Introduction
The progressive loss of extracellular matrix (ECM)
in cartilage is the characteristic lesion in osteoarthritis (OA).
Type II collagen and aggrecan are the main components of cartilage
ECM, and decreased production of either may irreversibly lead to
cartilage degradation. Transforming growth factor-β (TGF-β) is an
important regulator of the anabolic metabolism of articular
cartilage. TGF-β enhances ECM production by increasing synthesis of
type II collagen and aggrecan, thereby preventing cartilage
degradation (1,2). Reduced type II collagen and aggrecan
expression is a characteristic of aged, osteoarthritic and passaged
chondrocytes. However, TGF-β can stimulate expression of type II
collagen and aggrecan in these cells (3). Furthermore, bone-marrow mesenchymal
stem cells exposed to TGF-β express chondro-genic markers (type II
collagen and aggrecan), implying their differentiation into
chondrocytes (4,5). In addition, levels of type II
collagen and aggrecan in bovine intervertebral disc cells increase
following TGF-β1 exposure, suggesting that it can also act in the
prevention of degenerative disc disease (6).
TGF-β activates the Smad2/3 signaling pathway,
stimulating expression of type II collagen α1, aggrecan and sex
determining region Y-box 9 expression in human OA and bovine
chondrocytes (7). TGF-β can also
activate extracellular signal-regulated kinase (ERK)1/2 to regulate
anabolic activity through upregulation of TIMP-3 expression and
chondrocyte cell growth (8,9).
Furthermore, TGF-β1 can activate Smad2 and ERK1/2 signaling
pathways in human mesenchymal stem cells, subsequently increasing
the expression of type II collagen and aggrecan (10). Only a few studies have described
the role of ERK1/2 and Smad2/3 signaling pathways in TGF-β1-induced
type II collagen and aggrecan expression in rat chondrocytes
(8,11).
The present study investigated the effect of TGF-β1
on type II collagen and aggrecan expression in rat chondrocytes as
well as the possible mechanistic involvement of ERK1/2 and Smad2/3
signaling pathways. For this, either ERK1/2 or Smad2/3 signaling
pathways were blocked with small molecules, and the effect on
TGF-β1-induced type II collagen and aggrecan expression was
investigated. A previous study by our group suggested that
inhibition of ERK1/2 signaling may be used for the pharmacological
intervention of cartilage breakdown in OA (12). In addition to the inhibition of the
ERK1/2 signaling pathway, subsequent suppression of TGF-β1-induced
anabolic activity should be considered according to the results of
the present study.
Materials and methods
Chondrocyte isolation and culture
A total of 20 female Sprague-Dawley rats (weight,
180–200 g) were supplied by the Animal Center, Shanghai Jiao Tong
University School of Medicine (Shanghai, China). The rats were
sacrificed by cervical dislocation prior to surgical removal of
normal rat knee cartilage from the tibial platform and the femoral
condyle. The experiments were conducted with approval from the
Shanghai Jiao Tong University School of Medicine ethics committee.
Cartilage pieces were washed with phosphate-buffered saline (PBS)
twice and treated with 0.2 mg/ml collagenase (Sigma-Aldrich, St.
Louis, MO, USA) in serum-free Dulbecco's modified Eagle's medium
(DMEM; Gibco Life Technologies, Carlsbad, CA, USA) overnight at
37°C. The cells were collected by filtering the disaggregated
tissue through a 200-mesh nylon cell strainer, centrifugation at
1,000 × g for 5 min and two washes with PBS. Finally, the cells
were re-suspended and cultured in DMEM supplemented with 10%
(vol/vol) fetal bovine serum (Invitrogen Life Technologies,
Carlsbad, CA, USA), plus 1% penicillin and streptomycin (Gibco Life
Technologies). The culture medium was replaced every second day.
The chondrocytic phenotype of the cultured cells was confirmed by
positive immunostaining for type II collagen. Briefly, cells
cultured on glass slides were incubated overnight at 4°C with
anti-rat type II collagen (1:300 dilution; cat. no. ab34712; Abcam,
Cambridge, UK), and immunostaining was performed using
immunostaining reagent (Dolichos bifows agglutini
Immunohistochemistry kit; Wuhan Boster Biological Technology, Ltd.,
Wuhan, China), according to the manufacturer's instructions.
Positive immunostaining of collagen II was detected using an
inverted fluorescence microscope (IX70; Olympus, Tokyo, Japan).
Toluidine blue staining (Shanghai Chemical Reagent Co., Ltd.,
Shanghai, China) of glycosaminoglycans was also conducted.
First-passage chondrocytes were used in all experiments.
Chondrocyte treatment
First-passage chondrocytes were trypsinized and
seeded into six-well plates (3×105 cells/well) for 24 h.
Chondrocytes were maintained in serum-deficient DMEM for 24 h and
subjected to a variety of treatments. Chondrocytes were cultured
with or without 10 µg/ml recombinant human TGF-β1
(Perprotech, Rocky Hill, NJ, USA) for 48 h. Type II collagen and
aggrecan expression was evaluated by reverse transcription
quantitative polymerase chain reaction (RT-qPCR) and western
blotting. Chondrocytes were cultured with 10 µg/ml TGF-β1.
Following 0, 1, 5, 10, 15, 30, 60 and 180 min of incubation,
phosphorylated (p)-Smad3, Smad2/3, p-ERK1/2, and ERK1/2 levels were
examined by western blotting. For inhibition studies, chondrocytes
were pre-treated in serum-free medium with or without TGF-β
receptor I (ALK5) kinase inhibitor SB525334 (1 µM;
Selleckchem, Houston, TX, USA), the pharmacological ERK1/2
inhibitor PD98059 (10 µM; Promega Corporation, Madison, WI,
USA), and pharmacological Smad3 phosphorylation inhibitor SIS3 (10
µM; Santa Cruz Biotechnology, Inc. Dallas, TX, USA) for 60
min and then stimulated with or without 10 µg/ml TGF-β1 for
48 h. Expression of type II collagen and aggrecan was evaluated by
RT-qPCR and western blotting. To examine the involvement of the
ERK1/2 and Smad signaling pathways, chondrocytes were pre-treated
in serum-free medium with SB525334, PD98059 or SIS3 for 60 min,
followed by stimulation with 10 µg/ml TGF-β1. Following 0,
1, 5, 10, 15, 30, 60 or 180 min of incubation, p-Smad3, Smad2/3,
p-ERK1/2, and ERK1/2 levels were examined by western blotting.
RNA extraction and RT-qPCR
Total RNA was extracted from chondrocytes using
TRIzol reagent (Invitrogen Life Technologies) according to the
manufacturer's instructions. For first-strand cDNA synthesis, 2
µg mRNA was reverse-transcribed using the Reverse
Transcriptase Moloney murine leukemia virus (M-MLV) cDNA Synthesis
kit (Takara, Tokyo, Japan) according to the manufacturer's
instructions. Briefly, a 12-µl reaction mixture containing 2
µl oligo d(T)18 primer (50 µM), 2 µg total RNA
and RNase-free dH2O was incubated at 70°C for 10 min.
Subsequently, 1 µl deoxynucleotide triphosphate mixture (10
mM each), 4 µl 5X M-MLV buffer, 0.5 µl ribonuclease
inhibitor (40 U/µl), 1 µl reverse transcriptase M-MLV
(RNase H-free; 200 U/µl) and RNase-free dH2O were
added to a final volume of 20 µl and the mixture was
incubated for 60 min at 42°C. Next, the reaction was inactivated by
heating at 70°C for 15 min. Subsequently, 2 ml of the 20 ml reverse
transcription reaction was subjected to PCR amplification using PCR
Master Mix (cat. no. M7502, Promega Corporation) with initial
denaturation at 95°C for 10 min, then 35 cycles of 94°C for 30 sec,
60°C for 30 sec and 72°C for 45 sec. Products were quantified using
a melting curve analysis. Amplification of GAPDH was performed to
quantify PCR products and confirm the use of equal amounts of RNA.
Relative gene expression was analyzed using the 2ΔΔCt
method (13). Primer sequences
(Shanghai Sangon Biotechnology Co., Ltd., Shanghai, China) of rat
genes were as follows: Type II collagen forward,
5′-TCCTAAGGGTGCCAATGGTGA-3′, and reverse,
5′-GGACCAACTTTGCCTTGAGGAC-3′; aggrecan forward,
5′-TCCGCTGGTCTGATGGACAC-3′, and reverse,
5′-CCAGATCATCACTACGCAGTCCTC-3′; and GAPDH forward,
5′-CAAGTTCAACGGCACAGTCAAG-3′, and reverse,
5′-ACATACTCAGCACCAGCATCAC-3′.
Protein extraction and western blot
analysis
Whole-cell lysates were prepared using
radioimmunoprecipitation assay buffer [50 mM Tris-HCl (pH 7.4), 150
mM NaCl, 1% Triton X-100, 0.1% SDS, 2 mM EDTA and 2 mM
phenylmethanesulfonylfluoride] in the presence of protease and
phosphatase inhibitors. For the detection of p-Smad3, Smad2/3,
p-ERK1/2, ERK1/2 and aggrecan protein levels, equal amounts of
total cellular extracts from rat chondrocytes were then loaded on a
10% SDS-polyacrylamide gel, subjected to electrophoresis and
transferred to polyvinylidene fluoride membranes (EMD Millipore,
Billerica, MA, USA) by electroblotting. For type II collagen
protein levels, equal amounts of total cellular extracts were
separated by 8% SDS-polyacrylamide gel electrophoresis. Membranes
were then blocked with 5% skimmed milk. Polyclonal anti-rat type II
collagen (1:5,000 dilution; cat. no. ab34712; Abcam), polyclonal
anti-rat aggrecan (1:200 dilution; cat. no. 251591; Abbiotec, San
Diego, CA, USA), monoclonal anti-rat p-Smad3 (1:1,000 dilution;
cat. no. ab52903; Abcam), polyclonal anti-rat Smad2/3 (1:500
dilution; cat. no. R-1002–100; Novus, Littleton, CO, USA),
monoclonal anti-rat p-ERK1/2 (1:1,000 dilution; cat. no. ab32538;
Abcam) and polyclonal anti-rat ERK1/2 antibody (1:1,000; cat. no.
9102; Cell Signaling Technology Inc., Danvers, MA, USA) were
incubated with the membranes overnight at 4°C. The membranes were
then washed and reacted with secondary horseradish
peroxidase-conjugated secondary antibody [donkey anti-rabbit
immunoglobulin G for type II collagen, aggrecan, p-Smad3, Smad2/3,
p-ERK1/2, and ERK1/2 (1:2,000; cat. no. sc-2305; Santa Cruz
Biotechnology, Inc.)] at room temperature for 2 h. Finally, protein
bands were visualized by chemiluminescence (EMD Millipore) using
the Tanon 4200 SF Imaging Analysis system (Tanon Science and
Technology Co., Ltd., Shanghai, China). Western blots were
re-probed with a monoclonal anti-GAPDH antibody (1:1,000 dilution;
cat. no. 2118; Cell Signaling Technology Inc.) as a control.
Statistical analysis
All experiments were performed three times and the
results are expressed as the mean ± standard deviation. Statistical
significance was determined using the Mann-Whitney U test or
Kruskal-Wallis analysis of variance test, when appropriate, using
SPSS 13.0 statistical software (SPSS, Inc., Chicago, IL, USA).
P<0.05 was considered to indicate a statistically significant
difference between values.
Results
ERK1/2 and Smad2/3 signaling pathways
mediate TGF-β1-stimulated type II collagen and aggrecan expression
in rat chondrocytes
To confirm that TGF-β1 induced upregulation of type
II collagen and aggrecan expression in chondrocytes, rat
chondrocytes were cultured with or without TGF-β1 for 48 h. RT-qPCR
and western blotting were then performed to examine the expression
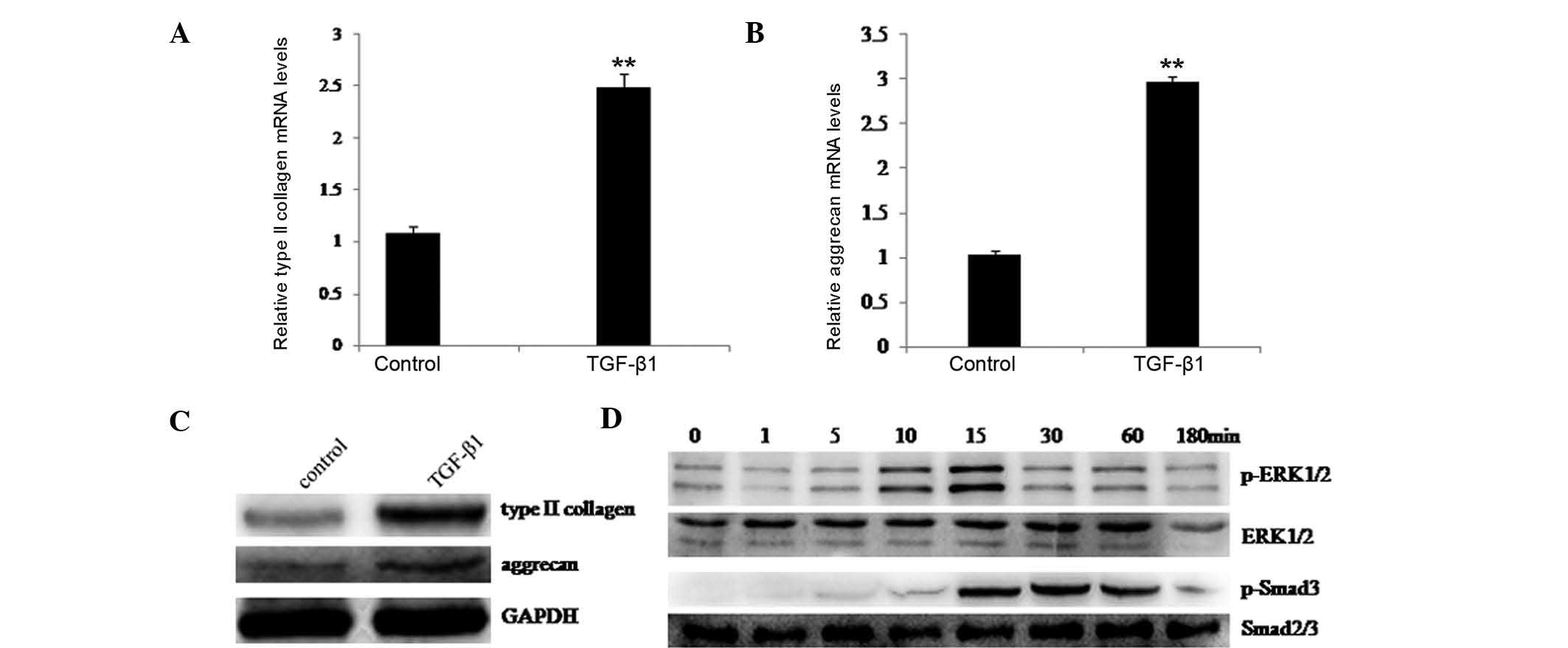
of type II collagen and aggrecan. As shown in Fig. 1A and B, significant upregulation of
type II collagen and aggrecan occurred following treatment with
TGF-β1. Western blotting also indicated that type II collagen and
aggrecan protein levels were increased following TGF-β1 treatment
(Fig. 1C). These results confirmed
that TGF-β1 stimulated type II collagen and aggrecan expression in
rat chondrocytes.
Furthermore, it was evaluated whether the ERK1/2 and
Smad2/3 signaling pathways were involved in TGF-β1 stimulation of
type II collagen and aggrecan. Rat chondrocytes were cultured in
the presence of TGF-β1, and at the indicated time-points, the
activation of ERK1/2 and phosphorylation of Smad3 were examined by
western blotting. Fig. 1D
demonstrates that TGF-β1 induced rapid phosphorylation of ERK1/2
and Smad3. These observations suggested that the ERK1/2 and Smad2/3
signaling pathways may have an important role in TGF-β1-stimulated
type II collagen and aggrecan induction in rat chondrocytes.
Blockade of TGF-β1 receptor represses the
stimulation of type II collagen and aggrecan expression
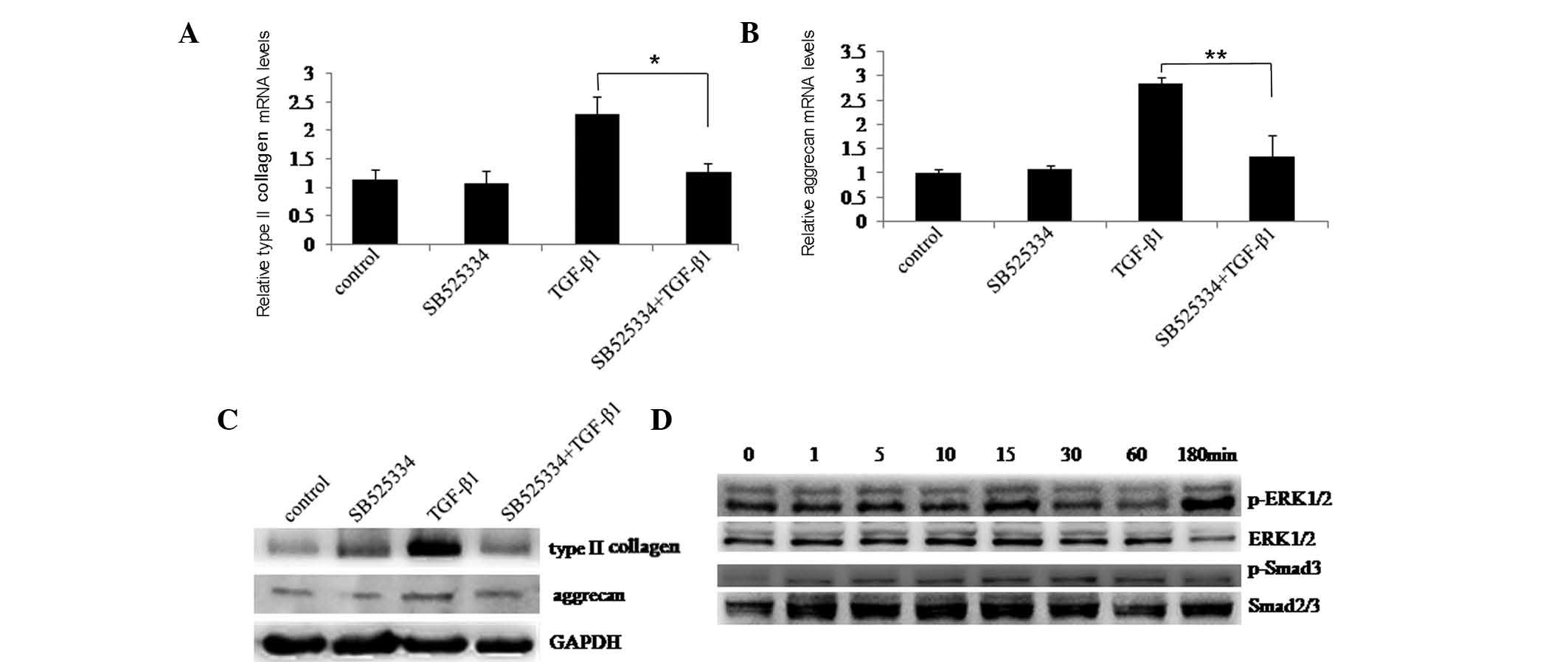
To further investigate the involvement of the ERK1/2
and Smad2/3 signaling pathways in TGF-β1-induced type II collagen
and aggrecan expression, chondrocytes were pre-treated with or
without ALK5 kinase inhibitor SB525334 and then stimulated with or
without TGF-β1. After 48 h, RT-qPCR and western blotting confirmed
that TGF-β1 stimulated type II collagen and aggrecan expression was
inhibited by SB525334 (Fig. 2A–C).
At the indicated time-points, western blotting demonstrated that
phosphorylation of ERK1/2 and Smad3 was blocked by SB525334
(Fig. 2D). These results further
validated the role of the ERK1/2 and Smad2/3 signaling pathways in
type II collagen and aggrecan induction by TGF-β1.
Blockade of TGF-β1-stimulated type II
collagen and aggrecan expression by inhibition of ERK1/2
signaling
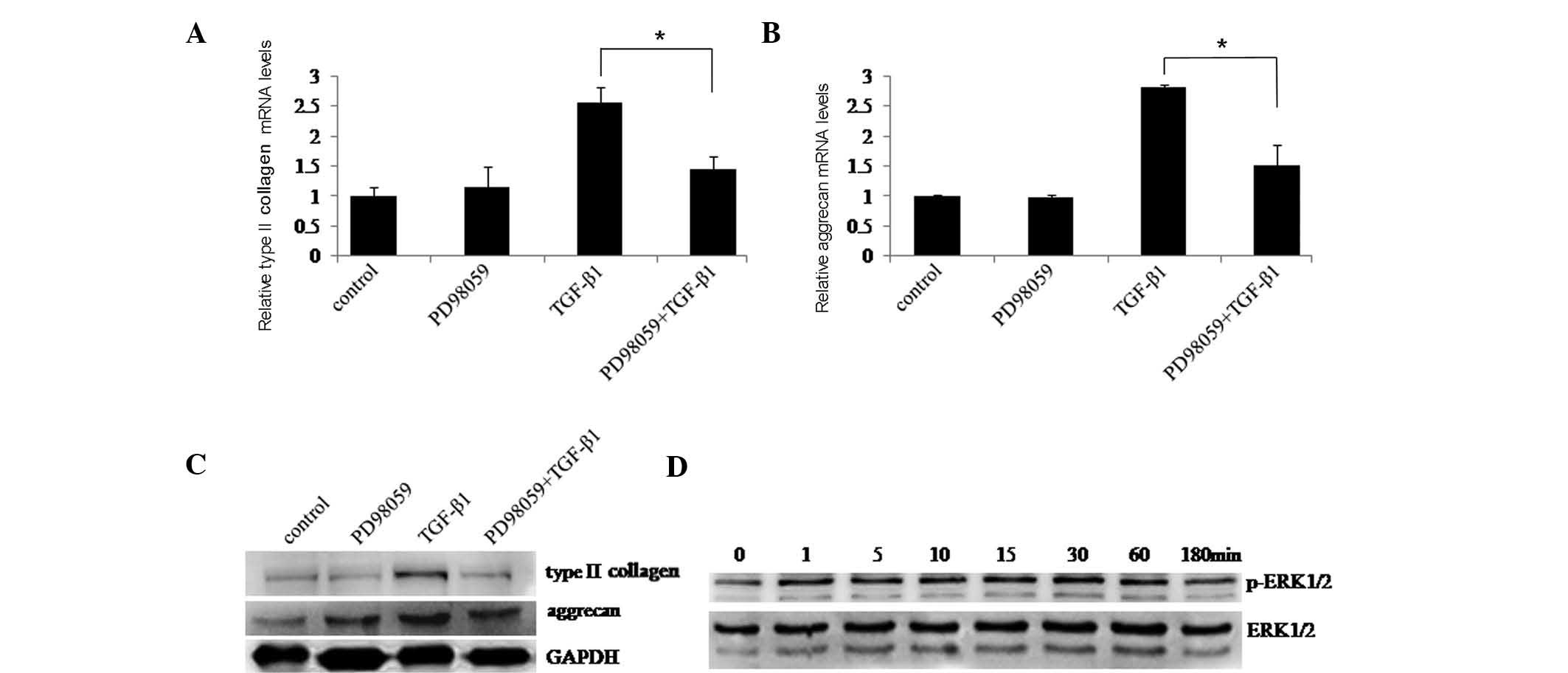
Next, the the role of ERK1/2 in type II collagen and
aggrecan induction by TGF-β1 was examined. Chondrocytes were
pre-treated with or without ERK1/2 inhibitor PD98059 followed by
culture with or without TGF-β1 for 48 h. RT-qPCR and western
blotting revealed that TGF-β1-induced expression of type II
collagen and aggrecan was significantly suppressed in the presence
of PD98059 (Fig. 3A–C). ERK1/2
signaling pathway activation was then assessed in chondrocytes
pre-treated with PD98059 followed by culture with TGF-β1.
Activation of the ERK1/2 signaling pathway was inhibited by PD98059
(Fig. 3D). These results suggested
that the ERK1/2 signaling pathway may have an important role in
type II collagen and aggrecan induction by TGF-β1.
Blockade of TGF-β1-stimulated type II
collagen and aggrecan expression by inhibition of p-Smad3
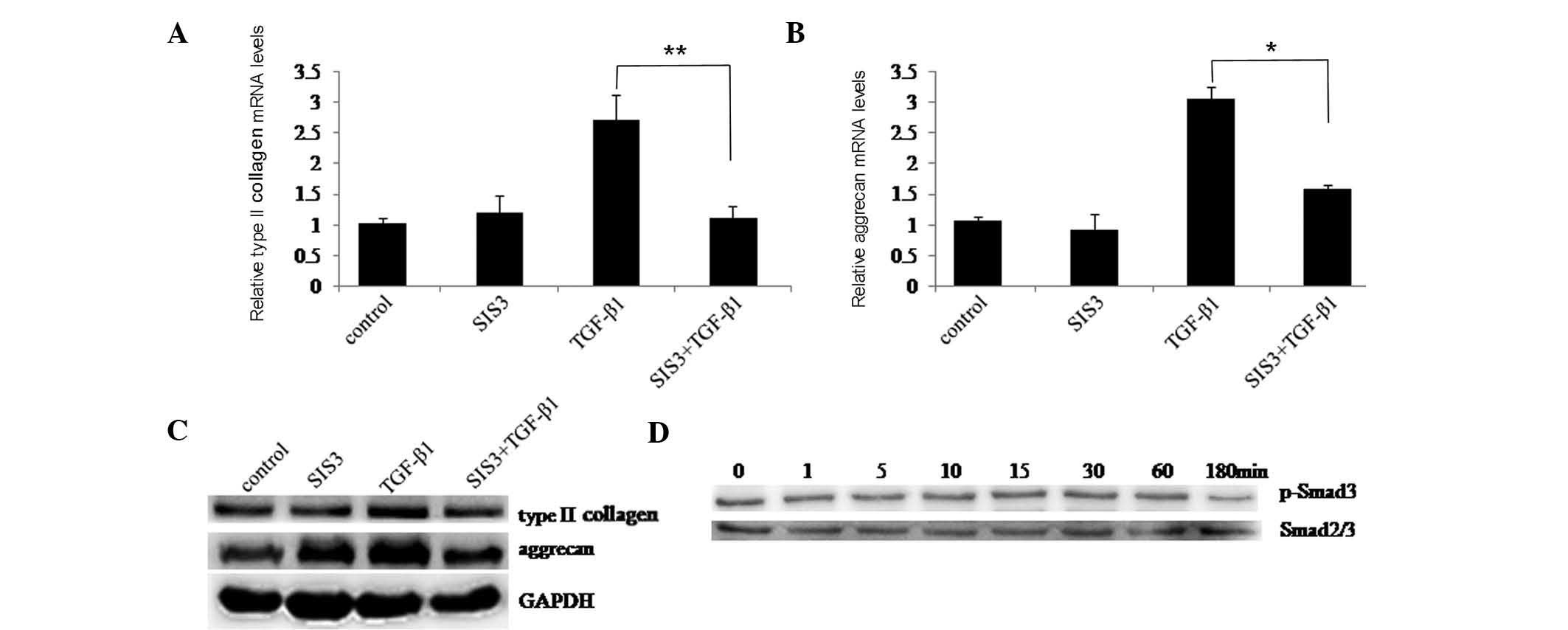
To determine whether the Smad2/3 signaling pathway
affected TGF-β1-induced type II collagen and aggrecan, SIS3 was
used to suppress Smad3 phosphorylation. First, chondrocytes were
cultured with or without TGF-β1 for 48 h following pre-treatment
with or without SIS3. RT-qPCR and western blotting indicated that
TGF-β1-induced type II collagen and aggrecan expression were
significantly inhibited by SIS3 (Fig.
4A–C). Next, chondrocytes were treated with TGF-β1 following
pre-treatment with SIS3. At the indicated time-points, western
blotting showed that Smad3 phosphorylation was markedly suppressed
by SIS3 (Fig. 4C). These results
suggested that the Smad2/3 signaling pathway may also have an
important role in type II collagen and aggrecan induction by
TGF-β1.
Discussion
TGF-β exerts profound effects on cellular
proliferation, differentiation and modulation of ECM, regulating
the anabolic process of articular cartilage and preventing
cartilage degradation. It can stimulate full-depth explants of
human OA knee articular cartilage to expression of type II collagen
and aggrecan (1). A study found
that adenoviral vector-mediated gene transduction of TGF-β1
increased type II collagen and aggrecan expression in normal and
osteoarthritic human chondrocytes (2). In addition, in bovine articular
chondrocytes, transgene-mediated TGF-β overexpression increased
type II collagen and aggrecan expression (14). In analogy with this the present
study showed that TGF-β1 significantly induced type II collagen and
aggrecan expression in rat chondrocytes. These results suggested
that TGF-β1 regulates the anabolic metabolism of articular
cartilage and prevents cartilage degradation.
TGF-β1 signals are transmitted through
phosphorylation of the Smad2/3 by activation of type I (TGF-βRI)
and type II (TGF-βRII) receptors (15–17).
Phosphorylated Smad2 and Smad3 form hetero-oligomeric complexes
with Smad4 and migrate to the nucleus, where they regulate the
transcription of target genes (17–19).
TGF-β also activates ERK1/2 to regulate the expression of target
genes and cellular proliferation (20–23).
Either or both the ERK1/2 or Smad2/3 signaling pathways are
involved in TGF-β1 sub-family-induced type II collagen gene
expression (24–26). The results of the present study
revealed that TGF-β1 rapidly induced phosphorylation of ERK1/2 and
Smad3, suggesting that activation of ERK1/2 as well as Smad2/3
signaling may account for TGF-β1-induced type II collagen and
aggrecan expression. Furthermore, the present study demonstrated
that the ALK5 inhibitor SB525334 significantly impaired
TGF-β1-induced type II collagen and aggrecan expression. In
addition, SB525334 suppressed TGF-β1-stimulated phosphorylation of
ERK1/2 and Smad3. In conclusion, these results strongly implicated
ERK1/2 and Smad2/3 signaling in TGF-β1-induced type II collagen and
aggrecan expression.
Additional evidence of a role for ERK1/2 signaling
in TGF-β1-induced type II collagen and aggrecan expression was
provided by the finding that PD98059 significantly inhibited
TGF-β1-induced type II collagen and aggrecan expression.
Furthermore, TGF-β1-stimulated phosphorylation of ERK1/2 was
inhibited by PD98059. This indicated that activation of ERK1/2
signaling is an essential mechanism by which TGF-β1 induces type II
collagen and aggrecan expression. Therefore, inhibition of ERK1/2
signaling may impair TGF-β1-stimulated anabolic metabolism of
articular cartilage. The present study also investigated inhibition
of Smad2/3 signaling pathway in TGF-β1-induced type II collagen and
aggrecan expression in rat chondrocytes using SIS3, a specific
inhibitor of Smad3 phosphorylation. The results showed that
TGF-β1-induced type II collagen and aggrecan expression was
significantly impaired by SIS3. Furthermore, TGF-β1-stimulated
phosphorylation of Smad3 was suppressed by SIS3. These results
further suggested a crucial role for Smad2/3 signaling in the
regulation of TGF-β1-induced type II collagen and aggrecan
expression.
The results of the present study strongly implicated
the ERK1/2 and Smad2/3 signaling pathways in TGF-β1-induced type II
collagen and aggrecan expression. The results provided a functional
link between ECM proteins, TGF-β1 activity and disease, suggesting
novel therapeutic opportunities for intervention in osteoarthritis.
The ERK1/2 signaling pathway transduces signals from the cell
membrane to the nucleus in response to a variety of stimuli,
controlling a wide spectrum of cellular processes, including
growth, differentiation and the expression of multiple genes. It is
well documented that interleukin (IL)-1β is an important mediator
of cartilage destruction during the OA process. IL-1β selectively
induces ERK1/2 activation and stimulates the production of matrix
metalloproteinases and aggrecanases (27,28),
which may contribute to cartilage loss and induce degradation of
cartilage. This suggests that ERK1/2 signaling may have an
important role in IL-1-induced degradation of cartilage. A previous
study by our group showed such a requirement of ERK1/2 pathways for
IL-β inhibition of type II collagen and aggrecan in human
chondrocytes (12). Therefore,
ERK1/2 represents a potential therapeutic drug target for OA.
PD98059 prevents cyclosporine A-induced nuclear
trans-location of Smad2/3 and apoptosis in renal proximal tubular
cells, suggesting the existence of crosstalk between ERK1/2 and
Smad2/3 (29). ERK1/2 is also
capable of phosphorylating Smads (30). Based on the results of the present
study, exploiting ERK1/2 as a therapeutic drug target for OA may
lead to decreased TGF-β1-induced anabolic metabolism or impaired
repair responses in chondrocytes through inhibition of ERK1/2 and
indirect inhibition of Smad2/3. Although TGF-β1 and IL-β have
opposing effects on type II collagen and aggrecan expression, they
exert their effects through the same signaling pathway, possibly by
activating different downstream transcription factors. Further
studies are required to examine the downstream transcription
factors influenced by ERK1/2 activation by various stimuli.
In conclusion, the present study showed that the
ERK1/2 and Smad2/3 signaling pathways are required for the
induction of type II collagen and aggrecan expression by TGF-β1.
This regulation of type II collagen and aggrecan expression may be
pivotal in TGF-β-induced ECM protein synthesis and maintenance, and
in the cartilage repair response. In addition, although inhibition
of ERK1/2 signaling represents a valuable potential therapeutic
drug target for OA (12), the side
effects of suppression of TGF-β1-induced metabolism of articular
cartilage should be considered.
Acknowledgments
The present study was supported by the National
Natural Science Foundation of China (grant no. 81101380).
References
|
1
|
Tchetina EV, Antoniou J, Tanzer M, Zukor
DJ and Poole AR: Transforming growth factor-beta2 suppresses
collagen cleavage in cultured human osteoarthritic cartilage,
reduces expression of genes associated with chondrocyte hypertrophy
and degradation, and increases prostaglandin E(2) production. Am J
Pathol. 168:131–140. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ulrich-Vinther M, Stengaard C, Schwarz EM,
Goldring MB and Soballe K: Adeno-associated vector mediated gene
transfer of transforming growth factor-beta1 to normal and
osteoarthritic human chondrocytes stimulates cartilage anabolism.
Eur Cell Mater. 10:40–50. 2005.PubMed/NCBI
|
|
3
|
Acosta CA, Izal I, Ripalda P,
Douglas-Price AL and Forriol F: Gene expression and proliferation
analysis in young, aged, and osteoarthritic sheep chondrocytes
effect of growth factor treatment. J Orthop Res. 24:2087–2094.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Huang CY, Hagar KL, Frost LE, Sun Y and
Cheung HS: Effects of cyclic compressive loading on chondrogenesis
of rabbit bone-marrow derived mesenchymal stem cells. Stem Cells.
22:313–323. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Morigele M, Shao Z, Zhang Z, Kaige M,
Zhang Y, Qiang W and Yang S: TGF-β1 induces a nucleus pulposus-like
phenotype in Notch 1 knockdown rabbit bone marrow mesenchymal stem
cells. Cell Biol Int. 37:820–825. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kwon YJ, Lee JW, Moon EJ, Chung YG, Kim OS
and Kim HJ: Anabolic effects of Peniel 2000, a peptide that
regulates TGF-β1 signaling on intervertebral disc degeneration.
Spine. 38:E49–E58. 2013. View Article : Google Scholar
|
|
7
|
Roman-Blas JA, Stokes DG and Jimenez SA:
Modulation of TGF-beta signaling by proinflammatory cytokines in
articular chondrocytes. Osteoarthritis Cartilage. 15:1367–1377.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Qureshi HY, Sylvester J, El Mabrouk M and
Zafarullah M: TGF-beta-induced expression of tissue inhibitor of
metalloproteinases-3 gene in chondrocytes is mediated by
extracellular signal-regulated kinase pathway and Sp1 transcription
factor. J Cell Physiol. 203:345–352. 2005. View Article : Google Scholar
|
|
9
|
Yonekura A, Osaki M, Hirota Y, Tsukazaki
T, Miyazaki Y, Matsumoto T, Ohtsuru A, Namba H, Shindo H and
Yamashita S: Transforming growth factor-beta stimulates articular
chon-drocyte cell growth through p44/42 MAP kinase (ERK)
activation. Endocr J. 46:545–553. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Re'em T, Kaminer-Israeli Y, Ruvinov E and
Cohen S: Chondrogenesis of hMSC in affinity-bound TGF-beta
scaffolds. Biomaterials. 33:751–761. 2012. View Article : Google Scholar
|
|
11
|
Qureshi HY, Ricci G and Zafarullah M: Smad
signaling pathway is a pivotal component of tissue inhibitor of
metalloproteinases-3 regulation by transforming growth factor beta
in human chondrocytes. Biochim Biophys Acta. 1783:1605–1612. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang X, Li F, Fan C, Wang C and Ruan H:
Effects and relationship of ERK1 and ERK2 in interleukin-1β-induced
alterations in MMP3, MMP13, type II collagen and aggrecan
expression in human chondrocytes. Int J Mol Med. 27:583–589.
2011.PubMed/NCBI
|
|
13
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative C(T) method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shi S, Mercer S, Eckert GJ and Trippel SB:
Regulation of articular chondrocyte aggrecan and collagen gene
expression by multiple growth factor gene transfer. J Orthop Res.
30:1026–1031. 2012. View Article : Google Scholar
|
|
15
|
Wrana JL, Attisano L, Wieser R, Ventura F
and Massagué J: Mechanism of activation of the TGF-beta receptor.
Nature. 370:341–347. 1994. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Heldin CH, Miyazono K and ten Dijke P:
TGF-beta signalling from cell membrane to nucleus through SMAD
proteins. Nature. 390:465–471. 1997. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shi Y and Massagué J: Mechanisms of
TGF-beta signaling from cell membrane to the nucleus. Cell.
113:685–700. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Itoh S, Itoh F, Goumans MJ and Ten Dijke
P: Signaling of transforming growth factor-beta family members
through Smad proteins. Eur J Biochem. 267:6954–6967. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Massagué J and Wotton D: Transcriptional
control by the TGF-beta/Smad signaling system. EMBO J.
19:1745–1754. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cailotto F, Bianchi A, Sebillaud S,
Venkatesan N, Moulin D, Jouzeau JY and Netter P: Inorganic
pyrophosphate generation by transforming growth factor-beta-1 is
mainly dependent on ANK induction by Ras/Raf-1/extracellular
signal-regulated kinase pathways in chondrocytes. Arthritis Res
Ther. 9:R1222007. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
O'Rear L, Longobardi L, Torello M, Law BK,
Moses HL, Chiarelli F and Spagnoli A: Signaling cross-talk between
IGF-binding protein-3 and transforming growth factor-(beta) in
mesenchymal chondroprogenitor cell growth. J Mol Endocrinol.
34:723–737. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shikhman AR, Brinson DC and Lotz MK:
Distinct pathways regulate facilitated glucose transport in human
articular chon-drocytes during anabolic and catabolic responses. Am
J Physiol Endocrinol Metab. 286:E980–E985. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Miyazaki Y, Tsukazaki T, Hirota Y,
Yonekura A, Osaki M, Shindo H and Yamashita S: Dexamethasone
inhibition of TGF beta-induced cell growth and type II collagen
mRNA expression through ERK-integrated AP-1 activity in cultured
rat articular chondrocytes. Osteoarthritis Cartilage. 8:378–385.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Schneiderbauer MM, Dutton CM and Scully
SP: Signaling “cross-talk” between TGF-beta1 and ECM signals in
chondrocytic cells. Cell Signal. 16:1133–1140. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang M, Zhou Q, Liang QQ, Li CG, Holz JD,
Tang D, Sheu TJ, Li TF, Shi Q and Wang YJ: IGF-1 regulation of type
II collagen and MMP-13 expression in rat endplate chondrocytes via
distinct signaling pathways. Osteoarthritis Cartilage. 17:100–106.
2009. View Article : Google Scholar
|
|
26
|
Park MS, Kim YH and Lee JW: FAK mediates
signal crosstalk between type II collagen and TGF-beta 1 cascades
in chon-drocytic cells. Matrix Biol. 29:135–142. 2010. View Article : Google Scholar
|
|
27
|
Kimura H, Yukitake H, Suzuki H, Tajima Y,
Gomaibashi K, Morimoto S, Funabashi Y, Yamada K and Takizawa M: The
chondroprotective agent ITZ-1 inhibits interleukin-1beta-induced
matrix metalloproteinase-13 production and suppresses nitric
oxide-induced chondrocyte death. J Pharmacol Sci. 110:201–211.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Megías J, Guillén MI, Bru A, Gomar F and
Alcaraz MJ: The carbon monoxide-releasing molecule
tricarbonyldichlororuth enium(II) dimer protects human
osteoarthritic chondrocytes and cartilage from the catabolic
actions of interleukin-1beta. J Pharmacol Exp Ther. 325:56–61.
2008. View Article : Google Scholar
|
|
29
|
Iwayama H, Sakamoto T, Nawa A and Ueda N:
Crosstalk between Smad and Mitogen-Activated Protein Kinases for
the Regulation of Apoptosis in Cyclosporine A- Induced Renal
Tubular Injury. Nephron Extra. 1:178–189. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Javelaud D and Mauviel A: Crosstalk
mechanisms between the mitogen-activated protein kinase pathways
and Smad signaling downstream of TGF-beta: Implications for
carcinogenesis. Oncogene. 24:5742–5750. 2005. View Article : Google Scholar : PubMed/NCBI
|