRNA, downregulation of SATB1 expression resulted in a
significant reduction in the CD44+/CD24−
population (Fig. 2B).
It has previously been suggested that tumor invasion
and metastasis may be mediated by the cancer stem cell population
(16,17), thus, the Matrigel-invading ability
of MCF-7 cells infected with GV287-SATB1 or BT-549 cells infected
with GV115-SATB1-shRNA was investigated. Overexpression of SATB1
resulted in a significant increase in invasiveness compared with
control groups in the MCF-7 cells (Fig. 2C), whilst downregulation of SATB1
resulted in a significant reduction in invasiveness compared with
control groups in BT-549 cells (Fig.
2D).
Knockdown or overexpression of SATB1
reduces or increases Notch1, Notch4, Hes1, Snail1 and Twist1
expression levels, respectively
To determine whether SATB1 affects Notch signaling
pathways, the expression levels of genes involved in stem cell
behavior, including Notch1, Notch4, Hes1, Snail1 and Twist1, were
investigated in MCF-7 and BT-549 cells transfected with GV287-SATB1
and GV115-SATB1-shRNA, respectively. Overexpression of SATB1
significantly increased the expression levels of Notch1, Notch4,
Hes1, Snail1 and Twist1 compared with control groups in MCF-7 cells
(Fig. 3A), which can be observed
by the results of the western blot analysis (Fig. 3B). Conversely, the downregulation
of SATB1 reduced the expression of these genes compared with
control groups in BT-549 cells (Fig.
3C), and can be observed by the results of the western blot
analysis (Fig. 3D).
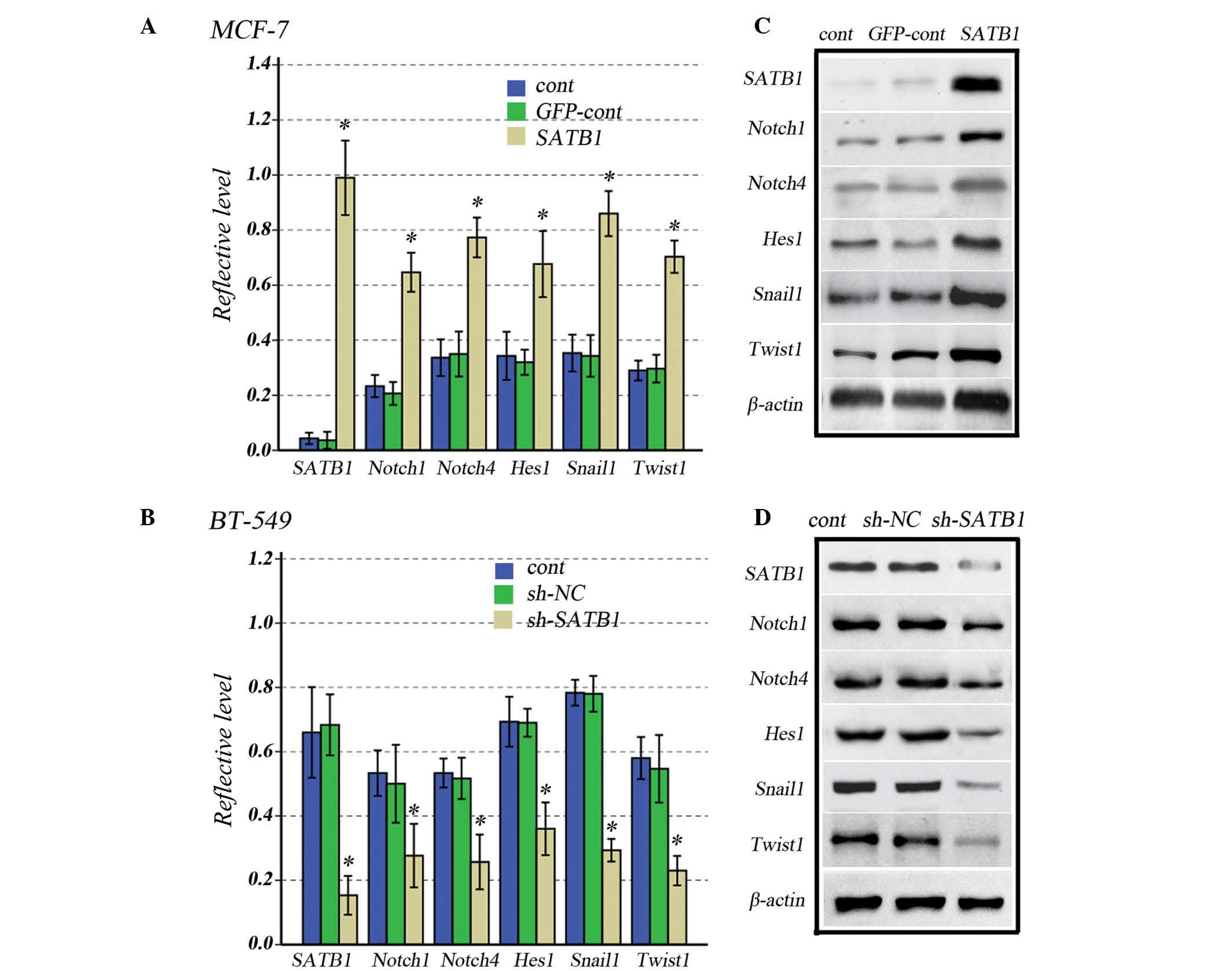 | Figure 3Effects of increased or reduced SATB1
expression on Notch1, Notch4, Hes1, Snail1 and Twist1 expression
levels in human breast cancer cells. (A) Graph and (B)
representative western blot of SATB1, Notch1, Notch4, Hes1, Snail1
and Twist1 expression levels in SATB1-overexpressing and control
MCF-7 cells. (C) Graph and (D) representative western blot of
SATB1, Notch1, Notch4, Hes1, Snail1 and Twist1 expression levels in
SATB1-knockdown and control BT-549 cells. All data are presented as
the mean ± standard deviation of experiments in triplicate;
*P<0.001 vs. controls. SATB1, special AT-rich
sequence-binding protein-1; cont, control; GFP, green fluorescent
protein; sh, small hairpin; NC, negative control. |
This suggests that SATB1 overexpression increases
the size of the stem cell pool, and also activates Notch signaling
pathways, increasing the expression levels of Snail1 and Twist1,
which drive EMT in breast cancer cells.
Effects of SATB1 expression on stem cell
population, tumorigenicity and Notch1, Notch4, Hes1, Snail1 and
Twist1 expression levels in breast cancer cells in vivo
A subcutaneous serial dilution transplantation
experiment was performed in female SCID mice using MCF-7 and BT-549
cells transfected with GV287-SATB1 and GV115-SATB1-shRNA,
respectively. A notable observation was that following the
subcutaneous implantation of 1×104 and 1×105
MCF-7 cells, SATB1 overexpression in the cells significantly
increased the success of engraftments (Fig. 4A), and subsequent to the
implantation of 1×106 MCF-7 cells, SATB1 overexpression
significantly increased tumor size after 21 days (Fig. 4B). Following the subcutaneous
implantation of 1×103 and 1×104 BT-549 cells,
SATB1 depletion in the cells significantly reduced the success of
engraftments (Fig. 4C), while
subsequent to the subcutaneous implantation of 1×106
BT-549 cells, SATB1 knockdown significantly reduced tumor size
after 21 days (Fig. 4D).
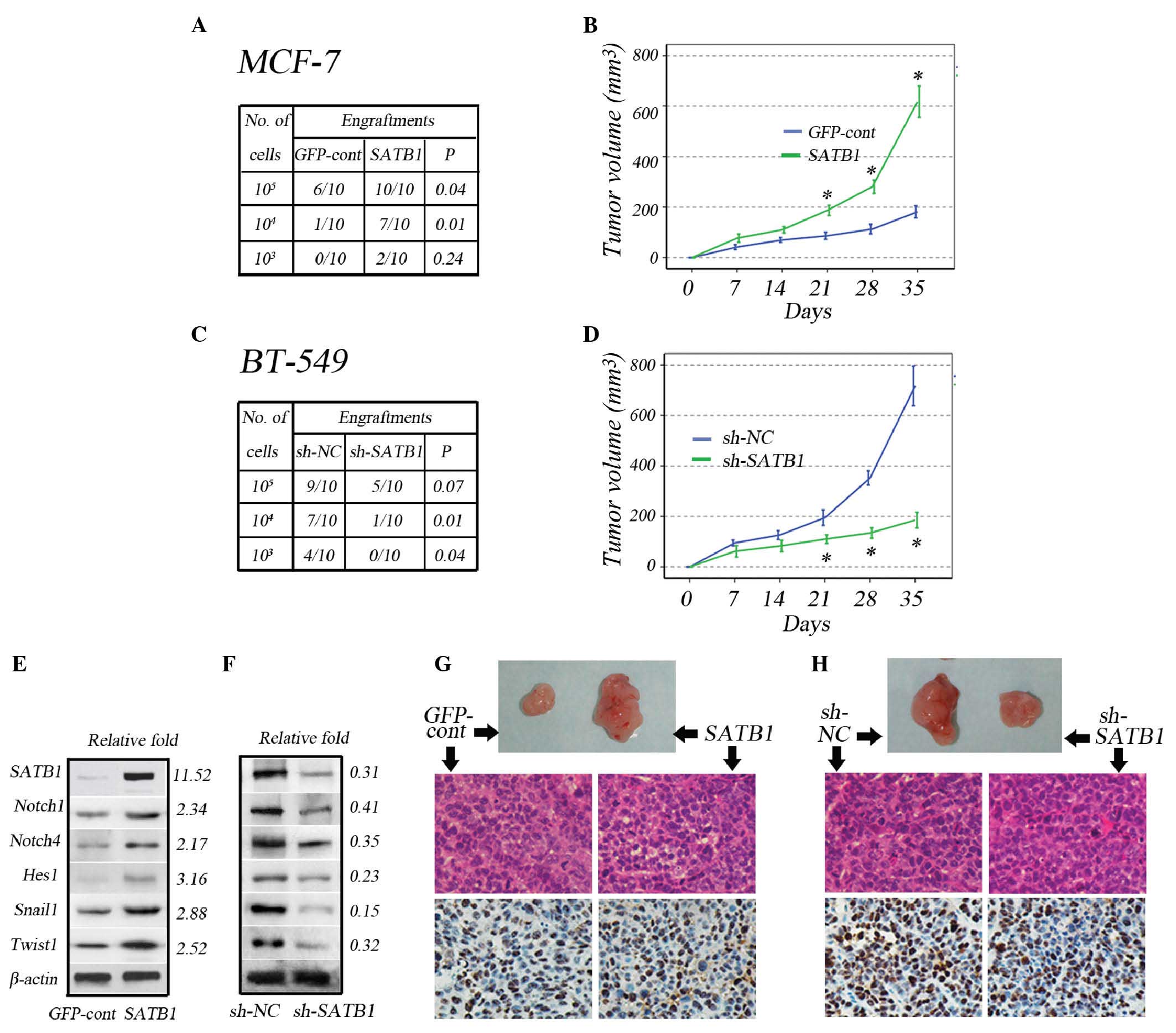 | Figure 4Effects of SATB1 expression on
tumorigenicity and expression levels of Notch1, Notch4, Hes1,
Snail1 and Twist1 in vivo. (A) The number of successful
tumor engraftments correlated with the number of MCF-7-GFP-cont and
MCF-7-SATB1 cells injected during serial dilution. (B) Tumor growth
size subsequent to injection of 1×106 MCF-7-GFP-cont and
MCF-7-SATB1 cells. (C) The number of successful tumor engraftments
correlated with the number of BT-549-sh-NC and BT-549-sh-SATB1
cells injected during serial dilution. (D) Tumor growth size
subsequent to injection of 1×106 BT-549-sh-NC and
BT-549-sh-SATB1 cells. Representative immunoblot analysis and
expression fold-change of SATB1, Notch1, Notch4, Hes1, Snail1 and
Twist1 in (E) MCF-7-GFP-cont and MCF-7-SATB1-derived and (F)
BT-549-sh-NC and BT-549-sh-SATB1-derived tumors relative to that of
the control group. Representative images of xenografts, hematoxylin
and eosin staining and immunohistochemical staining for Ki-67
xenografts, derived from injection of (G) 106
MCF-7-GFP-cont and MCF-7-SATB1 cells (magnification, ×10) and (H)
106 BT-549-sh-cont and BT-549-sh-SATB1 cells
(magnification, ×10). Quantitative data are presented as the mean ±
standard deviation for six tumors in each group;
*P<0.001 vs. controls. SATB1, special AT-rich
sequence-binding protein-1; GFP, green fluorescent protein; cont,
control; sh, small hairpin; NC, negative control; relative fold,
expression fold-change relative to the control group. |
In agreement with the in vitro results,
western blot analysis of the tumors revealed that in vivo
overexpression of SATB1 in MCF-7 cells increased the expression
levels of Notch1, Notch4, Hes1, Snail1 and Twist1 (Fig. 4E), whereas knockdown of SATB1 in
BT-549 cells reduced the expression levels (Fig. 4F). Immunohistochemical analyses of
the tumors revealed that the Ki67 proliferation index was
comparable between MCF-7-GFP and MCF-7-SATB1 tumors (Fig. 4G), despite an increase in tumor
size. In addition, the Ki67 proliferation index was comparable
between BT-549-GFP and BT-549-sh-SATB1 tumors, despite a reduction
in tumor size (Fig. 4H).
Discussion
Although SATB1 overexpression has been previously
demonstrated to increase breast cancer invasion and metastasis, its
effect on the stem cell population remains to be fully elucidated.
In the current study, SATB1 was identified to expand the cancer
stem cell population, accompanied by the activation of the Notch
signaling pathway, which promotes cancer stem cell self-renewal and
the expression of the Snail1 and Twist1 genes that drive EMT.
Overexpression of SATB1 in the breast cancer cell
line MCF-7 was demonstrated to increase the population, which was
measured by mammosphere formation and
CD44+/CD24− expression in vitro and
formation of tumors in SCID model mice (in vivo).
Downregulation of SATB1 in the breast cancer cell line BT-549
resulted in the opposite result. Previous studies have demonstrated
that the subpopulation of breast cancer cells exhibit increased
invasive properties (17). SATB1
overexpression increased the invasiveness of MCF-7 cells, and SATB1
knockdown reduced the invasiveness of BT-549 cells. These results
suggest that SATB1 expression in breast cancer may increase the
BCSC population, resulting in tumor progression.
Aberrant Notch signaling has been identified to be
associated with the development and progression of breast cancer
(18,19); the Notch signaling pathway
maintains the stemness of BCSCs (20). Previous studies have demonstrated
that Notch signaling activity increased in BCSCs, as demonstrated
by increased expression of the Notch receptors Notch1 and Notch4,
in addition to Hes-1, an immediate downstream target of Notch
signaling (8). In the current
study, SATB1 overexpression in MCF-7 cells was demonstrated to
significantly increase Notch1, Notch4 and Hes1 expression, whilst
its knockdown in BT-549 cells resulted in the opposite effect.
These data suggest that SATB1 mediates the BCSC population via the
Notch signaling pathway.
Additionally, it has been hypothesized that the
progression of the majority of carcinomas towards malignancy is
associated with EMT, during which breast cancer cells transition to
acquire stem cell-like properties (21,22).
In cancer EMT, signals emanate via factors, including transforming
growth factor-β, hepatocyte growth factor, epidermal growth factor
and hypoxia (23). The signals
then converge upon a limited set of transcriptional repressors,
including Snail1, Slug, ZEB1/2 and Twist1/2 (24,25).
One study suggested that Snail1 activity is required for EMT
initiation, whereas Twist1 is involved in the maintenance of EMT
during human breast cancer progression towards metastasis (13). In the current study, it was
observed that Snail1 and Twist1 were upregulated in MCF-7 cells
with SATB1 overexpression and inhibited in BT-549 cells with SATB1
knockdown. Therefore, in the present study, SATB1 expression was
established to affect the number of BCSCs, in addition to their
ability to propagate in conditions sustaining the undifferentiated
cell state.
In conclusion, the current study suggests that the
expression of SATB1 may increase the size of the BCSC population
via the activation of Notch signaling, which is required for
maintaining the stemness of BCSCs and increasing the expression
level of Snail1 and Twist1, which are required for EMT.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81060179 and
81260329). The authors would like to thank Dr. Jianjun Xioang at
the central laboratory of Jiujiang Medical College (Jiujiang,
China) for his technical assistance.
References
|
1
|
Youlden DR, Cramb SM, Dunn NA, Muller JM,
Pyke CM and Baade PD: The descriptive epidemiology of female breast
cancer: an international comparison of screening, incidence,
survival and mortality. Cancer Epidemiol. 36:237–248. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhou L, Jiang Y, Yan T, Di G, Shen Z, Shao
Z and Lu J: The prognostic role of cancer stem cells in breast
cancer: A meta-analysis of published literatures. Breast Cancer Res
Treat. 122:795–801. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Al-Hajj M, Wicha MS, Benito-Hernandez A,
Morrison SJ and Clarke MF: Prospective identification of
tumorigenic breast cancer cells. Proc Natl Acad Sci USA.
100:3983–3988. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ginestier C, Hur MH, Charafe-Jauffret E,
Monville F, Dutcher J, Brown M, Jacquemier J, Viens P, Kleer CG,
Liu S, et al: ALDH1 is a marker of normal and malignant human
mammary stem cells and a predictor of poor clinical outcome. Cell
Stem Cell. 1:555–567. 2007. View Article : Google Scholar
|
|
5
|
Liu X, Johnson S, Liu S, Kanojia D, Yue W,
Singh UP, Wang Q, Wang Q, Nie Q and Chen H: Nonlinear growth
kinetics of breast cancer stem cells: implications for cancer stem
cell targeted therapy. Sci Rep. 3:24732013.PubMed/NCBI
|
|
6
|
Kakarala M and Wicha MS: Cancer stem
cells: Implications for cancer treatment and prevention. Cancer J.
13:271–275. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Takebe N, Harris PJ, Warren RQ and Ivy SP:
Targeting cancer stem cells by inhibiting Wnt, Notch, and Hedgehog
pathways. Nat Rev Clin Oncol. 8:97–106. 2011. View Article : Google Scholar
|
|
8
|
Grudzien P, Lo S, Albain KS, Robinson P,
Rajan P, Strack PR, Golde TE, Miele L and Foreman KE: Inhibition of
Notch signaling reduces the stem-like population of breast cancer
cells and prevents mammosphere formation. Anticancer Res.
30:3853–3867. 2010.PubMed/NCBI
|
|
9
|
Harrison H, Farnie G, Howell SJ, Rock RE,
Stylianou S, Brennan KR, Bundred NJ and Clarke RB: Regulation of
breast cancer stem cell activity by signaling through the Notch4
receptor. Cancer Res. 70:709–718. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mani SA, Guo W, Liao MJ, et al: The
epithelial-mesenchymal transition generates cells with properties
of stem cells. Cell. 133:704–715. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Creighton CJ, Chang JC and Rosen JM:
Epithelial-mesenchymal transition in tumor-initiating cells and its
clinical implications in breast cancer. J Mammary Gland Biol
Neoplasia. 15:253–260. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Drasin DJ, Robin TP and Ford HL: Breast
cancer epithelial-to-mesenchymal transition: examining the
functional consequences of plasticity. Breast Cancer Res.
13:2262011. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tran DD, Corsa CA, Biswas H, Aft RL and
Longmore GD: Temporal and spatial cooperation of Snail1 and Twist1
during epithelial-mesenchymal transition predicts for human breast
cancer recurrence. Mol Cancer Res. 9:1644–1657. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Han HJ, Russo J, Kohwi Y and
Kohwi-Shigematsu T: SATB1 reprogrammes gene expression to promote
breast tumour growth and metastasis. Nature. 452:187–193. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ordinario E, Han HJ, Furuta S, et al: ATM
suppresses SATB1-induced malignant progression in breast epithelial
cells. PLoS One. 7:e517862012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wicha MS: Cancer stem cells and
metastasis: lethal seeds. Clin Cancer Res. 12:5606–5607. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sheridan C, Kishimoto H, Fuchs RK,
Mehrotra S, Bhat-Nakshatri P, Turner CH, Goulet R Jr, Badve S and
Nakshatri H: CD44+/CD24− breast cancer cells exhibit enhanced
invasive properties: an early step necessary for metastasis. Breast
Cancer Res. 8:R592006. View
Article : Google Scholar
|
|
18
|
Pece S, Serresi M, Santolini E, Capra M,
Hulleman E, Galimberti V, Zurrida S, Maisonneuve P, Viale G and Di
Fiore PP: Loss of negative regulation by Numb over Notch is
relevant to human breast carcinogenesis. J Cell Biol. 167:215–221.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Stylianou S, Clarke RB and Brennan K:
Aberrant activation of notch signaling in human breast cancer.
Cancer Res. 66:1517–1525. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Han J, Hendzel MJ and Allalunis-Turner J:
Notch signaling as a therapeutic target for breast cancer
treatment? Breast Cancer Res. 13:2102011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Battula VL, Shi Y, Evans KW, et al:
Ganglioside GD2 identifies breast cancer stem cells and promotes
tumorigenesis. J Clin Invest. 122:2066–2078. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Battula VL, Evans KW, Hollier BG, et al:
Epithelial-mesenchymal transition-derived cells exhibit
multilineage differentiation potential similar to mesenchymal stem
cells. Stem Cells. 28:1435–1445. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Peinado H, Olmeda D and Cano A: Snail, Zeb
and bHLH factors in tumour progression: an alliance against the
epithelial phenotype? Nat Rev Cancer. 7:415–428. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yang MH, Hsu DS, Wang HW, et al: Bmi1 is
essential in Twist1-induced epithelial-mesenchymal transition. Nat
Cell Biol. 12:982–992. 2010. View
Article : Google Scholar : PubMed/NCBI
|