Introduction
Inflammation is important in the development and
progression of atherosclerosis (1,2). The
inflammatory processes are characterized by increased circulating
levels of pro-inflammatory cytokines, including interleukin (IL)
-1β, IL-6, tumor necrosis factor-α (TNF-α) and inducible nitric
oxide synthase (iNOS), soluble adhesion molecules, and
cytokine-responsive acute phase protein such as C-reactive protein
(CRP) (3,4).
Fibrinogen, the acute phase reactant in humans, is
derived predominantly from hepatocytes in response to IL-6 and is
then secreted into the systemic circulation (5). Recent studies have reported that in
addition to its predictive role in determining cardiovascular risk,
fibrinogen also exerts a direct pro-inflammatory role on the
different vascular cells implicated in atherogenesis (6,7).
Fibrinogen stimulates the production of inflammatory cytokines
(IL-6, TNF-α, IL-1β, IL-8 and endothelin-1) in endothelial cells
(7,8), macrophages (9), monocytes (10) and neutrophils (11). In addition, fibrinogen participates
in the formation of atherosclerosis lesions through promoting
platelet aggregation (12),
increasing plasma viscosity (13),
injuring endothelial cells (14,15),
and stimulating migration and proliferation of vascular smooth
muscle cells (VSMCs) (16,17). Furthermore, fibrinogen is degraded
by plasmin to a series of fibrin degradation products (FDPs), which
share similar bioactivities to fibrinogen and also are associated
with the progression of atherosclerosis (17,18).
Our previous studies showed that fibrinogen and FDP upregulate the
expression of CRP, IL-6, TNF-α and iNOS in VSMCs (19,20).
3-Hydroxy-3-methylglutaryl-coenzyme A reductase
inhibitors (statins) effectively lower the serum cholesterol level
and reduce cardiovascular events and mortality in patients with or
without coronary artery disease (21). The beneficial effects of statins
have also been characterized by their non-lipid related mechanisms,
such as preventing inflammation, improving endothelial function,
inhibiting activation of VSMCs, stabilizing atherosclerotic
plaques, and preventing thrombosis formation (21,22).
Since chronic inflammation in the vessel wall is important in
atherogenesis, and hyperfibrinogenemia is associated with
cardiovascular disorders, including atherosclerosis, the aim of
this study was to observe whether pravastatin regulated fibrinogen-
and FDP-induced production of IL-6, TNF-α and iNOS, and to
highlight its anti-inflammatory effect and anti-atherosclerotic
action.
Materials and methods
Reagents
Plasminogen-depleted fibrinogen and plasmin were
obtained from Calbiochem (Merck, Darmstadt, Germany). Fetal bovine
serum (FBS) was purchased from Hyclone (Logan, UT, USA). Dulbecco's
modified Eagle's medium (DMEM), penicillin and streptomycin were
produced by Gibco-BRL (Carlsbad, CA, USA). Pravastatin and thrombin
were purchased from Sigma-Aldrich (St. Louis, MO, USA). Rat IL-6
and TNF-α ELISA kits were obtained from Alpha Diagnostics
International (San Antonio, TX, USA). Polyclonal rabbit anti-rat
iNOS antibody (cat. no. bs-0162R) was provided by Beijing
Biosynthesis Biotechnology (Beijing, China). TRIzol reagent kits
were provided by Invitrogen Life Technologies (Carlsbad, CA, USA).
The Revert Aid™ First Strand cDNA Synthesis kit was obtained from
Fermentas (St. Leon-Rot, Germany). Agarose gels were from Spanish
Biochemicals Corp. (Madrid, Spain).
Culture of rat VSMCs
Male Sprague-Dawley rats (n=12; age, 2–3 months;
weight, 100–150 g) were obtained from the Laboratory Animal Center
of Xi'an Jiaotong University School of Medicine (Xi'an, China).
VSMCs were prepared from the thoracic aorta of using the explant
method as described previously (23). Briefly, rats were anesthetized with
intraperitoneal injection of sodium pentobarbital (30 mg/kg;
Sigma-Aldrich). The thoracic aorta was removed and freed of
connective tissue and adherent fat. The endothelial cell layer of
the intima was removed mechanically, and the aortic artery was cut
into sections of ~3 mm. These were then placed in DMEM supplemented
with 10% FBS, 100 U/ml penicillin, 100 µg/ml streptomycin in
a humidified atmosphere of 5% CO2 and 95% air at 37°C
until VSMCs exhibited a typical 'hill and valley' growth pattern.
Finally, VSMCs were identified with the morphological examination,
and showed 99% purity as estimated with the immunocytochemical
staining for α-actin. The cells were observed using an optical
microscope (Eclipse E600; Nikon Corporation, Tokyo, Japan) and the
positive ratio of immunocytochemical staining for α-actin was
analyzed with Image-Pro Plus 5.0 software (Media Cybernetics Inc.,
Rockville, MD, USA). The cells were passaged by brief
trypsinization (MP Biomedicals, Santa Ana, CA, USA), and the cells
between passages 3 and 8 were used for the experiments. When the
cells were grown to confluence, they were starved for 24 h in
serum-free medium prior to the experiments. All experimental
procedures were performed in accordance with the international,
national and institutional rules, and approved by the Institutional
Animal Care Committee of Xi'an Jiaotong University (Xi'an,
China).
Experimental protocols
VSMCs were cultured in 6-well plates at a density of
5×106 cells/well. When cells had been starved for 24 h
in the serum-free medium, the cells were divided into the following
groups: Control, cells treated with fibrinogen or FDP, and cells
treated with fibrinogen or FDP with pravastatin (10, 30 or 100
µmol/l). Following pretreatment with pravastatin for 30 min,
the cells were stimulated with 5 µmol/l fibrinogen for 9 h,
or 1 ml FDP diluted in 30 ml physiological saline (1/30 FDP) for 3
h, as previously described by Naito et al (17). Then, the supernatant was collected.
IL-6 and TNF-α levels were determined using an ELISA, and iNOS
protein expression was detected with immunofluorescence. mRNA
expression of IL-6, TNF-α and iNOS were determined by reverse
transcription-polymerase chain reaction (RT-PCR).
Preparation of FDP
About 6.3 units of human thrombin were mixed with 3
ml fibrinogen (50 µmol/l), and incubated overnight at 37°C.
The formed clots were homogenized for 10 sec, and centrifuged at
600 × g for 10 min. Then, the supernatant was removed, and the
clots were washed three times with physiological saline. Digestion
of the clots was conducted in Tris-buffered saline containing
physiological levels of calcium (10 mmol/l Tris, 100 mmol/l NaCl
and 2.5 mmol/l CaCl2, pH 7). The clotted fibrin was
suspended in 5 ml buffer, and 2 units of human plasmin were added
to form FDP. Subsequently. the samples were placed on a rotator in
a 37°C incubator for 48 h, and sterilized by passing the samples
through a 0.22-µm pore sterilization filter (17). Finally, the samples containing FDP
were diluted with physiological saline. FDP concentrations were
expressed as the diluted proportion.
ELISA
VSMCs were cultured in 6-well plates at a density of
5×106 cells/well. The cells were stimulated with
fibrinogen (5 µmol/l) or FDP (1/30) for the indicated time
after pretreatment for 30 min with pravastatin at 10, 30 or 100
µmol/l. Then, the supernatant was collected for assay of
IL-6 and TNF-α by ELISA kits specific for rat IL-6 and TNF-α
according to the manufacturer's instructions.
RT-PCR
Total RNA was extracted and complementary DNA (cDNA)
was synthesized from 1 µg of total RNA with random hexamer
primers using Revert Aid™ First Strand cDNA Synthesis kit according
to the manufacturer's instructions. PCR amplification was performed
through 35 cycles at 94°C for 30 sec, 54°C for 30 sec, and 72°C for
30 sec, and the final extension of PCR products was performed for 5
min at 72°C. Primers for rat IL-6, TNF-α, iNOS and
glyceraldehyde-3-phosphate dehydrogenase (GAPDH) were designed with
Beacon designer 4.0 (Palo Alto, CA, USA) (Table I). GAPDH was used as an internal
control. The samples were run in triplicate. Equal volumes of the
reaction mixture from each sample were loaded on a 2% TAE agarose
gel (Spanish Biochemicals Corp., Madrid, Spain) containing ethidium
bromide, and resolved by electrophoresis. Images were digitally
captured using a gel analysis system (JS-680D; Shanghai Peiqing
Science & Technology Co., Ltd., Shanghai, China), and band
intensity was analyzed using Gel Pro Analyzer software, Version 4.0
(Media Cybernetics., Inc., Rockville, MD, USA). The relative amount
of each mRNA was normalized to the housekeeping gene (GADPH)
mRNA.
 | Table IPrimer sequence used for RT-PCR
analysis. |
Table I
Primer sequence used for RT-PCR
analysis.
| Gene | Primer sequence | Accession number |
|---|
| IL-6 |
5′-TCCAGCCAGTTGCCTTCTTG-3′ | NM _012589 |
|
5′-AGCCACTCCTTCTGTGACTC-3′ | |
| TNF-α |
5′-CCAGAACTCCAGGCGGTGTC-3′ | NM_012675 |
|
5′-GGCTACGGGCTTGTCACTCG-3′ | |
| iNOS |
5′-GGAAGAGACGCACAGGCAGAG-3′ | NM_012611 |
|
5′-GCAGGCACACGCAATGATGG-3′ | |
| GAPDH |
5′-GCCTTCTCCATGGTGGTGAA-3′ | NM_017008 |
|
5′-GGTCGGTGTGAACGGATTTG-3′ | |
Immunofluorescent analysis of iNOS
VSMCs were plated on cover slips in 6-well plates
until 90% confluency. Then, the treated cells were fixed with 4%
formaldehyde in phosphate-buffered saline (PBS) for 15 min. The
cell membrane was permeabilized with 0.2% Triton X-100-PBS
(Amresco, LLC, Solon, OH, USA) for 20 min, and the non-specific
binding sites were blocked with 10% goat serum. Following
pretreatment with rabbit anti-rat iNOS antibody (1:200 diluted in
PBS) overnight at 4°C, the cells were washed three times with
phosphate-buffered saline, and incubated with the secondary
antibodies conjugated to fluorescein isothiocyanate for 15 min at
room temperature. Finally, the immunolabeled cells were observed
under a fluorescent microscope (BX-51 Olympus, Tokyo, Japan), and
fluorescent intensity of iNOS was detected and analyzed with
Image-Pro Plus software (Media Cybernetics., Inc., Rockville, MD,
USA). Data were expressed as relative to the control.
Statistical analysis
Data are expressed as the mean ± standard deviation.
The experiments were repeated three times, and the differences
between means were examined for statistical significance
(P<0.05) by one-way analysis of variance followed by Fisher's
exact test. SPSS 13.0 (SPSS, Inc., Chicago, IL, USA) was used for
all statistical analyses.
Results
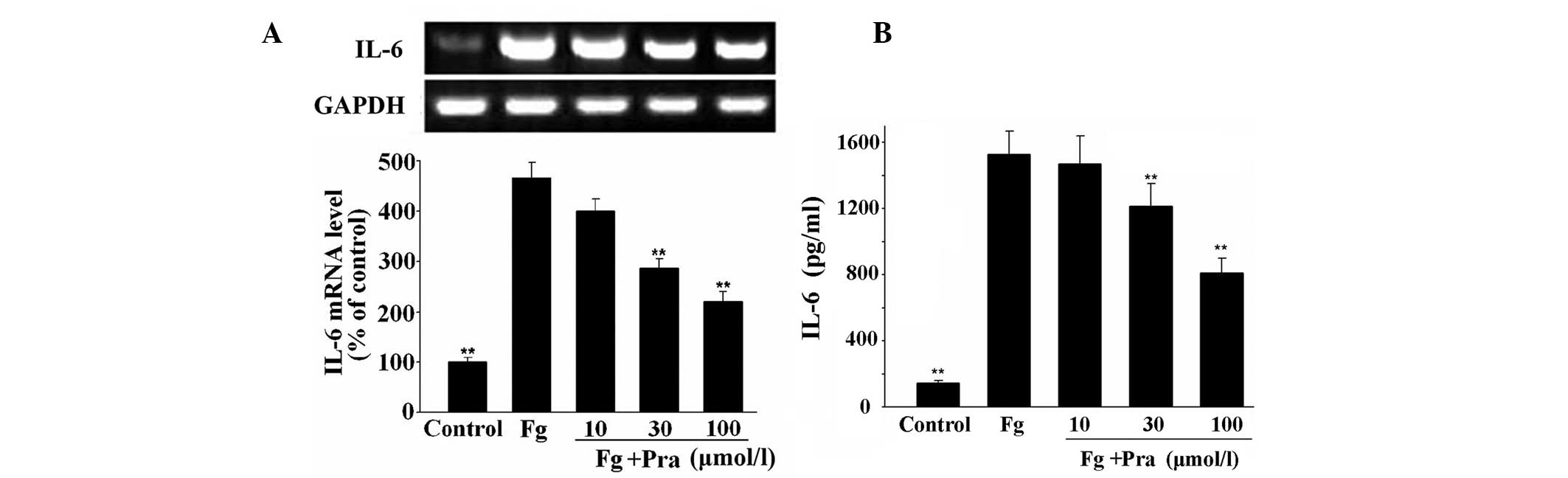
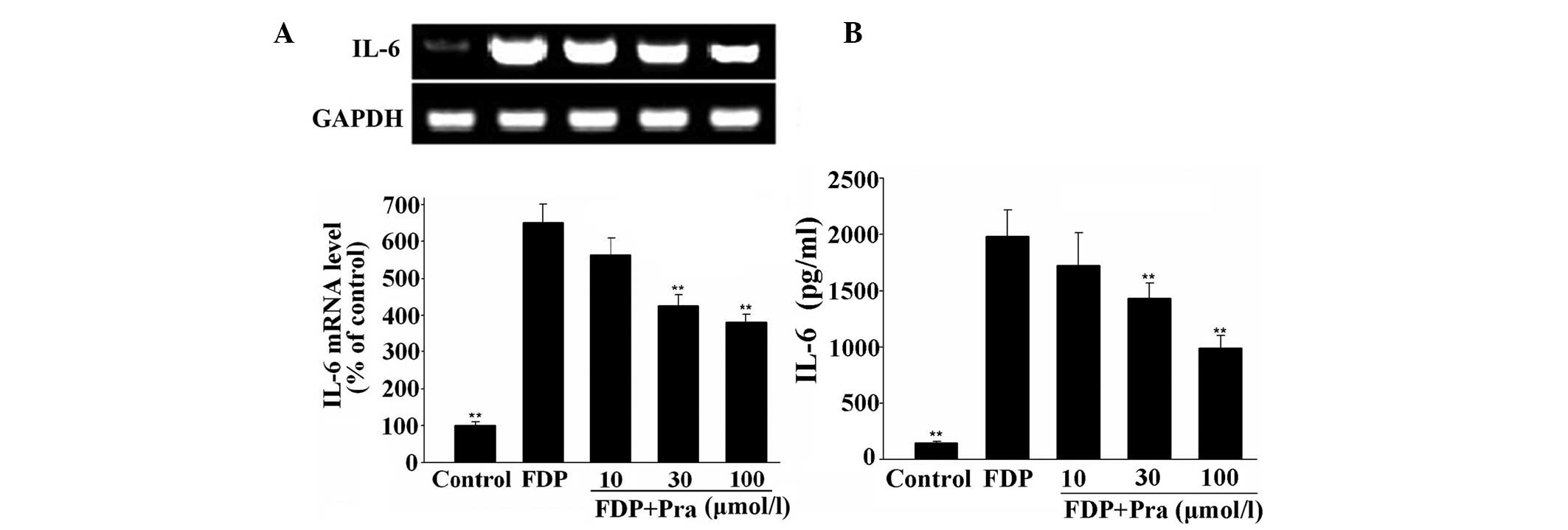
Pravastatin inhibits fibrinogen- and
FDP-induced IL-6 expression in VSMCs
As shown in Figs. 1
and 2, mRNA and protein expression
of IL-6 was low in the unstimulated VSMCs. However, mRNA and
protein expression of IL-6 was significantly increased following
stimulation of VSMCs with fibrinogen or FDP (P<0.01 vs.
control). However, pravastatin dose-dependently suppressed
fibrinogen- and FDP-stimulated IL-6 expression in VSMCs in protein
and mRNA levels (P<0.01 vs. fibrinogen or FDP alone). The
inhibitory rates in protein levels were 3.7, 20.7 and 46.9% in
fibrinogen-stimulated VSMCs, and were 13.1, 27.9 and 50.2% in
FDP-stimulated VSMCs with the increase of pravastatin
concentration.
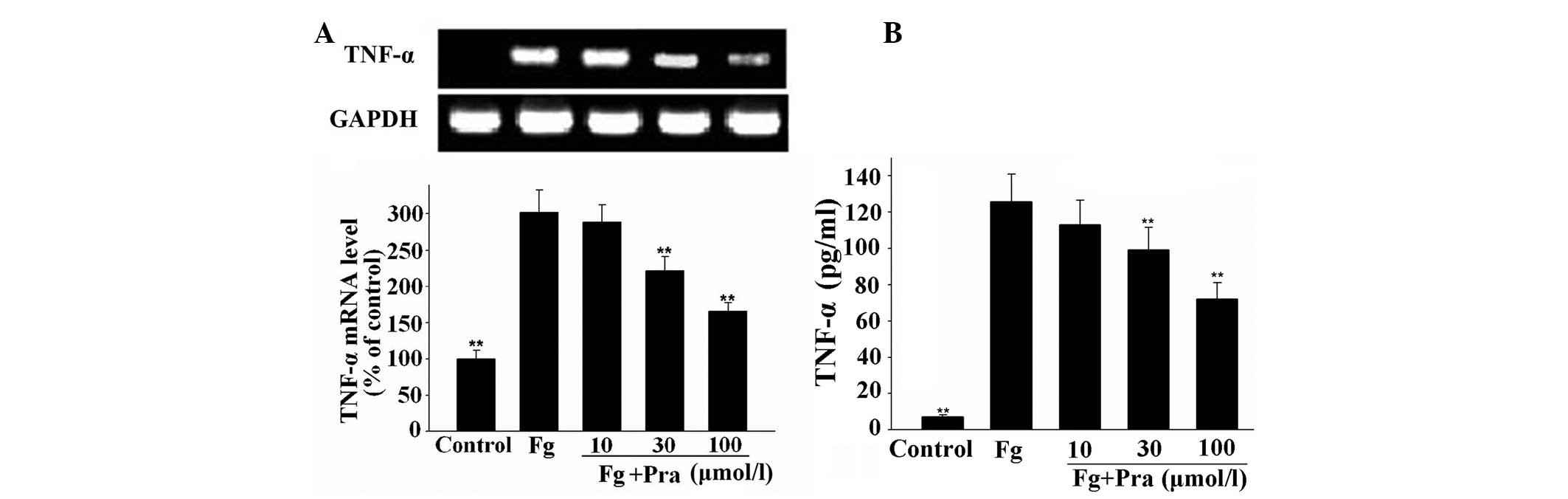
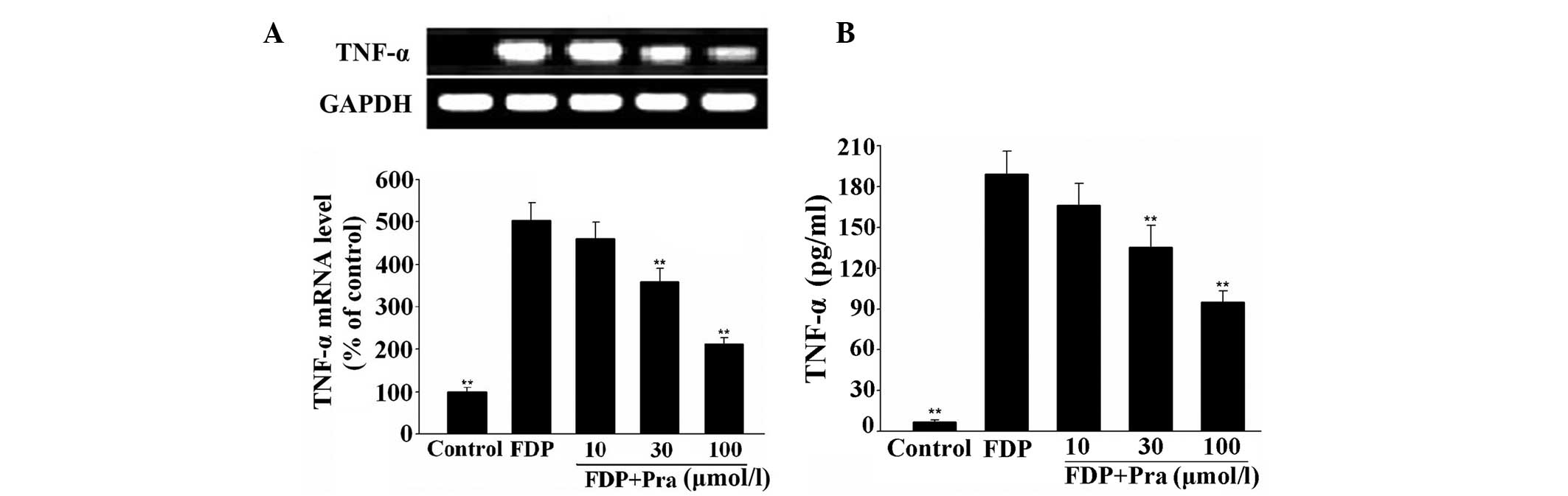
Pravastatin attenuates fibrinogen- and
FDP-induced TNF-α expression in VSMCs
As observed in Figs.
3 and 4, fibrinogen and FDP
produced a significant increase in the mRNA and protein expression
of TNF-α in VSMCs (P<0.01 vs. control), while pretreatment of
the cells with pravastatin inhibited fibrinogen- and FDP-induced
mRNA and protein expression of TNF-α in a concentration-dependent
manner (P<0.01 vs. fibrinogen or FDP alone). The maximal
inhibition of fibrinogen- and FDP-induced TNF-α protein expression
was 42.7 and 49.8%, respectively.
Pravastatin reduces fibrinogen- and
FDP-stimulated iNOS expression in VSMCs
The results shown in Figs. 5 and 6 indicate that mRNA and protein
expression of iNOS in VSMCs was significantly enhanced after
exposure of the cells to the same concentration of fibrinogen and
FDP (P<0.05 vs. control). However, pretreatment of the cells
with pravastatin antagonized the stimulatory effect of fibrinogen
and FDP on mRNA and protein expression of iNOS in a dose-dependent
manner (P<0.05 or P<0.01 vs. fibrinogen or FDP alone). The
maximal inhibition reached 49.2% for fibrinogen-induced iNOS
protein expression and 53.6% for FDP-induced iNOS protein
expression.
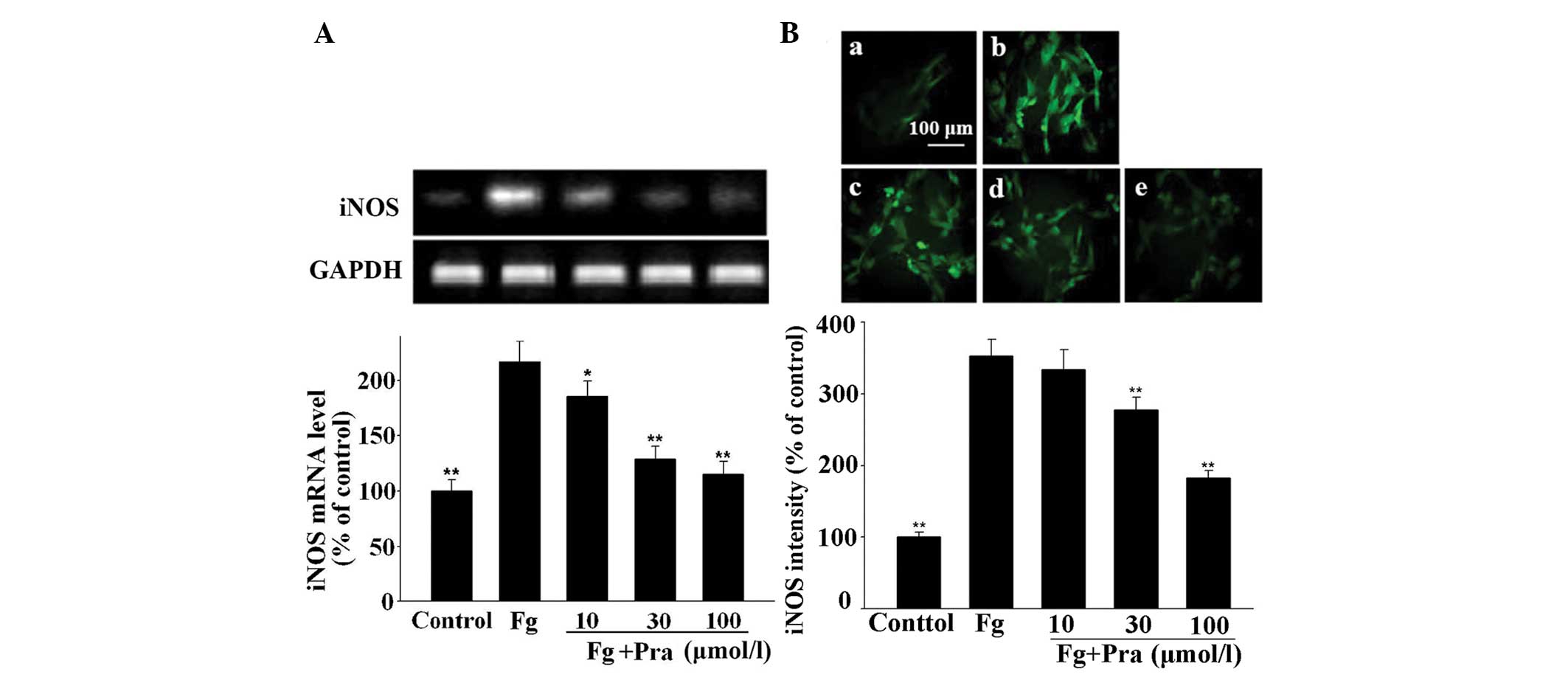 | Figure 5Effect of Pra on Fg-induced expression
of iNOS in VSMCs. VSMCs were stimulated with 5 µmol/l Fg for
9 h after pretreatment for 30 min with different concentrations of
Pra. Then, mRNA and protein expression of iNOS was determined by
reverse transcription-polymerase chain reaction and
immunofluorescence, respectively. (A) mRNA expression of iNOS.
Protein expression of iNOS in (Ba) control, (Bb) Fg alone, (Bc)
Fg+10 µmol/l Pra, (Bd) Fg+30 µmol/l Pra and (Be)
Fg+100 µmol/l Pra. Values from three independent experiments
are expressed as the mean ± standard deviation.
*P<0.05 and **P<0.01 vs. Fg. Scale bar,
100 µm. Pra, pravastatin; Fg, fibrinogen; iNOS, inducible
nitric oxide synthase; VSMCs, vascular smooth muscle cells. |
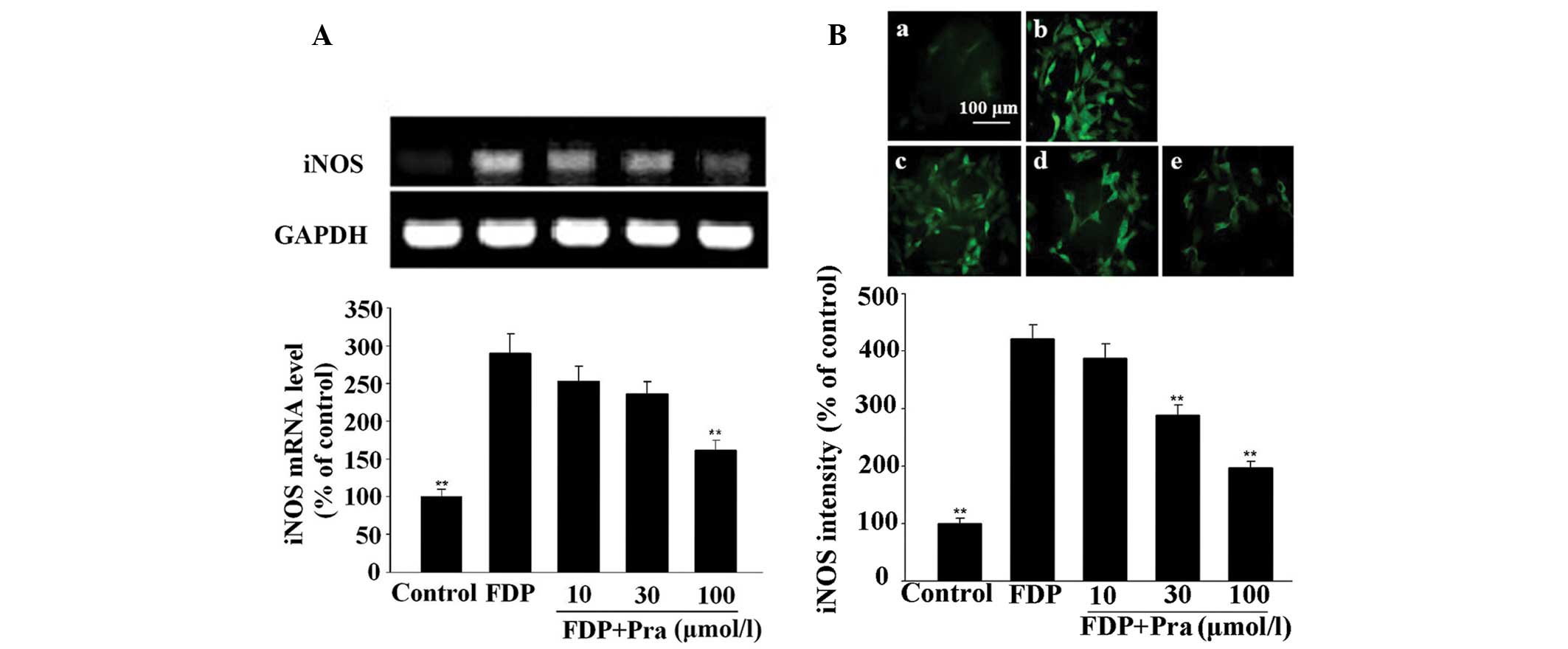 | Figure 6Effect of Pra on FDP-induced
expression of iNOS in VSMCs. VSMCs were stimulated with 1/30 FDP
for 3 h after pretreatment for 30 min with the different
concentrations of Pra. Then, mRNA and protein expression of iNOS
was determined by reverse transcription-polymerase chain reaction
and immunofluorescence, respectively. (A) iNOS mRNA expression.
iNOS protein expression in (Ba) control, (Bb) FDP alone, (Bc)
FDP+10 µmol/l Pra, (Bd) FDP+30 µmol/l Pra and (Be)
FDP+100 µmol/l Pra. Values from three independent
experiments are expressed as the means ± standard deviation.
**P<0.01 vs. FDP. Scale bar, 100 µm. Pra,
pravastatin; FDP, fibrin degradation products; iNOS, inducible
nitric oxide synthase; VSMCs, vascular smooth muscle cells. |
Discussion
Atherosclerosis is considered to be a chronic
inflammatory disease that leads to a number of cardiovascular
diseases (CVDs). Statins have been demonstrated to decrease
cardiovascular events through their pleiotropic properties,
including anti-inflammatory action (24). However, the precise mechanisms by
which statins inhibit inflammatory responses remain unclear.
VSMCs are the major players implicated in
atherogenesis. Migration and proliferation of VSMCs are the
critical events in the initiation and progression of
atherosclerosis as well as in the inflammatory processes (6,2).
IL-6 and TNF-α are the important pro-inflammatory cytokines
involved in atherogenesis, which stimulate the release of other
inflammatory mediators, increase expression of adhesion molecules
in endothelial cells, promote release of various chemokines to
recruit monocytes to the injury site and enhance their adhesiveness
to endothelium (25,26). A clinical study suggests that high
serum levels of IL-6 and TNF-α is associated with a worse prognosis
in patients after acute myocardial infarction (27).
It is known that iNOS exists in human
atherosclerotic lesions as several factors present in
atherosclerotic plaques, such as TNF-α, IL-1 and γ-interferon, are
able to induce iNOS expression in VSMCs. High iNOS activity
produces large quantities of NO, which may lead to cellular damage,
inflammation, apoptosis and peroxynitrite formation. Furthermore,
peroxynitrite may cause oxidative damage to endothelial cells and
VSMCs, and therefore is important in the pathology of
atherosclerosis (28–30).
Although the role of fibrinogen in the clotting
cascade has been well defined, its role in the inflammatory
response is less well understood. It has long been demonstrated
that fibrinogen and its fragments are incorporated into
atherosclerotic plaques as the plaques develop (31). Fibrinogen is also converted into a
fibrin matrix at the site of inflammation, and fibrin(ogen)
deposition may directly participate in the inflammatory response by
providing a scaffold for inflammatory cell adhesion and migration,
and for subsequent remodeling of the tissue with extracellular
matrix (32). Our previous studies
found that fibrinogen and FDP are able to produce a
pro-inflammatory effect on VSMCs via stimulating the generation of
CRP, IL-6, TNF-α and iNOS, which suggests a novel mechanism
involving fibrinogen in atherogenesis (19,20).
In view of the pro-inflammatory effect of fibrinogen
on VSMCs, which is involved in its pro-atherosclerotic process, the
inhibitory effect of pravastatin on fibrinogen- and FDP-stimulated
generation of inflammatory cytokines, such as IL-6, TNF-α and iNOS,
was observed in VSMCs. The results showed that pravastatin at the
concentrations used in the present study significantly inhibited
fibrinogen- and FDP-stimulated expression of IL-6, TNF-α and iNOS
in VSMCs at the mRNA and protein leves, suggesting that pravastatin
is able to exert an anti-inflammatory effect via decreasing
fibrinogen- and FDP-induced expression of IL-6, TNF-α and iNOS in
VSMCs.
Our previous study demonstrated that lovastatin
reduces nuclear factor-κB activation induced by CRP in human
vascular endothelial cells (33),
and Han et al in turn demonstrated that simvastatin inhibits
angiotensin II-induced CRP expression in human aortic endothelial
cells through interfering with AT1-reactive oxygen
species-mitogen activated protein kinase (MAPK) signaling pathway
(unpublished data). The MAPK signal pathway mediates the
inflammatory response induced by inflammatory cytokines in the
atherogenesis-related cells (34–37).
Thus, it was inferred that pravastatin, analogous to lovastatin and
simvastatin, reduces fibrinogen- and FDP-stimulated generation of
IL-6, TNF-α and iNOS in VSMCs possibly by inhibiting the MAPK
signaling pathway. Further studies are required to characterize the
mechanisms responsible for the inhibitory effect of
pravastatin.
In conclusion, the present results demonstrate that
pravastatin is able to inhibit fibrinogen- and FDP-stimulated
generation of IL-6, TNF-α and iNOS in VSMCs thus relieving the
inflammatory response in the vessel wall involved in atherogenesis.
This may aid in further explaining the beneficial effects of
pravastatin on atherosclerosis and CVDs. It also suggests that
application of pravastatin may be beneficial for the prevention of
atherosclerosis in hyperfibrinogenemia.
Acknowledgments
This study was supported by the Doctoral Fund of the
Ministry of Education of China (grant no. 20100201110053).
References
|
1
|
Hansson GK: Inflammation, atherosclerosis
and coronary artery disease. N Engl J Med. 352:1685–1695. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Libby P: Inflammation in atherosclerosis.
Nature. 420:868–874. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bermudez EA, Rifai N, Buring J, Manson JE
and Ridker PM: Interrelationships among circulating interleukin-6,
C-reactive protein and traditional cardiovascular risk factors in
women. Arterioscler Thromb Vasc Biol. 22:1668–1673. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lind L: Circulating markers of
inflammation and atherosclerosis. Atherosclerosis. 169:203–214.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Huber SA, Sakkinen P, Conze D, Hardin N
and Tracy R: Interleukin-6 exacerbates early atherosclerosis in
mice. Arterioscler Thromb Vasc Biol. 19:2364–2367. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ross R: Atherosclerosisan inflammatory
disease. N Engl J Med. 340:115–126. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lominadze D, Dean WL, Tyagi SC and Roberts
AM: Mechanisms of fibrinogen-induced microvascular dysfunction
during cardiovascular disease. Acta Physiol (Oxf). 198:1–13. 2010.
View Article : Google Scholar
|
|
8
|
Sen U, Tyagi N, Patibandla PK, Dean WL,
Tyagi SC, Roberts AM and Lominadze D: Fibrinogen-induced
endothelin-1 production from endothelial cells. Am J Physiol Cell
Physiol. 296:C840–C847. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Szaba FM and Smiley ST: Roles for thrombin
and fibrin(ogen) in cytokine/chemokine production and macrophage
adhesion in vivo. Blood. 99:1053–1059. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Robson SC, Shephard EG and Kirsch RE:
Fibrin degradation product D-dimer induces the synthesis and
release of biologically active IL-lbeta, IL-6 and plasminogen
activator inhibitors from monocytes in vitro. Br J Haematol.
86:322–326. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kuhns DB, Nelson EL, Alvord WG and Gallin
JI: Fibrinogen induces IL-8 synthesis in human neutrophils
stimulated with formyl-methionyl-leucyl-phenylalanine or
leukotriene B(4). J Immunol. 167:2869–2878. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tsakadze NL, Zhao Z and D'Souza SE:
Interaction of inter-cellular adhesion molecule-1 with fibrinogen.
Trends Cardiovasc Med. 12:101–108. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mosesson MW: Fibrinogen and fibrin
structure and functions. J Thromb Haemost. 3:1894–1904. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tyagi N, Roberts AM, Dean WL, Tyagi SC and
Lominadze D: Fibrinogen induces endothelial cell permeability. Mol
Cell Biochem. 307:13–22. 2008. View Article : Google Scholar
|
|
15
|
Patibandla PK, Tyagi N, Dean WL, Tyagi SC,
Roberts AM and Lominadze D: Fibrinogen induces alterations of
endothelial cell tight junction proteins. J Cell Physiol.
221:195–203. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Rauch BH, Müschenborn B, Braun M, Weber AA
and Schrör K: ICAM-1 and p38 MAPK mediate fibrinogen-induced
migration of human vascular smooth muscle cells. Eur J Pharmacol.
577:54–57. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Naito M, Stirk CM, Smith EB and Thompson
WD: Smooth muscle cell outgrowth stimulated by fibrin degradation
products: The potential role of fibrin fragment E in restenosis and
atherogenesis. Thromb Res. 98:165–174. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hicks RC, Golledge J, Mir-Hasseine R and
Powell JT: Vasoactive effects of fibrinogen on saphenous vein.
Nature. 379:818–820. 1996. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Guo F, Liu JT, Wang CJ, Liu N and Lu PP:
Fibrinogen, fibrin and FDP induce C-reactive protein generation in
rat vascular smooth muscle cells: Pro-inflammatory effect on
atherosclerosis. Biochem Biophy Res Commun. 390:942–946. 2009.
View Article : Google Scholar
|
|
20
|
Lu PP, Liu JT, Liu N, Guo F, Ji YY and
Pang X: Pro-inflammatory effect of fibrinogen and FDP on vascular
smooth muscle cells by IL-6, TNF-α and iNOS. Life Sci. 88:839–845.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Veillard NR and Mach F: Statins: The new
aspirin? Cell Mol Life Sci. 59:1771–1786. 2002. View Article : Google Scholar
|
|
22
|
Gaugler MH, Vereycken-Holler V, Squiban C,
Vandamme M, Vozenin-Brotons MC and Benderitter M: Pravastatin
limits endothelial activation after irradiation and decreases the
resulting inflammatory and thrombotic responses. Radiat Res.
163:479–487. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hadrava V, Tremblay J and Hamet P:
Abnormalities in growth characteristics of aortic smooth muscle
cells in spontaneously hypertensive rats. Hypertension. 13:589–597.
1989. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Takemoto M and Liao JK: Pleiotropic
effects of 3-hydroxy-3-meth-ylglutaryl coenzyme a reductase
inhibitors. Arterioscler Thromb Vasc Biol. 21:1712–1719. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tedgui A and Mallat Z: Cytokines in
atherosclewrosis: Pathogenic and regulatory pathways. Physiol Rev.
86:515–581. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yudkin JS, Kumari M, Humphries SE and
Mohamed-Ali V: Inflammation, obesity, stress and coronary heart
disease: Is interleukin-6 the link? Atherosclerosis. 148:209–214.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kosmala W, Przewlocka-Kosmala M and
Mazurek W: Proinflammatory cytokines and myocardial viability in
patients after acute myocardial infarction. Int J Cardiol.
101:449–456. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Buttery LD, Springall DR, Chester AH,
Evans TJ, Standfield EN, Parums DV, Yacoub MH and Polak JM:
Inducible nitric oxide synthase is present within human
atherosclerotic lesions and promotes the formation and activity of
peroxynitrite. Lab Invest. 75:77–85. 1996.PubMed/NCBI
|
|
29
|
Luoma JS, Strålin P, Marklund SL, Hiltunen
TP, Särkioja T and Ylä-Herttuala S: Expression of extracellular SOD
and iNOS in macrophages and smooth muscle cells in human and rabbit
atherosclerotic lesions: Colocalization with epitopes
characteristic of oxidized LDL and peroxynitrite-modified proteins.
Arterioscler Thromb Vasc Biol. 18:157–167. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Radomski MW and Salas E: Nitric
oxide-biological mediator, modulator and factor of injury: its role
in the pathogenesis of atherosclerosis. Atherosclerosis.
118(Suppl): S69–S80. 1995. View Article : Google Scholar
|
|
31
|
Smith EB, Thompson WD, Crosbie L and Stirk
CM: Fibrinogen/fibrin in atherogenesis. Eur J Epidemiol. 8(Suppl
1): 83–87. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Perez RL and Roman J: Fibrin enhances the
expression of IL-lbeta by human peripheral blood mononuclear cells.
Implications in pulmonary inflammation. J Immunol. 154:1879–1887.
1995.PubMed/NCBI
|
|
33
|
Lin R, Liu J, Peng N, Yang G, Gan W and
Wang W: Lovastatin reduces nuclear factor kappaB activation induced
by C-reactive protein in human vascular endothelial cells. Biol
Pharm Bull. 28:1630–1634. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Peng N, Liu JT, Gao DF, Lin R and Li R:
Angiotensin II-induced C-reactive protein generation: Inflammatory
role of vascular smooth muscle cells in atherosclerosis.
Atherosclerosis. 193:292–298. 2007. View Article : Google Scholar
|
|
35
|
Wang C, Liu J, Guo F, Ji Y and Liu N:
Endothelin-1 induces the expression of C-reactive protein in rat
vascular smooth muscle cells. Biochem Biophys Res Commun.
389:537–542. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Han C, Liu J, Liu X and Li M: Angiotensin
II induces C-reactive protein expression through ERK1/2 and JNK
signaling in human aortic endothelial cells. Atherosclerosis.
212:206–212. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Li M, Liu J, Han C, Wang B, Pang X and Mao
J: Angiotensin II induces the expression of c-reactive protein via
MAPK-dependent signal pathway in U937 macrophages. Cell Physiol
Biochem. 27:63–70. 2011.PubMed/NCBI
|