Introduction
Trihydroxyethylrutoside (troxerutin) is one of the
flavonoid rutoside derivatives. It exhibits non-mutagenic
properties and has a functional role in the treatment of chronic
venous insufficiency (CVI) (1,2). A
number of studies have demonstrated other beneficial effects of
troxerutin, in vitro and in vivo, and it may be
effective in reducing different cytotoxicities. In particular,
troxerutin has been observed to exhibit an inhibitory effect on the
neurotoxicity induced by high cholesterol mediated cognitive
deficits, kainic acid-triggered excitotoxic damage and β-amyloid
oligomerization (3–5). In addition, troxerutin has a
photoprotective effect against ultraviolet B (UVB) radiation in
human skin cells, including dermal fibroblasts and keratinocytes
(6,7). Troxerutin also exerts a protective
effect against γ-radiation in mice (8,9).
Although the precise cellular mechanisms underlying the effects of
troxerutin remain to be fully elucidated, these reports can be
summarized as a single meaningful finding, that troxerutin inhibits
the production of reactive oxygen species (ROS). In vivo
investigations have demonstrated that CVI-bearing patients have
increased levels of ROS, and troxerutin has a protective effect
against oxygen-derived free radical scavengers on the endothelium
in these patients (10,11). In addition, the aforementioned
neurotoxicities are inhibited following troxerutin application by
reducing the production of ROS (3,4,12).
UVB and γ-radiation are known ROS stimulators (13,14),
and a previous study demonstrated that troxerutin protects against
radiation-induced lipid peroxidation (9). These studies suggest that this
toxerutin may offer a novel therapeutic strategy for ROS-induced
diseases.
Dermal papilla (DP) cells are located at the base of
hair follicles and are important in the induction of growth and
maintenance of epithelial cells, which are the predominant
components of hair follicles (15). In response to hormonal changes, DP
cells direct the follicular epithelial cells to enter the hair
growth cycle, which involves anagen, an active growing phase;
catagen, a short transitionary regressive phase; and telogen, a
dormant resting phase (15). An
increasing body of evidence has demonstrated excessive loss of
viability and death of DP cells in balding regions of the scalp,
compared with non balding regions, due to increased levels of
5α-reductase (16), a converting
enzyme for androgenic hormones and intracellular ROS (17). In addition, previous reports have
indicated that oxidative stress is generated by the exposure of
androgen sensitive prostate cancer cells to high levels of
androgens (18), and that lipid
peroxides increase the levels of ROS and apoptosis of the hair
follicle cells (19). Furthermore,
DP cells in the balding scalp grow more slowly in vitro,
compared with cells from the non balding scalp. The reduced
proliferative activity of balding DP cells is associated with
changes in the expression levels of senescence-associated (SA)
β-galactosidase, oxidative stress markers, superoxide dismutase and
catalase (20). These findings
indicate that oxidative stress is important in the loss of DP cells
and in hair production.
In the present study, the hypothesis that troxerutin
inhibits ROS-mediated cellular dysfunction in human DP (HDP) cells
was investigated. In addition, using micro (mi)RNA microarrays and
bioinformatics analysis, the role of troxerutin in the regulation
of the expression and mechanisms of specific miRNAs was evaluated.
The present study aimed to examine troxerutin as a potential novel
chemical agent for the preven tion and/or treatment of
alopecia.
Materials and methods
Cell culture and viability
The HDP cells were purchased from Innoprot (Biscay,
Spain) and cultured in Dulbecco’s modified Eagle’s medium,
containing 10% fetal bovine serum (FBS; Thermo Fisher Scientific,
Waltham, MA, USA) and 1% penicillin streptomycin (Gibco Life
Technologies, Grand Island, NY, USA) at 37°C and 5% CO2.
The cells were plated at a density of 4×103/well in a
96-well plate. At 70–80% confluence, the cells were treated with
troxerutin (Sigma-Aldrich, St. Louis, MO, USA) at concentrations
ranging between 0 and 60 μM for 24 h at 37°C. Subsequently,
10 μl water soluble tetrazolium salt assay solution
(EZ-Cytox Cell Viability Assay kit; Itsbio, Seoul, Korea) was added
to each well and, following incubation for 30 min at 37°C, the
optical density was measured at 490 nm using an iMark microplate
reader (Bio Rad Laboratories, Inc., Hercules, CA, USA). To examine
troxerutin mediated ROS protection, the cells were pretreated with
troxerutin at the following concentrations: 0, 5, 10 and 15
μM for 8 h. Subsequently, 750 μM
H2O2 was added to each well. Following
incubation for 24 h at 37°C, cell viability was evaluated using an
EZ-Cytox Cell Viability Assay kit. The level of cell viability (%)
was normalized to that of 0.1% dimethyl-sulfoxide (DMSO;
Sigma-Aldrich)-treated cells. Each experiment was repeated at least
three times. The P-value was determined using Student’s t-test and
P<0.05 was considered to indicate a statistically significant
difference.
Analysis of cell cycle
The HDP cells (2×106), which had been
treated with troxerutin and/or H2O2 were
trypsinized using 0.25% trypsin-EDTA (Gibco Life Technologies),
washed once with phosphate-buffered saline (PBS), and used for
analysis. The cell cycle distribution was measured using propidium
iodide (PI; Sigma-Aldrich) staining solution, containing 50
μg/ml PI, 0.5% Triton X-100 (Sigma-Aldrich) and 100
μg/ml RNase (Qiagen, Hilden, Germany). The cells were fixed
in 70% cold ethanol (Merck Millipore, Darmstadt, Germany) and
incubated for 1 h at −20°C. Subsequently, PI staining solution was
added to the fixed cells, followed by incubation for 1 h in the
dark at 37°C. The PI fluorescence intensity was detected using a BD
FACSCalibur flow cytometer (BD Biosciences, San Jose, CA, USA). The
mean PI fluorescence intensity was calculated based on the
measurements of 10,000 cells using the FL2-H channel.
Analysis of intracellular levels of
ROS
The HDP cells (2×106), which had been
treated with troxerutin and/or H2O2 were
washed with PBS and trypsinized. Intracellular ROS levels were
measured using 2′7′-dichlorofluorescein diacetate fluorescent dye
(DCF-DA; Sigma-Aldrich), as described previously (21). The cells were resuspended in 10
μM DCF-DA and further incubated at room temperature for 1 h
in the dark. The intensity of the resulting fluorescence was
measured using a BD FACSCalibur flow cytometer (BD Biosciences).
The mean DCF fluorescence intensity was calculated based on
measurements of 10,000 cells using the FL1-H channel. The M1 range
was calculated as the percentage of each subpopulation of cells
exhibiting increased DCF-DA fluorescence.
Analysis of cellular senescence
The HDP cells (2×106), which had been
treated with troxerutin and/or H2O2 were
washed in PBS and fixed for 5 min at room temperature in 2%
formaldehyde/0.2% glutaraldehyde (Sigma-Aldrich). The cells were
washed with PBS and incubated for 15 h at 37°C with SA
β-galactosidase staining solution (BioVision, Milpitas, CA, USA).
The stained cells (blue) were observed using a bright-field
microscope (CKX41; Olympus Corporation, Tokyo, Japan;
magnification, ×200), counted in three different fields and the
percentage of stained cells was determined.
Analysis of miRNA expression
profiles
The HDP, which had been cells treated with
troxerutin and/or H2O2 were washed in cold
PBS and trypsinized for RNA purification. The total RNA was
extracted and purified from the cells using TRIzol®
reagent (Invitrogen Life Technologies, Carlsbad, CA, USA),
according to the manufacturer’s instructions. The integrity (RNA
integrity number >8.0) and purity (A260/280 and A260/230 values
>1.8) were confirmed using an Agilent 2100
Bioanalyzer® (Agilent Technologies, Inc., Santa Clara,
CA, USA) and a MaestroNano® microvolume
spectrophotometer (Maestrogen, Las Vegas, NV, USA), respectively.
Samples (100 ng) of RNA meeting these criteria were first
dephosphorylated by incubation with calf intestinal alkaline
phosphatase (Agilent Technologies, Inc.) at 37°C for 30 min.
Subsequently, cyanine 3-pCp labeling solution (Agilent
Technologies, Inc.) and T4 RNA ligase (Agilent Technologies, Inc.)
were added to the dephosphorylated RNA samples and incubated at
16°C for 2 h. Following the labeling reaction, the samples were
dried and treated with GE Blocking Agent (Agilent Technologies,
Inc.). The samples were hybridized to the SurePrint G3 Human v16
miRNA 8×60 K (Agilent Technologies, Inc.) microarray at 55°C, with
constant rotation at 20 rpm in an Agilent Microarray Hybridization
Chamber(Agilent Technologies, Inc.) for 20 h. The array was then
washed and scanned using an Agilent SureScan Microarray scanner and
the images captured were quantified using Agilent Feature
Extraction software (version 10.7; Agilent Technologies, Inc.). The
data were analyzed with the assistance of GeneSpring GX software
version 7.3 (Agilent Technologies). In addition, fold-change
analysis was performed to select those with ≥2.0-fold between the
H2O2-treated control cells and those treated
with troxerutin and H2O2.
Bioinformatic analysis of altered
miRNAs
Analysis of the biological significance of the
altered miRNAs in the present study was performed, as previously
described (21). First, the
putative target genes of the altered miRNAs were predicted using
MicroCosm Targets Version 5 (http://www.ebi.ac.uk/enright-srv/microcosm/htdocs/targets/v5/).
Subsequently, the target genes were grouped into four categories:
Aging, skin development, apoptosis and cell proliferation, based on
the AmiGo 2 Gene Ontology (GO) analysis tool (amigo.geneontology.org/cgi-bin/amigo/browse.cgi). The
putative target genes of each miRNA were further analyzed for
biologic function using the Kyoto Encyclopedia of Genes and Genomes
(KEGG) pathway within the Database for Annotation, Visualization
and Integrated Discovery, (DAVID; http://david.abcc.ncifcrf.gov/home.jsp) bioinformatics
resources (version 6.7), according to the standard procedures
(22). The ‘KEGG_pathway’ category
was processed by setting the threshold of the EASE score, a
modified Fisher’s exact P-value, to 0.1. The KEGG pathways,
identified as having a percentage of involved target genes / total
target genes in each pathway >1% were selected.
Results
H2O2-induced cell
damage is inhibited by troxerutin in HDP cells
HDP cells are a major component of skin and direct
hair growth, and loss or senescence of these cells is a key cause
of hair loss (15). Previous
reports have demonstrated that cisplatin-and androgen
overexposure-mediated cellular dysfunction, including apoptosis,
can induce alopecia and occur predominantly by stimulating the
production ROS in HDP cells (16,23,24).
In our previous study, pretreatment of human dermal fibroblasts and
HaCaT keratinocytes with troxerutin was observed to protect against
UVB-mediated cell death (6,7). UVB
light is an important inducer of ROS production in several types of
cell (13); therefore, the present
study examined the possible role of troxerutin in protecting
against ROS-induced cell stress and damage in HDP cells. Initially,
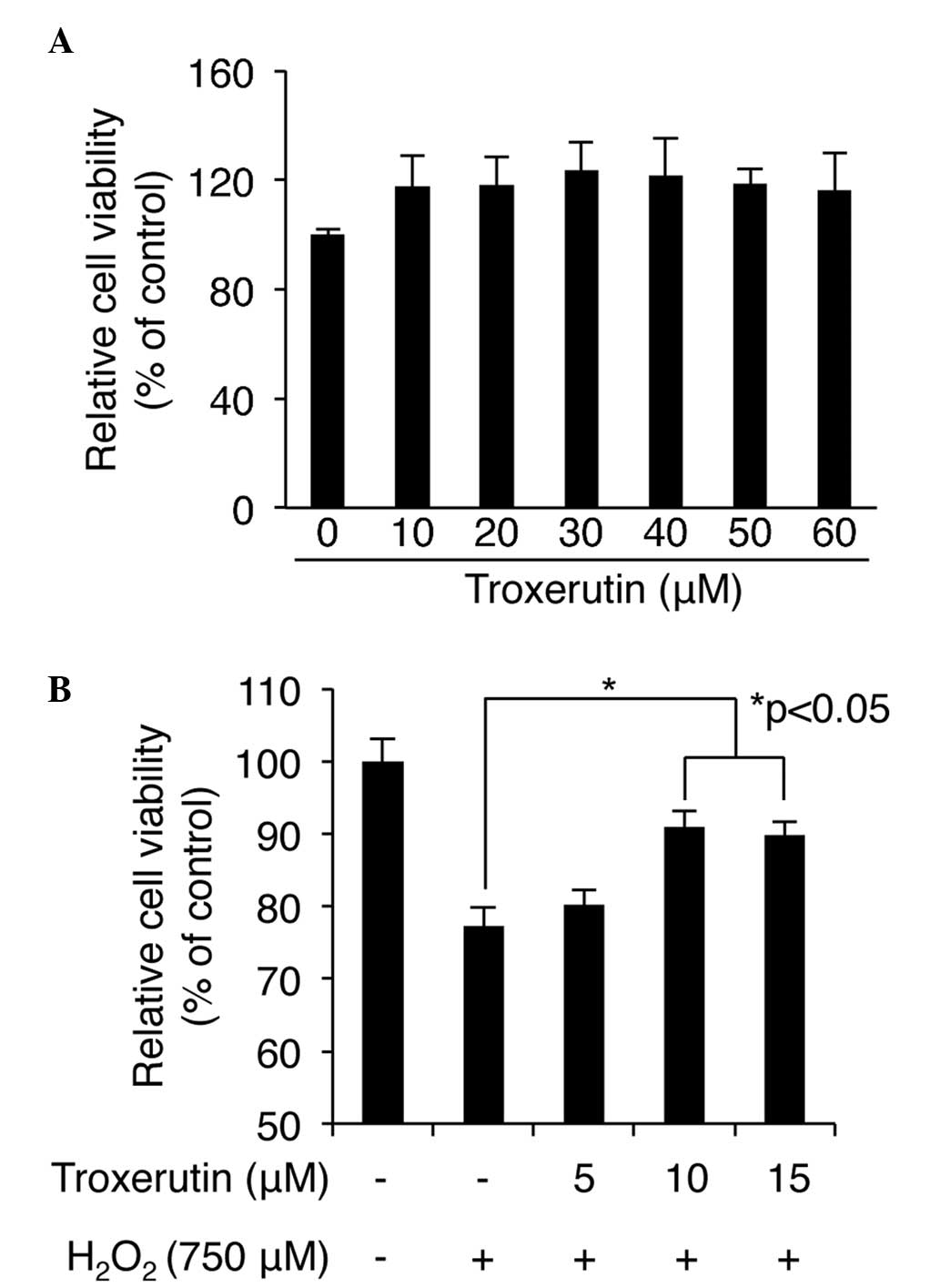
troxerutin-mediated cytotoxicity in HDP cells was screened for. No
significant changes in cell viability were detected following
treatment with between 0 and 60 μM troxerutin for 24 h
(Fig. 1A), indicating that
troxerutin was non toxic in the HDP cells. Subsequently, the
protective role of trox erutin against ROS-induced cell damage was
determined using H2O2, a ROS inducer. The HDP
cells were pretreated with several concentrations (0–60 μM)
of troxerutin for 6 h, followed by the addition of 750 μM
H2O2 and incubation for an additional 24 h.
The results revealed that the maximum protective effect against ROS
induced cell damage in the HDP cells occured folowing pretreatment
with 10 μM troxerutin (Fig.
1B). Treatment with H2O2 alone decreased
cell viability to 77.33±2.44%; however, pretreatment with 10
μM troxerutin maintained cell viability at 90.88±2.24%
following H2O2 exposure (P<0.05; Fig. 1B). Concentrations of troxerutin
>15 μM did not significantly enhance the protective
effect of 10 μM troxerutin (Fig. 1B and data not shown). These results
suggested that pretreatment with troxerutin inducedresistance
against H2O2-mediated cytotoxicity in the HDP
cells.
H2O2-induced cell
death is inhibited by troxerutin in HDP cells
Our previous study demonstrated that troxerutin
effects the level of cell death induced by UVB irradiation
(6,7) and Fig.
1 shows the protective role of troxerutin against ROS-induced
cell damage, observed in the present study. Therefore, to examine
whether troxerutin is involved in the response to ROS stress, which
is known to inhibit cell-cycle progression and induce cell death,
the present study investigated changes in the cell cycle and in
cell death in the HDP cells pretreated with different
concentrations of troxerutin prior to H2O2
exposure. The HDP cells were pretreated with 0, 5 or 10 μM
troxerutin for 6 h, followed by treat ment of the cells with 750
μM H2O2 for an additional 24 h.
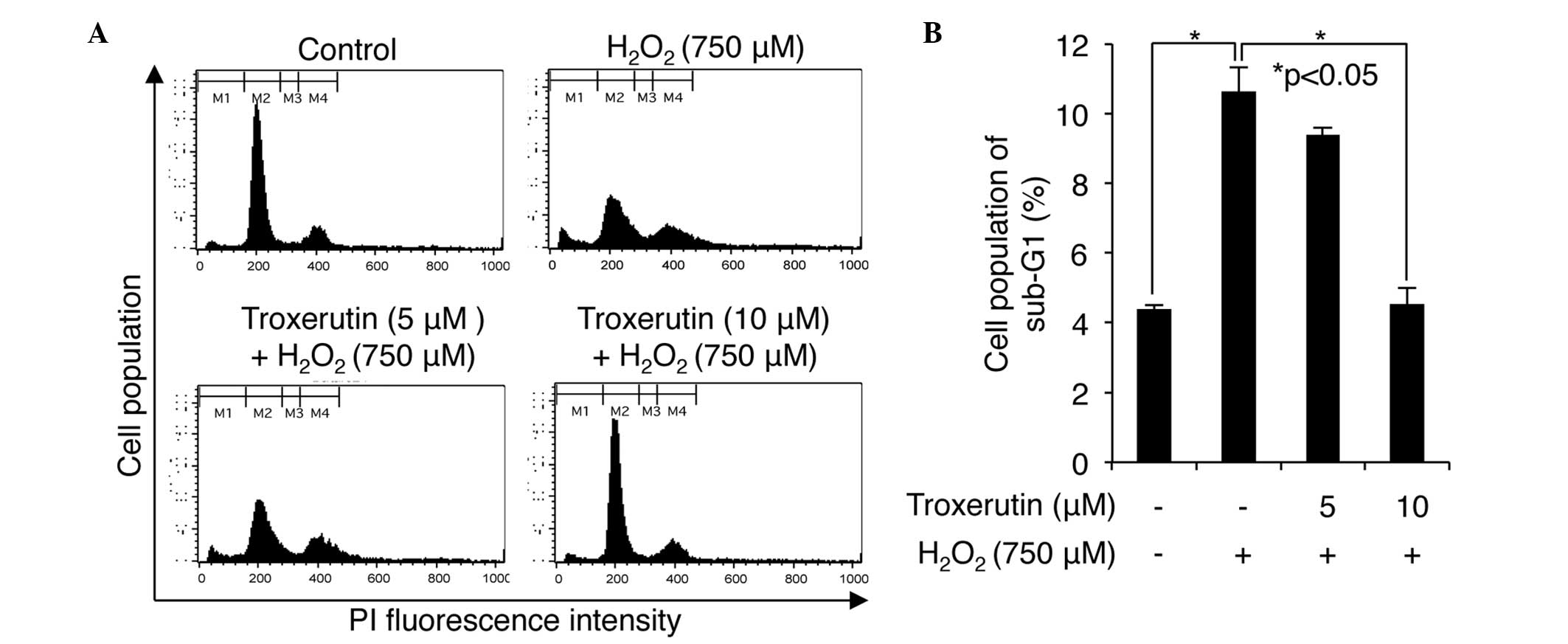
Subsequently, the cells were analyzed by flow cytometry. At
concentrations of 5 and 10 μM, pretreatment with troxerutin
caused a decrease in the number of cells in the sub G1 phase,
indicative of cell death (Fig.
2A). H2O2 increased the percentage of the
non-pretreated cells in the sub-G1 phase to 10.63±0.43%; however,
this value increased to only 9.38±0.11 and 4.53±0.53% in the cells
pretreated with 5 and 10 μM troxerutin, respectively
(P<0.05; Fig. 2A and B).
Therefore, these results suggested that troxerutin overcame the
effect of H2O2-mediated cell death, resulting
in a diminution of cells in the sub-G1 phase.
Hydrogen peroxide-induced ROS production
is inhibited by troxerutin in HDP cells
Although the above results indicated that troxerutin
had a protective effect against H2O2-mediated
cell death, it remained to be elucidated whether troxerutin also
regulates the level of intracellular
H2O2-induced ROS in HDP cells. To examine
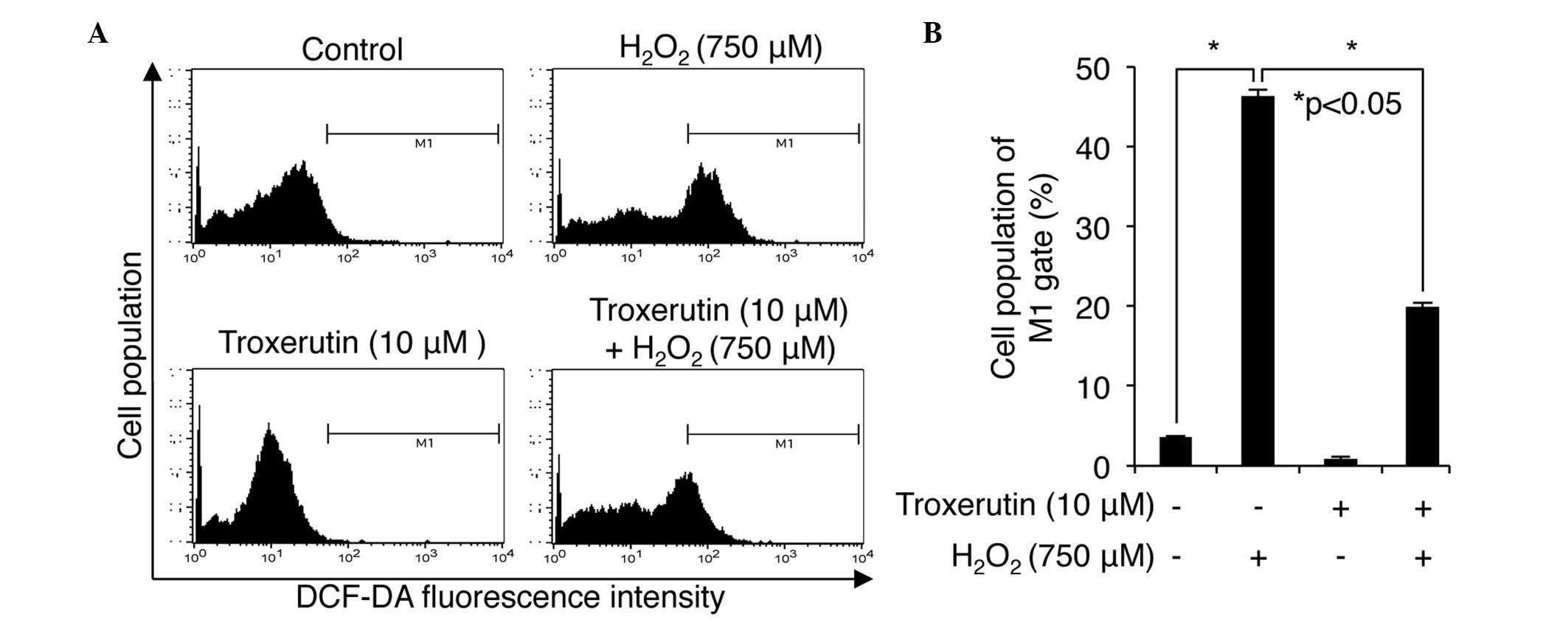
this, the presents study performed a fluorescent DCF-DA staining
assay following treatment with troxerutin and
H2O2 in the HDP cells. The DCF-positive cells
were then analyzed using flow cytometry to determine and compare
the levels of intracellular ROS in the control cells,
non-troxerutin pretreated cells, troxeruti only treated cells and
troxerutin pretreated/H2O2 treated cells. As
shown in Fig. 3, in the control
and troxerutin-only-treated cells, 3.58±0.15 and 0.89±0.11% were
DCF-positive (P<0.05; Fig. 3B),
suggestive of ROS respectively, whereas treatment with
H2O2 alone increased the level of ROS to
46.36±2.33%. The cells pretreated with troxerutin were 19.92±1.95%
DCF-positive following H2O2 treatment,
indicating that troxerutin reduced the
H2O2-induced production of ROS in the HDP
cells.
Hydrogen peroxide-induced senescence is
inhibited by troxerutin in HDP cells
Increased ROS are one of the key mediators of
cellular senescence (25). A
previous report demonstrated that the premature senescence of
balding DP cells is associated with changes in the expression of SA
β-galactosidase and also suggested that oxidative stress may be
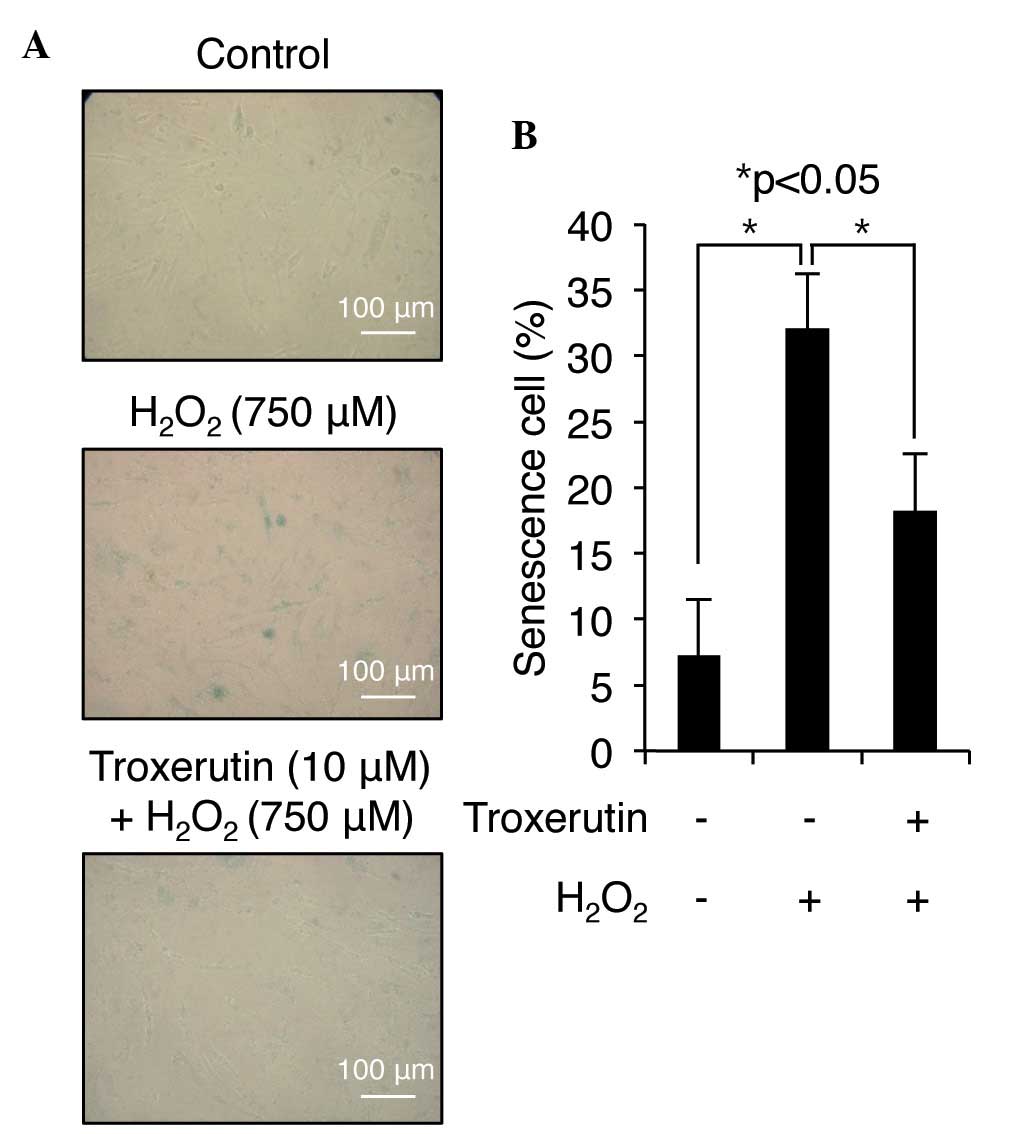
involved in the premature senescence of these cells (20). The present study assayed for the
presence of SA β-galactosidase activity to investigate whether
troxerutin affects H2O2-induced senescence,
thereby contributing to its protective effect against ROS-mediated
cell damage. H2O2 treatment increased the
number of SA β-galactosidase-positive cells to 32.11±3.32% compared
with the control; however, only 18.22±5.21% of the cells pretreated
with troxerutin were SA β-galactosidase-positive following
treatment with H2O2 (Fig. 4). These data indicated that
troxerutin has the potential to inhibit cellular senescence in HDP
cells.
Troxerutin-mediated protective effects
against ROS are involved in changes in miRNA expression in HDP
cells
The present study also analyzed the miRNA expression
profiles of non-pretreated and troxerutin-pretreated HDP cells
treated with H2O2, as miRNAs can be involved
in cell death, ROS scavenging and senescence (21,26–28).
In the microarray, a total of 24 miRNAs were detected with a
≥2.0-fold change in expression levels between the two groups. Among
these, 10 miRNAs were upregulated and 14 were downregulated in the
H2O2-treated cell, which had been pretreated
with troxerutin (Table I). To
investigate the biological value of the microarray data, several
bioinformatic analyses were performed to predict the putative
target genes of the altered miRNAs, and the GO and signaling
pathways of the target genes. The putative target genes of each
miRNA were deterfmined using MicroCosm Targets Version 5, following
which GO analysis was performed for the target genes. Subsequently,
the target genes of each miRNA were categorized into four
biological functions: Aging, skin development, apoptosis and cell
proliferation. Several target genes of each miRNA were found to be
involved in these four biological functions at different levels
(Tables II and III). For example, has-miR-602, which
was the most highly upregulated miRNA (6.91-fold) based on the
microarray data, potentially targets 34 genes, six of which were
involved in aging, and one of the remaining 28 target genes was
involved in skin development (Table
II). Similarly, the target genes of the downregulated miRNAs
were also differentially involved in the four functions (Table III), indicating that the altered
miRNAs identified by the microarray analysis had distinct
biological roles associated with the protective effect of
troxerutin in H2O2-treated HDP cells.
Therefore, the present study further analyzed the signaling
pathways associated with the upregulated and downregulated miRNAs
using KEGG pathway analysis and the DAVID bioinformatics tool
(22), the results of which are
presented in Tables IV and
V, respectively. The results
demonstrated that the altered miRNAs are functionally involved in
shared and unique pathways among the miRNAs. For example,
hsa-miR-602 was identified to be functionally involved in MAPK,
insulin, and calcium signaling pathways, whereas has-miR-205 3p was
found to be involved in cancer, MAPK, Wnt and cell adhesion
signaling pathways. Overall, these results indicated that the miRNA
expression patterns of non-pretreated and troxerutin-pretreated
H2O2-treated HDP cells can be distinguished,
and those which are significant changed may be involved in
troxerutin-mediated protection against
H2O2-induced cellular stress through the
regulation of multiple signaling pathways.
 | Table IMicroRNAs with ≥2-fold change in
expression in troxerutin pretreated
H2O2-treated human dermal papilla cells. |
Table I
MicroRNAs with ≥2-fold change in
expression in troxerutin pretreated
H2O2-treated human dermal papilla cells.
| microRNA | Change relative to
control | Direction of
regulation | Chromosome |
|---|
| hsa-miR-150-3p | 4.13 | Up | 19 |
|
hsa-miR-181a-2-3p | 2.31 | Up | 9 |
| hsa-miR-205-3p | 4.28 | Up | 1 |
| hsa-miR-21-3p | 2.92 | Up | 17 |
|
hsa-miR-29b-1-5p | 3.72 | Up | 7 |
|
hsa-miR-3127-5p | 2.19 | Up | 2 |
|
hsa-miR-371a-5p | 2.30 | Up | 19 |
|
hsa-miR-3663-3p | 2.53 | Up | 10 |
| hsa-miR-4298 | 2.01 | Up | 11 |
| hsa-miR-602 | 6.91 | Up | 9 |
| hsa-miR-1181 | −3.14 | Down | 19 |
| hsa-miR-1202 | 2.78 | Down | 6 |
|
hsa-miR-1224-5p | −4.66 | Down | 3 |
| hsa-miR-1290 | 2.15 | Down | 1 |
|
hsa-miR-135a-3p | 5.61 | Down | 3 |
| hsa-miR-28-5p | 2.95 | Down | 3 |
|
hsa-miR-378a-5p | 2.01 | Down | 5 |
| hsa-miR-4271 | −2.16 | Down | 3 |
| hsa-miR-452-5p | −2.51 | Down | X |
| hsa-miR-572 | −4.26 | Down | 4 |
| hsa-miR-575 | −8.01 | Down | 4 |
| hsa-miR-629-3p | 3.12 | Down | 15 |
| hsa-miR-939 | 2.22 | Down | 8 |
| hsa-miR-940 | −2.30 | Down | 16 |
| Table IIPredicted targets of microRNAs
upregulated in response to troxerutin pretreatment in
H2O2-exposed human dermal papilla cells. |
Table II
Predicted targets of microRNAs
upregulated in response to troxerutin pretreatment in
H2O2-exposed human dermal papilla cells.
| microRNA | Aging | Skin
development | Apoptosis | Cell
proliferation |
|---|
| hsa-miR-150-3p | – | – | BCL3, INHBA,
ARHGEF2, RHOA, ATG7, MAP3K5, MECOM, PLAGL1 | BCL3, INHBA, MECOM,
ARHGEF2, RHOA, NDN, PROX1, BTRC, TGFBI |
|
hsa-miR-181a-2-3p | LMNA, SRF | SRF | LMNA, MED1, BDNF,
TIAL1, SRPK2, CITED2, AGAP2, PSMD7, MAPK8 | SRF, MED1, BDNF,
TIAL1, SRPK2, CITED2, SOX11, FBXW7 |
| hsa-miR-205-3p | TBX3, CDK6, MNT,
IL1B, TECP2L1, PNPT1, ATR | APC, DBI | BRCA1, HDAC2, SOS2,
SIX4, GRAM, MDM4, CUL5, NBN, MNT, IL1B, TBX3, WNT5A, RAD21, MAP3K5,
RASSF6, CREB1, GLO1, API5, SOS1, APC, MSX2, FGF2, SOX2, DUSP1,
GSK3B, PSMA5,MITF, HiPK2, HOXA13, PARK7, NAIP, BCLAF1 | MITF, GRAM, MDM4,
CUL5, NBN, CDK13, CASK, PURA, MNT, IL1B, TBX3, HDAC2, CDK6, MSX2,
FGF2, SOX2, HiPK2, BRCA1, WNT5A, EVI5, TOB1, NUMB |
| hsa-miR-21-3p | CDK6 | – | MAP2K4, MAP3K1,
BCL2L11, SMAD3, CUL3, SOX4, BAG4, RNF41, AMIGO2, SLC11A2, KDM28,
DAB2IP, FOXO3, CCAR1, ROBO2, TRIM32, DSG1 | CUL3, SOX4, NR6A1,
FTO, TRIM32, FOXO3, SMAD3, CDK6, KDM28, DAB2IP, CD274, PBRM1 |
|
hsa-miR-29b-1-5p | NR3C1, SIRT1 | – | NR3C1, SIRT1, REST,
PTK2, SOS2, NUAK2, PSMD7 | NR3C1, SIRT1, REST,
PTK2, FGF18, INSR, PBRM1 |
| hsa-miR-3127 | – | – | – | – |
|
hsa-miR-371a-5p | – | LEF1, ATP7A,
COL8A1 | LEF1, SOX2, CITED2,
STK4, RB1CC1, BARD1, GSK3B, PSMF1, NR4A2, DYRK2, RPS6KA1, ITSN1,
MAP3K1 | LEF1, SOX2, CITED2,
STK4, COL8A1, RNF10, MAPRE1, BTG3, CCR2, FRS2, PRMT5 |
|
hsa-miR-3663-3p | FAS, CASP2, CDKN1A,
PTH1R | ADAMTS2, BCL11B,
COL3A1, COL1A1 | FAS, CASP2, BCL11B,
USP28, TGFB2, DDX5, COMP, PIGT, CDKN1A, TIAL1, PPP2R1B, PSMA2,
MEF2D | FAS, TIAL1, TGFB2,
USP28, CDKN1A, BCL11B, VSIG |
| hsa-miR-4298 | HMGA1, AMFR | – | MED1, FGF2, TRAF5,
CCAR1 | HMGA1, MED1, WT1,
FGF2 |
| hsa-miR-602 | EDN1, VDR, SOD2,
HTT, SLC34A2, CHEK1 | APC | NOG, ERBB4, PSMD2,
PIM1, EDN1, VDR, SOD2, DYRK2, ALDH1A2, CLI2, SEMA3A, HTT, APC,
H1F0, PPARG, BCL2L15, JMY, TP53BP2, MYO18A, SHF | NOG, ERBB4, PIM1,
PPARG, CLI2, CDC27, CDK13, LIFR, EDN1, VDR, SOD2, STAT3, APC,
ALDH1A2, ACSL6, PPP1R8, EMX2, CDK9, RTKN2, ID4, ZEB1 |
| Table IIIPredicted targets of microRNAs
downregulated in response to troxerutin pretreatment in
H2O2-exposed human dermal papilla cells. |
Table III
Predicted targets of microRNAs
downregulated in response to troxerutin pretreatment in
H2O2-exposed human dermal papilla cells.
| microRNA | Aging | Skin
development | Apoptosis | Cell
proliferation |
|---|
| hsa-miR-1181 | – | – | – | – |
| hsa-miR-1202 | CLNB, PNPT1,
SLC18A2 | – | CLN8, RRN3, PIK3CG,
ETS1, DRAM1, DNAJC10, STEAP3, IKBKG, SOS1, NOD1 | RRN3, PIK3CG, ETS1,
CDC6, BCAT1, NRP1, ERG, SESN1, FZD6, CD276, GAS8, RPS15A |
|
hsa-miR-1224-5p | HMGA2, AQP2,
SLC1A2 | APC | HMGA2, AQP2, APC,
FGFR1, ADORA1, SATB1, STAT5B | RBBP7, APC, CD160,
RC3H1, HMGA2, FGFR1, ADORA1, SATB1, STAT5B, NOLC1 |
| hsa-miR-1290 | HMGA2, NUAK1,
TERF2, SLC1A2, FADS1, DDC | APC, COL8A1 | HMGA2, APC, RRN3,
ITGAV, CSE1L, NOTCH1, GAS, BMI1, FOXC1, ROBO1, USP28 | HMGA2, BMI1, NUAK1,
APC, MLL2, RRN3, ITGAV, CSE1L, NOTCH1, GAS, HES1, NPR3, CDC27,
COL8A1, CDKN2B, FOXC1, ROBO1, USP28, FIGF, NRAS |
|
hsa-miR-135a-3p | – | TFAP2A | TFAP2A, POU3F3,
RRP8, PEG3, DYRK2, | TFAP2A, POU3F3,
DERL2, RERG, COL8A1, CEP120 |
| hsa-miR-28-5p | – | – | MST4 CNTFR, STK4,
BAG1, SON, NR4A3, PAK2 | CNTFR, STK4, HTR4,
FTSJ2, SESN1, TNS3, RAP18, DERL2 |
|
hsa-miR-378a-5p | PML | – | DFFA, ITSN1, CTSB,
ROBO2, DEPTOR, RAG1, RFFL, IL24, PML, VHL, FRZB, STK4, BAG1,
ITGB2 | FZD3, RAC2, CCND2,
FER, PML, VHL, FRZB, STK4, NUDC, PDAP1, ITGAL, PELI1, HNF4A,
CD33 |
| hsa-miR-4271 | HMGA1, AMFR,
SLC6A3 | – | ALDH1A2, SPN,
EIF2AK3, FOXO3, WNT7B, MAPK1, CYLD, MAPT, MEF2D, DAPL1, EP300 | COL4A3BP, FOXO4,
PDGFB, WNT7B, MAPK1, ALDH1A2, CDK2, SPN, MXD1, FOXO3, TGFBR3,
CNOT8, MBD2, CD209, CDON, HOXD13, |
| hsa-miR-452-5p | TIMP3 | – | SPRY2, PAX3, SOX7,
LRP6, SNAI2, CSNK2A2, FGD4, PKN2, ITGA6, PDCD6IP | SPRY2, PAX3, SOX7,
LRP6, SNAI2, RPA1, EPS8, NFIB, MAPRE1, ODZ1, CDCA7L, CD47, E2F3,
PURA, |
| hsa-miR-572 | – | – | HIP1, CASP10, E2F2,
MAP3K1 | RUNX1, CDC27,
ROS1 |
| hsa-miR-575 | – | – | ZBTB16, HIP1,
PDPK1, BRAF, CASP10, E2F2, MAP3K1, DNM1L | ZBTB16, NR3C2,
NDEL1, ROS1, BRFOX2, KIF15 |
| hsa-miR-629-3p | SOD2, VDR, EDN1,
CHEK1, SLC34A2 | – | THOC1, MYO18A,
TP53BP2, APC, PPARG, PIM1, PSMD2, SOD2, VDR, EDN1, ERBB4, PERP,
BCL2L15 | DLG3, RTKN2, CDK9,
STAT3, EPHB1, ACSL6, LIFR, EREG, APC, PPARG, PIM1, STAT6, PDGFC,
ZEB1, NOLC1, ID4, SOD2, VDR, EDN1, ERBB4, CDK13, CDC27 |
| hsa-miR-939 | TIMP1, ATM, CDKN1A,
NEK6, SCL34A2, PRELP, SLC1A2 | NGFR, COL1A1 | TNF, BCL6, BTC,
NRG1, IHH, TIMP1, ATM, WNK3, CLIP3, NEK6, NGFR, MT3, TRAIP, CDKN1A,
NACC1, IP6K2, PAX7, CAMK1D, CASP10, USP7, CSNK2A2, THRA, INHBB,
BCL2L2 | BCL6, BTC, NRG1,
IHH, GRN, TRAIP, CDKN1A, TNF, E2F8, RXRB, RARA, DRD2, CSF1, TIMP1,
ATM, NGFR, MT3, NOS2, AGGF1, ELN |
| hsa-miR-940 | – | – | – | – |
| Table IVFunctional annotation chart for
miRNAs upregulated in response to troxerutin pretreatment in
H2O2-exposed human dermal papilla cells. |
Table IV
Functional annotation chart for
miRNAs upregulated in response to troxerutin pretreatment in
H2O2-exposed human dermal papilla cells.
| microRNA | Putative target
genes (n) | KEGG pathway | Genes involved in
the term (n) | Involved
genes/total genes (%) | P-value |
|---|
| miR-150-3p | 184 | Wnt signaling
pathway | 5 | 2.7 | 6.00E-02 |
| Neurotrophin
signaling pathway | 4 | 2.2 | 1.20E-01 |
| Ubiquitin mediated
proteolysis | 4 | 2.2 | 1.50E-01 |
| Adherens
junction | 3 | 1.6 | 1.70E-01 |
| miR-181a-2-3p | 189 | Endocytosis | 6 | 3.2 | 2.90E-02 |
| Chemokine signaling
pathway | 6 | 3.2 | 3.00E-02 |
| Ubiquitin mediated
proteolysis | 5 | 2.6 | 3.90E-02 |
| Pancreatic
cancer | 4 | 2.1 | 3.00E-02 |
| Adherens
junction | 4 | 2.1 | 3.50E-02 |
| Nucleotide excision
repair | 3 | 1.6 | 6.40E-02 |
| miR-205-3p | 944 | Pathways in
cancer | 19 | 2.0 | 2.50E-01 |
| MAPK signaling
pathway | 17 | 1.8 | 1.70E-01 |
| Wnt signaling
pathway | 15 | 1.6 | 9.20E-03 |
| miR-21-3p | 210 | Cell adhesion
molecules | 7 | 3.3 | 4.70E-03 |
| Ubiquitin mediated
proteolysis | 6 | 2.9 | 2.30E-02 |
| Long-term
potentiation | 5 | 2.4 | 8.60E-03 |
| Oocyte meiosis | 5 | 2.4 | 4.20E-02 |
| miR-29b-1-5p | 265 | Insulin signaling
pathway | 5 | 1.9 | 8.50E-02 |
| Cell cycle | 4 | 1.5 | 2.00E-01 |
| Wnt signaling
pathway | 4 | 1.5 | 2.90E-01 |
| Jak-STAT signaling
pathway | 4 | 1.5 | 3.00E-01 |
| mir-3127-5p | 205 | – | – | – | – |
| miR-371a-5p | 351 | Spliceosome | 8 | 2.3 | 4.20E-03 |
| Wnt signaling
pathway | 7 | 2.0 | 3.60E-02 |
| mir-3663-3p | 305 | MAPK signaling
pathway | 12 | 3.9 | 5.90E-03 |
| Pathways in
cancer | 11 | 3.6 | 5.50E-02 |
| Neurotrophin
signaling pathway | 7 | 2.3 | 2.00E-02 |
| Pancreatic
cancer | 5 | 1.6 | 3.50E-02 |
| Chronic myeloid
leukemia | 5 | 1.6 | 4.00E-02 |
| mir-4298 | 185 | Oocyte meiosis | 5 | 2.7 | 8.70E-03 |
| Neuroactive ligand
receptor interaction | 5 | 2.7 | 1.20E-01 |
| Calcium signaling
pathway | 4 | 2.2 | 1.40E-01 |
|
Phosphatidylinositol signaling system | 3 | 1.6 | 1.10E-01 |
| miR-602 | 302 | MAPK signaling
pathway | 7 | 2.3 | 2.20E-01 |
| Insulin signaling
pathway | 6 | 2.0 | 5.30E-02 |
| Alzheimer’s
disease | 6 | 2.0 | 1.00E-01 |
| Calcium signaling
pathway | 6 | 2.0 | 1.30E-01 |
| Table VFunctional annotation chart for
miRNAs downregulated in response to troxerutin in
H2O2-exposed HDP cells. |
Table V
Functional annotation chart for
miRNAs downregulated in response to troxerutin in
H2O2-exposed HDP cells.
| microRNA | Putative target
genes (n) | KEGG pathway | Genes involved in
the term (n) | Involved
genes/total genes (%) | P-value |
|---|
| miR-1181 | 2 | – | – | – | – |
| miR-1202 | 241 | Pathways in
cancer | 8 | 3.3 | 1.50E-01 |
| Insulin signaling
pathway | 5 | 2.1 | 1.10E-01 |
|
Phosphatidylinositol signaling system | 4 | 1.7 | 7.60E-02 |
| ABC
transporters | 3 | 1.2 | 1.20E-01 |
| mTOR signaling
pathway | 3 | 1.2 | 1.50E-01 |
| Inositol phosphate
metabolism | 3 | 1.2 | 1.60E-01 |
| miR-1224-5p | 213 | Axon guidance | 4 | 1.9 | 1.00E-01 |
| miR-1290 | 593 | Pathways in
cancer | 17 | 2.9 | 4.00E-02 |
| Insulin signaling
pathway | 13 | 2.2 | 7.60E-04 |
| Regulation of actin
cytoskeleton | 12 | 2.0 | 6.30E-02 |
| MAPK signaling
pathway | 12 | 2.0 | 1.90E-01 |
| ErbB signaling
pathway | 11 | 1.9 | 2.80E-04 |
| miR-135a-3p | 140 | – | – | – | – |
| miR-28-5p | 157 | MAPK signaling
pathway | 7 | 4.5 | 1.20E-02 |
| Axon guidance | 4 | 2.5 | 6.60E-02 |
| miR-378a-5p | 366 | Wnt signaling
pathway | 7 | 1.9 | 3.60E-02 |
| TGF-β signaling
pathway | 4 | 1.1 | 1.70E-01 |
| miR-4271 | 361 | Jak-STAT signaling
pathway | 7 | 1.9 | 7.80E-02 |
| Lysine
degradation | 4 | 1.1 | 5.20E-02 |
| miR-452-5p | 327 | Oocyte meiosis | 8 | 2.3 | 1.30E-03 |
| Wnt signaling
pathway | 7 | 2.0 | 2.60E-02 |
| ECM-receptor
interaction | 5 | 1.4 | 3.80E-02 |
| Small cell lung
cancer | 5 | 1.4 | 3.80E-02 |
| miR-572 | 6 | – | – | – | – |
| miR-575 | 241 | MAPK signaling
pathway | 8 | 3.3 | 7.70E-02 |
| Prostate
cancer | 6 | 2.5 | 7.70E-03 |
| Melanoma | 5 | 2.1 | 1.70E-02 |
| Cell cycle | 5 | 2.1 | 9.60E-02 |
|
Aldosterone-regulated sodium
reabsorption | 4 | 1.7 | 1.90E-02 |
| mTOR signaling
pathway | 4 | 1.7 | 3.50E-02 |
| Androgen and
estrogen metabolism | 3 | 1.2 | 9.40E-02 |
| miR-629-3p | 441 | PPAR signaling
pathway | 6 | 1.4 | 1.20E-02 |
| miR-939 | 365 | Calcium signaling
pathway | 10 | 2.4 | 1.30E-02 |
| Regulation of actin
cytoskeleton | 9 | 2.1 | 8.90E-02 |
| ErbB signaling
pathway | 5 | 1.2 | 1.20E-01 |
| p53 signaling
pathway | 4 | 0.9 | 1.80E-01 |
| Wnt signaling
pathway | 6 | 1.4 | 2.20E-01 |
| miR-940 | – | – | – | – | – |
Discussion
In the present study, the protective effect of
troxerutin against H2O2-induced oxidative
stress in HDP cells was confirmed using biochemical assays.
Notably, pretreatment with troxerutin decreased the cell death, ROS
production and cellular senescence, which was mediated by exposure
to H2O2. Although the specific signaling
pathways involved in the protective effect were not demonstrated,
the findings of the present study are important in that they
identify troxerutin as a candidate agent for use in the prevention
and/or treatment of alopecia. A growing body of evidence suggests
the role of oxidative stress in alopecia, and that the prevention
of oxidative stress may offer novel strategies for the intervention
and reversal of alopecia and even graying of hair (17). A previous case study confirmed
increased oxidative stress in alopecia areata patients compared
with healthy individuals (29). In
addition, a study using a mouse model demonstrated that hair
dye-induced hair loss is caused predominantly by
H2O2-induced oxidative stress (30). Oxidative stress stimulates the
production of a known inhibitor of hair follicles, tumor growth
factor-β, in DPC cells, which induces the onset of androgenic
alopecia (24). Our previous
studies demonstrated that troxerutin has a photoprotective effect
against UV radiation on dermal fibroblasts and keratinocytes
(6,7), and several clinical and theoretical
reports have revealed that UV radiation has negative effects on
hair growth through the induction of oxidative stress, acute
telogen effluvium and follicular micro-inflammation in follicular
stem cells (31–33). Therefore, countering oxidative
stress can be considered an important strategy to overcome
stress-or androgen-dependent alopecia, and the results of the
present study confirmed that troxerutin inhibited oxidative
stress-induced cellular damage in the DPC cells. In addition, our
previous studies and the present studies demonstrated low levels of
cytotoxicity of troxerutin on dermal fibroblasts, keratinocytes and
DP cells (6,7). Therefore, further investigation of
the clinical effect of topical application of troxerutin to the
scalp is required.
Using miRNA microarray analysis, the present study
identified 24 miRNAs in the HDP cells treated with troxerutin and
H2O2, which were differentially expressed
compared with the cells treated with H2O2
only. Of these, has-miR-602 was the most markedly upregulated by
troxerutin in the H2O2-treated HDP cells
(6.91-fold), and has been reported to downregulate the expression
of the RASSF1A and TP73 tumor suppressor genes (34). Several reports have revealed that
H2O2 induces the expression of TP73 (35) and that the anticancer drug
cisplatin, which has been reported to induce alopecia in patients,
stimulates ROS-induced apoptosis and functionally upregulates the
expression of p73 (36–38). Although the cellular functions of
miR-602, RASSF1A and TP73 have not been investigated in DP cells,
the data of the present study suggested that the interaction
between miR-602 and the two genes may be functionally involved in
ROS-induced cellular stress and even alopecia-associated
mechanisms. The biological functions of has-miR-575, which was the
most downregulated miRNA in the results of the present study, have
not been reported previously; however, it may regulate
H2O2-mediated cellular stress. PDPK1, also
termed PDK1, is a putative target of miR-575 (Table III) and a well-known kinase,
which phosphorylates and activates Akt1 kinase, induces cell
proliferation and protects against
H2O2-induced apoptosis (39,40).
The present study also classified the biological functions of
differentially expressed miRNAs by troxerutin in the
H2O2-treated HDP cells. KEGG pathway analysis
of the target genes of the upregulated and downregulated miRNAs
revealed 18 and 23 pathways, respectively, were statistically
enriched. Among these, the WNT and MAPK signaling pathways, which
were the most markedly enriched pathways associated with the target
genes of the upregulated and downregulated miRNAs, are involved in
the regulation of H2O2-mediated cellular
stress, including apoptosis and antioxidative mechanisms (41–45),
suggesting that the miRNAs altered by troxerutin may be involved in
protective mechanisms against H2O2-induced
damage through the regulation of these pathways.
In conclusion, the present study revealed a novel
role of troxerutin as a putative antioxidant agent in HDP cells. In
addition, the results revealed 24 differentially expressed miRNAs
and determined the putative involvement of 18 signaling pathways
associated with upregulated miRNAs and 23 signaling pathways
associated with downregulated miRNAs in the troxerutin mediated
protective effect against H2O2-induced cell
damage. Although further experiments are required to confirm the
differentially expressed miRNAs and their target genes, the results
of the present study may assist in elucidating the mechanism
underlying the troxerutin-mediated protection and miRNA-associated
signaling pathways in HDP cells.
Acknowledgments
This study was supported by a grant from the Korean
Health Technology R&D Project, Ministry of Health and Welfare,
Republic of Korea (grant no. HN13C0075). Dr Seunghee Bae was
supported by the KU Research Professor Program of Konkuk
University.
References
|
1
|
Marzin D, Phi HV, Olivier P and Sauzieres
J: Study of mutagenic activity of troxerutin, a flavonoid
derivative. Toxicol Lett. 35:297–305. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Boisseau MR, Taccoen A, Garreau C, Vergnes
C, Roudaut MF and Garreau-Gomez B: Fibrinolysis and hemorheology in
chronic venous insufficiency: a double blind study of troxerutin
efficiency. J Cardiovasc Surg (Torino). 36:369–374. 1995.
|
|
3
|
Lu J, Wu DM, Zheng ZH, Zheng YL, Hu B and
Zhang ZF: Troxerutin protects against high cholesterol-induced
cognitive deficits in mice. Brain. 134:783–797. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lu J, Wu DM, Zheng YL, et al: Troxerutin
counteracts domoic acid induced memory deficits in mice by
inhibiting CCAAT/enhancer binding protein beta-mediated
inflammatory response and oxidative stress. J Immunol.
190:3466–3479. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Babri S, Amani M, Mohaddes G, Alihemmati A
and Ebrahimi H: Protective effects of troxerutin on beta-amyloid
(1–42)-induced impairments of spatial learning and memory in rats.
Neurophysiology. 44:387–393. 2012. View Article : Google Scholar
|
|
6
|
Lee KS, Cha HJ, Lee GT, et al: Troxerutin
induces protective effects against ultraviolet B radiation through
the alteration of microRNA expression in human HaCaT keratinocyte
cells. Int J Mol Med. 33:934–942. 2014.PubMed/NCBI
|
|
7
|
Cha HJ, Lee KS, Lee GT, et al: Altered
miRNA expression profiles are involved in the protective effects of
troxerutin against ultraviolet B radiation in normal human dermal
fibroblasts. Int J Mol Med. 33:957–963. 2014.PubMed/NCBI
|
|
8
|
Ping X, Junqing J, Junfeng J and Enjin J:
Radioprotective effects of troxerutin against gamma irradiation in
V79 cells and mice. Asian Pac J Cancer Prev. 12:2593–2596.
2011.
|
|
9
|
Maurya DK, Salvi VP and Krishnan Nair CK:
Radioprotection of normal tissues in tumor bearing mice by
troxerutin. J Radiat Res. 45:221–228. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Flore R, Gerardino L, Santoliquido A, et
al: Enhanced oxidative stress in workers with a standing
occupation. Occup Environ Med. 61:548–550. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hladovec J: Protective effect of
oxygen-derived free radical scavengers on the endothelium in vivo.
Physiol Bohemoslov. 35:97–103. 1986.PubMed/NCBI
|
|
12
|
Lu J, Wu DM, Hu B, Zheng YL, Zhang ZF and
Wang YJ: NGF-Dependent activation of TrkA pathway: A mechanism for
the neuroprotective effect of troxerutin in D-galactose-treated
mice. Brain Pathol. 20:952–965. 2010.PubMed/NCBI
|
|
13
|
Heck DE, Vetrano AM, Mariano TM and Laskin
JD: UVB light stimulates production of reactive oxygen species:
unexpected role for catalase. J Biol Chem. 278:22432–22436. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Leach JK, Van Tuyle G, Lin PS,
Schmidt-Ullrich R and Mikkelsen RB: Ionizing radiation induced,
mitochondria-dependent generation of reactive oxygen/nitrogen.
Cancer Res. 61:3894–3901. 2001.PubMed/NCBI
|
|
15
|
Driskell RR, Clavel C, Rendl M and Watt
FM: Hair follicle dermal papilla cells at a glance. J Cell Sci.
124:1179–1182. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hibberts NA, Howell AE and Randall VA:
Balding hair follicle dermal papilla cells contain higher levels of
androgen receptors than those from non balding scalp. J Endocrinol.
156:59–65. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Trüeb RM: Oxidative stress in ageing of
hair. Int J Trichology. 1:6–14. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ripple MO, Henry WF, Rago RP and Wilding
G: Prooxidant-antioxidant shift induced by androgen treatment of
human prostate carcinoma cells. J Natl Cancer Inst. 89:40–48. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Naito A, Midorikawa T, Yoshino T and
Ohdera M: Lipid peroxides induce early onset of catagen phase in
murine hair cycles. Int J Mol Med. 22:725–729. 2008.PubMed/NCBI
|
|
20
|
Bahta AW, Farjo N, Farjo B and Philpott
MP: Premature senescence of balding dermal papilla cells in vitro
is associated with p16 (INK4a) expression. J Invest Dermatol.
128:1088–1094. 2008. View Article : Google Scholar
|
|
21
|
Lee EJ, Cha HJ, Ahn KJ, An IS, An S and
Bae S: Oridonin exerts protective effects against hydrogen
peroxideinduced damage by altering microRNA expression profiles in
human dermal fibroblasts. Int J Mol Med. 32:1345–1354.
2013.PubMed/NCBI
|
|
22
|
Huang DW, Sherman BT, Tan Q, et al: The
DAVID Gene Functional Classification Tool: a novel biological
module-centric algorithm to functionally analyze large gene lists.
Genome Biol. 8:R1832007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Luanpitpong S, Nimmannit U, Chanvorachote
P, et al: Hydroxyl radical mediates cisplatin-induced apoptosis in
human hair follicle dermal papilla cells and keratinocytes through
Bcl-2-dependent mechanism. Apoptosis. 16:769–782. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shin H, Yoo HG, Inui S, et al: Induction
of transforming growth factor-beta 1 by androgen is mediated by
reactive oxygen species in hair follicle dermal papilla cells. BMB
Rep. 46:460–464. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Colavitti R and Finkel T: Reactive oxygen
species as mediators of cellular senescence. IUBMB Life.
57:277–281. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Baehrecke EH: miRNAs: micro managers of
programmed cell death. Curr Biol. 13:R473–R475. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang Z, Liu Y, Han N, et al: Profiles of
oxidative stress-related microRNA and mRNA expression in auditory
cells. Brain Res. 1346:14–25. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Faraonio R, Salerno P, Passaro F, et al: A
set of miRNAs participates in the cellular senescence program in
human diploid fibroblasts. Cell Death Differ. 19:713–721. 2012.
View Article : Google Scholar :
|
|
29
|
Bakry OA, Elshazly RM, Shoeib MA and Gooda
A: Oxidative stress in alopecia areata: a case-control study. Am J
Clin Dermatol. 15:57–64. 2014. View Article : Google Scholar
|
|
30
|
Seo JA, Bae IH, Jang WH, et al: Hydrogen
peroxide and monoethanolamine are the key causative ingredients for
hair dye-induced dermatitis and hair loss. J Dermatol Sci.
66:12–19. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Trüeb RM: Is androgenetic alopecia a
photoaggravated dermatosis? Dermatology. 207:343–348. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Camacho F, Moreno JC and García-Hernández
MJ: Telogen alopecia from UV rays. Arch Dermatol. 132:1398–1399.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Johnsson A, Kjeldstad B and Melø TB:
Fluorescence from pilosebaceous follicles. Arch Dermatol Res.
279:190–193. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yang L, Ma Z, Wang D, Zhao W, Chen L and
Wang G: MicroRNA-602 regulating tumor suppressive gene RASSF1A is
overexpressed in hepatitis B virus-infected liver and
hepatocellular carcinoma. Cancer Biol Ther. 9:803–808. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Singh M, Sharma H and Singh N: Hydrogen
peroxide induces. apoptosis in HeLa cells through mitochondrial
pathway. Mitochondrion. 7:367–373. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Surendiran A, Balamurugan N, Gunaseelan K,
Akhtar S, Reddy KS and Adithan C: Adverse drug reaction profile of
cisplatin-based chemotherapy regimen in a tertiary care hospital in
India: An evaluative study. Indian J Pharmacol. 42:40–43. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Casares C, Ramírez-Camacho R, Trinidad A,
Roldán A, Jorge E and García-Berrocal JR: Reactive oxygen species
in apoptosis induced by cisplatin: review of physiopathological
mechanisms in animal models. Eur Arch Otorhinolaryngol.
269:2455–2459. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Gong JG, Costanzo A, Yang HQ, et al: The
tyrosine kinase c-Abl regulates p73 in apoptotic response to
cisplatin-induced DNA damage. Nature. 399:806–809. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang Z, Cui M, Sun L, et al:
Angiopoietin-1 protects H9c2 cells from
H2O2-induced apoptosis through AKT signaling.
Biochem Biophys Res Commun. 359:685–690. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Mora A, Komander D, van Aalten DM and
Alessi DR: PDK1, the master regulator of AGC kinase signal
transduction. Semin Cell Dev Biol. 15:161–170. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Finkel T: Signal transduction by reactive
oxygen species. J Cell Biol. 194:7–15. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Shin SY, Kim CG, Jho EH, et al: Hydrogen
peroxide negatively modulates Wnt signaling through downregulation
of beta-catenin. Cancer Lett. 212:225–231. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Korswagen HC: Regulation of the
Wnt/beta-catenin pathway by redox signaling. Dev Cell. 10:687–688.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Song X, Xu A, Pan W, et al: Minocycline
protects melanocytes against H2O2-induced
cell death via JNK and p38 MAPK pathways. Int J Mol Med. 22:9–16.
2008.PubMed/NCBI
|
|
45
|
Ismail NS, Pravda EA, Li D, Shih SC and
Dallabrida SM: Angiopoietin-1 reduces
H2O2-induced increases in reactive oxygen
species and oxidative damage to skin cells. J Invest Dermatol.
130:1307–1317. 2010. View Article : Google Scholar : PubMed/NCBI
|