Introduction
Colon cancer is a malignant tumor arising from the
inner wall of the large intestine. It is aggressive and progresses
rapidly (1,2). Previous epidemiological studies have
indicated that the incidence rate increases with age (3). Almost 25% of cases present with
metastases at the initial diagnosis and approximately half of
patients with colon cancer will develop metastases, contributing to
the high mortality rate of this disease (4). Furthermore, the majority of
chemotherapeutic/chemopreventive agents including imatinib,
doxorubicin and methotrexate have detrimental effects on patients'
health (5–7). Certain natural compounds from Chinese
medicinal herbs have been identified to be cytotoxic to cancer
cells whilst remaining non-toxic to normal cells, including
taxinol, taxanes, camptothecins, epipodophyllo, curcumin, genistein
and perillyl alcohol (8,9). Therefore, the evaluation and
development of the anticancer activity of medicinal herbs is
important for the improvement of anticancer agents. To date, the
majority of polysaccharides and polysaccharide-protein complexes
isolated from fungi have been reported to account for their
anticancer activities in clinical therapy for patients with cancer
(10,11).
G. lucidum is a fungi that has been used in
Asia as a health tonic to promote longevity for thousands of years
(12). Polysaccharides derived
from G. lucidum (GLPs) have a wide range of biological and
pharmacological properties, including anticarcinoma effects, and as
such have attracted attention in the biochemical and medical
fields. GLPs have been demonstrated to induce suppression of cell
growth and apoptosis in different types of cancer cells, with
minimal negative effects on normal cells (13). Further studies have demonstrated
that GLPs directly or indirectly mediate cancer cell death through
diverse mechanisms (14–16). GLPs act as potential
immunomodulators that indirectly inhibit cancer cells by increasing
the activity of host immune cells (14,17).
GLPs have been demonstrated to be able to directly induce cell
differentiation of leukemia cells by caspase cleavage and p53
activation (18). Additionally,
GLPs may suppress Hep2 cells via the regulation of hepatic miRNAs
and immune-associated miRNAs (15), and may reverse drug resistance by
inhibition of the expression of resistance genes in the SKOV-3/DDP
resistant ovarian cancer cell line (19). Therefore, polysaccharides from
G. lucidum may represent a safe and effective agent in
cancer therapy. In a previous study, combination treatment with
GLPs and 5-fluorouracil (5-FU) resulted in synergistic
cytotoxicity, apoptosis and cell cycle arrest in human colon cancer
cells (20). However, the detailed
apoptotic mechanism mediated by GLPs in human colon cancer cells
remains to be fully elucidated.
In the present study, the inhibitory effect of
high-molecular-weight GLPs (>10 kDa) on LoVo human colon cancer
cells and the possible associated molecular mechanisms were
investigated.
Materials and methods
Drugs and chemicals
Dulbecco's modified Eagle's medium (DMEM) high
glucose and fetal bovine serum (FBS) were obtained from Gibco Life
Technologies (Carlsbad, CA, USA). 5-FU (>99% purity) was
purchased from Shanghai Bangcheng Chemical Co., Ltd. (Shanghai,
China). MTT, the Lactate Dehydrogenase (LDH) Cytotoxicity Assay kit
and the Caspase-3, -8 and -9 Activity Assay kits were purchased
from Beyotime Institute of Biotechnology (Nantong, China). The
rabbit polyclonal primary antibodies against β-actin (1:4,000;
20536-1-AP), Fas (1:1,000; 13098-1-AP) and caspase-3 (1:500;
19677-1-AP) were purchased from Proteintech Group, Inc. (Chicago,
IL, USA) and the goat anti-rabbit horseradish peroxidase
(HRP)-labeled secondary antibody (1:3,000; sc-2004) was purchased
from Santa Cruz Biotechnology, Inc. (Dallas, TX, USA). All
chemicals were of analytical grade unless otherwise specified.
Isolation and characterization of
GLPs
Slices of G. lucidum (Leyss. ex. Fr.) Karst.
were provided by the State Key Laboratory of Sub-Health
Intervention Technology, State Administration of Traditional
Chinese Medicine (Changsha, China). Samples were ground to a fine
powder and refluxed in three volumes of methanol/chloroform solvent
(1:2, v/v; Sinopharm Chemical Reagent Co., Ltd., Shanghai, China)
and 80% ethanol three times (6 h/each time). The residues were
dried, suspended in ~15 vol distilled water and were concentrated
with vacuum rotary evaporator (EYELA Auto Jack NAJ-100; Tokyo
Rikakikai Co., Ltd., Tokyo, Japan) vacuum filtration three times
for 6 h each time at 95°C. The combined concentrated solution was
ethanol precipitated and then deproteinized using a Sevage assay
(21). The extracts were purified
using DEAE-cellulose (DE 52; Sigma-Aldrich, St. Louis, MO, USA),
dialyzed with 3000 MWCO regenerated cellulose dialysis tubing
(Spectrum Laboratories, Inc., Rancho Dominguez, CA, USA),
concentrated and freeze-dried. The crude polysaccharides extraction
yield was calculated by dividing the final weight of crude extract
by the initial weight of the samples. Purified GLPs (10 mg/ml) were
ultrafiltered using Biomax-500, Biomax-300, Ultracel PL-100 and
Ultracel PL-30 membranes (EMD Millipore, Billerica, MA, USA) with a
molecular weight cut-off of 50, 30, 10 and 3 kDa, and the five
freeze-dried fractions were weighed for analysis of molecular
weight composition. Three fractions of polysaccharides with high
molecular weight (>10 kDa) were pooled and further analyzed.
Polysaccharide content was calculated by the phenol-sulfuric acid
method (22). Protein and nucleic
acid were determined by UV spectrum analysis (UV-1800; Shimadzu
Corporation, Kyoto, Japan), and uronic acid was determined by the
3,5-dimethylphenol method (23).
The infrared spectra were recorded with a Fourier
Transform-Infrared Spectrometer (Thermo Fisher Scientific, Madison,
WI, USA) in the range of 400–4,000 cm−1. GLPs were
hydrolyzed with 1 mol/l H2SO4 (Sinopharm
Chemical Reagent Co., Ltd.) for 8 h at 100°C, and cooled prior to
centrifugation at 2,400 x g for 30 min at room temperature using
centrifugal filters (Amicon Ultra-4; EMD Millipore). The GLPs were
then dried in the rotary evaporator, then monosaccharide components
were analyzed by high-performance anion exchange chromatography on
a Dionex ICS-3000 Ion Chromatograph System (Thermo Fisher
Scientific).
Cell culture
The human LoVo colon carcinoma cell line was
obtained from the Institute of Basic Medical Sciences (Beijing,
China). Cells were maintained in DMEM high glucose supplemented
with 10% FBS in an incubator at 37°C and 5% CO2.
Cell viability assay
LoVo cells at a density of 3×104
cells/well were cultured in 96-well plates. Following culture for
24 h, cells were incubated with fresh medium containing various
concentrations of GLPs (0.313, 0.625, 1.25, 2.5, 5 and 10 mg/ml) at
the predetermined time intervals (24, 48 and 72 h). A total of 50
µg/ml 5-FU was used as a positive control, while untreated
cells served as a negative control. Subsequently, 20 µl MTT
solution (5 mg/ml in phosphate-buffered saline, PBS; pH 7.4;
Sinopharm Chemical Reagent Co., Ltd.) was added, followed by
incubation for 4 h in the dark. Subsequent to the removal of the
media, purple formazan crystals were dissolved in 150 µl
dimethyl sulfoxide (Beijing Solarbio Science & Technology Co.,
Ltd., Beijing, China). The absorbance at 492 nm was measured using
a microplate reader (MK3; Thermo Fisher Scientific, Waltham, MA,
USA).
Scratch-wound assay
Following the formation of a cell mono-layer in the
24-well plate, lines were scratched in the layer using 10 µl
pipette tips. Following the removal of non-adherent cells by PBS,
adherent cells were incubated with various concentrations of GLPs
(0, 0.625, 1.25, 2.5, 5 and 10 mg/ml) for 48 h at 37°C (50
µg/ml 5-FU as a positive control). Three different views at
each concentration were imaged using an inverted microscope (IMT-2;
Olympus, Tokyo, Japan) and analyzed using ImageJ software, version
1.42 (National Institutes of Health, Bethesda, MD, USA).
Electron microscopy
The GLP-treated cell suspension was fixed with 2.5%
glutaraldehyde (Sigma-Aldrich) for 2 h, followed by fixation with
1% osmium tetroxide (OsO4; Sigma-Aldrich) for a further
2 h. Cells were rinsed in PBS three times prior to dehydration in
an ascending ethanol series (50, 70, 80, 90, 95 and 100%).
Following infiltration in 50% Quetol 651 (Nisshin EM, Tokyo, Japan)
for 1 h and 100% Quetol 651 for 6 h, cells were polymerized at 60°C
for 39 h. Samples were subsequently sectioned into 100 nm-thick
sections with a Leica UC6 micro-tome (Leica Microsystems, Inc.,
Buffalo Grove, IL, USA) and dyed with 4% uranyl acetate
(Sigma-Aldrich) for 10 min. Transmission electron microscopy (TEM)
was performed using a JEOL JEM-1230 (JEOL, Ltd., Tokyo, Japan)
microscope. For scanning electron microscopy (SEM; JSM-6380LV;
JEOL, Ltd.), LoVo cells were exposed to different concentrations of
GLPs in six-well plates for 48 h. Subsequently, cells were fixed in
1% glutaraldehyde and 1% OsO4, dehydrated in an ethanol
series (50–100%, v/v) and dried by critical-point drying with the
JSM-6380LV microscope. Following gold sputtering also with the
JSM-6380LV microscope, digital images were obtained using the
scanning electron microscope.
Measurement of DNA fragmentation
Cells were incubated with GLPs (2.5, 5 and 10 mg/ml)
or 5-FU (50 µg/ml) for 24 h, followed by resuspension in 70%
ethanol for 4 h at −20°C. Following centrifugation at 1,000 x g for
5 min at 4°C, the precipitate was dissolved in 40 µl
phosphate-citric acid buffer (pH 7.8; 91.5 parts 0.5 M
Na2HPO4 and 8.5 parts of 0.23 M
NaH2PO4) and continuously incubated at room
temperature for 45 min with intermittent agitation. The supernatant
was mixed with 3 µl 1 mg/ml RNaseA and 3 µl 0.25%
NP40 (Beijing Solarbio Science & Technology Co., Ltd.) at 37°C
for 30 min. DNA fragmentation was analyzed by gel electrophoresis
on a 1.2% (w/v) agarose gel (Sigma-Aldrich). The gel was visualized
by staining with ethidium bromide (Amresco LLC, Solon, OH, USA) and
the images were captured using the UVP GDS-8000 Bioimaging System
(UVP, Inc. Upland, CA, USA).
LDH release assay
Following culturing for 48 h with or without
different concentrations of GLPs (0, 1.25, 2.5 and 5 mg/ml), the
supernatant was collected and added to a new 96-well plate (200
µl/well). Subsequently, LDH release was calculated using a
LDH Cytotoxicity Assay kit according to the manufacturer's
instructions. The plates were analyzed using the MK3 micro-plate
reader at a wavelength of 450 nm.
Measurement of caspase-3, -8 and -9
activity
The activity of caspases-3, -8 and -9 was quantified
using the Caspase-3, -8 and -9 Activity kits according to the
manufacturer's instructions. Briefly, cells were treated for 24 h
with GLPs (0, 2.5 and 5 mg/ml). Following production of the pellet
by centrifugation at 600 x g for 5 min at 4°C, cells were washed
with PBS two times and subsequently lysed using lysis reagent
(Beyotime Institute of Biotechnology) in an ice bath for 15 min.
The supernatants were incubated with 80 µl reaction buffer
(1% NP-40, 20 mM Tris-HCl, pH 7.5, 137 mM NaCl and 10% glycerol;
Beyotime Institute of Biotechnology) and 10 µl substrate at
37°C for 2 h. At the end of the incubation, the absorbance of
cleavage (pNA) was measured with the MK3 microplate reader at 405
nm.
Western blot analysis
GLP-treated cells were collected, washed with
ice-cold PBS and lysed in radioimmunoprecipitation assay buffer
(Applygen Technologies, Inc., Beijing, China) for 30 min at 4°C.
Protein concentration was determined using the Bradford assay
(Bio-Rad Laboratories, Inc., Hercules, CA, USA). Total protein was
subjected to electrophoresis on 12% SDS-PAGE (Beyotime Institute of
Biotechnology) and transferred onto polyvinylidene fluoride
membranes (EMD Millipore). Subsequently, the membranes were
incubated with rabbit anti-Fas (1:1,000), rabbit anti-caspase-3
(1:500) and mouse anti-β-actin (1:4,000) at 4°C overnight.
Membranes were then incubated with HRP-labeled secondary antibodies
(1:3,000) at room temperature for 1.5 h. The signal was detected
using enhanced chemiluminescence reagents (Thermo Fisher
Scientific) for western blot analysis. The relative abundance of
bands was quantified using ImageJ software.
Statistical analysis
All data are presented as the mean ± standard
deviation. Statistical analyses were performed with Student's
t-test using SPSS software, version 18.0 (SPSS, Inc., Chicago, IL,
USA). P<0.01 was considered to indicate a statistically
significant difference.
Results
Preparation of GLPs
The yield of crude polysaccharides was ~3.08%. The
molecular weight composition of polysaccharides >10 kDa was:
10–30 kDa, 32.1%; 30–50 kDa, 21.8%; and >50 kDa, 46.1%. Using UV
scanning, the absence of nucleic acids and proteins in the GLPs was
indicated by minimal absorption at 260 and 280 nm. GLPs were
comprised of 89% carbohydrate and 11% uronic acid. The peaks of
infrared spectroscopy at 847 and 922 cm−1 indicated that
the purified GLPs predominantly consisted of α-polysaccharides
(24). The results of the
monosaccharide composition analysis were in agreement and indicated
that GLPs were composed of arabinose, galactose, glucose and
cellose at the molar ratios of 11:3:3:1. GLPs were prepared as a
stock of 10 mg/ml in DMEM high glucose containing 10% FBS and
stored at −20°C.
Cytotoxic effects of GLPs on LoVo
cells
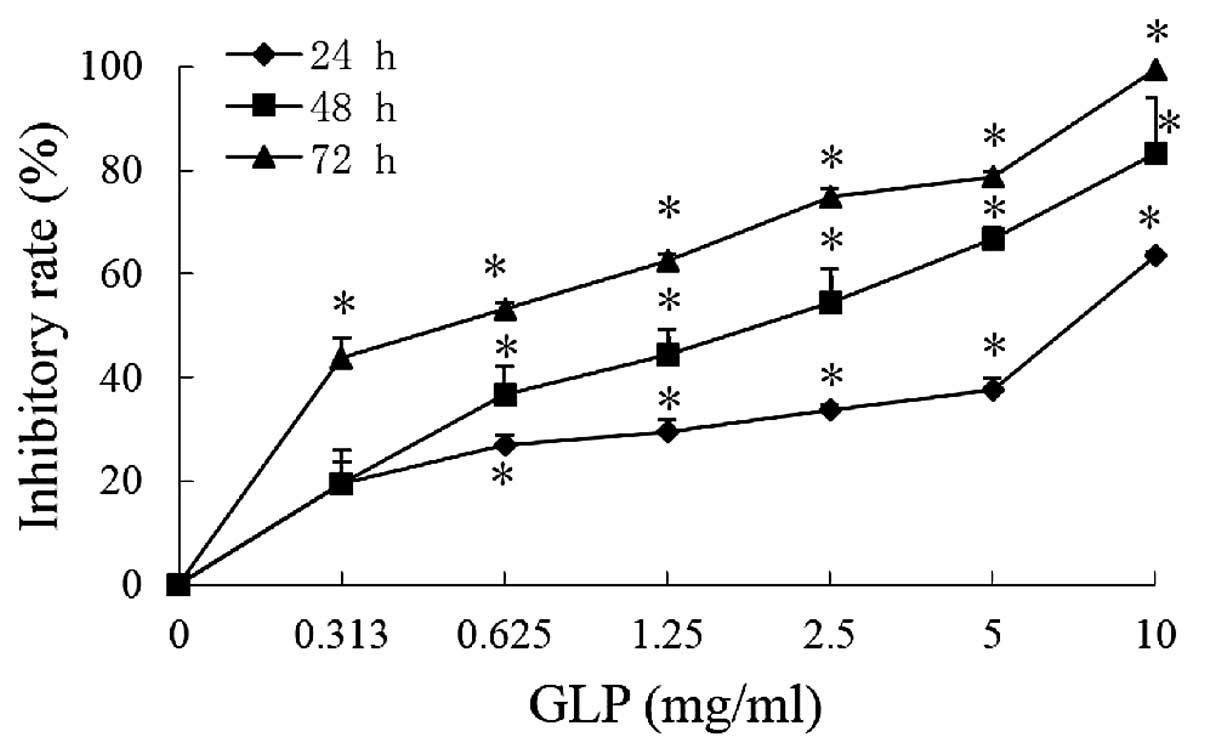
As presented in Fig.
1, the inhibitory rates of GLPs (0.313, 0.625, 1.25, 2.5, 5 and
10 mg/ml) at 72 h were 43.6, 53.0, 62.5, 74.9, 78.6 and 99.3%,
respectively, compared with the control. The half maximal
inhibitory concentration (IC50) values at 24, 48 and 72
h were 6.94, 1.67 and 0.63 mg/ml, respectively. The results
demonstrate that GLPs significantly reduced cell viability in a
dose- and time-dependent manner (P<0.01), suggesting that GLPs
possess a potent cytotoxicity against human colon cancer cells.
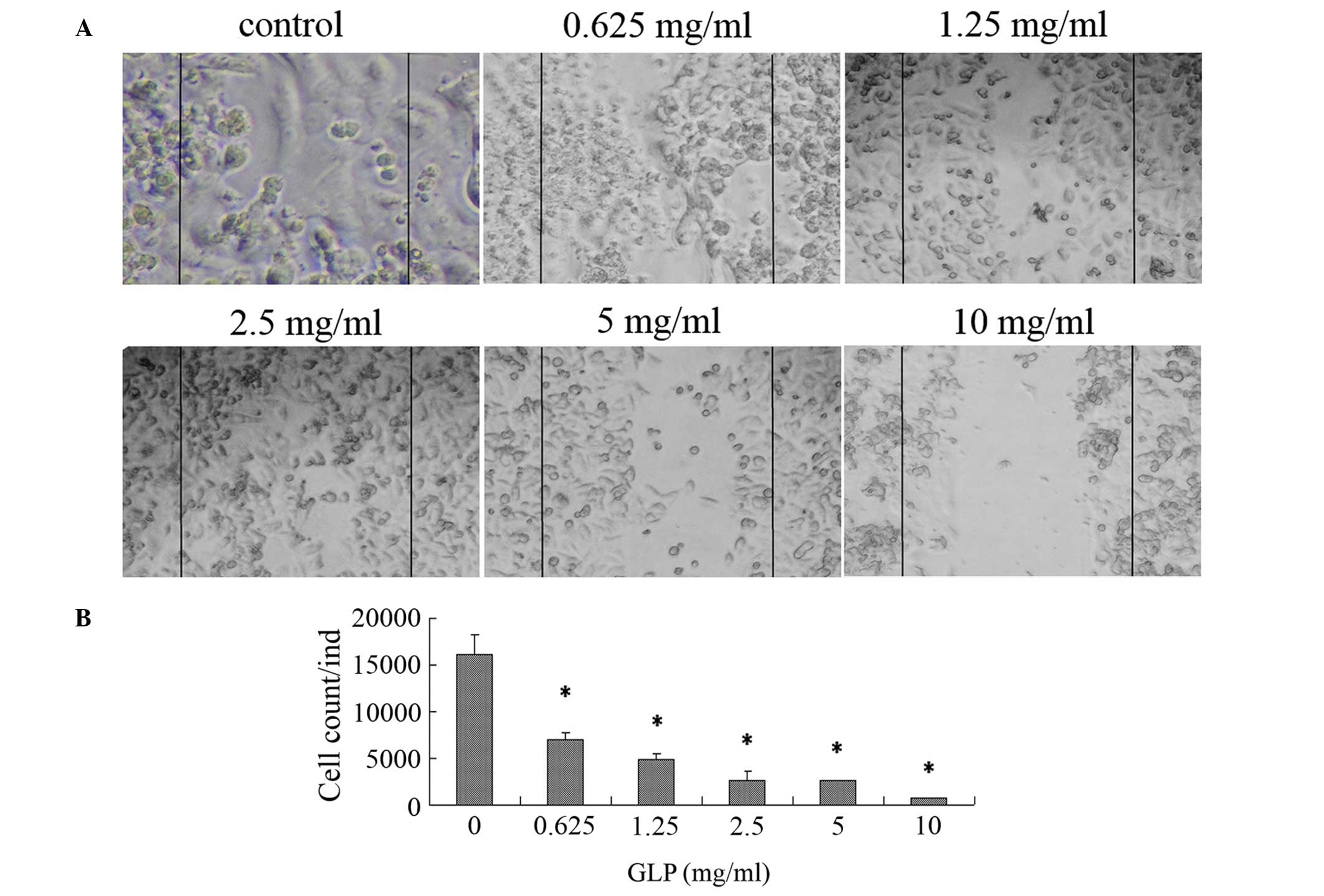
GLPs inhibit cell migration in LoVo
cells
At 48 h, the scratch wounds were completely repaired
by the migration of untreated cells, whilst the wounds were
maintained in GLP-incubated cells as indicated by examination using
light microscopy (Fig. 2A). In
particular, LoVo cells treated with GLP concentrations of 5 and 10
mg/ml did not migrate towards the wound area compared with the
untreated cells. As presented in Fig.
2B, exposure to GLPs (0.625–10 mg/ml) resulted in a reduction
in the number of migratory cells from 95.9–56.9% (P<0.01),
indicating that GLPs are able to dose-dependently suppress cell
migration in LoVo cells.
GLP-mediated morphological alterations in
LoVo cells
To investigate the cell death induced by GLPs,
morphological characteristics of cells were observed by SEM and
TEM. SEM observations demonstrated that untreated cells exhibited a
uniform distribution and numerous microvilli on the cell surface
(Fig. 3A–a and A–d). In
GLP-treated cells, morphological alterations were observed
including low cellular density (Fig.
3A–b and A–c), a reduction in the number or disappearance of
microvilli and the formation of apoptotic bodies (Fig. 3A–e and A–f). In the TEM study,
untreated cells displayed microvilli on the cell surface and a
dispersed chromatin pattern (Fig.
3B–a). Following incubation with 5 mg/ml GLPs, the induction of
cell death was indicated by cell shrinkage, microvilli loss, cell
membrane blebbing (arrow in Fig.
3B–b) and cytoplasm marginalization (Fig. 3B–b). Subsequently, the cells broke
up into membrane-bound apoptotic bodies (arrow in Fig. 3B–c). Following exposure to 10 mg/ml
GLPs, apoptotic bodies were degraded by lysosomal enzymes (Fig. 3B–d).
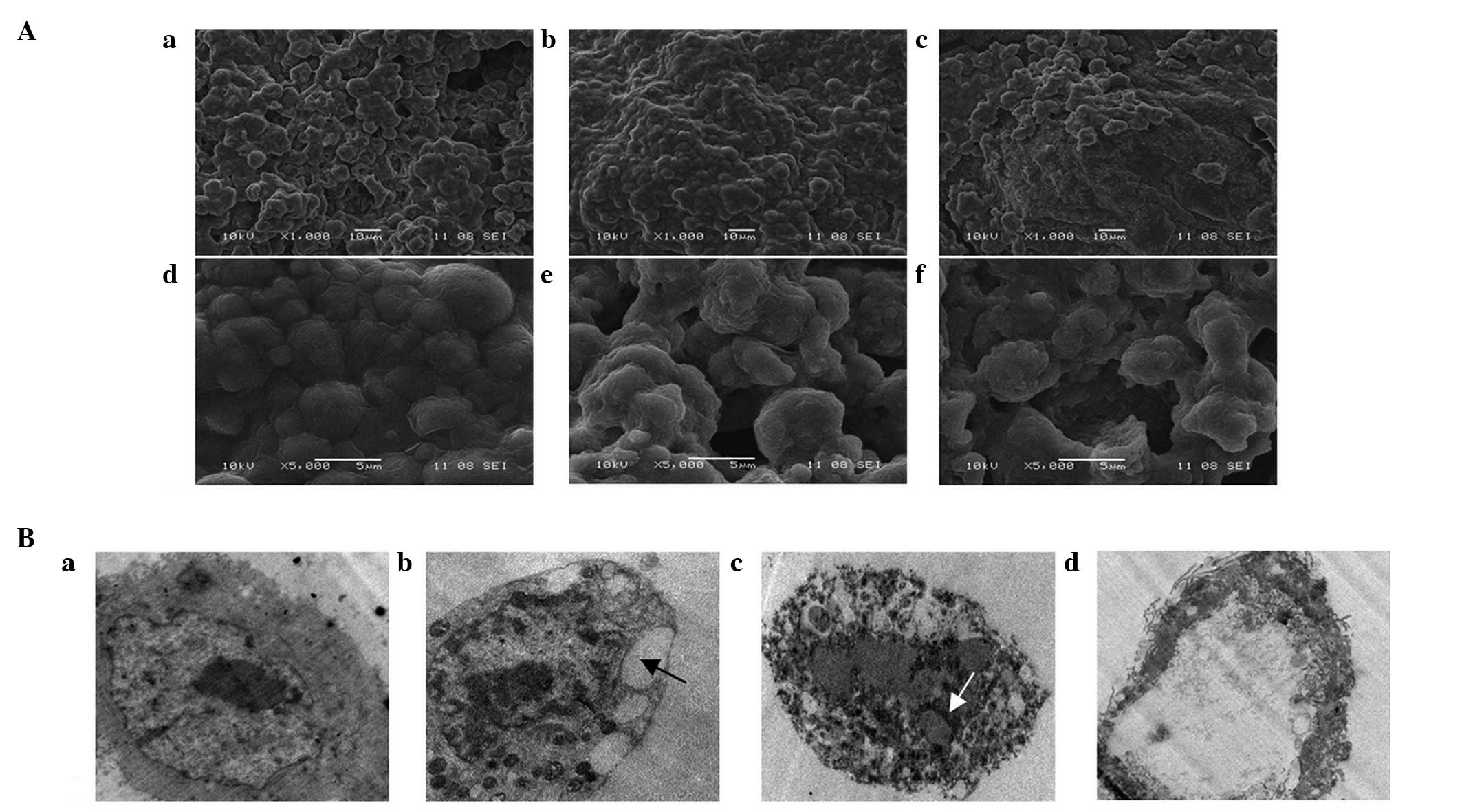 | Figure 3Effect of GLPs on the morphology of
LoVo cells. Cells were exposed to 0, 5 and 10 mg/ml GLP for 48 h.
(A) Appearance of LoVo cells under scanning electron microscopy.
(A-a-A-c) Magnification, ×1,000; (A-d-A-f) magnification, ×5,000.
(B) Morphological observation of LoVo cells by transmission
electron microscopy (magnification, ×10,000). Black arrows, empty
vacuole; white arrows, apoptotic bodies. GLP, G. lucidum
polysaccharide. |
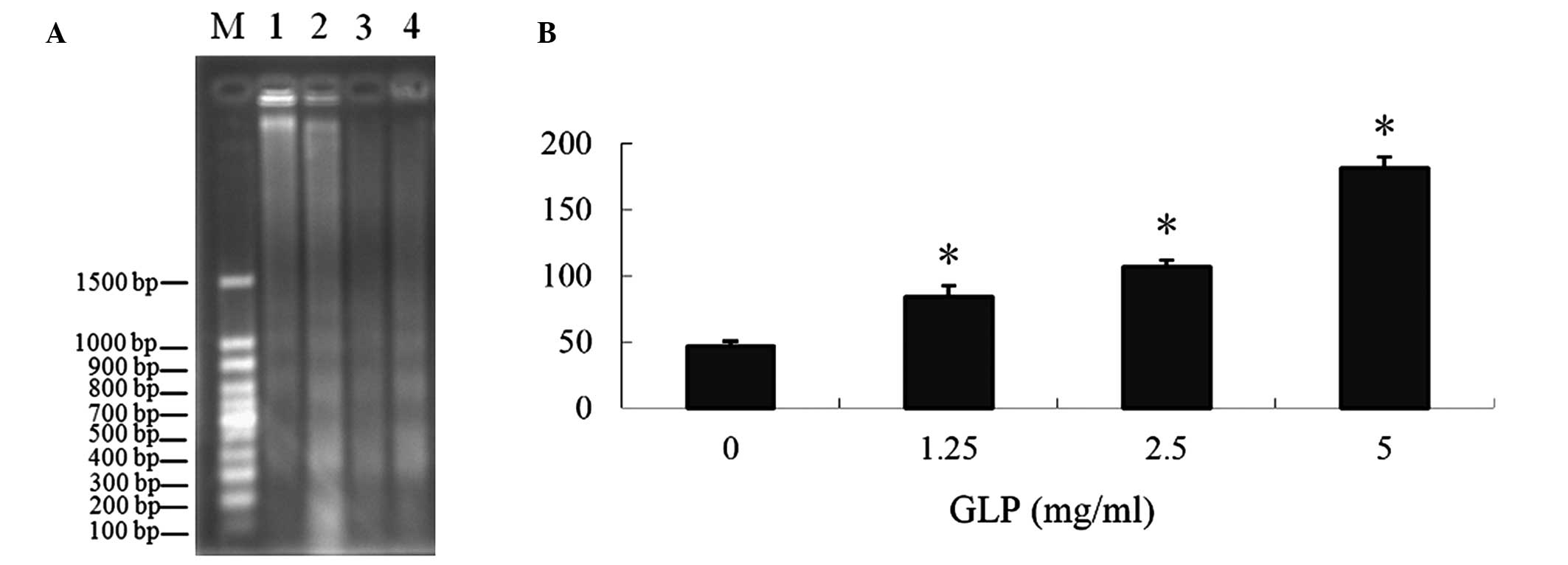
Induction of DNA fragmentation and LDH
release by GLPs in LoVo cells
DNA fragmentation and the level of LDH released into
the culture medium were used as indexes for evaluating apoptosis.
The results demonstrated that GLPs induced DNA fragmentation
following treatment with various doses of GLPs (1.25–10 mg/ml),
presented in Fig. 4A. The LDH
release assay demonstrated that GLP-induced LDH release was
significantly increased (P<0.01), and this effect was
dose-dependent (Fig. 4B), further
supporting the role of apoptosis in GLP-mediated cytotoxicity.
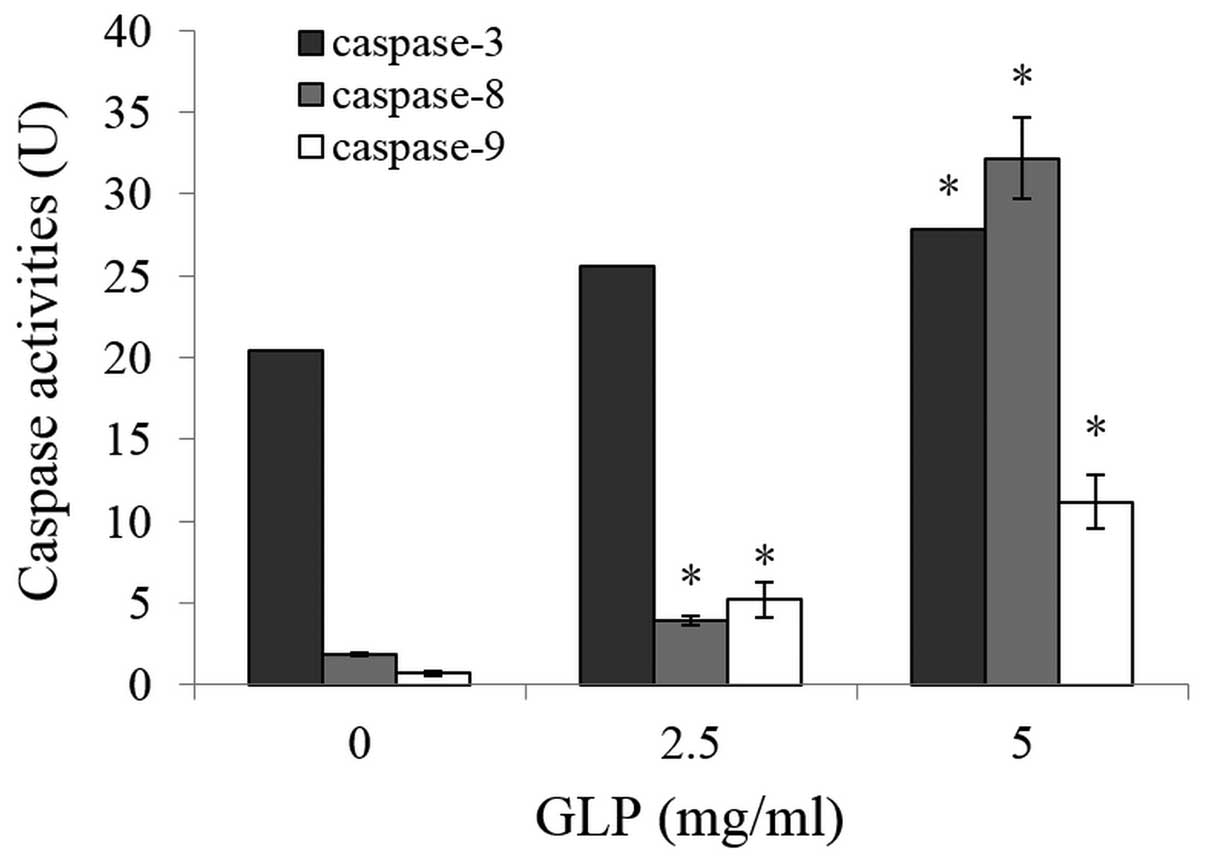
Analysis of GLP-induced apoptotic
pathways involved in LoVo cells
The addition of GLPs to cultured cells increased the
activity of caspases-3, -8 and -9 in a dose-dependent manner
(P<0.01), indicating GLP-mediated apoptosis occurred via the
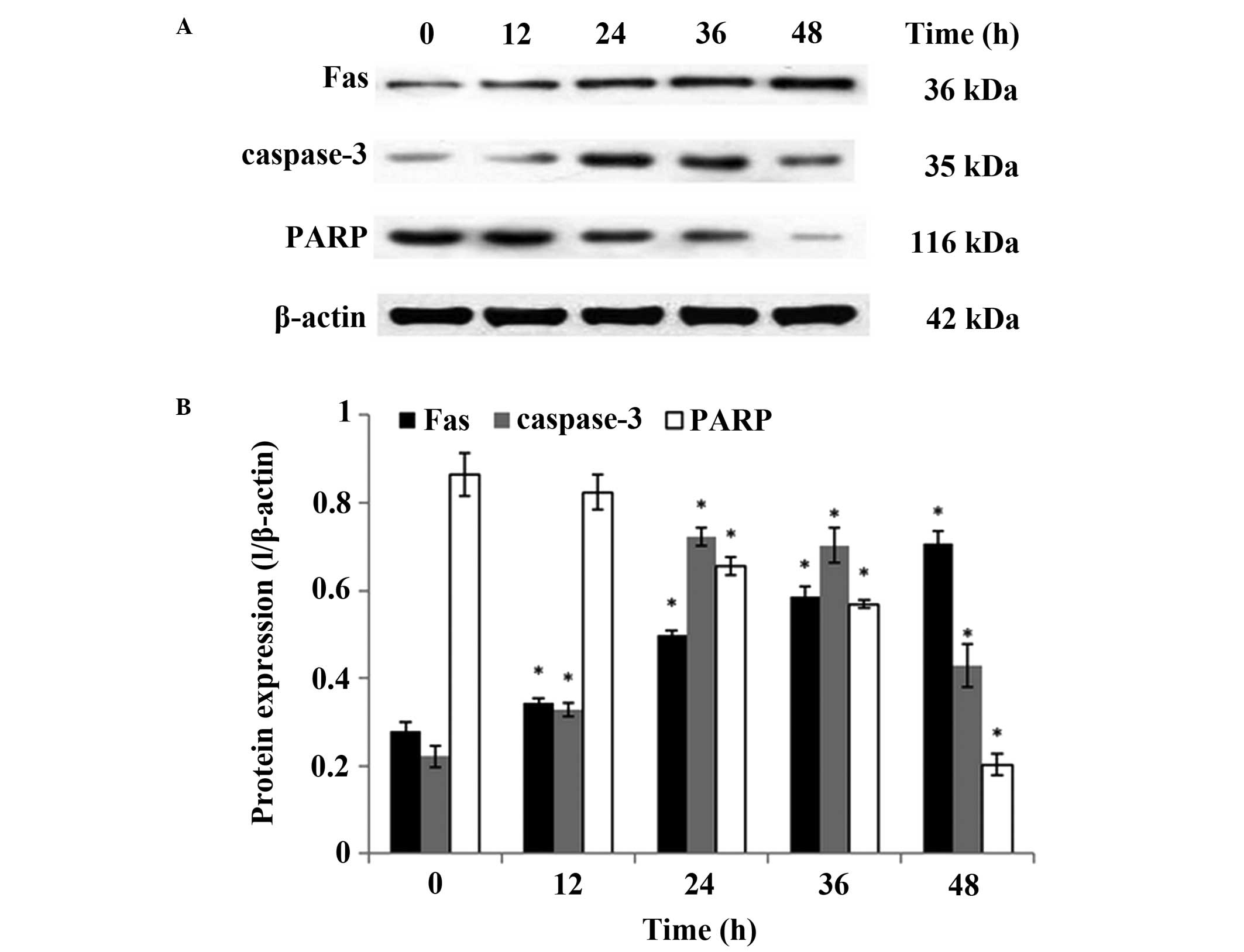
caspase-dependent pathway in LoVo cells (Fig. 5). Using western blotting, the
effect of GLPs on the expression of proteins associated with
apoptosis, including Fas, caspase-3 and poly(ADP-ribose) polymerase
(PARP) was further investigated. As presented in Fig. 6, the level of caspase-3 protein was
significantly increased compared with the control group at 12 h and
reached its maximum level at 24 h, which is in agreement with the
results of caspase activity assay. In addition, the levels of Fas
and PARP were increased and reduced, respectively, in a
dose-dependent manner (P<0.01). These results suggest a possible
association between GLP-induced apoptosis and the activation of the
Fas/caspase-dependent pathway.
Discussion
As important ingredients in G. lucidum, GLPs
have been demonstrated to reduce drug toxicity caused by
chemotherapy (25), whilst
additionally possessing antitumor activity without exhibiting
adverse effects. These effects have been reported to be mediated
through the activation of the immune response and the induction of
apoptosis in a range of cancer cells (26,27).
The anticancer activity of polysaccharides is known to be affected
by their chemical structure, chemical components, conformation,
molecular mass and solubility in water (28,29).
In general, GLPs with a backbone of β-(1,3)-D-glucan and a high branching ratio,
have been implicated to exhibit anticancer activity (30,31).
Additionally, high molecular weight GLPs exhibit greater
antineoplastic activity than low molecular weight GLPs (32). Previous studies have demonstrated
that GLPs, in addition to other chemical structures, possess
anticancer effects in vitro and in vivo (33–35).
GLPs comprising of 1,3-, 1,4- and 1,6-linked β-D-glucopyranosyl
have been demonstrated to enhance the proliferation of T- and
B-lymphocytes in vitro, and GLPs consisting of 1,4-linked
α-D-glucopyranosyl residues and 1,6-linked β-D-galactopyranosyl
residues exhibited immune-stimulating activity in mice (36). Pan et al (37) isolated GLPs with a molecular weight
of 78 kDa, which were composed of D-galactose, D-glucose and
L-rhamnose in a molar ratio of 1.00:3.22:1.15. In the current
study, GLPs (molecular weight >10 kDa) with α-glycosidic
linkages were obtained from G. lucidum. The purified GLPs
were predominantly composed of four kinds of monosaccharides,
arabinose, galactose, glucose and cellose, at a molar ratio of
11:3:3:1, which was similar to a previous study (38). The anticancer effect on human colon
cancer LoVo cells was investigated in the current study. In an MTT
assay, 0.313–10 mg/ml GLPs demonstrated a time- and
concentration-dependent inhibition of LoVo cells (Fig. 1), suggesting GLPs possess potential
cytotoxic properties against LoVo cancer cells.
Cell migration serves an important role in numerous
physiological processes, such as cell differentiation and
proliferation (39). Unregulated
cell migration is responsible for cancer formation, relapse and
metastasis, ultimately leading to development of malignant tumors
(40). Therefore, the inhibition
of this process is a potential therapeutic strategy for controlling
tumor growth. In the current study, significant suppression of
migration was observed in the scratch-wound assay in LoVo cells
following incubation with GLPs at doses of 0.625–10 mg/ml (Fig. 2). This demonstrated that GLPs
additionally act as an inducer for migration inhibition in
vitro.
Apoptosis serves a role in the suppression of tumor
progression and elimination of unwanted cells in various biological
systems, and serves a key role in cell death induced by anticancer
drugs (41). Apoptotic cells
exhibit characteristic morphological alterations including cell
shrinkage, membrane blebbing, chromatin cleavage, formation of
apoptotic bodies and nuclear condensation (42). In the current study, using electron
microscopy (TEM and SEM), the ultrastructural alterations that
indicate apoptosis were examined, including the appearance of
pyknotic nuclei (chromatin condensation and marginalization),
nuclear fragmentation and apoptotic body formation (Fig. 3). Following exposure of LoVo cells
to GLPs, DNA fragments were observed, exhibiting exhibiting clear
fragmentation by GLP treatment, particularly at 5 and 10 mg/ml GLP
(Fig. 4A). Additionally, GLPs
induced LDH release into the culture medium (Fig. 4B) (43). These observations indicate that
apoptosis occurred in the LoVo human cancer cells upon treatment
with GLPs, suggesting that GLPs may exert their anticancer effects
on LoVo cells through an apoptotic pathway.
The organized process of apoptosis is controlled by
a signaling network, which includes the following pathways:
Mitogen-activated protein kinase/extracellular signal-related
kinase, calpain/apoptosis inducing factor, mitochondrial and the
death receptor pathways (44). Fas
serves an important role in mediating apoptosis in the death
receptor pathway. It stimulates an apoptotic signal by binding to
Fas ligand (FasL) expressed on the surface of neighboring cells,
which in turn results in the recruitment of specialized adaptor
proteins and the activation of caspase cascades (45,46).
Increasing evidence suggests that activation of FasL/Fas may
mediate activation downstream of caspase-8, followed by the
activation of caspases-9 and -3 (47). Additionally, apoptosis is able to
occur by the caspase-8 independent FasL/Fas-mediated death receptor
pathway (48). Mitochondria
contain several potentially apoptogenic factors including
cytochrome c and pro-caspases-3 and -9, which are associated
with the mitochondrial apoptotic pathway (49,50).
Selenium-enriched GLP-induced apoptosis has been demonstrated to be
associated with a reduction in the mitochondrial membrane
potential, an increase in cytosolic levels of cytochrome c
and an increase in the activity of PARP, caspase-3 and caspase-9,
with no obvious activation of caspase-8 in human breast cancer
cells (27). In the current study,
to further understand the mechanisms of GLP-induced apoptosis in
human colon cancer LoVo cells, the activity of caspases and the
expression levels of proteins associated with apoptosis were
investigated. These data indicated that GLPs significantly
increased the activity of caspases-3, -8 and -9 in a time-dependent
manner (Fig. 5). Additionally,
GLP-induced apoptosis was observed to involve cleavage of PARP and
the upregulation of Fas and caspase-3 (Fig. 6). It is conceivable that GLPs may
exert their anticancer activity through the death receptor
signaling pathway in LoVo cells.
In conclusion, GLPs exhibited significant
cytotoxicity in LoVo cells in a time- and concentration-dependent
manner. The anticancer effects of GLPs were associated with the
induction of apoptosis and suppression of cell migration. In
addition, these data demonstrate that GLPs induced apoptosis via a
Fas/caspase-8 dependent pathway. The current study provides a new
perspective for further research on the toxicology and pharmacology
of GLPs as a potential candidate for the treatment or prevention of
colon cancer.
Acknowledgments
The authors would like to thank to Dr Zebin Huang
who supplied the human colon cancer cells and would like to thank
Professor Dongbo Liu and Professor Zhilan Xia for providing the
G. lucidum.
References
|
1
|
Tokunaga T, Oshika Y, Abe Y, Ozeki Y,
Sadahiro S, Kijima H, Tsuchida T, Yamazaki H, Ueyama Y, Tamaoki N,
et al: Vascular endothelial growth factor (VEGF) mRNA isoform
expression pattern is correlated with liver metastasis and poor
prognosis in colon cancer. Br J Cancer. 77:998–1002. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Resnick MB, Routhier J, Konkin T, Sabo E
and Pricolo VE: Epidermal growth factor receptor, c-MET,
beta-catenin, and p53 expression as prognostic indicators in stage
II colon cancer: A tissue microarray study. Clin Cancer Res.
10:3069–3075. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel R, Naishadham D and Jermal A:
Cancer Statistics, 2013. Ca Cancer J Clin. 63:11–30. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Van Cutsem E and Nordlinger B: Advanced
colorectal cancer: ESMO Clinical Practice Guidelines for treatment.
Ann Oncol. 21(Suppl 5): v93–v97. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Verweij J, Casali PG, Zalcberg J, LeCesne
A, Reichardt P, Blay JY, Issels R, van Oosterom A, Hogendoorn PC,
Van Glabbeke M, et al: Progression-free survival in
gastrointestinal stromal tumours with high-dose imatinib:
Randomised trial. Lancet. 36:1127–1134. 2004. View Article : Google Scholar
|
|
6
|
Gordon KB, Tajuddin A, Guitart J, Kuzel
TM, Eramo LR and Vonroenn J: Hand-foot syndrome associated with
liposome-encapsulated doxorubicin therapy. Cancer. 75:2169–2173.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Berkun Y, Levartovsky D, Rubinow A, Orbach
H, Aamar S, Grenader T, Abou Atta I, Mevorach D, Friedman G and
Ben-Yehuda A: Methotrexate related adverse effects in patients with
rheumatoid arthritis are associated with the A1298C polymorphism of
the MTHFR gene. Ann Rheum Dis. 63:1227–1231. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Venditto VJ and Simanek EE: Cancer
therapies utilizing the camptothecins: A review of the in vivo
literature. Mol Pharm. 7:307–349. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bao B, Ali S, Banerjee S, Wang Z, Logna F,
Azmi AS, Kong D, Ahmad A, Li Y, Padhye S, et al: Curcumin analogue
CDF inhibits pancreatic tumor growth by switching on suppressor
microRNAs and attenuating EZH2 expression. Cancer Res. 72:335–345.
2012. View Article : Google Scholar
|
|
10
|
Tan W, Lu J, Huang M, Li Y, Chen M, Wu G,
Gong J, Zhong Z, Xu Z, Dang Y, et al: Anti-cancer natural products
isolated from chinese medicinal herbs. Chin Med. 6:272011.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Harvey AL: Natural products in drug
discovery. Drug Discov Today. 13:894–901. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu GT: Recent advances in research of
pharmacology and clinical applications of Ganoderma P. Karst
species (Aphyllophoromycetideae) in China. Int J Med Mushrooms.
17:63–67. 2015.
|
|
13
|
Liu YJ, Shen J, Xia YM, Zhang J and Park
HS: The polysaccharides from Ganoderma lucidum: Are they always
inhibitors on human hepatocarcinoma cells? Carbohydr Polym.
90:1210–1215. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xu Z, Chen X, Zhong Z, Chen L and Wang Y:
Ganoderma lucidum polysaccharides: Immunomodulation and potential
anti-tumor activities. Am J Chin Med. 39:15–27. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shen J, Park HS, Xia YM, Kim GS and Cui
SW: The polysaccharides from fermented Ganoderma lucidum mycelia
induced miRNAs regulation in suppressed HepG2 cells. Carbohydr
Polym. 103:319–324. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li A, Shuai X, Jia Z, Li H, Liang X, Su D
and Guo W: Ganoderma lucidum polysaccharide extract inhibits
hepatocellular carcinoma growth by downregulating regulatory T
cells accumulation and function by inducing microRNA-125b. J Transl
Med. 13:1002015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Guo L, Xie J, Ruan Y, Zhou L, Zhu H, Yun
X, Jiang Y, Lü L, Chen K, Min Z, et al: Characterization and
immunostimulatory activity of a polysaccharide from the spores of
Ganoderma lucidum. Int Immunopharmacol. 9:1175–1182. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhu XL, Chen AF and Lin ZB: Ganoderma
lucidum polysaccharides enhance the function of immunological
effector cells in immunosuppressed mice. J Ethnopharmacol.
111:219–226. 2007. View Article : Google Scholar
|
|
19
|
Qu HG, Gao L, He D, et al: Effect of
reversion of Ganoderma lucidum polysaccharides on cisplatin
resistant in ovarian cancer cells and its mechanism. J Jilin Univ
(Medicine Edition). 37:251–255. 2011.
|
|
20
|
Liang Z, Yi YJ, Guo YT, Wang RC and Xiong
XY: Effect of combined Ganoderma lucidum polysaccharides and
flurouracil proliferation and apoptosis in human colon carcinoma
HCT-116 cells. Food Sci. 33:310–314. 2012.In Chinese.
|
|
21
|
Staub AM: Removal of proteins Sevage
method. Methods Carbohydr Chem. 5:5–6. 1965.
|
|
22
|
Kochert G: Carbohydrate determination by
the phenol-sulfuric acid method. Handbook of Phycological Methods,
Vol II, Physiological and Biochemical Methods. Hellebust JA and
Craigie JS: Cambridge University Press; Cambridge: pp. 95–97.
1978
|
|
23
|
Scott RW: Colorimetric determination of
hexuronic acids in plant materials. Anal Chem. 51:936–941. 1979.
View Article : Google Scholar
|
|
24
|
Li YQ, Fang L and Zhang KC: Structure and
bioactivities of a galactose rich extracellular polysaccharide from
submergedly cultured Ganoderma lucidum. Carbohydr Polym.
77:323–328. 2008.
|
|
25
|
Hu Z, Chen X, Yang X, Gao Y and Zhou S:
Water-soluble polysaccharides of Ganoderma lucidum (W. Curt:Fr) P
Karst (Aphyllophoromycetideae) alleviate the dose-limiting
toxicities of irinotecan (CPT-11). Int J Med Mushrooms. 8:321–328.
2006. View Article : Google Scholar
|
|
26
|
Zhou GQ, Zhao HY and Lu C: Effect of
Ganoderma lucidum polysaccharides on intestinal mucosal immune
system in H22 liver cancer bearing mice. Zhongguo Zhong Xi Yi Jie
He Za Zhi. 29:335–339. 2009.In Chinese. PubMed/NCBI
|
|
27
|
Shang D, Li Y, Wang C, Wang X, Yu Z and Fu
X: A novel polysaccharide from Se-enriched Ganoderma lucidum
induces apoptosis of human breast cancer cells. Oncol Rep.
25:267–272. 2011.
|
|
28
|
Ooi VE and Liu F: Immunomodulation and
anti-cancer activity of polysaccharide-protein complexes. Curr Med
Chem. 7:715–729. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xu W, Zhang F, Luo Y, Ma L, Kou X and
Huang K: Antioxidant activity of a water-soluble polysaccharide
purified from Pteridium aquilinum. Carbohydr Res. 344:217–222.
2009. View Article : Google Scholar
|
|
30
|
Miyazaki T and Nishijima M: Studies on
fungal polysaccharides. XXVII Structural examination of a
water-soluble, antitumor polysaccharide of Ganoderma lucidum. Chem
Pharm Bull (Tokyo). 29:3611–3616. 1981. View Article : Google Scholar
|
|
31
|
Mizuno T, Wang G, Zhang J, Kawagishi H,
Nishitoba T and Li J: Reishi, Ganoderma lucidum and Ganoderma
tsugae: Bioactive substances and medicinal effects. Food Rev Int.
11:151–166. 1995. View Article : Google Scholar
|
|
32
|
Zhang L, Zhang M, Zhou Q, Chen J and Zeng
F: Solution properties of antitumor sulfated derivative of
α-(1-->3)-D-glucan from Ganoderma lucidum. Biosci Biotechnol
Biochem. 64:2172–2178. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kimura Y, Taniguchi M and Baba K:
Antitumor and antimetastatic effects on liver of triterpenoid
fractions of Ganoderma lucidum: Mechanism of action and isolation
of an active substance. Anticancer Res. 22:3309–3318. 2002.
|
|
34
|
Tang W, Liu JW, Zhao WM, Wei DZ and Zhong
JJ: Ganoderic acid T from Ganoderma lucidum mycelia induces
mitochondria mediated apoptosis in lung cancer cells. Life Sci.
80:205–211. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Weng CJ and Yen GC: The in vitro and in
vivo experimental evidences disclose the chemopreventive effects of
Ganoderma lucidum on cancer invasion and metastasis. Clin Exp
Metastas. 27:361–369. 2010. View Article : Google Scholar
|
|
36
|
Bao XF, Wang XS, Dong Q, Fang JN and Li
XY: Structural features of immunologically active polysaccharides
from Ganoderma lucidum. Phytochemistry. 59:175–181. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Pan D, Wang L, Chen C, Teng B, Wang C, Xu
Z, Hu B and Zhou P: Structure characterization of a novel neutral
polysaccharide isolated from Ganoderma lucidum fruiting bodies.
Food Chem. 135:1097–1103. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chan WK, Cheung CC, Law HK, Lau YL and
Chan GC: Ganoderma lucidum polysaccharides can induce human
monocytic leukemia cells into dendritic cells with
immunostimulatory function. J Hematol Oncol. 1:92008. View Article : Google Scholar
|
|
39
|
Doyle AD, Petrie RJ, Kutys ML and Yamada
KM: Dimensions in cell migration. Curr Opin Cell Biol. 25:642–649.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Condeelis J and Pollard JW: Macrophages:
Obligate partners for tumor cell migration, invasion, and
metastasis. Cell. 124:263–266. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Surh YJ: Cancer chemoprevention with
dietary phytochemicals. Nat Rev Cancer. 3:768–780. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Doonan F and Cotter TG: Morphological
assessment of apoptosis. Methods. 44:200–204. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Sun B, Cai Y, Li Y, Li J, Liu K, Li Y and
Yang Y: The nonstructural protein NP1 of human bocavirus 1 induces
cell cycle arrest and apoptosis in Hela cells. Virology. 440:75–83.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Nguyen TTT, Tran E, Nguyen TH, Do PT,
Huynh TH and Huynh H: The role of activated MEK-ERK pathway in
quercetin-induced growth inhibition and apoptosis in A549 lung
cancer cells. Carcinogenesis. 25:647–659. 2004. View Article : Google Scholar
|
|
45
|
Chu K, Niu X and Williams LT: A
Fas-associated protein factor, FAF1, potentiates Fas-mediated
apoptosis. Proc Natl Acad Sci USA. 92:11894–11898. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Huang DC, Hahne M, Schroeter M, Frei K,
Fontana A, Villunger A, Newton K, Tschopp J and Strasser A:
Activation of Fas by FasL induces apoptosis by a mechanism that
cannot be blocked by Bcl-2 or Bcl-x(L). Proc Natl Acad Sci USA.
96:14871–14876. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Li H, Zhu H, Xu CJ and Yuan J: Cleavage of
BID by caspase 8 mediates the mitochondrial damage in the Fas
pathway of apoptosis. Cell. 94:491–501. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Xiong J, Cheng G, Tang H, Zhen HN and
Zhang X: Ardipusilloside I induces apoptosis in human glioblastoma
cells through a caspase-8-independent FasL/Fas-signaling pathway.
Environ Toxicol Pharmacol. 27:264–270. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Larrosa M, Tomás-Barberán FA and Espín JC:
The dietary hydrolysable tannin punicalagin releases ellagic acid
that induces apoptosis in human colon adenocarcinoma Caco-2 cells
by using the mitochondrial pathway. J Nutr Biochem. 17:611–625.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Fulda S and Debatin KM: Extrinsic versus
intrinsic apoptosis pathways in anticancer chemotherapy. Oncogene.
25:4798–4811. 2006. View Article : Google Scholar : PubMed/NCBI
|