Introduction
Acute lung injury (ALI) and its more severe form,
acute respiratory distress syndrome (ARDS), contribute to morbidity
and mortality in critically ill patients, with rates of morbidity
and mortality of 75/100,000 and 40–60%, respectively (1,2). In
particular, gram-negative sepsis often leads to ALI/ARDS (3). ALI/ARDS are characterized by an
extensive inflammatory process leading to diffuse alveolar damage,
an influx of neutrophils, activation of proinflammatory cytokines
and chemokines, macrophages and protein-rich exudate in the
alveolar space due to the disruption of the alveolar epithelial
barrier (1,2).
The active form of vitamin D, termed calcitriol or
1,25-dihydroxyvitamin D3
[1,25-(OH)2D3], binds to the vitamin D
receptor (VDR), a member of the nuclear hormone receptor
superfamily (4). In addition to
regulating calcium, vitamin D acts as a regulator of multiple
biological processes, including anti-inflammation, immunomodulation
and barrier function maintenance (5). Furthermore, it has been suggested
that the production of 1,25-(OH)2D3 is
evidence of a local paracrine/autocrine action in various tissues
(6), including pulmonary
epithelial cells (7). There is
growing evidence in support of associations between vitamin D
deficiency, and impaired pulmonary function (8), an increased incidence of ALI/ARDS
(9–12) and inflammatory diseases, including
asthma (13), tuberculosis
(14) and chronic obstructive
pulmonary disease (COPD) (8,15).
Previous studies have suggested that vitamin D deficiency is also
common in critically ill patients (9,10),
and is often associated with increased morbidity and mortality,
including that caused by ALI/ARDS (9–12,16,17).
However, the underlying mechanism of vitamin D/VDR signaling and
sepsis-induced ALI/ARDS has yet to be investigated.
Vitamin D/VDR signaling is important for the
integrity of tissue barriers and anti-inflammatory functions
(5,6). It has been demonstrated to regulate
the components of tight junctions and maintain the integrity of
epithelial barriers in multiple organs, including the skin
(18), eyes (19) and large intestine (20). In lungs, the permeability of the
alveolar epithelial barrier is largely regulated by the
intercellular junctions that seal the paracellular space (21,22).
Disruption of the epithelial barrier may increase alveolar
permeability and result in paracellular movements of fluid from the
interstitium to the pulmonary airspace, as well as infiltration of
inflammatory cells. Subsequent pulmonary edema impairs blood-gas
exchange, leading to ARDS. Preserving or restoring these barriers
may provide a therapeutic strategy for preventing or treating
ALI/ARDS (23,24). However, to the best of our
knowledge, there has been no previous investigation regarding
vitamin D/VDR signaling regulation of the alveolar epithelial
barrier in the lungs.
The aim of the present study was to investigate the
effect of VDR knockout on lipopolysaccharide (LPS)-induced ALI and
to assess the effect of vitamin D treatment on LPS-induced ALI in a
mouse model. The results suggested that mice lacking VDR had a
compromised alveolar epithelial barrier and aggravated ALI.
Furthermore, it was identified that vitamin D treatment may sustain
the integrity of the barrier and thus attenuate ALI.
Materials and methods
Ethics statement
All experimental procedures were reviewed and
approved by the Institutional Ethics Committee of China Medical
University (Shenyang, China).
Animals
VDR heterozygous (VDR+/−) mice with a
C57BL/6J background (6 weeks old) were purchased from The Jackson
Laboratory (Bar Harbor, ME, USA). VDR heterozygous males and
females were bred to generate wild-type (WT; VDR+/+) and
VDR knockout (KO; VDR−/−) mice for the experiment. All
mice were kept in pathogen-free cages with a 12-h light/dark cycle,
and fed a rescue diet high in calcium, phosphate and lactose
(Envigo; Madison, WI, USA) to maintain a normal plasma calcium
level in VDR KO mice (25).
Genotyping
Total DNA was harvested from the mouse tails and
primers were provided by The Jackson Laboratory. Polymerase chain
reaction (PCR) was performed as follows: Pre-denaturation at 94°C
for 5 min; followed by 35 cycles of 94°C for 30 sec, 55°C for 30
sec, 72°C for 40 sec, 72°C for 10 min, and held at 4°C until
electrophoresis. Agarose gel electrophoresis was performed with 2%
agarose (in TAE buffer; Beyotime Institute of Biotechnology) at 100
V until the bands were clearly separated.
Treatment groups
Two mice (age, 8–12 weeks) per gender were selected
for each treatment group. LPS [Lipopolysaccharides from
Escherichia coli 0111:B4, dissolved in phosphate-buffered
saline (PBS)] from Sigma-Aldrich (St. Louis, MO, USA) was injected
once intratracheally (10 mg/kg) in WT and KO mice. The control
group was comprised of KO and WT mice injected with the same volume
of sterile PBS. Bronchoalveolar lavage fluid (BALF) was collected.
Briefly, following anesthesia by intraperitoneal injection with a
cocktail of xylazine (Rompun 2%; Bayer AG, Leverkusen, Germany) and
ketamine (Ketavest; 100 mg/ml; Pfizer, Inc., New York, NY, USA),
the trachea was exposed and the lungs were lavaged three times with
0.2 ml sterile saline per wash. The BALF was stored at 4°C and
samples of lung tissue were harvested for RNA, protein and
histological studies 12 h after treatment, and refrigerated at
−80°C.
WT mice were randomly divided into four treatment
groups. Two vitamin D treatment groups (one PBS group and one LPS
group) were treated with the vitamin D analog paricalcitol
(Sigma-Aldrich) dissolved in propylene (glycol:ethanol, 90:10; 0.5
µg/kg body weight) through intra-peritoneal injection 30 min
prior to LPS or PBS treatment. The two vehicle groups received the
dissolvent only prior to LPS or PBS treatment.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was isolated from the lung tissues using
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). First-strand cDNAs were synthesized using a
PrimeScript RT reagent kit (Takara Biotechnology Co., Ltd., Dalian,
China) and PCR was performed with the 20-µl volume reaction
mixture using a SYBR-Green PCR reagent kit (Clontech Laboratories,
Inc., Mountainview, CA, USA) on a LightCycler 480 Real-Time PCR
system (Roche Diagnostics, Basel, Switzerland). The cycling
conditions were as follows: 50°C for 2 min and 95°C for 2 min,
followed by 40 cycles of: 95°C for 15 sec, 55°C for 15 sec, 72°C
for 1 min and 72°C for 1 min. Relative transcripts of mRNA were
calculated using the quantification cycle (Cq) 2−ΔΔCq
formula (26). β-2 micro-globulin
served as an internal control. Sequences of the PCR primers are
provided in Table I.
 | Table IPrimer sequences for reverse
transcription-quantitative polymerase chain reaction. |
Table I
Primer sequences for reverse
transcription-quantitative polymerase chain reaction.
| Primer name | Forward
(5′→3′) | Reverse
(5′→3′) |
|---|
| Mouse TNF-α |
ATGAGCACAGAAAGCATGA |
AGTAGACAGAAGAGCGTGGT |
| Mouse IL-6 |
CCTCTGGTCTTCTGGAGTACC |
ACTCCTTCTGTGACTCCAGC |
| Mouse IL-1β |
AATGAAAGACGGCACACCCA |
TGCTTGTGAGGTGCTGATGT |
| Mouse IFN-γ |
TTCTTCAGCAACAGCAAGGC |
TCAGCAGCGACTCCTTTTCC |
| Mouse MCP-1 |
GCTCAGCCAGATGCAGTTAA |
TCTTGAGCTTGGTGACAAAAACT |
| Mouse MIP-2 |
TGAACTGCGCTGTCAATGC |
GCTTCAGGGTCAAGGCAAAC |
| Mouse occludin |
CTACGGAGGTGGCTATGGAG |
AGCGCTGACTATGATCACGA |
| Mouse ZO-1 |
ACGATCTCCTGACCAACGTT |
GCTTTGGGTGGATGATCGTC |
| Mouse B2M |
CGGCCTGTATGCTATCCAGA |
GGGTGAATTCAGTGTGAGCC |
Western blot analysis
Following anesthesia, the chests of the mice were
opened and the lung was harvested and homogenized in RIPA buffer
(Beyotime Institute of Biotechnology, Haimen, China). The
supernatant was used to measure protein concentration according to
the BCA method (Biorad Laboratories, Inc., Hercules, CA, USA). SDS
(3X) was subsequently added and the mixture was heated to 95°C for
5 mins. The proteins (50 µg per lane) were electrophoresed
on 10% SDS-PAGE (80 V for 30 min in condensed gel and 120 V for 90
min in dissociated gel), separated by 10% SDS-PAGE gels and
electroblotted onto polyvinylidene difluoride membranes (EMD
Millipore, Billerica, MA, USA) at 90 V for 90 min. The density of
the bands were quantitated using ImageJ software (version 1.47;
National Institutes of Health, Bethesda, MD, USA) and normalized to
that of β-actin. The following primary antibodies were used:
Polyclonal, anti-rabbit VDR (cat. no. C20; 1:1,000 dilution; Santa
Cruz Biotechnology, Inc., Dallas, TX, USA), monoclonal, anti-mouse
β-actin (cat. no. A5316; 1:10,000 dilution; Sigma-Aldrich),
monoclonal, anti-mouse zonula occludens-1 (ZO-1; cat. no. 339100;
1:2,000 dilution; Cell Signaling Technology, Inc., Danvers MA, USA)
and monoclonal, anti-mouse occludin (cat. no. 331500; 1:2,000
dilution; Cell Signaling Technology, Inc.). The PVDF membranes were
incubated at room temperature for 1 h in 5% Tris-buffered saline
and Tween-20 (TBST) with non-fat milk to block non-specific
binding, then incubated overnight at 4°C with the primary
antibodies. After washing three times in 0.1% TBST, the membranes
were incubated at room temperature for 60 min with horseradish
peroxidase-conjugated secondary antibodies (goat anti-rabbit and
anti-mouse IgG-HRP; cat. nos. sc-2004 and sc-2005, respectively;
1:2,000 dilution; Santa Cruz Biotechnology, Inc.). The membrane was
then washed three times in 0.1% TBST.
Histology and immunofluorescence
The mice lungs were harvested and the right lung was
placed in 4% formalin overnight, dehydrated with graded alcohol,
placed in xylene for 1 h and then embedded in paraffin at 60°C.
Sections of the lung tissues (4 µm) were stained with
hematoxylin and eosin (H&E; Beyotime Institute of
Biotechnology) at room temperature. The lung morphology resulting
from the different treatments was scored according to H&E
stained slides using a novel acute lung injury scoring system
(27). A total of five random
fields were selected and the evaluation was completed by two
pathologists blinded to the study design. To localize the
expression of tight junction proteins, sections were incubated with
the anti-ZO-1 (1:200 dilution) or anti-occludin (1:200 dilution)
antibodies, and were subsequently conjugated with Alexa Fluor 555
or 488 secondary antibodies (Invitrogen; Thermo Fisher Scientific,
Inc.). Antigens were visualized using a Leica DFC425 fluorescence
microscope [Leica Microsystems (Schweiz) AG, Heerbrugg,
Switzerland].
Assessment of lung injury
The wet-dry ratio of lung weight was measured, as
previously described (21).
Briefly, the left lung was excised and the wet weight was measured
using a Genimi-20 Portable Milligram Scale (American Weigh Scales,
Inc., Norcross, GA, USA), the lung was then dried in an oven at
85°C for 72 h and weighed dry. The wet-dry ratio was calculated as
follows: Wet lung weight/dry lung weight. The BALF protein
concentration was determined using a DC Protein Assay kit (Bio-Rad
Laboratories, Inc.), according to the manufacturer's protocol.
Myeloperoxidase (MPO) activity was analyzed using a Myeloperoxidase
Activity Assay kit, according to the manufacturer's protocol
(CytoStore, Inc., Alberta, Canada) and an M200 microplate system
(Tecan Group Ltd., Männedorf, Switzerland) was used to measure the
optical density. In addition, the BALF to serum fluorescence ratio
of fluorescein isothiocyanate-conjugated 4 kDa dextran (FD4;
Sigma-Aldrich) was calculated to evaluate the pulmonary
permeability, as previously described (21).
Statistical analysis
All continuous data are presented as the mean ±
standard deviation. Statistical comparison of continuous variables
between groups was performed using Student's t-test or one-way
analysis of variance with GraphPad Prism (version 6.0; GraphPad
Software, Inc., La Jolla, CA, USA) and Statistical Product and
Service Solutions (version 17.0; SPSS, Inc., Chicago, IL, USA).
P<0.05 indicated a statistically significant difference.
Results
VDR KO mice exhibit more severe
LPS-induced ALI than VDR WT mice
VDR WT and KO mice were bred and genotyped according
to the protocol of The Jackson Laboratories. Genotype and VDR
expression were confirmed using the lung lysates (Fig. 1A and B). The lungs of VDR KO mice
appeared more injured and congested following LPS treatment
compared with VDR WT mice (Fig.
1C). VDR KO mice also exhibited more severe interstitial edema,
alveolar wall thickening and inflammatory cells infiltration
compared with WT mice (Fig. 1D).
Furthermore, ALI scores of VDR KO mice were significantly higher
than VDR WT mice following LPS treatment (P<0.001; Fig. 1E).
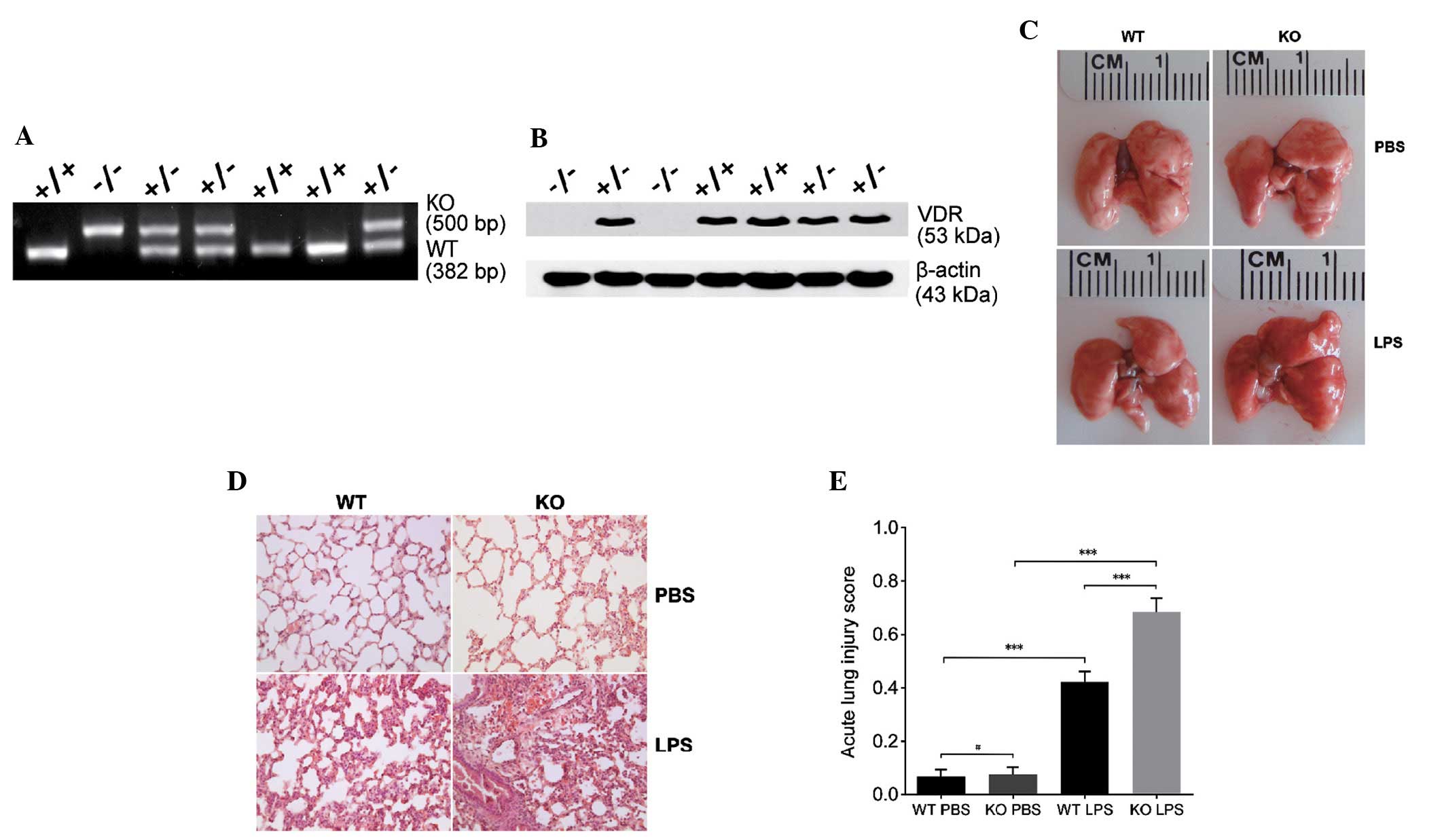 | Figure 1VDR KO mice exhibit more severe acute
lung injury (ALI) than CDR WT mice after LPS treatment. (A) Mouse
genotyping of VDR [WT(+), KO(−)]. (B) Western blotting of lung
lysates from WT (VDR+/+), heterozygous
(VDR+/−) and KO (VDR−/−) mice with anti-VDR
antibody. Following exposure to LPS or PBS, the (C) morphology and
(D) histology of lung tissue sections from WT and VDR KO mice were
investigated. Hematoxylin and eosin stain; magnification, x200. (E)
ALI scores according to microscopic examination for the two mouse
models. Data are presented as mean ± standard deviation (n=5).
#P>0.05, ***P<0.001. KO, knockout; WT,
wild-type; VDR, vitamin D receptor; PBS, phosphate buffered saline;
LPS, lipopolysaccharides. |
In mice lacking VDR, pulmonary permeability was
increased and MPO activity was higher. More specifically, a
significantly higher wet-dry lung ratio was observed (P<0.05;
Fig. 2A) in VDR KO mice subsequent
to LPS exposure compared with WT mice, indicating that more fluid
was retained in the VDR KO lung. VDR KO mice were also identified
to have higher expression levels of BALF protein (Fig. 2B; P<0.001), and greater FD4
leakage from the lung interstitium or capillaries into the alveolar
space (Fig. 2C; P<0.001)
compared with VDR WT mice. Furthermore, exposure to LPS induced
significantly higher MPO activity in the lungs of VDR KO mice
compared with VDR WT mice (P<0.001; Fig. 2D).
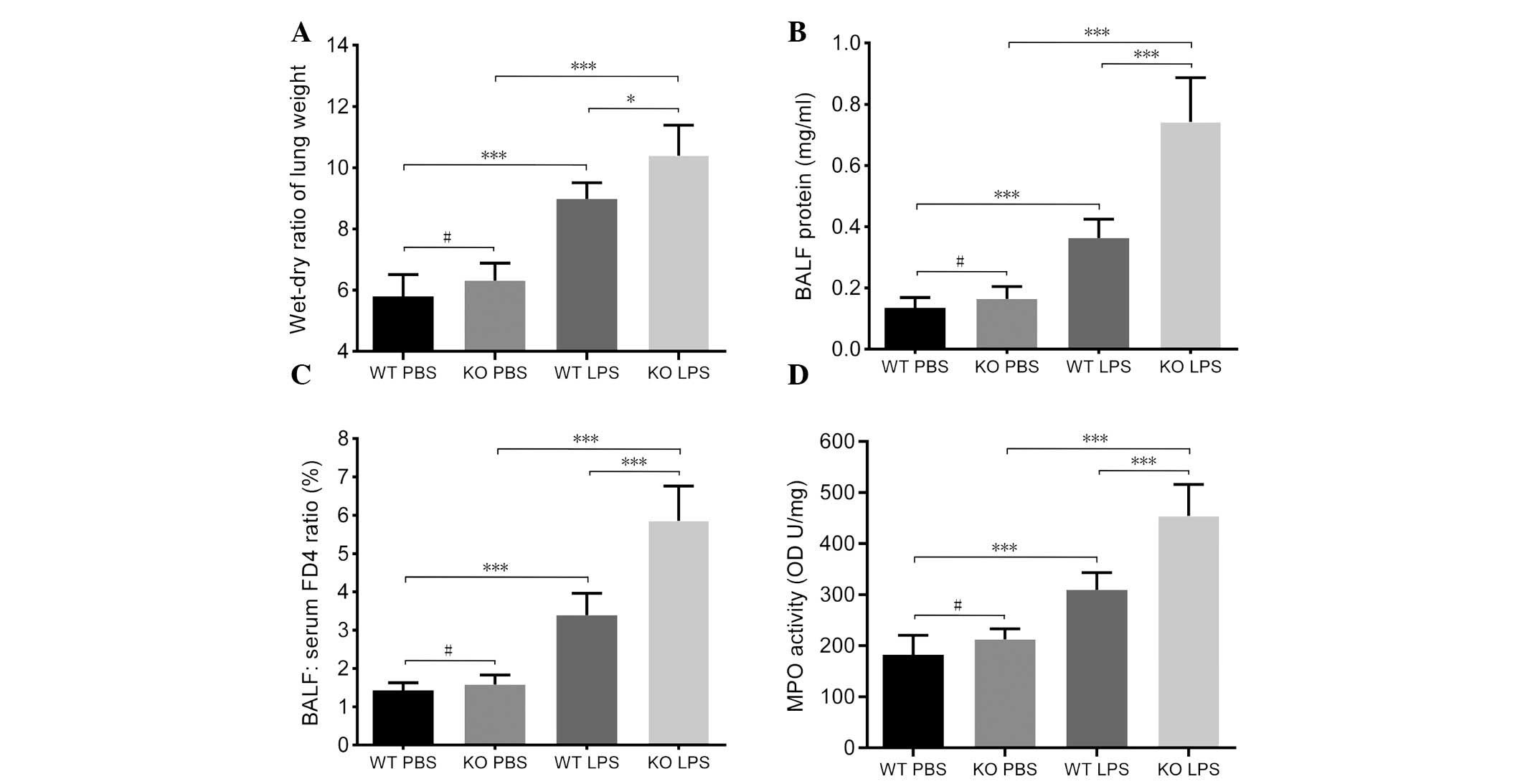 | Figure 2VDR KO mice exhibit greater pulmonary
edema than VDR WT mice. WT and KO mice were treated with PBS or LPS
intratracheally. Lung samples were harvested and analyzed 12 h
later. (A) Wet-dry ratio of lung weight, (B) BALF protein
concentration, (C) BALF to serum FD4 ratio and (D) MPO activity of
lung lysates of different mice. Data are presented as mean ±
standard deviation. #P>0.05, *P<0.05,
***P<0.001. (n=5–6 per group). WT, wild-type; PBS,
phosphate-buffered saline; KO, knockout; LPS, lipopolysaccharides;
FD4, fluorescein isothiocyanate-conjugated 4 kDa dextran; MPO,
myeloperoxidase. |
VDR KO leads to severe lung
inflammation
To further investigate the anti-inflammatory effect
of VDR in lungs, the levels of proinflammatory cytokines and
chemokines were determined. RT-qPCR indicated that the mRNA
expression levels of proinflammatory cytokines and chemokines were
significantly higher in VDR KO mice compared with VDR WT mice
following LPS treatment (Fig.
3).
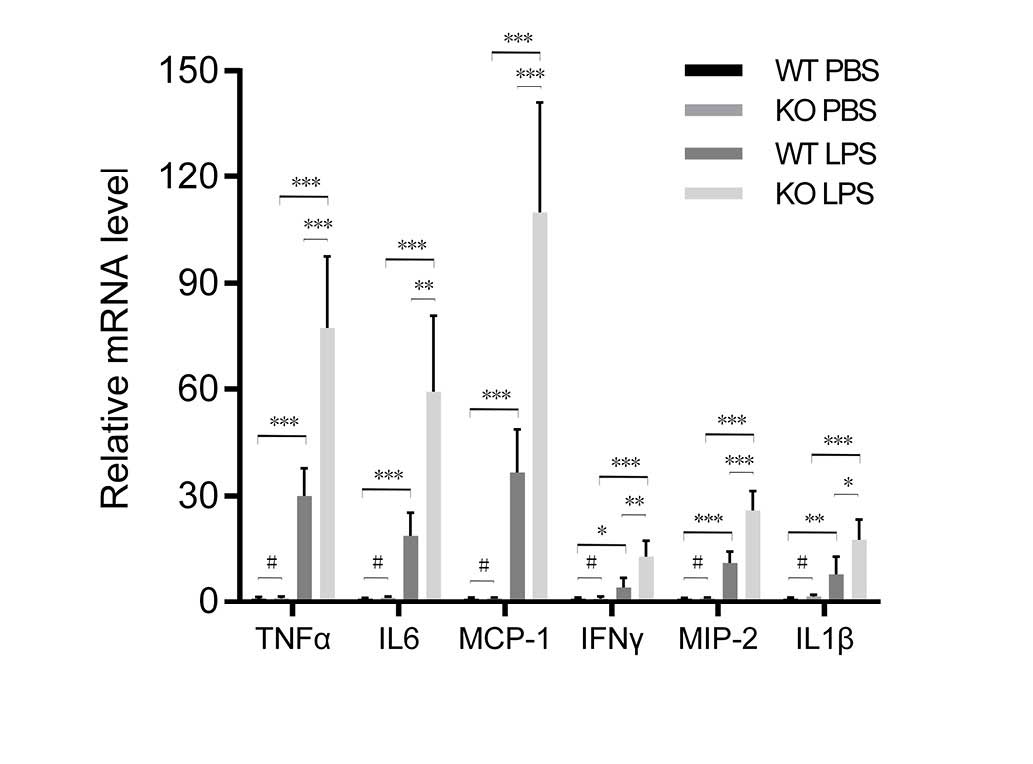 | Figure 3Quantified mRNA expression of
proinflammatory cytokines and chemokines is higher in KO compared
with WT mice following LPS or PBS treatment. Data are presented as
mean value ± standard deviation. #P>0.05,
*P<0.05, **P<0.01,
***P<0.001 (n=6). WT, wild-type; PBS,
phosphate-buffered saline; KO, knockout; LPS, lipopolysaccharides;
TNF-α, tumor necrosis factor-α; IL-6, interleukin-6; MCP-1,
monocyte chemotactic protein-1; IFNγ, interferon γ; MIP-2,
macrophage inflammatory protein-2; IL-1β, interleukin-1β. |
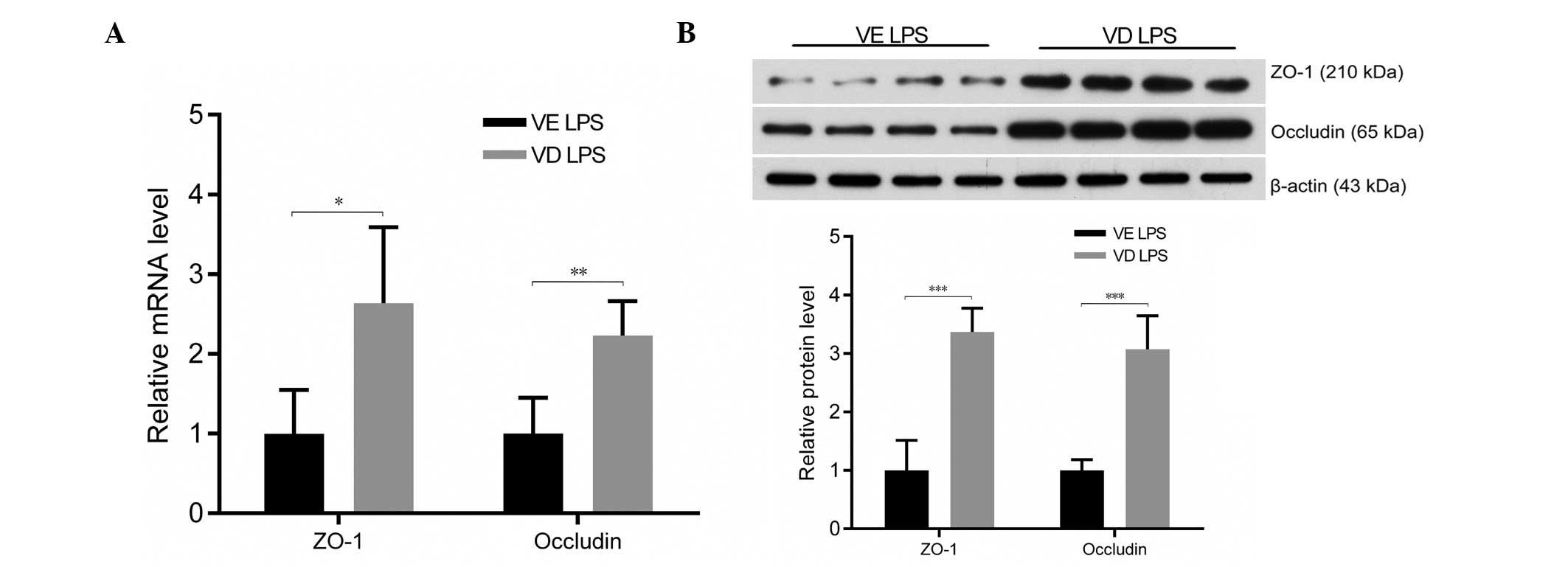
VDR KO decreases the expression of
pulmonary epithelial tight junction proteins, occludin and
ZO-1
Following exposure to LPS, mRNA (Fig. 4A) and protein (Fig. 4B) expression levels of occludin and
ZO-1 were significantly lower in VDR KO mice compared with VDR WT
mice (P<0.01). Furthermore, LPS treatment resulted in lower
levels of expression of occludin (Fig.
4C) and ZO-1 (Fig. 4D) in the
alveolar epithelial cells of all mice treated with LPS, with VDR KO
mice demonstrating a more severely disrupted expression pattern
than VDR WT mice. Therefore, the lack of VDR may compromise the
function of the pulmonary barrier by decreasing the expression of
occludin and ZO-1.
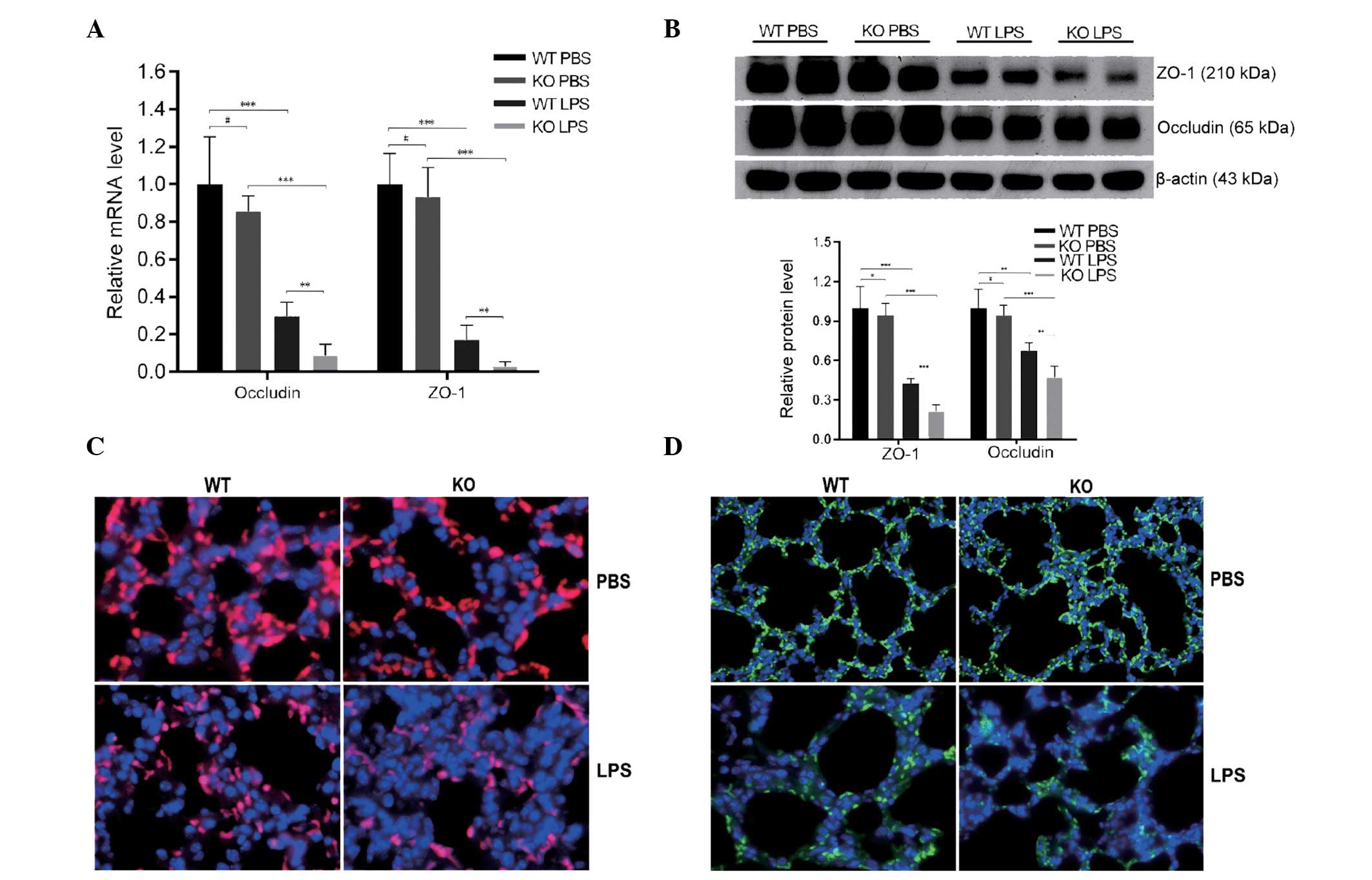 | Figure 4KO mice exhibit reduced occludin and
ZO-1 expression following LPS treatment. (A) Quantification of
occludin and ZO-1 mRNA expression in WT and KO mice following LPS
or PBS treatment. (B) Western blots and quantification of ZO-1 and
occludin protein levels in lung lysates. (C) Occludin (red,
occludin and Alexa Fluor 488; blue, DAPI) and (D) ZO-1 (blue, DAPI;
green, ZO-1 and Alexa Fluor 488) immunofluorescence staining. Data
are presented as the mean ± standard deviation
#P>0.05, **P<0.01,
***P<0.001 (n=5–6 per group). WT, wild-type; PBS,
phosphate-buffered saline; KO, knockout; LPS, lipopolysaccharides;
ZO-1, zonula occludens-1. |
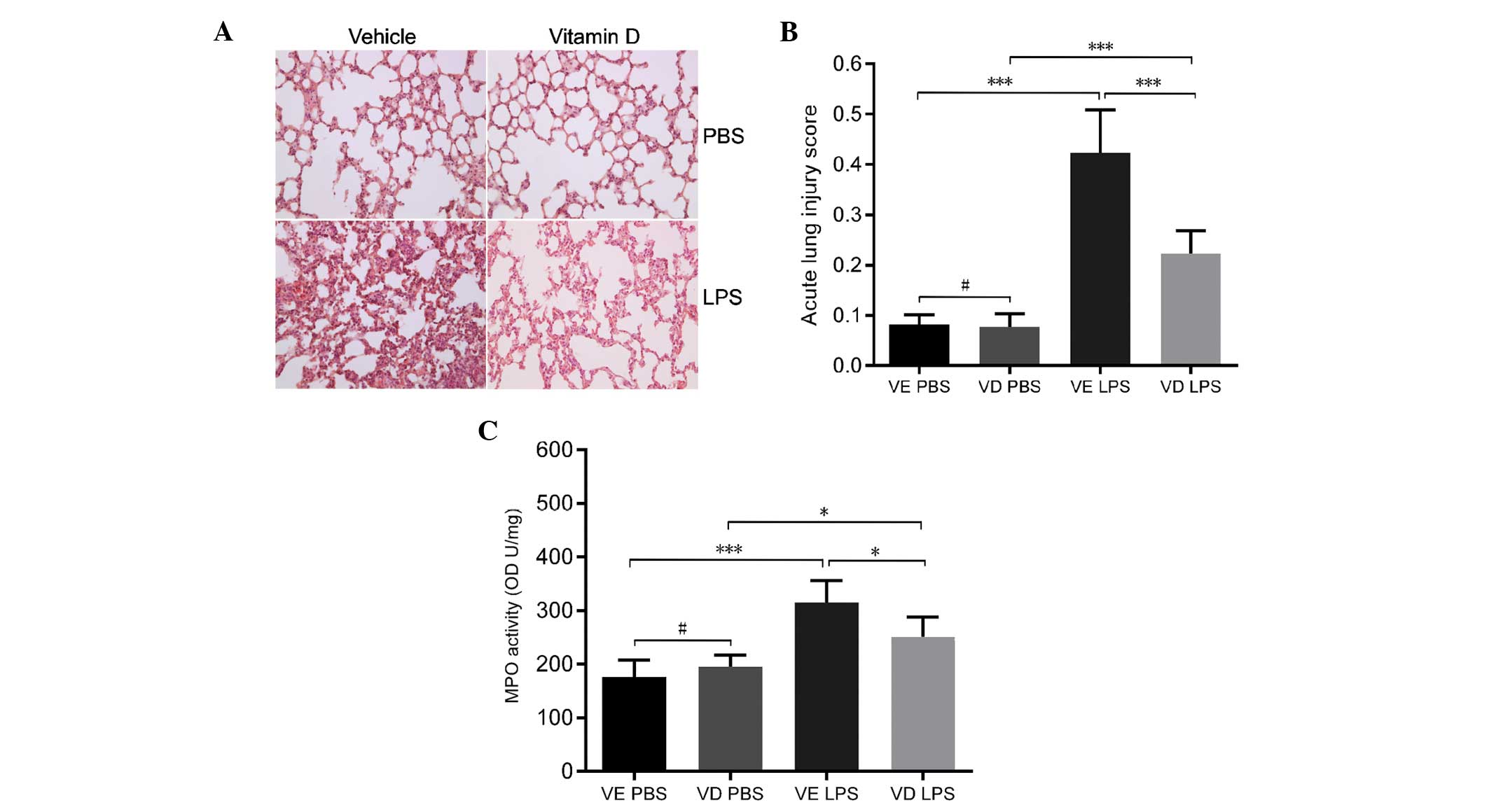
Vitamin D analog treatment alleviates
LPS-induced ALI by preserving occludin and ZO-1 expression
Histological examination of LPS-exposed vehicle mice
revealed a considerable infiltration of inflammatory cells and
thickening of the alveolar wall. However, in mice treated with
vitamin D analog prior to LPS exposure, pulmonary inflammation and
the thickening of the alveolar wall was less apparent (Fig. 5A). This was accompanied by a
significantly decreased ALI score in the vitamin D pretreated mice
compared with the vehicle treated mice (P<0.001; Fig. 5B). MPO activity was also
significantly lower in VD mice (P<0.05; Fig. 5C). The induction of proinflammatory
cytokine and chemokine expression was significantly suppressed by
vitamin D treatment (Fig. 6A and
B). Furthermore, animals pretreated with vitamin D experienced
significantly less severe pulmonary edema (P<0.05; Fig. 6C) and lower levels of BALF protein
debris entering the alveolar space (P<0.01; Fig. 6D) compared with VE mice. Vitamin D
pretreated mice exhibited significantly higher mRNA (Fig. 7A) and protein (Fig. 7B) expression levels of occludin and
ZO-1 compared with vehicle treated mice. This may be due to VDR
decreasing the permeability of the pulmonary epithelial barrier,
thus limiting the pathological changes that occur during ALI.
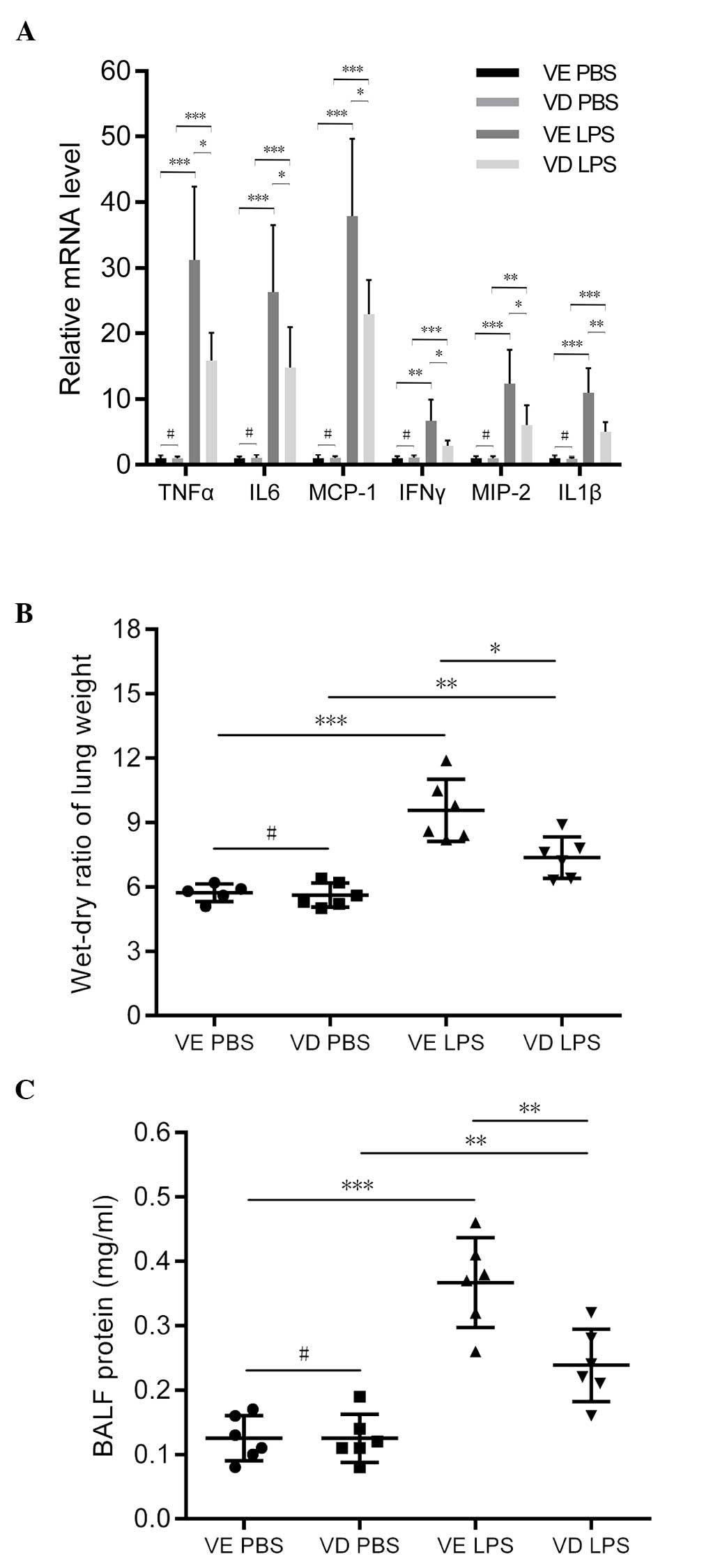 | Figure 6Vitamin D treatment inhibits lung
inflammation and alleviated pulmonary edema. (A) Reverse
transcription-polymerase chain reaction quantitation of
proinflammatory cytokines and chemokines. (B) Wet-dry ratio of lung
weight. (C) BALF protein concentration. Data are presented as mean
value ± standard deviation. #P>0.05,
*P<0.05, **P<0.01,
***P<0.001, (n=4–6 per group). VE, vehicle; VD,
vitamine D (paricalcitol); PBS, phosphate-buffered saline; LPS,
lipopolysaccharides; TNF-α, tumor necrosis factor-α; IL-6,
interleukin-6; MCP-1, monocyte chemotactic protein-1; IFN γ,
interferon γ; MIP-2, macrophage inflammatory protein-2; IL-1β,
interleukin-1β; BALF, bronchoalveolar lavage fluid. |
Discussion
Vitamin D deficiency impairs lung function and has
been associated with a number of lung diseases, including COPD,
asthma, tuberculosis, ALI and its severe form, ARDS (8–17);
however the underlying mechanisms of these associations remain
unclear. The present study demonstrated that VDR null mice exhibit
more severe LPS-induced ALI, primarily due to deterioration of the
alveolar epithelial tight junctions through a decrease in occludin
and ZO-1 expression. By contrast, vitamin D treatment alleviated
lung injury through maintenance of the pulmonary barrier. To the
best of our knowledge, this was the first study to investigate the
protective function of the vitamin D/VDR signaling pathway on the
pulmonary epithelial barrier.
ARDS remains a major cause of morbidity and
mortality in critically ill patients (1–3). It
is characterized by the disruption of the endothelial and
epithelial barriers of alveoli, leading to increased barrier
permeability (1,2). The alveolar epithelial barrier
consists of a monolayer of epithelial cells with intercellular
junctions that seal the paracellular space and regulate barrier
permeability (21,22). Preserving or restoring the barrier
function of alveolar epithelial cells may be a novel treatment for
sepsis-induced ARDS. Tight junction complexes are composed of
integral membrane proteins, cytoplasmic plaque proteins and
cytoskeletal proteins (28). Among
these, occludin and ZO-1 are key components that regulate
paracellular permeability. They are indispensible in alveolar
epithelial barrier function and fluid clearance (21). Reduced or dysmorphic expression of
occludin and ZO-1 may compromise the alveolar barrier function and
result in increased alveolar permeability, thus impairing blood-gas
exchange. In a previous study, hyperoxia was identified to disrupt
the pulmonary epithelial barrier in newborn rats by decreasing
occludin and ZO-1 levels (21).
Therefore, disruption of the alveolar epithelial barrier is a
critical factor in the pathogenesis of lung injuries and subsequent
pathological changes; however, the molecular mechanisms that
influence components of the tight junction remain poorly
understood.
The active form of vitamin D is
1,25-(OH)2D3, and is important in maintaining
the structure and function of epithelial barriers in multiple
tissues (18–20). The administration of
1,25-(OH)2D3 as part of a therapeutic regimen
may revert proteinuria and inhibit glomerular podocytes injury,
partially through improving the barrier function in the kidneys
(29). VDR null mice are more
susceptible to colonic injury induced by dextran sulfate sodium
compared with WT mice, as the lack of VDR compromises the
intestinal epithelial barrier structure (20). Thus, the vitamin D/VDR signaling
pathway may be a target in the treatment of various inflammatory
diseases through preserving or restoring epithelial barrier
function. In the present study, VDR KO mice exhibited more severe
ALI induced by LPS compared with WT mice. Furthermore, occludin and
ZO-1 levels in VDR KO mice were lower compared with WT mice. This
led to greater infiltration of inflammatory cells, release of
proinflammatory cytokines and chemokines, and fluid retention.
However, vitamin D treatment may partly reverse this pathological
process. The present study indicates that vitamin D/VDR signaling
may inhibit endotoxin-induced ALI through maintaining the integrity
of pulmonary epithelial barrier.
Neutrophil recruitment is important in the
progression of ALI and ARDS (30).
The infiltration of neutrophils may be evaluated through MPO
activity. In the present study, VDR KO mice exposed to LPS
exhibited higher MPO activity in the lung lysates compared with WT
mice. When vitamin D pretreatment was applied, lower MPO activity
was observed. In agreement with these results, a previous study
used a hamster model of ALI to identify that
1,25-(OH)2D3 inhibits neutrophil recruitment
by 40%, due to its inhibitory effect on interleukin-8 (IL-8)
(30). The alveolar barrier limits
neutrophil infiltration, thus the inhibitory effect of neutrophil
recruitment by vitamin D/VDR signaling may be partly due to the
maintenance of the integrity of the alveolar barrier.
Furthermore, lung epithelial cells had been
identified to express high baseline levels of 1α-hydroxylase, which
activates vitamin D, and low levels of inactivating 24-hydroxylase
(7). Active vitamin D generated by
lung epithelial cells is important for innate immune functions
(7). During sepsis,
pro-inflammatory cytokines and chemokines are produced by activated
alveolar macrophages in the air space. Tumor necrosis factor-α
(TNF-α), IL-6 and IL-1β proteins are also important as they
mediate, amplify and promote the process of lung inflammation
(1). These cascades of immune
responses result in various stages of alveolar epithelial injury
(1). Furthermore,
1,25-(OH)2D3 may ameliorate
seawater-aspiration induced ALI through the inhibition of nuclear
factor-κB and the RhoA/Rho kinase signaling pathways (31). In addition, it may also alleviate
lung damage secondary to ischemia reperfusion injury (32). The present study identified that
LPS treatment increased the expression levels of the following
cytokines and chemokines in WT mice: TNF-α, IL-6, IL-1β,
interferon-γ, monocyte chemoattractant protein-1 and macrophage
inflammatory protein-2. This effect was even more apparent in VDR
KO mice, thus confirming that the VDR KO lung was markedly more
inflamed than the WT lung. By contrast, vitamin D treatment
substantially inhibited the build-up of chemokines and attenuated
LPS-induced lung injury. Furthermore, the present study determined
that active vitamin D was capable of triggering an
anti-inflammatory defense and preserving the structural barrier in
the lungs.
In conclusion, the current study highlights that
lack of VDR may compromise the pulmonary epithelial barrier
defense, leading to a more severe LPS-induced lung injury.
Furthermore, vitamin D treatment may preserve the alveolar barrier
function and therefore alleviate LPS-induced lung injury. These
observations provide further insight into the pathogenesis of
ALI/ARDS and emphasize that vitamin D may be a novel treatment for
ALI/ARDS.
Acknowledgments
This study was supported by the National Natural
Science Foundation of China (grant nos. 81170605, 81471489,
81271938, 81270726) and the Outstanding Scientific Fund of Sheng
Jing Hospital (Shenyang, China).
References
|
1
|
Ware LB and Matthay MA: The acute
respiratory distress syndrome. N Engl J Med. 342:1334–1339. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Parekh D, Dancer RC and Thickett DR: Acute
lung injury. Clin Med. 11:615–618. 2011. View Article : Google Scholar
|
|
3
|
Matthay MA, Zimmerman GA, Esmon C,
Bhattacharya J, Coller B, Doerschuk CM, Floros J, Gimbrone MA Jr,
Hoffman E, Hubmayr RD, et al: Future research directions in acute
lung injury: Summary of a National Heart, Lung, and Blood Institute
working group. Am J Respir Crit Care Med. 167:1027–1035. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Haussler MR, Whitfield GK, Haussler CA,
Hsieh JC, Thompson PD, Selznick SH, Dominguez CE and Jurutka PW:
The nuclear vitamin D receptor: Biological and molecular regulatory
properties revealed. J Bone Miner Res. 13:325–349. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bouillon R, Carmeliet G, Verlinden L, van
Etten E, Verstuyf A, Luderer HF, Lieben L, Mathieu C and Demay M:
Vitamin D and human health: Lessons from vitamin D receptor null
mice. Endocr Rev. 29:726–776. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Adams JS and Hewison M: Update in vitamin
D. J Clin Endocrinol Metab. 95:471–478. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hansdottir S, Monick MM, Hinde SL, Lovan
N, Look DC and Hunninghake GW: Respiratory epithelial cells convert
inactive vitamin D to its active form: Potential effects on host
defense. J Immunol. 181:7090–7099. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Afzal S, Lange P, Bojesen SE, Freiberg JJ
and Nordestgaard BG: Plasma 25-hydroxyvitamin D, lung function and
risk of chronic obstructive pulmonary disease. Thorax. 69:24–31.
2014. View Article : Google Scholar
|
|
9
|
Rippel C, South M, Butt WW and
Shekerdemian LS: Vitamin D status in critically ill children.
Intensive Care Med. 38:2055–2062. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lee P, Eisman JA and Center JR: Vitamin D
deficiency in critically ill patients. N Engl J Med. 360:1912–1914.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Barnett N, Zhao Z, Koyama T, Janz DR, Wang
CY, May AK, Bernard GR and Ware LB: Vitamin D deficiency and risk
of acute lung injury in severe sepsis and severe trauma: A
case-control study. Ann Intensive Care. 4:5–14. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Parekh D, Dancer RC, Lax S, Cooper MS,
Martineau AR, Fraser WD, Tucker O, Alderson D, Perkins GD,
Gao-Smith F and Thickett DR: Vitamin D to prevent acute lung injury
following oesophagectomy (VINDALOO): Study protocol for a
randomised placebo controlled trial. Trials. 14:100–106. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hollams EM, Hart PH, Holt BJ, Serralha M,
Parsons F, de Klerk NH, Zhang G, Sly PD and Holt PG: Vitamin D and
atopy and asthma phenotypes in children: A longitudinal cohort
study. Eur Respir J. 38:1320–1327. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wilkinson RJ, Llewelyn M, Toossi Z, Patel
P, Pasvol G, Lalvani A, Wright D, Latif M and Davidson RN:
Influence of vitamin D deficiency and vitamin D receptor
polymorphisms on tuberculosis among Gujarati Asians in west London:
A case-control study. Lancet. 355:618–621. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Persson LJ, Aanerud M, Hiemstra PS, Hardie
JA, Bakke PS and Eagan TM: Chronic obstructive pulmonary disease is
associated with low levels of vitamin D. PLoS One. 7:e38934–e38941.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Amrein K, Zajic P, Schnedl C,
Waltensdorfer A, Fruhwald S, Holl A, Purkart T, Wünsch G, Valentin
T, Grisold A, et al: Vitamin D status and its association with
season, hospital and sepsis mortality in critical illness. Crit
Care. 18:R472014. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Parekh D, Thickett DR and Turner AM:
Vitamin D deficiency and acute lung injury. Inflamm Allergy Drug
Targets. 12:253–261. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hartmann B, Riedel R, Jörss K,
Loddenkemper C, Steinmeyer A, Zügel U, Babina M, Radbruch A and
Worm M: Vitamin D receptor activation improves allergen-triggered
eczema in mice. J Invest Dermatol. 132:330–336. 2012. View Article : Google Scholar
|
|
19
|
Yin Z, Pintea V, Lin Y, Hammock BD and
Watsky MA: Vitamin D enhances corneal epithelial barrier function.
Invest Ophthalmol Vis Sci. 52:7359–7364. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kong J, Zhang Z, Musch MW, Ning G, Sun J,
Hart J, Bissonnette M and Li YC: Novel role of the vitamin D
receptor in maintaining the integrity of the intestinal mucosal
barrier. Am J Physiol Gastrointest Liver Physiol. 294:G208–G216.
2008. View Article : Google Scholar
|
|
21
|
You K, Xu X, Fu J, Xu S, Yue X, Yu Z and
Xue X: Hyperoxia disrupts pulmonary epithelial barrier in newborn
rats via the deterioration of occludin and ZO-1. Respir Res.
13:36–46. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Overgaard CE, Mitchell LA and Koval M:
Roles for claudins in alveolar epithelial barrier function. Ann N Y
Acad Sci. 1257:167–174. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Budinger GR and Sznajder JI: The
alveolar-epithelial barrier: A target for potential therapy. Clin
Chest Med. 27:655–669, abstract ix. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bhattacharya J and Matthay MA: Regulation
and repair of the alveolar-capillary barrier in acute lung injury.
Annu Rev Physiol. 75:593–615. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li YC, Pirro AE, Amling M, Delling G,
Baron R, Bronson R and Demay MB: Targeted ablation of the vitamin D
receptor: An animal model of vitamin D-dependent rickets type II
with alopecia. Proc Natl Acad Sci USA. 94:9831–9835. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Golan MA, Liu W, Shi Y, Chen L, Wang J,
Liu T and Li YC: Transgenic expression of vitamin D receptor in gut
epithelial cells ameliorates spontaneous colitis caused by
interleukin-10 deficiency. Dig Dis Sci. 60:1941–19477. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Matute-Bello G, Downey G, Moore BB,
Groshong SD, Matthay MA, Slutsky AS and Kuebler WM: Acute Lung
Injury in Animals Study Group: An official American Thoracic
Society workshop report: Features and measurements of experimental
acute lung injury in animals. Am J Respir Cell Mol Biol.
44:725–738. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Schneeberger EE and Lynch RD: The tight
junction: a multifunctional complex. Am J Physiol Cell Physiol.
286:C1213–1228. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Migliori M, Giovannini L, Panichi V,
Filippi C, Taccola D, Origlia N, Mannari C and Camussi G: Treatment
with 1,25-dihy-droxyvitamin D3 preserves glomerular slit
diaphragm-associated protein expression in experimental
glomerulonephritis. Int J Immunopathol Pharmacol. 18:779–790.
2005.
|
|
30
|
Takano Y, Mitsuhashi H and Ueno K:
1α,25-dihydroxyvitamin D3 inhibits neutrophil
recruitment in hamster model of acute lung injury. Steroids.
76:1305–1309. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang M, Dong M, Liu W, Wang L, Luo Y, Li
Z and Jin F: 1α,25-dihydroxyvitamin D3 ameliorates seawater
aspiration-induced acute lung injury via NF-κB and RhoA/Rho kinase
pathways. PLoS One. 9:e104507–e104516. 2014. View Article : Google Scholar
|
|
32
|
Shih PK, Chen YC, Huang YC, Chang YT, Chen
JX and Cheng CM: Pretreatment of vitamin D3 ameliorates lung and
muscle injury induced by reperfusion of bilateral femoral vessels
in a rat model. J Surg Res. 171:323–328. 2011. View Article : Google Scholar
|