Introduction
Nasopharyngeal carcinoma (NPC) is one of the most
common types of cancer derived from epithelial cells located in the
nasopharynx (1). NPC has a
distinct epidemiology and distribution, with the highest incidence
in southern China and Southeast Asia (2). Radiotherapy is the primary
therapeutic strategy for patients with NPC (3). Although numerous patients with NPC
undergo radiotherapy treatment, advanced NPC patients tend to
experience therapy failure due to local recurrence and distant
metastasis (4). The poor prognosis
of NPC patients may be due to its radioresistance (5). An improved understanding of the
molecular mechanisms that contribute to the radioresistance of NPC
may provide novel therapeutic strategies and therefore improved
clinical outcomes.
microRNAs (miRNAs) are a class of small (19–25
nucleotides) endogenous non-coding RNA molecules that regulate gene
expression at the post-transcriptional level by translational
arrest or messenger RNA (mRNA) cleavage. These effects are achieved
via miRNA-binding to the 3′-untranslated region (3′UTR) of their
target mRNA, which decreases expression of the associated protein
(6). An increasing body of
evidence suggests that miRNAs have crucial roles in numerous
biological processes, including cellular differentiation,
proliferation, apoptosis, autophagy and metabolism (7,8). The
dysregulation of miRNAs may be associated with numerous human
diseases, including certain types of cancer (9). Specific miRNAs function as tumor
suppressors or oncogenes and are influential in tumor progression
and therapeutic resistance (10).
Of note, recent studies have indicated an association between the
expression of certain miRNAs, including miR-608 and miR-29c, and
the success of radiotherapy treatment, particularly in NPC.
(11,12). Further studies have indicated that
miR-101 is downregulated in numerous types of cancer, including
gastric (13), lung (14) and colon cancer (15), and a loss of miR-101 expression is
involved in carcinogenesis (16)
and angiogenesis (17). Studies
have also indicated that the ectopic expression of miR-101 is able
to sensitize non-small cell lung cancer cells to radiation by
targeting DNA-dependent protein kinase and ataxia telangiectasia
mutated (ATM) (18). However, the
association between miR-101 and the modulation of radioresistance
in NPC remains to be elucidated.
The present study aimed to elucidate the function of
miR-101 in NPC by analyzing miR-101 expression in NPC cell lines
and investigating the effects of ectopic expression on NPC-cell
proliferation and radiosensitivity. Additionally, the present study
aimed to identify functional targets of miR-101 in order to
elucidate the mechanism by which it exerts its effects in NPC and
thereby propose a strategy for enhancing NPC cell radiosensitivity
and improving treatment of NPC.
Materials and methods
Cell culture
Three NPC cell lines (CNE-2, 5-8F and 6-10B) were
provided by Prof. Xia Yunfei (Sun Yat-sen University Cancer Center,
Guangzhou, China) and maintained in the State Key Laboratory of
Oncology in South China (Guangzhou, China). The CNE-1 and NP69 cell
lines were maintained in the lab at Nanfang Hospital, Southern
Medical University (Guangzhou, China) and were purchased from the
Cell Bank of Sun Yat-Sen University in 2010. The passage number of
all five cell lines used in the present study was <20. All of
the cell lines were tested against mycoplasmic infection. CNE-1,
CNE-2, 5-8F and 6-10B cells were maintained in RPMI-1640 medium
(Invitrogen Life Technologies, Carlsbad, CA, USA) supplemented with
10% fetal bovine serum (FBS; Invitrogen Life Technologies), 100
U/ml penicillin (Sigma-Aldrich, St. Louis, MO, USA) and 50 μg/ml
streptomycin (Sigma-Aldrich). NP69 cells were cultured in
keratinocyte/serum-free medium (Invitrogen Life Technologies)
supplemented with bovine pituitary extract (BD Biosciences, San
Jose, CA, USA). All cell lines were incubated at 37°C in a
humidified atmosphere of 5% CO2.
Quantitative polymerase chain reaction
(qPCR)
Total RNA was isolated from cells using Trizol
reagent (Invitrogen) according to the manufacturer’s instructions.
The reverse-transcription and PCR primers for miR-101 and U6 were
purchased from Ribobio (Guangzhou, China). The PCR primers for
STMN1 were forward, 5′-CCTCTGTTTGGCGCTTTTGTGCG-3′ and reverse,
5′-GGCACGCTTCTCCAGTTCTTTCACC-3′. The PCR primers for β-actin were
forward, 5′-TCGACAACGGCTCCGGCAT-3′ and reverse,
5′-AAGGTGTGGTGCCAGATTTTC-3′. The cDNA library was synthesized using
the PrimeScript RT reagent kit (Takara Bio, Inc., Dalian, China).
For mature miRNA quantification, cDNA was generated using specific
stem-loop universal primers. Aliquots of cDNA were amplified for 40
cycles, which were perofmred as follows: Denaturing at 95°C for 5
sec, annealing at 60°C for 34 sec and extension at 60°C for 1 min.
Real-time qPCR for miRNA and mRNA was performed using
SYBR® Premix Ex Taq II (Takara) and quantified in an ABI
7500 Sequence Detection system (Perkin Elmer/Applied Biosystems).
Either U6 or β-actin were used as an internal control.
Oligonucleotide and small interfering RNA
(siRNA) transfection
miR-101 mimic, miRNA mimic negative control
oligonucleotides, STMN1 siRNA and siRNA negative control were all
purchased from Ribobio. Oligonucleotide and siRNA transfection were
performed using Lipofectamine® 2000 reagent (Invitrogen)
according to the manufacturer’s instructions.
Cell proliferation assay
Cell proliferation was measured using the MTT dye
reduction method (19). Briefly,
48 h following transfection, cells were seeded in 96-well plates at
a low density (2×103 cells/well) in RMPI-1640 medium
supplemented with 10% FBS and incubated for 1–5 days. Following
incubation, 50 μl MTT solution (2 mg/ml; Sigma-Aldrich) was added
to each well, and the cells were incubated for a further 2 h at
37°C. The media containing MTT solution was removed, and the dark
blue crystals were dissolved by the addition of 100 μl
dimethylsulfoxide. The absorbance value was measured with a
microplate reader (SpectraMax M5; Molecular Devices, Sunnyvale, CA,
USA) at a test and reference wavelength of 570 nm. The percentage
of growth was determined relative to untreated controls. Each
experiment was performed ≥three times with triplicate samples.
Clonogenic survival assays
CNE-2 and 5-8F cells were pretreated by either
miR-101 mimic or STMN1 siRNA transfection for 48 h and subsequently
seeded onto six-well plates in triplicate at specific cell
densities, followed by exposure to the indicated doses of radiation
(0, 2, 4, 6 or 8 Gy) using 6 MV X-rays generated from linear
accelerators (Varian 2300EX; Varian, Palo Alto, CA, USA) at a dose
rate of 3 Gy/min. Following 10–14 days of incubation at 37°C, the
cells were fixed using 100% methanol and stained using 1% crystal
violet (Sigma-Aldrich). Colonies containing ≥50 normal-appearing
cells were counted via microscopic inspection (Olympus IX71;
Olympus, Tokyo, Japan). The surviving fraction was calculated as
described previously (20). The
multi-target single-hit model was fitted to the data to generate
survival curves using the formula:
SF=1−(1−e−D/D0)N. The sensitization
enhancement ratio at a survival fraction of 10% (SER10) was
subsequently calculated. Each experiment was independently
performed ≥three times.
Immunofluorescent staining for
γ-H2AX
Forty-eight hours following transfection with
miR-101 mimic or miRNA mimic negative control, 1×105
cells were seeded in chamber slides and incubated overnight. The
cells were subsequently exposed to 6 Gy irradiation (IR).
Twenty-four hours following IR, the cells were fixed in 4%
paraformaldehyde (Sigma-Aldrich), permeabilized in 0.1% Triton
X-100 (Sigma), blocked in 2% bovine serum albumin (Roche,
Stockholm, Sweden) and incubated with a primary antibody against
γ-H2AX (Abcam, San Francisco, CA, USA) overnight at 4°C. The
primary antibody was subsequently washed off, and a secondary
antibody conjugated to fluorescein isothiocyanate (Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA) was applied to the
slides. Cells were washed with phosphate-buffered saline
(Sigma-Aldrich) and counterstained with DAPI (Invitrogen Life
Technologies). The γ-H2AX foci were observed under a fluorescence
microscope (Olympus BX51, Olympus). For each group, the γ-H2AX foci
were counted in ≥50 cells.
Antibodies and western blot analysis
For the western blot analysis, cells were lysed in
radio-immunoprecipitation assay buffer (Cell Signaling Technology,
Beverly, MA, USA) containing phosphatase and proteinase inhibitor
cocktails (Sigma-Aldrich). The protein concentrations were
determined using a bicinchoninic acid protein assay kit (Pierce
Biotechnology, Rockfold, IL, USA). Equal amounts of total protein
were resolved via SDS-PAGE (Bio-Rad, Hercules, CA, USA), and the
proteins were transferred onto polyvinylidene difluoride membranes
(Bio-Rad). The membranes were blocked in 5% non-fat milk for 1 h at
room temperature and then incubated overnight at 4°C with primary
antibodies to anti-STMN1 (Abcam), anti-LC3B (Novus Biological,
Littleton, CO, USA), anti-p62/SQSTM1 (Cell Signaling Technology) or
anti-β-actin (ProteinTech, Chicago, IL, USA). Following three
washes, the membranes were incubated with secondary antibodies
[species-specific horseradish peroxidase(HRP)-conjugated] for 1 h
at room temperature. The immunoreactive bands were visualized with
the Immobilon Western chemiluminescent HRP substrate (Millipore,
Temecula, CA, USA). Each experiment was independently performed
≥three times.
Generation of stable green fluorescent
protein (GFP)-light chain (LC)3-expressing cells
The lentiviral vector containing the GFP-LC3
reporter was purchased from GenePharma (Shanghai, China). CNE-2
cells were infected with recombinant lentivirus and purified using
flow cytometry (BD Biosciences) to generate populations that stably
expressed GFP-LC3.
Luciferase reporter assay
The STMN1 wild-type (wt) and mutant (mut) 3′UTRs,
which contain the putative miR-101 binding site, were created and
cloned into the Renilla luciferase vector (pLUC-REPORT vector;
Promega, Madison, WI, USA). For the luciferase reporter assay,
CNE-2 and 5-8F cells were co-transfected with a luciferase reporter
vector (either pLUC-3′UTR-STMN1 or pLUC-3′UTR-mut-STMN1) and
negative control miRNA or the miR-101 mimic. Forty-eight hours
following transfection, the cells were assayed for luciferase
activity using the Dual-Luciferase assay kit (Promega) according to
the manufacturer’s instructions. For each sample, the relative
luciferase activity was normalized to firefly luciferase activity.
Three independent experiments were performed in triplicate.
STMN1 rescue experiments
The pCMX-IRES2-eGFP-STMN1 (PCA-STMN1) plasmid and
empty vector were synthesized by GenePharma (Shanghai, China).
CNE-2 cells were co-transfected with miR-101 mimic or miRNA mimic
negative control and with pCMX-IRES2-eGFP-STMN1 plasmid or the
empty vector. Forty-eight hours following transfection, the cells
were analyzed for proliferation and clonogenic survival as
described. The cells were also analyzed for radiation-induced
autophagy activity by western blotting and confocal microscopy
(Olympus FV1000; Olympus). STMN1 expression was verified by western
blot analysis.
Statistical analysis
All values were expressed as the mean ± standard
deviation and were obtained from experiments that were repeated
≥three times. Significant differences between the means were
measured using a two-tailed unpaired Student’s t-test or one-way
analysis of variance. All statistical analyses were performed using
SPPS version 13.0 software (SPSS Inc., Chicago, IL, USA). P<0.05
was considered to indicate a statistically significant difference
between values.
Results
miR-101 is downregulated in NPC cell
lines and affects the radiation response of NPC cells
In the present study, the expression of miR-101 in
four NPC cell lines and the human immortalized nasopharyngeal
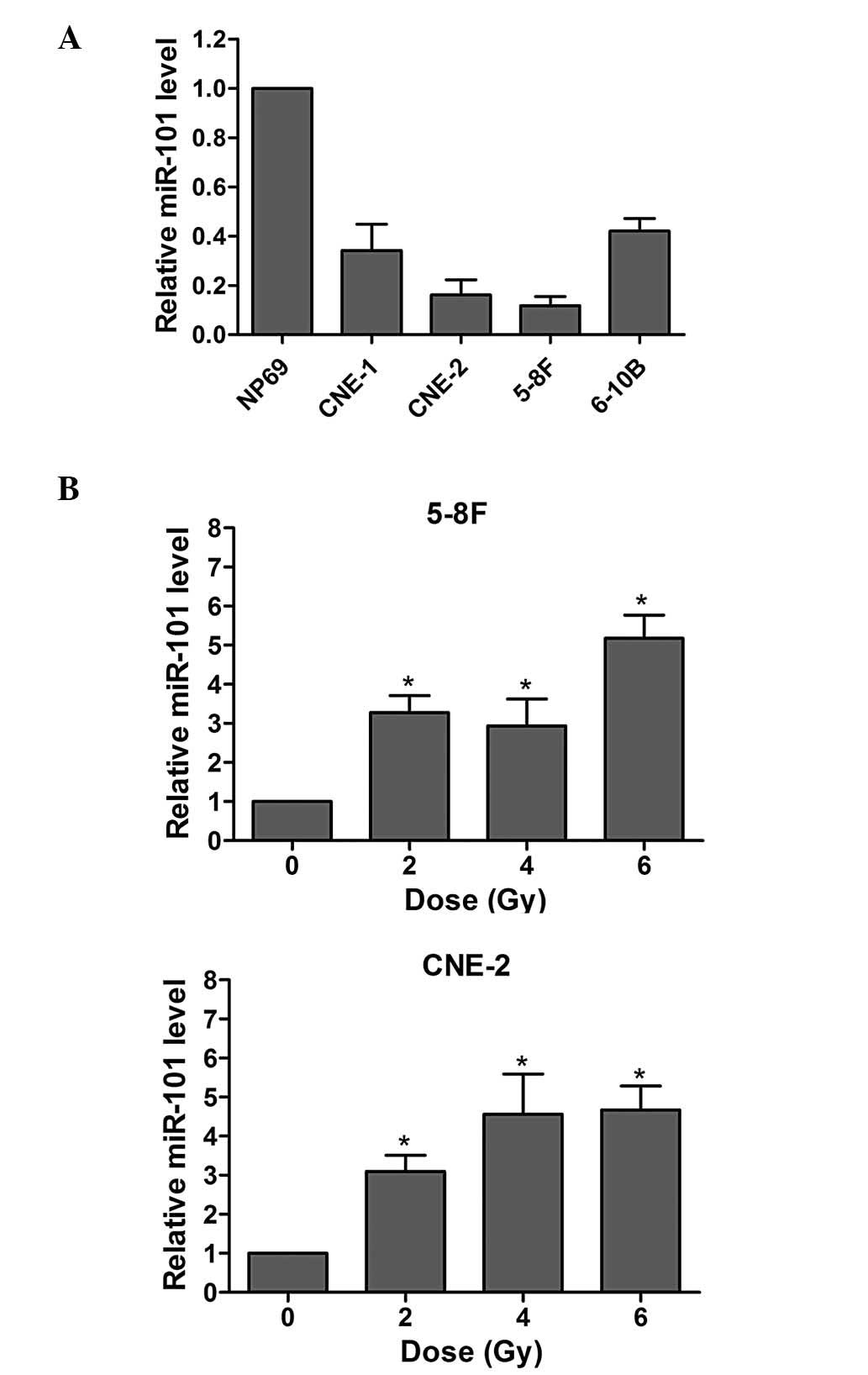
epithelial cell line NP69 was investigated. As indicated in
Fig. 1A, the miR-101 levels were
significantly decreased in all four NPC cell lines (P<0.01),
particularly in the 5-8F and CNE-2 cell lines.
The effects of radiation on miR-101 expression of
NPC cells were also examined. As demonstrated in Fig. 1B, the levels of IR-induced miR-101
expression in both cell lines increased upon IR. These results
indicated that miR-101 may influence the IR response of NPC
cells.
miR-101 suppresses NPC-cell viability and
sensitizes NPC cells to radiation
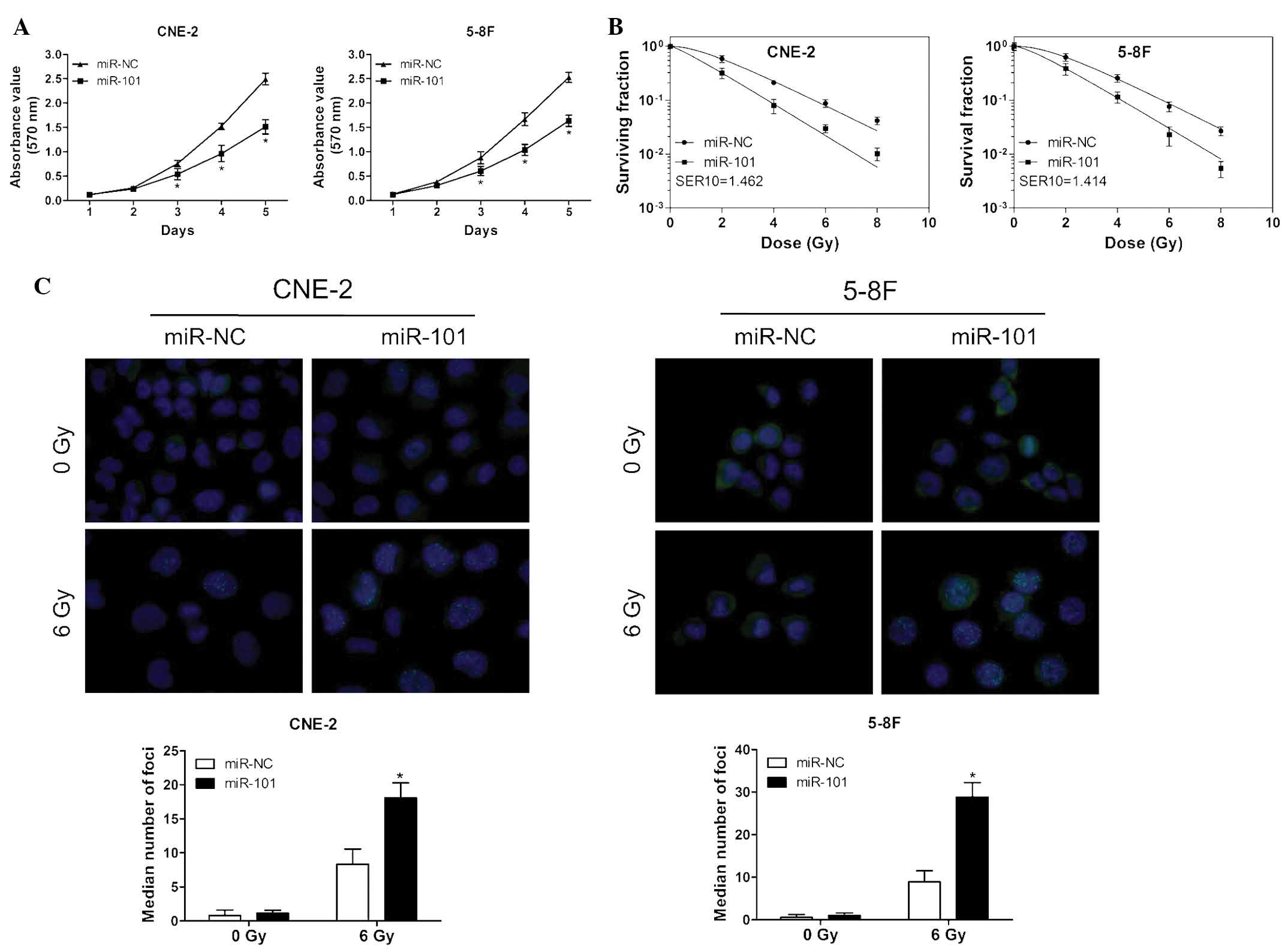
To evaluate the effects of miR-101 overexpression on
the cell viability and radiosensitivity of NPC cells, an MTT assay
and a clonogenic survival assay were performed following
transfection of CNE-2 and 5-8F cells with miR-101 mimic or negative
control. The ectopic expression of miR-101 significantly reduced
the proliferation of NPC cells compared with that of the controls
(Fig. 2A). Moreover, the survival
fraction of cells transfected with miR-101 mimics was significantly
decreased following various doses of irradiation compared with that
of cells transfected with negative controls (Fig. 2B).
To assess the effect of miR-101 on DNA damage in NPC
cells, the number of γ-H2AX foci following IR was measured. The
γ-H2AX foci-number is an established molecular marker of DNA damage
and repair (21). As indicated in
Fig. 2C, the ectopic expression of
miR-101 led to a markedly increased persistence of γ-H2AX foci 24 h
post-IR compared with that of the control groups. These results
suggested that miR-101 enhanced the radiosensitivity of NPC cells
in a manner that may be associated with the suppression of cell
viability and persistence of DNA damage.
STMN1 is a direct target of miR-101 and
is involved in NPC-cell radioresistance and growth
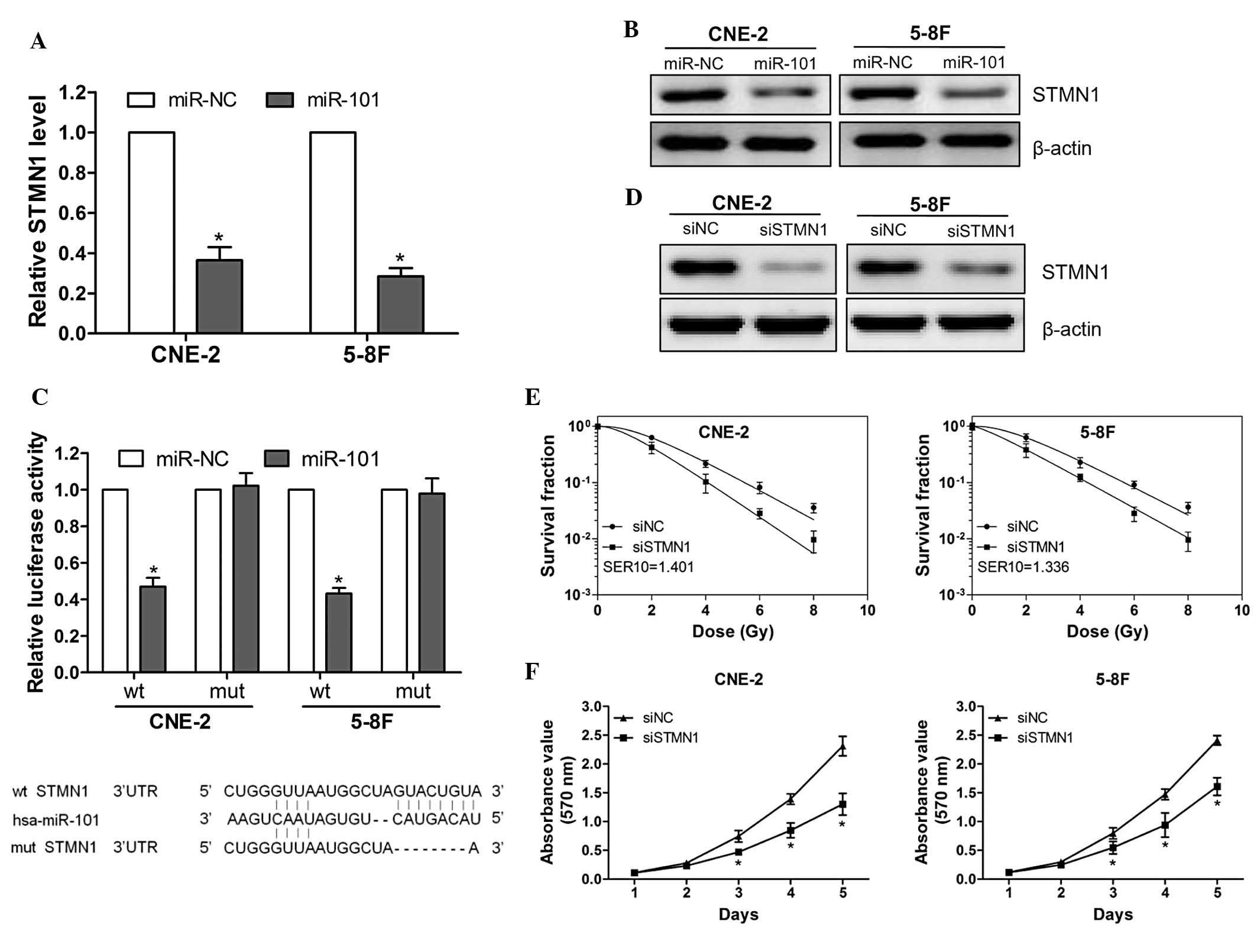
To investigate the molecular mechanism by which
miR-101 increased the radiosensitivity of NPC cells, STMN1 was
identified as a potential target of miR-101 based on the three
publicly available databases [TargetScan (http://www.targetscan.org), miRanda (http://www.microrna.org/microrna/home.do) and Pictar
(http://pictar.mdc-berlin.de/)] and a
previous study (7). The ectopic
expression of miR-101 was able to significantly suppress the mRNA
and protein expression of STMN1 (Fig.
3A and B). Subsequently, luciferase reporter vectors that
contained wild-type or mutant miR-101 target sequences of the STMN1
3′UTR were constructed (Fig. 3C,
lower panel) and a luciferase reporter assay was performed to
determine whether STMN1 was a direct target of miR-101. It was
demonstrated that the overexpression of miR-101 significantly
suppressed the luciferase activity of the wt 3′UTR of STMN1 but not
the mut reporter gene (Fig. 3C,
upper panel), indicating the specificity of miR-101 to target the
STMN1 3′UTR. These results indicated that STMN1 is a direct target
of miR-101 in NPC cells.
To confirm that the miR-101-enhanced
radiosensitivity is due to the direct targeting of STMN1, CNE-2 and
5-8F cells were transfected with STMN1 siRNA or control siRNA.
Knocking down the expression of STMN1 significantly enhanced the
radiosensitivity of CNE-2 and 5-8F cells (Fig. 3D and E). Furthermore, the
proliferation assay indicated that silencing STMN1 expression
significantly suppressed CNE-2 and 5-8F cell-growth (Fig. 3F). These data demonstrated that
miR-101 enhanced radiosensitivity by directly targeting STMN1.
Restoration of STMN1 expression rescues
the effect of miR-101 on cell viability, radiosensitivity and
radiation-induced autophagy
To determine the functional relevance of STMN1
regulation by miR-101, the effect of STMN1 on miR-101-mediated
growth suppression and radiosensitivity was evaluated. STMN1
expression was markedly increased following transfection with
PCA-STMN1 plasmid (Fig. 4A).
Furthermore, the co-expression of STMN1 markedly rescued the growth
suppression and radiosensitivity of CNE-2 cells transfected with
the miR-101 mimic (Fig. 4B and C).
Additionally, forced expression of STMN1 rescued the downregulation
of autophagy-associated protein LC3II and upregulation of p62
protein (Fig. 4D), and decreased
the lipidation of LC3 (Fig. 4E),
which represented decreased activity of autophagy in the presence
of miR-101 24 h following irradiation. These results indicated that
STMN1 is one of the key functional targets of miR-101, with respect
to the effect of miR-101 on the growth inhibition, radiosensitivity
enhancement and autophagy inhibition of NPC cells.
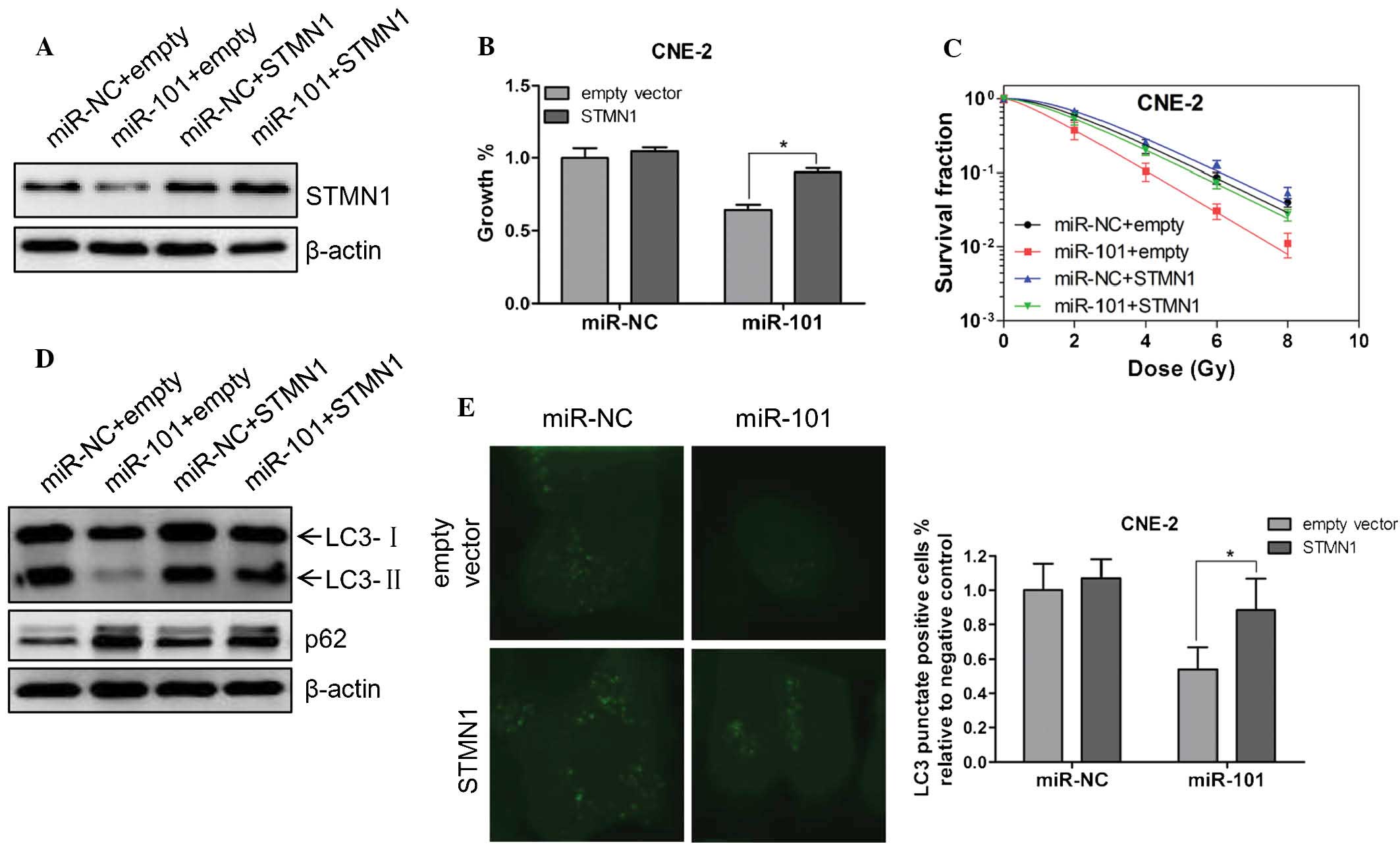 | Figure 4Restoration of STMN1 expression
rescues the effect of miR-101 on cell viability, radiosensitivity
and radiation-induced autophagy. (A) Either miR-101 mimic or
miRNA-NC was co-transfected into CNE-2 cells with PCA-STMN1 or the
empty vector. Western blot analysis was performed to verify the
expression of STMN1 48 h following co-transfection. β-Actin was
used as an internal control. (B) CNE-2 cells were treated as in A
and cell viability was determined with an MTT assay 72 h following
co-transfection. (C) Clonogenic survival assays of CNE-2 cells
treated as in A followed by various doses of radiation. Surviving
fractions were calculated as described. SER10 for miR-101+empty
vector, miR-NC+STMN1 and miR-101+STMN1 were 1.42, 0.91 and 1.02,
respectively. (D) CNE-2 cells were treated as in A followed by 6 Gy
irradiation. Twenty-four hours following IR, western blot analysis
was performed to detect the expression of LC3 and p62. β-Actin was
used as an internal control. (E) A CNE-2 cell line stably
expressing GFP-LC3 was established. These cells were treated as in
A followed by 6 Gy IR. Twenty-four hours following IR, cells were
fixed in 4% paraformaldehyde and subsequently observed under a
confocal microscope (magnification, ×600). The percentage of cells
with GFP-LC3 in puncta was calculated in five random fields. Values
are presented as the mean ± standard deviation.
*P<0.05 vs. empty vector. STMN1, stathmin 1; miR,
miRNA; NC, negative control; IR, irradiation. |
Discussion
The results of the present study demonstrated that
miR-101 was downregulated in NPC cell lines and that IR induced the
expression of miR-101. The ectopic expression of miR-101 suppressed
the viability of, and enhanced the radiosensitivity of NPC cells.
STMN1 was additionally identified as a direct functional target of
miR-101 involved in cell growth, radiosensitivity and
radiation-induced autophagy. These results suggested that miR-101
has significant roles in the development and radiosensitivity of
NPC.
Previous studies have indicated that numerous miRNAs
may function as oncogenes, for example miR-155 and miR-21 (22,23),
or as tumor suppressors, including the miR-200 family and let-7
(24,25). A loss of miR-101 expression is
frequently observed in certain types of human cancer and is
associated with therapeutic resistance, which suggests that miR-101
may act as a tumor suppressor (18). In the present study, it was
demonstrated that miR-101 was downregulated in NPC cell lines and
that the ectopic expression of miR-101 significantly suppressed
NPC-cell viability and enhanced their radiosensitivity. This
further supported the hypothesized anti-tumor effects of miR-101 in
NPC.
A study by Frankel et al (7) indicated that miR-101 may act as an
inhibitor of autophagy, a catabolic pathway which involves
self-degradation and the recycling of macromolecules and cellular
organelles. This mechanism has been shown to be a critical adaptive
response for tumor cell-survival under stressful conditions,
including hypoxia, chemotherapy, radiotherapy or nutrient
deprivation, which results in therapeutic resistance (26). Studies have also suggested that
blocking autophagy may enhance NPC radiosensitivity (27). In the present study, it was
demonstrated that expression of miR-101 was increased in response
to IR. This behavior may be an adaptive feedback response to
regulate autophagy.
miRNAs mainly regulate their target gene expression
via translational repression, mRNA degradation or both (3). Several targets of miR-101, such as
histone-lysine N-methyltransferase (28) and ATM (29), have been identified. In the present
study, a luciferase reporter gene assay verified STMN1 as a direct
target of miR-101. In addition, the overexpression of miR-101
significantly reduced the mRNA and protein expression of STMN1. A
recent study demonstrated that the expression of STMN1 was
upregulated in malignant cancers and correlated with poor prognosis
and therapeutic resistance (30).
In the present study, it was demonstrated that silencing STMN1
expression was able to suppress NPC-cell proliferation and enhance
their radiosensitivity. Furthermore, the forced expression of STMN1
rescued the growth suppression, radiosensitivity and decreased
activity of radiation-induced autophagy in NPC cells. This
suggested that miR-101 may suppress cell proliferation and enhance
radioresistance of NPC cells by directly targeting STMN1. Further
investigation is required to elucidate the association between
miR-101-mediated autophagy and the radiosensitivity of NPC.
In conclusion, the results of the present study
indicated that the downregulation of miR-101 in NPC cell lines and
ectopic expression of miR-101 suppressed the cell viability and
enhanced the radiosensitivity of NPC cells by directly targeting
STMN1. This identified an miR-101/STMN1 pathway which may
contribute to the elucidation of the molecular mechanisms by which
miR-101 regulates the radiosensitivity of NPC cells. Further
investigation will be performed to determine whether
miR-101-enhanced radiosensitivity is correlated with the inhibition
of autophagy in NPC.
Acknowledgements
The authors would like to thank Professor Xia Yunfei
for providing the nasopharyngeal carcinoma cell lines. This study
was supported by the National Natural Science Foundation of China
Grant (nos. 81272508, 81172243 and 81201961) and the Guangdong
Natural Science Foundation (no. S2011040003465).
References
|
1
|
Wei WI and Sham JS: Nasopharyngeal
carcinoma. Lancet. 365:2041–2054. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lo KW, Chung GT and To KF: Deciphering the
molecular genetic basis of NPC through molecular, cytogenetic, and
epigenetic approaches. Semin Cancer Biol. 22:79–86. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Feng XP, Yi H, Li MY, Li XH, Yi B, Zhang
PF, Li C, Peng F, Tang CE, Li JL, et al: Identification of
biomarkers for predicting nasopharyngeal carcinoma response to
radiotherapy by proteomics. Cancer Res. 70:3450–3462. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lai SZ, Li WF, Chen L, Luo W, Chen YY, Liu
LZ, Sun Y, Lin AH, Liu MZ and Ma J: How does intensity-modulated
radiotherapy versus conventional two-dimensional radiotherapy
influence the treatment results in nasopharyngeal carcinoma
patients? Int J Radiat Oncol Biol Phys. 80:661–668. 2011.
View Article : Google Scholar
|
|
5
|
Gupta AK, McKenna WG, Weber CN, Feldman
MD, Goldsmith JD, Mick R, Machtay M, Rosenthal DI, Bakanauskas VJ,
Cerniglia GJ, et al: Local recurrence in head and neck cancer:
relationship to radiation resistance and signal transduction. Clin
Cancer Res. 8:885–892. 2002.PubMed/NCBI
|
|
6
|
Kim VN, Han J and Siomi MC: Biogenesis of
small RNAs in animals. Nat Rev Mol Cell Biol. 10:126–139. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Frankel LB, Wen J, Lees M, Høyer-Hansen M,
Farkas T, Krogh A, Jäättelä M and Lund AH: microRNA-101 is a potent
inhibitor of autophagy. EMBO J. 30:4628–4641. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bartel DP: microRNAs: genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Esquela-Kerscher A and Slack FJ: Oncomirs
- microRNAs with a role in cancer. Nat Rev Cancer. 6:259–269. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
O’Day E and Lal A: MicroRNAs and their
target gene networks in breast cancer. Breast Cancer Res.
12:2012010. View
Article : Google Scholar
|
|
11
|
Zheng J, Deng J, Xiao M, Yang L, Zhang L,
You Y, Hu M, Li N, Wu H, Li W, et al: A sequence polymorphism in
miR-608 predicts recurrence after radiotherapy for nasopharyngeal
carcinoma. Cancer Res. 73:5151–5162. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang JX, Qian D, Wang FW, Liao DZ, Wei
JH, Tong ZT, Fu J, Huang XX, Liao YJ, Deng HX, et al: microRNA-29c
enhances the sensitivities of human nasopharyngeal carcinoma to
cisplatin-based chemotherapy and radiotherapy. Cancer Lett.
329:91–98. 2013. View Article : Google Scholar
|
|
13
|
He XP, Shao Y, Li XL, Xu W, Chen GS, Sun
HH, Xu HC, Xu X, Tang D, Zheng XF, et al: Downregulation of miR-101
in gastric cancer correlates with cyclooxygenase-2 overexpression
and tumor growth. FEBS J. 279:4201–4212. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Luo L, Zhang T, Liu H, Lv T, Yuan D, Yao
Y, Lv Y and Song Y: miR-101 and Mcl-1 in non-small-cell lung
cancer: expression profile and clinical significance. Med Oncol.
29:1681–1686. 2012. View Article : Google Scholar
|
|
15
|
Schee K, Boye K, Abrahamsen TW, Fodstad Ø
and Flatmark K: Clinical relevance of microRNA miR-21, miR-31,
miR-92a, miR-101, miR-106a and miR-145 in colorectal cancer. BMC
Cancer. 12:5052012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang R, Wang HB, Hao CJ, Cui Y, Han XC, Hu
Y, Li FF, Xia HF and Ma X: miR-101 is involved in human breast
carcinogenesis by targeting Stathmin1. PLoS One. 7:e461732012.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang J, Han C, Zhu H, Song K and Wu T:
miR-101 inhibits cholangiocarcinoma angiogenesis through targeting
vascular endothelial growth factor (VEGF). Am J Pathol.
182:1629–1639. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen S, Wang H, Ng WL, Curran WJ and Wang
Y: Radiosensitizing effects of ectopic miR-101 on non-small-cell
lung cancer cells depend on the endogenous miR-101 level. Int J
Radiat Oncol Biol Phys. 81:1524–1529. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Green LM, Reade JL and Ware CF: Rapid
colorimetric assay for cell viability: application to the
quantitation of cytotoxic and growth inhibitory lymphokines. J
Immunol Methods. 70:257–268. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xie G, Zhan J, Tian Y, Liu Y, Chen Z, Ren
C, Sun Q, Lian J, Chen L, Ruan J, et al: Mammosphere cells from
high-passage MCF7 cell line show variable loss of tumorigenicity
and radioresistance. Cancer Lett. 316:53–61. 2012. View Article : Google Scholar
|
|
21
|
Mah LJ, El-Osta A and Karagiannis TC:
gammaH2AX: a sensitive molecular marker of DNA damage and repair.
Leukemia. 24:679–686. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kong W, He L, Richards EJ, Challa S, Xu
CX, Permuth-Wey J, Lancaster JM, Coppola D, Sellers TA, Djeu JY and
Cheng JQ: Upregulation of miRNA-155 promotes tumour angiogenesis by
targeting VHL and is associated with poor prognosis and
triple-negative breast cancer. Oncogene. 33:679–689. 2014.
View Article : Google Scholar :
|
|
23
|
Oue N, Anami K, Schetter AJ, Moehler M,
Okayama H, Khan MA, Bowman ED, Mueller A, Schad A, Shimomura M, et
al: High miR-21 expression from FFPE tissues is associated with
poor survival and response to adjuvant chemotherapy in colon
cancer. Int J Cancer. 134:1926–1934. 2014. View Article : Google Scholar
|
|
24
|
Schliekelman MJ, Gibbons DL, Faca VM,
Creighton CJ, Rizvi ZH, Zhang Q, Wong CH, Wang H, Ungewiss C, Ahn
YH, et al: Targets of the tumor suppressor miR-200 in regulation of
the epithelial-mesenchymal transition in cancer. Cancer Res.
71:7670–7682. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hu X, Guo J, Zheng L, Li C, Zheng TM,
Tanyi JL, Liang S, Benedetto C, Mitidieri M, Katsaros D, Zhao X,
Zhang Y, Huang Q and Zhang L: The heterochronic microRNA let-7
inhibits cell motility by regulating the genes in the actin
cytoskeleton pathway in breast cancer. Mol Cancer Res. 11:240–250.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Choi AM, Ryter SW and Levine B: Autophagy
in human health and disease. N Engl J Med. 368:651–662. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang Y, Yin W and Zhu X: Blocked autophagy
enhances radiosensitivity of nasopharyngeal carcinoma cell line
CNE-2 in vitro. Acta Otolaryngol. 134:105–110. 2014. View Article : Google Scholar
|
|
28
|
Luo C, Merz PR, Chen Y, Dickes E, Pscherer
A, Schadendorf D and Eichmüller SB: miR-101 inhibits melanoma cell
invasion and proliferation by targeting MITF and EZH2. Cancer Lett.
341:240–247. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yan D, Ng WL, Zhang X, Wang P, Zhang Z, Mo
YY, Mao H, Hao C, Olson JJ, Curran WJ and Wang Y: Targeting
DNA-PKcs and ATM with miR-101 sensitizes tumors to radiation. PLoS
One. 5:e113972010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wik E, Birkeland E, Trovik J, Werner HM,
Hoivik EA, Mjos S, Krakstad C, Kusonmano K, Mauland K, Stefansson
IM, et al: High phospho-Stathmin(Serine38) expression identifies
aggressive endometrial cancer and suggests an association with PI3K
inhibition. Clin Cancer Res. 19:2331–2341. 2013. View Article : Google Scholar : PubMed/NCBI
|