Introduction
Micro (mi)RNA is a small, non-coding RNA regulatory
molecule of 18–24 nucleotides in length, which
post-transcriptionally silences genes by base-pairing with the mRNA
for degradation and/or translational repression (1). miRNA has been shown to be involved in
cell proliferation, differentiation and apoptosis, and is therefore
closely associated with the occurrence and progression of a variety
of human diseases (2). It has been
reported that miRNAs are involved in inflammatory reactions and
immune responses to various neurological pathologies underlying
central nervous system injuries (3).
It was previously revealed that the highly conserved
miR-9 (homologous to miR-79) is involved in microglial inflammatory
responses following cerebral injuries (4,5).
miR-9 is encoded by three distinct genomic loci (miR-9-1, -2 and
-3), which give rise to mature miR-9 species with identical
sequences. It has also been found that miR-9 directly acts on the
3′-untranslated region (UTR) of the inflammatory regulator of
nuclear factor (NF)-κB, an integral part of the inflammatory
reaction of tumor cells (6,7).
Similarly, miR-9 has been shown to be involved in the NF-κB
signaling pathway in which microglial cells are activated by
lipopolysaccharide stimuli, and a lack of miR-9 results in the
upregulation of proinflammatory cytokines/chemokines [interleukin
(IL)-1β, tumor-necrosis factor (TNF)-α, IL-6 and monocyte
chemoattractant protein (MCP)-1] (5). NF-κB may be singularly important in
regulating genetic responses to brain stresses through the innate
immune response, since it belongs to the category of 'pre-formed'
primary transcription factors, which are already present in cells
in an 'inactive' state. In unstimulated cells, NF-κB is sequestered
in the cytosol via interacting with inhibitor of NF-κB (IκB)
proteins; however, when cells receive pathological stimuli, IκB
proteins can be phosphorylated by IκB kinase (IKK), which is
activated, leading to its translocation to the nucleus, where it
induces the expression of various proinflammatory cytokines,
including TNF-α, IL-1β and IL-6. NF-κB-regulated miR-9 has the
potential to contribute to the regulation of neuroinflammation and
innate immune signaling in stressed primary neural cells of the
human brain (8). In addition,
stroke downregulated the expression of miR-9 in neural progenitor
cells (9). A study reported that
the serum levels of miR-9 were suppressed to facilitate
neuroinflammation and brain injury, and miR-9 was negatively
correlated with the infarct volume and plasma high-sensitivity
C-reactive protein levels in patients with acute ischemic stroke
(10).
Stroke, also termed cerebrovascular disease, is
characterized by high mortality, morbidity and recurrence rates. As
an important medical methodology, electroacupuncture (EA) is a
treatment modality of disease by inserting needles along specific
pathways or meridians attaching a micro-electro-stimulation in
oriental medicine, and has been widely used in clinical practice
for motor impairment in patients with stroke (11). It has been previous demonstrated
that EA stimulation of Zusanli (ST36) and Quchi (LI11) acupoints
attenuates inflammatory injury and infarct volume in a rat model of
middle cerebral artery occlusion (MCAO) (12). EA has been shown to significantly
reduce the expression of proinflammatory cytokines, IL-1β and
TNF-α, via the NF-κB signaling pathway in ischemic cerebral tissues
(12,13); however, the affect of EA on NF-κB
mediated regulators in ischemic stroke remains to be
elucidated.
The present study hypothesized that EA regulates the
expression of miR-9 to suppress the activation of the NF-κB
signaling pathway and contribute to the release of proinflammatory
cytokines, IL-1β, IL-6 and TNF-α, in MCAO injury.
Materials and methods
Statement of ethics
All experimental procedures were approved by the
Animal Care and Use Committee of Fujian University of Traditional
Chinese Medicine (FUTCM) and followed Chinese Specifications for
the Production, Care and Use of the Laboratory Animals. All animal
experiments were permitted, according to the License no. SYXK (Min)
2012-007. Throughout the experiments, the utmost effort was made to
minimize animal suffering.
MCAO rat model for focal cerebral
ischemia
Adult male Sprague-Dawley rats (n=90; weight 230–250
g) were housed in an environmentally controlled room at FUTCM
(22±2°C, with a 12 h light/dark cycle). The rats were provided
ad libitum access to standard rat chow and water. Focal
cerebral ischemia was induced by MCAO, as previously described
(14), with minor modifications.
Briefly, left MCAO was performed using an occluding suture
(diameter, 0.26 mm) for 2 h, and following 2 h of MCAO-evoked
ischemia, the suture was slowly drawn back to allow reperfusion.
The ipsilateral cerebral blood flow was measured using laser
Doppler flowmetry (MoorDRT4; Biopac Systems, Inc. Goleta, CA, USA).
The MCAO model was considered successful only when the drop in
cerebral blood flow was ≥80% of baseline during occlusion. This
blood flow rate was maintained for at least 1 h, with the exception
of the 0 h time-point.
Rat groups and EA treatments
The rats were randomly divided into the following
five groups (n=18/group): i) Sham operation group (Sham); ii) MCAO
model group (MCAO); iii) EA group (MCAO + EA); iv) EA combined with
dimethyl sulfoxide (DMSO) group (EA + DMSO); v) EA combined with
miR-9 inhibitors using a random number table method (EA + miR-9
inhibitors) (15).
The MCAO + EA, EA + DMSO and EA + miR-9 inhibitors
groups were stimulated with an electric potential difference of 4
V, and dense disperse wave of 1- or 20-Hz (adjusted to the muscle
twitch threshold) using an EA apparatus (G6805; SMIF, Shanghai,
China) for 30 min once daily. The rats were immobilized using a
surgical fixation device (Beijing Science and Technology
Development Co., Ltd., Beijing, China) (12). EA was performed by stimulating the
right paralyzed limb at Zusanli (ST36) and Quchi (LI11) acupoints
with a depth of 2–3 mm (12,14).
The treatment was performed 24 h following the surgery. All animals
were sacrificed by cervical dislocation following anesthesia with
10% chloral hydrate (Fujian Academy of Integrative Medicine,
Fuzhou, China) at 72 h following the surgery.
Intraventricular injections
Intraventricular injections were performed in the EA
+ DMSO and EA + miR-9 inhibitors groups 30 min prior to MCAO. The
animals were anesthetized with 10% chloral hydrate and were
subsequently placed in a stereotaxic apparatus (68001; RWD Life
Science Co., Ltd., Shenzhen China). A volume of 7 μl miR-9
inhibitors (10 mM/l in DMSO) or DMSO were injected into the left
lateral ventricle in rats (16).
Stereotactic coordinates were as follows: Anteroposterior, 0.8 mm;
mediolateral, 1.5 mm; depth, 3.5 mm.
Scoring of neurological deficits
Neurological deficits in the MCAO rat model were
scored using the Longa et al (17) method following resuscitation. The
scores for the neurological behavioral were as follows: 0, Absence
of neurological injury symptoms; 1, flexion of the right front paw;
2, body turning towards the right side while walking; 3, falling
down on the right side; 4, complete loss of consciousness with the
inability to walk. Rats scoring 0 and 4 were excluded from the
present study.
2,3,5-Triphenyltetrazolium chloride (TTC)
staining
Brain tissues were dissected using surgical
apparatus (Fujian Academy of Integrative Medicine). The brain
tissues were frozen at −20°C for 20 min and cut into five 2 mm
sections in the coronal plane at the level of the optic chiasm. The
sections were preserved in 2% TTC (Sigma-Aldrich, St. Louis, MO,
USA) phosphoric acid buffer [0.2 mol/l (pH 7.4)], as previously
described (12) and placed in an
incubation box at 37°C in the dark. The sections were alternately
overturned twice and were removed following two 15 min long
alternations. The normal tissues were stained red using TTC; the
infarct loci were not stained and were white in color. Images of
the sections were captured using a digital camera (Canon SX20;
Canon, Inc., Tokyo, Japan). The percentage of the infarct volume in
the total brain volume was calculated using a Motic Med 6.0 image
analysis system (Motic Group Co., Ltd., Xiamen, China).
Hematoxylin and eosin (HE) staining
The tissue sections were embedded with paraffin and
subjected to conventional gradient dewaxing, followed by HE
staining for 3 min at 25°C. Following rinsing with flowing water
for 1–2 min, the tissue sections were placed in a 75% hydrochloric
acid/alcohol mixture to highlight hematoxylin for 30–45 sec. The
tissue sections were subsequently rinsed with flowing water for
10–20 min until the blue color returned. Successive treatment with
95% ethanol and acidified eosin-ethanol staining were performed for
1 min at 25°C. Finally, the tissue sections were dehydrated with
gradient ethanol, rendered transparent with xylene and sealed with
neutral balsam. The morphological changes were observed under a
light microscope (DM500; Leica Microsystems, Wetzlar, Germany).
Immunofluorescence
The paraffin-embedded brain-tissue sections were
treated with microwave heat-induced epitope retrieval. Following
three washes in phosphate-buffered saline (PBS; pH 7.4), the
specimens were incubated for 1 h at 37°C in a 1:120 dilution of
fluorescein isothiocyanate-labeled rabbit anti-rat NF-κB p65
antibody (cat. no. ab16502; Abcam, Cambridge, UK). Washing was
repeated, as above. The nuclei of all cells were counterstained
with 0.1 μg/ml 4′,6-diamidino-2-phenylindole (DAPI) and
incubated for 5 min at 25°C. Following a further three washes in
PBS, the tissues were mounted in Prolong Gold Antifade reagent
(Invitrogen; Thermo Fisher Scientific, Inc.). Images were captured
using a fluorescent microscope (Olympus BX61; Olympus Corp., Tokyo,
Japan) at a magnification of ×200.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
The total RNA was extracted from the ischemic cortex
using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). A SYBR Green® microRNA Reverse
Transcription kit (Guangzhou RiboBio Co., Ltd., Guangzhou, China)
was used to reverse transcribe the RNA into cDNA prior to qPCR. The
PCR primer kit used for miR-9 was rno-miR-9 (miRQP0825;
GeneCopoeia, Guangzhou, China) and the PCR primer kit used for U6
was rno-miR-U6 (RQP047936; GeneCopoeia). The GeneAmp PCR System
9600 (Thermo Fisher Scientific, Inc.) was used. The PCR cycling
conditions were 95°C for 10 sec and 38 cycles at 56.5°C for 20 min.
The relative expression of miR-9 was quantified using the
2−ΔΔCq method. The reference gene was rno-miR-U6
(RQP047936; GeneCopoeia, Guangzhou, China).
Western blotting
Cell lysis buffer (1 ml; Invitrogen; Thermo Fisher
Scientific, Inc.) and PMSF (10 μl) were added to 100 mg
brain tissue at 25°C for 15 min for protein extraction. The protein
concentration was determined using Coomassie brilliant blue
staining (Invitrogen; Thermo Fisher Scientific, Inc.). The proteins
were denatured by heating and 50 μg protein was used for 12%
sodium dodecyl sulfate-polyacrylamide gel (Promega Corp., Madison,
WI, USA) electrophoresis. The proteins were transferred onto
nitrocellulose or polyvinylidene difluoride membranes (Promega
Corp.) and were sealed with 5% non-fat milk at room temperature for
2 h. The membranes were incubated with NF-κB p65 (dilution, 1:800;
cat. no. ab16502; Abcam), IκBα (dilution, 1:800; cat. no. ab7217;
Abcam), TNF-α (1:800; cat. no. ab6671; Abcam), IL-1β (1:800; cat.
no. ab200478; Abcam) and β-actin (1:1,000; cat. no. ab189073;
Abcam) primary antibodies at 4°C overnight. The membranes were
washed and horseradish peroxide-labeled secondary antibodies were
added. The cells were incubated in an oscillation incubator at 37°C
for 1 h. The color was developed by the addition of
electrochemiluminescence solution (Invitrogen; Thermo Fisher
Scientific, Inc.), and the images were visualized using a Bio-Image
system (Bio-Rad Laboratories, Inc., Hercules, USA). Gray scale
analysis was performed for the target bands using Image-Pro Plus
software (version 7.0; UVP, LLC, Upland, CA, USA).
Luciferase reporter assay
The psiCHECK2-NF-κB-luc and psiCHECK2-NF-κB-Mut-luc
firefly luciferase reporter vectors (GeneCopoeia Inc.), which
contain the intact or mutated putative miR-9 recognition sequence
from the 3′-UTR of NF-κB, respectively, cloned downstream of the
firefly luciferase gene were constructed. The following primers
were designed (Shanghai Sangon Biological Engineering Co., Ltd.,
Shanghai, China) to PCR the 3′-UTR of human NF-κB from the total
RNA extracted from primary cortical neuronal cells: NF-κB, forward:
5′-gcuuuaaaaaaaggagaaaa-3′ and reverse: 5′-acccatctcacccattcttg-3′;
nf-κb mutant, forward: 5′-cuggcuuuaaaaaauccucuuaa-3′ and reverse:
5′-tgacacagcaactcctttgg-3′.
Each PCR product was ~1 kb and covers the putative
miR-9 recognition sequence or mutated sequence from the 3′-UTR of
NF-κB. Clones were selected following screening by restriction
digestion with XhoI and NotI. Primary cortical
neuronal cells were seeded into 96-well white assay plates (Corning
Inc., Corning, NY, USA) at a density of 30,000 cells/well 1 day
prior to transfection. A total of 10 ng of each reporter construct
was co-transfected with the miR-9 mimics or a mimic negative
control (GeneCopoeia Inc.) at a final concentration of 100 nM into
primary cortical neuronal cells using Lipofectamine 2000 (Thermo
Fisher Scientific, Inc.), according to the manufacturer's protocol.
Following incubation for 48 h, firefly and Renilla
luciferase activities were measured using the Dual-Glo luciferase
assay system, according to the manufacturer's protocol (Promega
Corp.). Renilla luciferase activity was normalized against
firefly luciferase activity for each sample. All experiments were
performed in triplicate and three wells were used for each
condition in each experiment.
Statistical analysis
All quantitative data are expressed as the mean ±
standard error and were analyzed using SPSS 17.0 statistical
analysis software (IBM, Corp., Armonk, NY, USA). The data were
subjected to a one-way analysis of variance between different
groups, followed by the least significant difference t-test.
P<0.05 was considered to indicate a statistically significant
difference. All final results were analyzed in a blinded
manner.
Results
Effects of EA on neurological deficit
scores
In the present study, the neuroprotective effect of
EA treatment was examined using neurological deficit scores.
Table I shows that all MCAO rats
demonstrated obvious neurological signs, as compared with the rats
in the Sham group (P<0.05), suggesting successful model
construction. No significant differences were observed in the
neurological signs among the MCAO, EA + DMSO and EA + miR-9
inhibitors groups at 24 h following MCAO and reperfusion. However,
72 h after MCAO and reperfusion, the neurological deficits of all
rats in the EA + DMSO group exhibited a significant improvement
compared with the MCAO and EA + inhibitors groups (P<0.05).
These results suggested that EA treatment effectively alleviated
the symptoms of neurological deficit of rats with cerebral
ischemia. Furthermore, it was shown that miR-9 inhibitors
suppressed the EA-triggered recovery of neurological function,
while DMSO did not.
 | Table INeurological deficit scores. |
Table I
Neurological deficit scores.
| Group (n=18) | Deficit score at
different durations following MCAO
|
|---|
| 2 h | 24 h | 72 h |
|---|
| Sham | 0 | 0 | 0 |
| MCAO | 1.87±0.23a | 2.25±0.18 | 2.08±0.20 |
| MCAO + EA | 1.85±0.24a | 2.07±0.28 | 1.14±0.25b |
| EA + DMSO | 1.83±0.24a | 2.17±0.28 | 1.08±0.23b |
| EA + miR-9
inhibitors | 1.87±0.18a | 2.21±0.30 | 1.84±0.20 |
Effects of EA on infarct volume
TTC-stained brain sections were evaluated 72 h after
MCAO and reperfusion. The results showed that the sections from the
Sham group were stained red, whereas the unstained white infarct
area was visible on the left side of the brain in the other groups.
Compared with the Sham group, the infarct volume in the MCAO+EA
group was significantly decreased (P<0.05; Fig. 1). Furthermore, the infarct volume
of the EA + DMSO group was lower compared with that of the MCAO and
EA + miR-9 inhibitors groups (P<0.05; Fig. 1), suggesting that the miR-9
inhibitors suppressed the cerebral protective efficacy of EA
treatment.
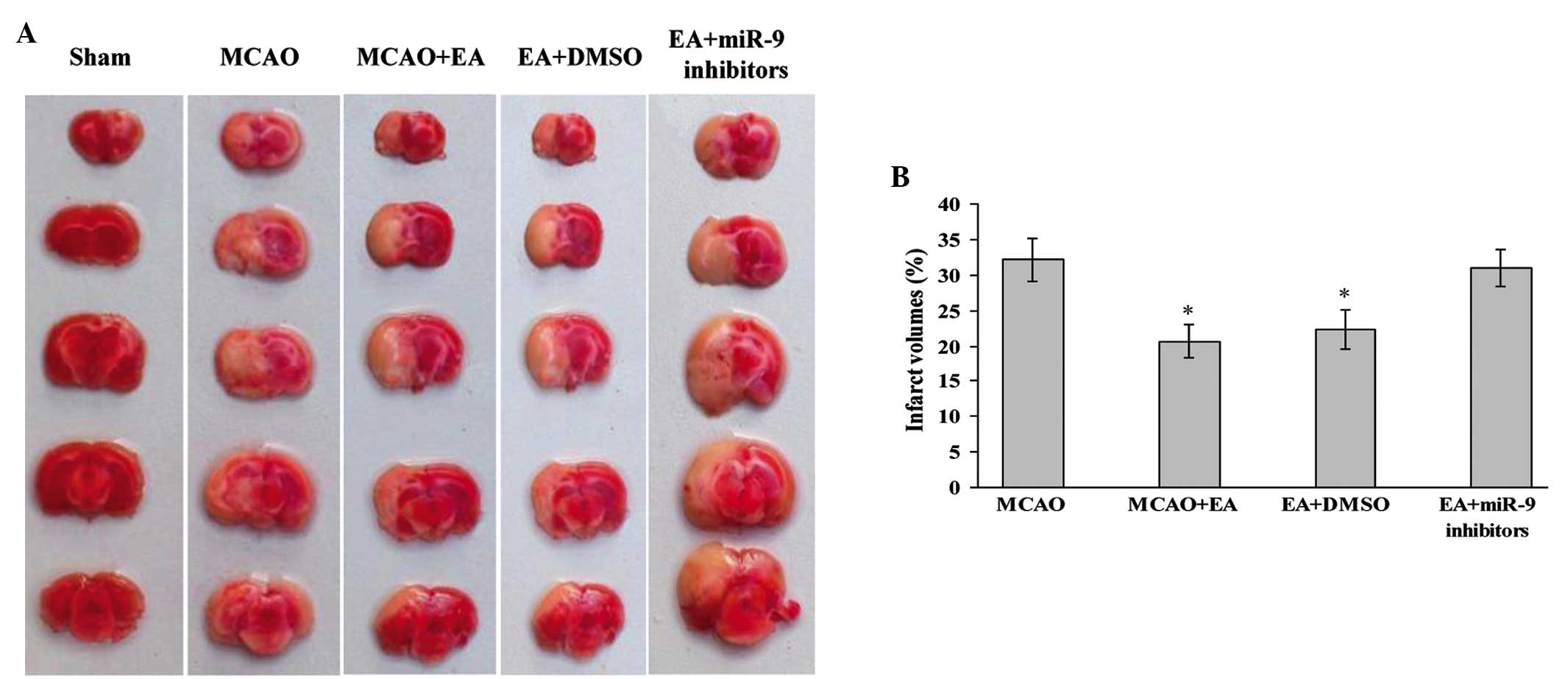 | Figure 1Effect of EA on infarct volume and
morphological structure of rat brains. (A)
2,3,5-Triphenyltetrazolium chloride staining indicating cerebral
infarct volumes of Sham, MCAO, MCAO + EA, EA + DMSO and EA + miR-9
inhibitors groups. (B) Bar graph showing the percentage of total
brain volume in each group (n=4; *P<0.05, vs. the
MCAO and the EA + miR-9 inhibitors groups). MCAO, middle cerebral
artery occlusion; EA, electroacupuncture; miR-9, microRNA-9; DMSO,
dimethyl sulfoxide. |
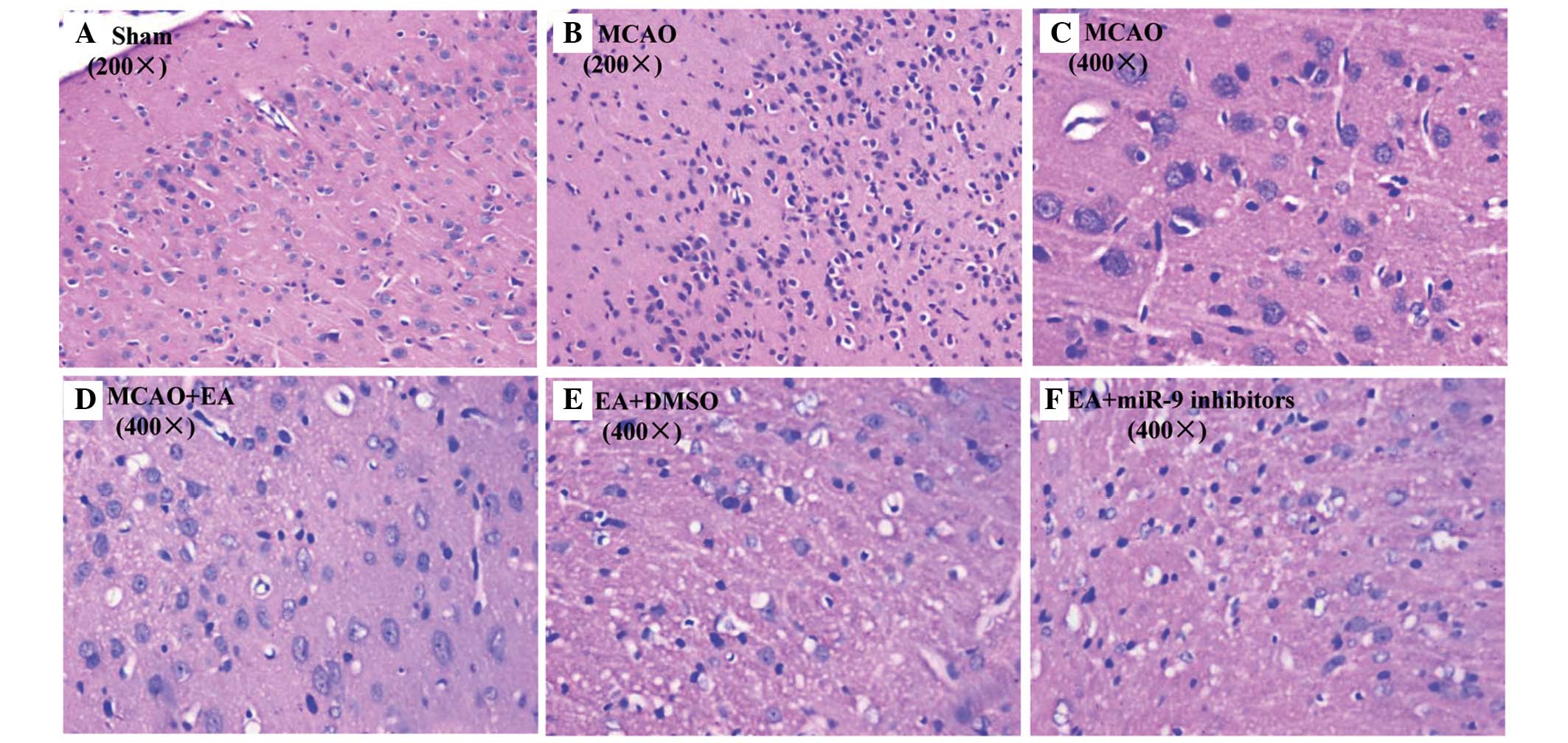
Effects of EA on inflammation
To further investigate the neuroprotective efficacy
of EA treatment, its anti-inflammatory effect was examined using HE
staining. As expected, no histopathological abnormalities and
inflammatory cells were observed in the Sham group (Fig. 2A). By contrast, in the infarct core
and bounding region of the cerebral cortical area of the MCAO
group, the glial and neuron cells exhibited interstitial edema,
were shrunken and showed condensed nuclei (Fig. 2B and C), which was ameliorated by
EA treatment (Fig. 2D).
Furthermore, compared with the MCAO group, considerably fewer
inflammatory cells were infiltrated into the cerebral infarct areas
in the EA + DMSO group (Fig. 2E),
whereas inflammatory cells were infiltrated in the EA + miR-9
inhibitors group (Fig. 2F),
suggesting that miR-9 inhibitors suppressed EA-alleviated cerebral
inflammation in MCAO rats.
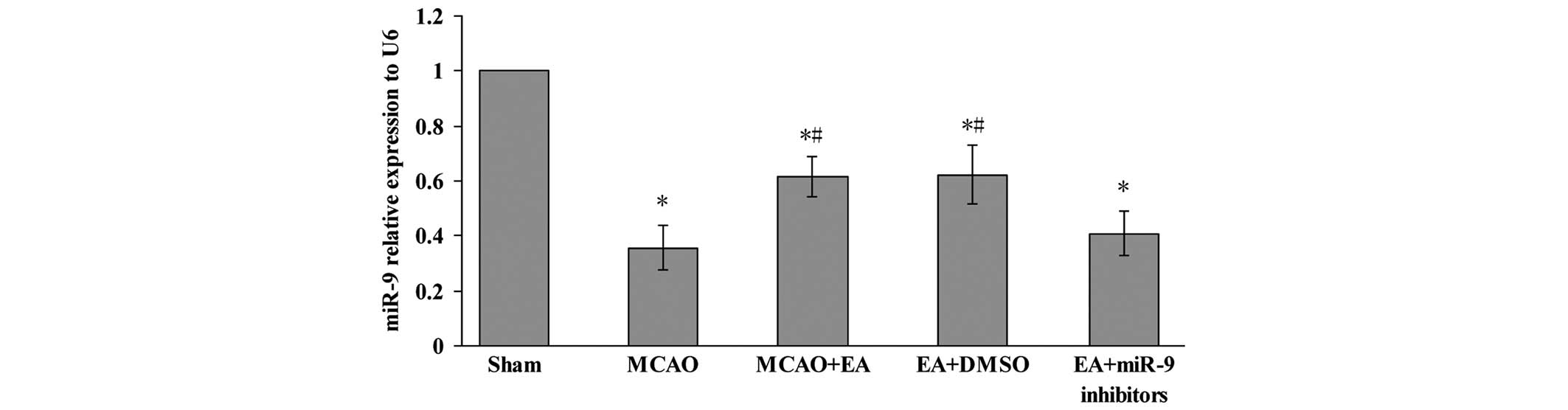
Effects of EA on the expression of miR-9
in the cerebral cortex
In order to investigate the effect of EA on the
miR-9, the expression of miR-9 in the peri-infarct cerebral cortex
was assessed using qPCR. Fig. 3
shows that the relative expression of the miR-9 in MCAO, MCAO + EA,
EA + DMSO and EA + miR-9 inhibitors groups were significantly
decreased compared with the Sham group 72 h after MCAO and
reperfusion (P<0.05). In the MCAO + EA and EA + DMSO groups,
however, a significant increase was observed compared with the MCAO
group (P<0.05). In addition, the expression of miR-9 in the EA +
miR-9 inhibitors group was found to be lower compared with that in
the MCAO + EA and EA + DMSO groups (P<0.05).
Effect of EA on the NF-κB signaling
pathway
To investigate the mechanism of the
anti-inflammatory effect of EA, the localization and expression of
NF-κB signaling pathway-associated factors in the peri-infarct
cortex was evaluated. The NF-κB p65 subunit was visualized using
immunofluorescent staining, and the cells were counterstained with
DAPI. NF-κB nuclear translocation was recognized by the
colocalization of the p65 subunit with DAPI. The cerebral ischemic
injury resulted in the nuclear translocation of the NF-κB p65
subunit in the MCAO group, which was not observed in the sham
operation group. However, EA inhibited the NF-κB nuclear
translocation and reduced the number of NF-κB p65-positive cells,
whereas of NF-κB p65-positive cells decreased in the EA + miR-9
inhibitors group, as compared with the EA and EA + DMSO groups
(Fig. 4A). The levels of
expression of NF-κB p65, TNF-α and IL-1β in the EA group were
significantly decreased compared with the Sham group (P<0.05).
By contrast, the IκBα expression levels were increased in the EA
group compared with those in the MCAO group (P<0.05). In the EA
+ miR-9 inhibitors group, the expression levels of NF-κB p65 not
only increased, but also exceeded those of the MCAO group,
suggesting that the administration of miR-9 inhibitors
significantly promoted the expression of NF-κB p65. Furthermore,
the differences in IκBα expression among the EA, EA + DMSO and EA +
miR-9 inhibitors groups were found to be statistically significant
(P>0.05), suggesting that miR-9 inhibitors did not alter the
expression of NF-κB upstream-related protein IκBα.
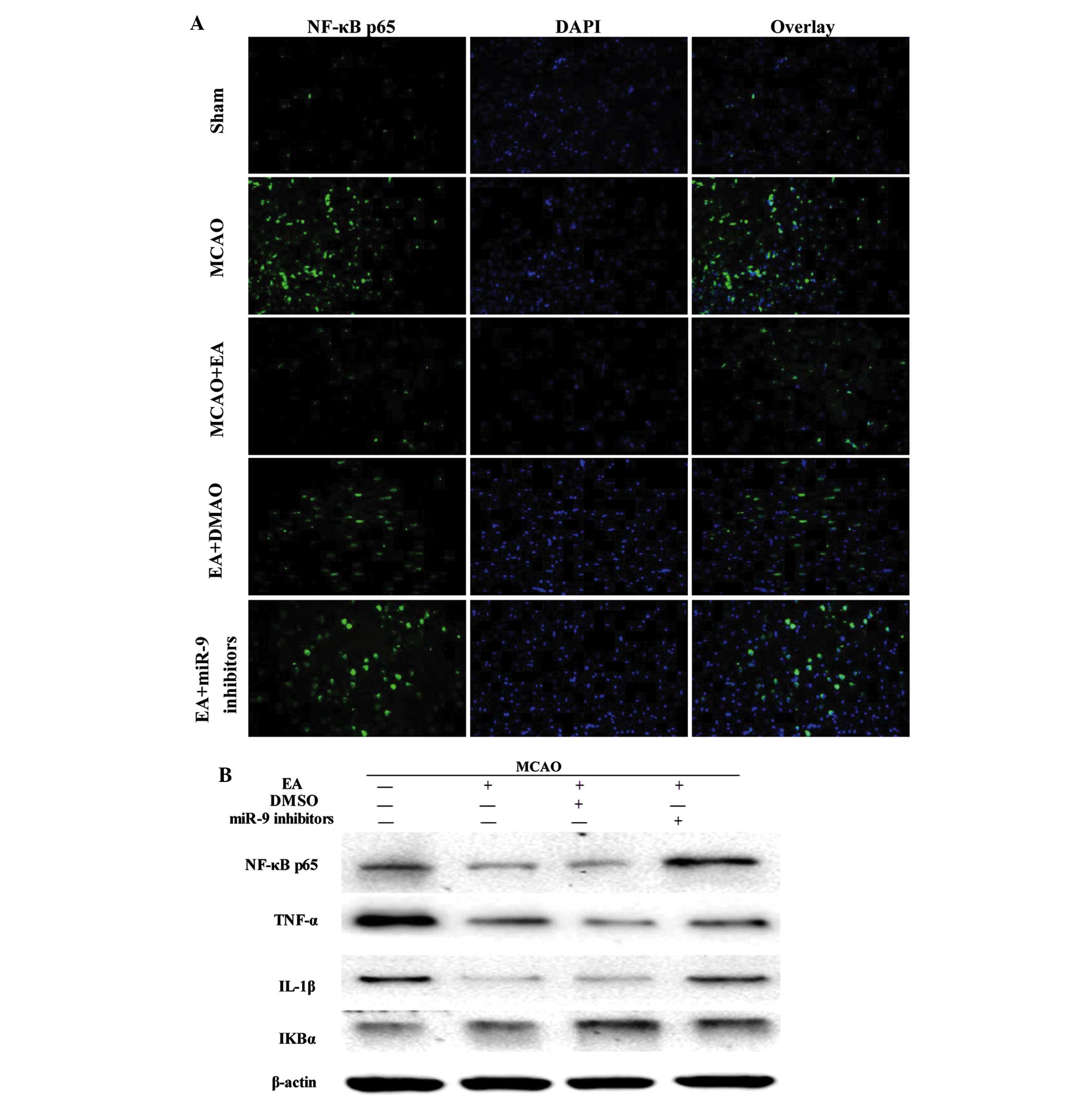 | Figure 4Effect of EA on NF-κB signaling
pathway-associated factors. (A) Evaluation of NF-κB p65 (green),
DAPI (blue) and the colocalization positive cells in peri-infarct
cortical tissue of the Sham, MCAO, MCAO + EA, EA + DMSO and EA +
miR-9 inhibitor groups (n=4). (B) Western blot analysis of the
protein expression levels of NF-κB p65, TNF-α, IL-1β and IκBα in
the peri-infarct cortical tissue of the MCAO, MCAO + EA, EA + DMSO
and EA + miR-9 inhibitors groups. (C) Bar graph showing the fold
change of NF-κB p65, TNF-α, IL-1β and IκBα in each group (n=4)
(*P<0.05, vs. MCAO group; #P<0.05, vs.
MCAO + EA and EA + DMSO groups). EA, electroacupuncture; DAPI,
4′,6-diamidino-2-phenylindole; MCAO, middle cerebral artery
occlusion; DMSO, dimethyl sulfoxide; miR-9, microRNA-9; NF-κB,
nuclear factor-κB; TNF-α, tumor necrosis factor-α; IL-1β,
interleukin-1β; IκBα, inhibitor of κBα. |
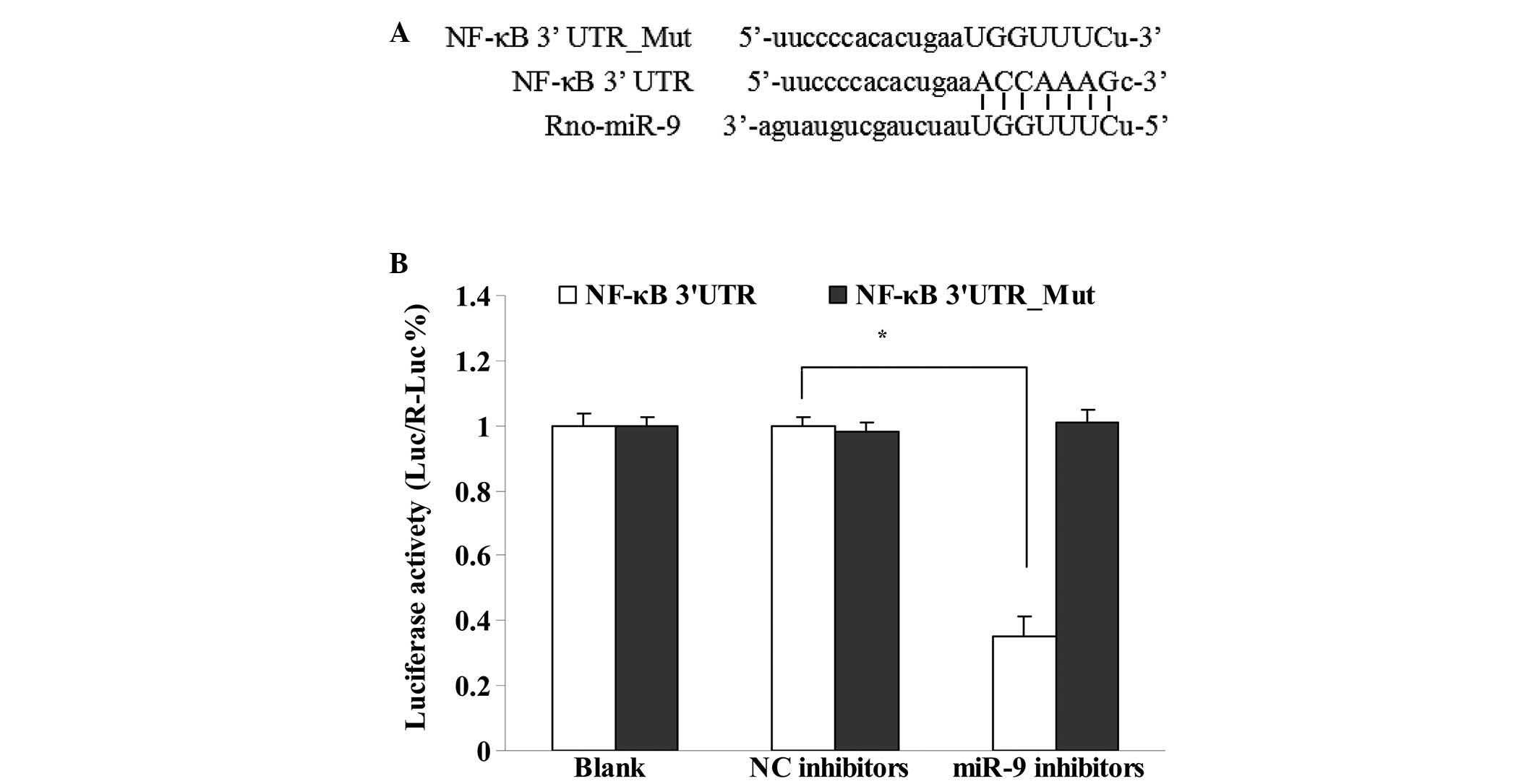
Interaction of miR-9 and NF-κB
The target genes of miR-9 were predicted with an
online algorithm using miRWalk database. Fig. 5A shows that the site where miR-9
binds to NF-κB was predicted and identified as ACCAAAG. The
interaction of miR-9 and NF-κB was observed using a dual-luciferase
reporter vector assay (Fig. 5B).
The results showed that miR-9 bound to 3′UTR of the NK-κB gene
(P<0.05), however, not to the mutant 3′UTR (P>0.05).
Discussion
For >3,000 years, practitioners in China have
used acupuncture to treat various diseases, including stroke.
Acupuncture is widely used for the improvement of motor, sensation,
speech and other neurological functions in patients with stroke.
Compared with other conventional interventions, acupuncture is
relatively simple, inexpensive and safe. For those reasons,
acupuncture is not only widely accepted by Chinese patients, but
also increasingly practiced in certain Western countries (18). Electrical stimulation was added to
traditional acupuncture and termed electro-acupuncture (EA), which
quantified the inserting dose and enhanced treatment efficiency
(19). A number of indices that
evaluate the quality of life of stroke patients have suggested that
EA treatment may improve patient self care (20,21).
Studies have reported that the application of EA treatment on Quchi
(LI11) and Zusanli (ST36) acupoints results in a decrease in the
neurological deficit scores of patients with hemiplegia (22). Our previous animal studies on the
underlying mechanism of EA treatment on Quchi (LI11) and Zusanli
(ST36) demonstrated that EA stimulation of those particular
acupoints considerably improved the neurological deficit scores of
rats, as well as reduce ischemic infarct volume-mediated
anti-inflammatory pathway (12,14).
The present study demonstrated that EA treatment on Quchi (LI11)
and Zusanli (ST36) acupoints clearly decreased the infarct volume
of MCAO rats and reduced alleviated cerebral neurocyte interstitial
edema and inflammatory invasion at 72 h after MCAO and
reperfusion.
It has been reported that NF-κB is involved in
regulating ischemic stroke injury (23). The classical pathway of NF-κB
activation is through a heterodimer of p50 and p65 of two subunits
of NF-κB. NF-κB dimers are maintained in the inactive state by a
family of inhibitors called IκB. Receptor signaling leads to the
activation of a multisubunit IKK complex, which phosphorylates IκB
on two key serine residues. Phosphorylation of IκB marks it for
degradation by the ubiquitin pathway, so that the NF-κB dimer is
liberated to translocate to the nucleus, bind DNA and activate
transcription. It has been shown that EA causes the downregulation
of NF-κB signaling pathway-assocaited factors, NF-κB p65, TNF-α,
IL-1β and MCP-1, and the upregulation of IκBα in the cortex,
hippocampus and corpus striatum of MCAO rats (12,24,25).
Consistent with previous reports, the results of the present study
demonstrated that EA treatment on Quchi (LI11) and Zusanli (ST36)
acupoints reduced the expression of NF-κB p65 and inhibited nuclear
translocation of NF-κB p65 in the cerebral peri-infarct cortex of
rats with ischemia-reperfusion injuries.
miRNA is a small, non-coding RNA molecule involved
in the growth and development of various diseases, including stroke
(26). By recognizing the 3′UTR of
target mRNA, mature miRNA degrades the target gene or inhibits its
translation, therefore, regulating gene expression and protein
synthesis (27). The
miRNA-mediated immune response is achieved by a highly complex
regulatory network in the presence of stimuli and pathogens
(3). For example, miR-146a,
miR-21, miR-221, miR-579, miR-125b and miR-155 are involved in the
negative regulation of inflammatory reactions (28–30).
Few studies have focused on the involvement of miRNA in the acute
or sub-acute phase of inflammatory reactions induced by ischemic
strokes. It has been reported that the level of miR-155 was
significantly downregulated in the brain and blood following
inflammatory damage (31,32). A previous study revealed that miR-9
is crucial in the NF-κB signaling pathway, and the activation of
microglia by targeting MCPIP1, the latter of which is associated
with inflammatory reactions following cerebral ischemia (5). miR-9 has been shown to regulate the
expression of peroxisome proliferator-activated receptor δ in human
monocytes during the inflammatory response (33). miR-9 is also induced by the
proinflammatory cytokines, TNF-α and IL-1β (34).
miR-9 is a tissue-specific miRNA, which is
predominantly expressed in the region close to the
midbrain-hindbrain boundary and is involved in normal brain
development (35,36). The present study found that miR-9
was associated with inflammatory response and a significant
reduction of miR-9 expression in the peri-infarct cortex following
ischemia-reperfusion injury; however, the expression of miR-9 was
found to be upregulated following EA treatment on Quchi (LI11) and
Zusanli (ST36) acupoints. It has been shown that NF-κB is a direct
target of miR-9 in tumors (6,37).
The present study demonstrated that miR-9 negatively regulated the
expression of NF-κB in primary cortical neuronal cells.
Furthermore, miR-9 inhibitors suppressed EA-alleviated cerebral
inflammation and the expression of NF-κB signaling pathway
downstream-associated factors, NF-κB p65, TNF-α, IL-1β, without
altering the level of NF-κB upstream-associated protein, IκBα.
These results demonstrated that EA treatment on Quchi (LI11) and
Zusanli (ST36) acupoints targeted the miR-9-NF-κB downstream
pathway following ischemic stroke, therefore suggesting a cerebral
protective effect of EA.
In conclusion, miR-9 has been shown to be involved
in the inflammatory reactions induced by cerebral
ischemia-reperfusion injuries by binding to NF-κB. miR-9 may serve
as a potential therapeutic target for the treatment of cerebral
ischemic injuries. EA treatment on Quchi (LI11) and Zusanli (ST36)
acupoints regulated the miR-9-mediated NF-κB signaling pathway and
reduced the secretion of the proinflammatory cytokines, TNF-α and
IL-1β. It is possible, however, that miR-9 is involved in multiple
signaling pathways, which are associated with inflammatory injury
following cerebral ischemia, however, this requires further
elucidation.
Acknowledgments
The present study was supported by the Natural
Science Foundation of China (nos. 81403450, 81373778 and 81273835),
and by the Key Subjects of Fujian university of Traditional Chinese
Medicine (no. X2014069-xueke).
References
|
1
|
Cullen BR: Transcription and processing of
human microRNA precursors. Mol Cell. 16:861–865. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Doench JG and Sharp PA: Specificity of
microRNA target selection in translational repression. Genes Dev.
18:504–511. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
O'Connell RM, Rao DS and Baltimore D:
MicroRNA regulation of inflammatory responses. Annu Rev Immunol.
30:295–312. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Akerblom M, Sachdeva R, Quintino L,
Wettergren EE, Chapman KZ, Manfre G, Lindvall O, Lundberg C and
Jakobsson J: Visualization and genetic modification of resident
brain microglia using lentiviral vectors regulated by microRNA-9.
Nat Commun. 4:17702013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yao H, Ma R, Yang L, Hu G, Chen X, Duan M,
Kook Y, Niu F, Liao K, Fu M, et al: MiR-9 promotes microglial
activation by targeting MCPIP1. Nat Commun. 5:43862014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Guo LM, Pu Y, Han Z, Liu T, Li YX, Liu M,
Li X and Tang H: MicroRNA-9 inhibits ovarian cancer cell growth
through regulation of NF-kappaB1. FEBS J. 276:5537–5546. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang J, Gu Z, Ni P, Qiao Y, Chen C, Liu X,
Lin J, Chen N and Fan Q: NF-kappaB P50/P65 hetero-dimer mediates
differential regulation of CD166/ALCAM expression via interaction
with micoRNA-9 after serum deprivation, providing evidence for a
novel negative auto-regulatory loop. Nucleic Acids Res.
39:6440–6455. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lukiw WJ: NF-κB-regulated micro RNAs
(miRNAs) in primary human brain cells. Exp Neurol. 235:484–490.
2012. View Article : Google Scholar :
|
|
9
|
Liu XS, Chopp M, Zhang RL, Tao T, Wang XL,
Kassis H, Hozeska-Solgot A, Zhang L, Chen C and Zhang ZG: MicroRNA
profiling in subventricular zone after stroke: MiR-124a regulates
proliferation of neural progenitor cells through Notch signaling
pathway. PLoS One. 6:e234612011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu Y, Zhang J, Han R, Liu H, Sun D and
Liu X: Downregulation of serum brain specific microRNA is
associated with inflammation and infarct volume in acute ischemic
stroke. J Clin Neurosci. 22:291–295. 2015. View Article : Google Scholar
|
|
11
|
Chen W, Gu HW, Ma WP, Li QS, Yu Q, Liu XQ,
Liu SH, Li WH, Liu HL and Dai MT: Multicentral randomized
controlled study on effects of acupuncture at Zusanli (ST 36) and
Xuanzhong (GB 39) on cerebrovascular function in the patient of
ischemic stroke. Zhongguo Zhen Jiu. 26:851–853. 2006.In
Chinese.
|
|
12
|
Lan L, Tao J, Chen A, Xie G, Huang J, Lin
J, Peng J and Chen L: Electroacupuncture exerts anti-inflammatory
effects in cerebral ischemia-reperfusion injured rats via
suppression of the TLR4/NF-κB pathway. Int J Mol Med. 31:75–80.
2013.
|
|
13
|
Jin Z, Liang J, Wang J and Kolattukudy PE:
Delayed brain ischemia tolerance induced by electroacupuncture
pretreatment is mediated via MCP-induced protein 1. J
Neuroinflammation. 10:632013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xue X, You Y, Tao J, Ye X, Huang J, Yang
S, Lin Z, Hong Z, Peng J and Chen L: Electro-acupuncture at points
of Zusanli and Quchi exerts anti-apoptotic effect through the
modulation of PI3K/Akt signaling pathway. Neurosci Lett. 558:14–19.
2014. View Article : Google Scholar
|
|
15
|
Lu T, Song QH, Xu RM, Guo YH, Wang F, Hu
JP, Wang Y and Zhang LY: Dance combined with magnetic pulse
stimulates the ability of walk and balance in elder people. Int J
Clin Exp Med. 8:4381–4386. 2015.PubMed/NCBI
|
|
16
|
Zhao H, Wang J, Gao L, Wang R, Liu X, Gao
Z, Tao Z, Xu C, Song J, Ji X and Luo Y: MiRNA-424 protects against
permanent focal cerebral ischemia injury in mice involving
suppressing microglia activation. Stroke. 44:1706–1713. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Longa EZ, Weinstein PR, Carlson S and
Cummins R: Reversible middle cerebral artery occlusion without
craniectomy in rats. Stroke. 20:84–91. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhuang L, He J, Zhuang X and Lu L: Quality
of reporting on randomized controlled trials of acupuncture for
stroke rehabilitation. BMC Complement Altern Med. 14:1512014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chang L, He PL, Zhou ZZ and Li YH:
Efficacy observation of dysphagia after acute stroke treated with
acupuncture and functional electric stimulation. Zhongguo Zhen Jiu.
34:737–740. 2014.In Chinese. PubMed/NCBI
|
|
20
|
Sze FK, Wong E, Or KK, Lau J and Woo J:
Does acupuncture improve motor recovery after stroke? A
meta-analysis of randomized controlled trials. Stroke.
33:2604–2619. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shen PF, Kong L, Ni LW, Guo HL, Yang S,
Zhang LL, Zhang ZL, Guo JK, Xiong J, Zhen Z, et al: Acupuncture
intervention in ischemic stroke: A randomized controlled
prospective study. Am J Chin Med. 40:685–693. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang H, Xie Y, Zhang Q, Xu N, Zhong H,
Dong H, Liu L, Jiang T, Wang Q and Xiong L: Transcutaneous electric
acupoint stimulation reduces intra-operative remifentanil
consumption and alleviates postoperative side-effects in patients
undergoing sinusotomy: A prospective, randomized,
placebo-controlled trial. Br J Anaesth. 112:1075–1082. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Harari OA and Liao JK: NF-κB and innate
immunity in ischemic stroke. Ann N Y Acad Sci. 1207:32–40. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Feng X, Yang S, Liu J, Huang J, Peng J,
Lin J, Tao J and Chen L: Electroacupuncture ameliorates cognitive
impairment through inhibition of NF-κB-mediated neuronal cell
apoptosis in cerebral ischemia-reperfusion injured rats. Mol Med
Rep. 7:1516–1522. 2013.PubMed/NCBI
|
|
25
|
Wang ZK, Ni GX, Liu K, Xiao ZX, Yang BW,
Wang J and Wang S: Research on the changes of IL-1 receptor and
TNF-alpha receptor in rats with cerebral ischemia reperfusion and
the chronergy of acupuncture intervention. Zhongguo Zhen Jiu.
32:1012–1018. 2012.In Chinese. PubMed/NCBI
|
|
26
|
Alvarez-Garcia I and Miska EA: MicroRNA
functions in animal development and human disease. Development.
132:4653–4662. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ambros V: MicroRNA pathways in flies and
worms: Growth, death, fat, stress and timing. Cell. 113:673–676.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Quinn SR and O'Neill LA: A trio of
microRNAs that control Toll-like receptor signalling. Int Immunol.
23:421–425. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sheedy FJ, Palsson-McDermott E, Hennessy
EJ, Martin C, O'Leary JJ, Ruan Q, Johnson DS, Chen Y and O'Neill
LA: Negative regulation of TLR4 via targeting of the
proinflammatory tumor suppressor PDCD4 by the microRNA miR-21. Nat
Immunol. 11:141–147. 2010. View
Article : Google Scholar
|
|
30
|
El Gazzar M and McCall CE: MicroRNAs
distinguish translational from transcriptional silencing during
endotoxin tolerance. J Biol Chem. 285:20940–20951. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tili E, Michaille JJ, Cimino A, Costinean
S, Dumitru CD, Adair B, Fabbri M, Alder H, Liu CG, Calin GA and
Croce CM: Modulation of miR-155 and miR-125b levels following
lipopolysaccharide/TNF-alpha stimulation and their possible roles
in regulating the response to endotoxin shock. J Immunol.
179:5082–5089. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu DZ, Tian Y, Ander BP, Xu H, Stamova
BS, Zhan X, Turner RJ, Jickling G and Sharp FR: Brain and blood
microRNA expression profiling of ischemic stroke, intracerebral
hemorrhage and kainate seizures. J Cereb Blood Flow Metab.
30:92–101. 2010. View Article : Google Scholar
|
|
33
|
Thulin P, Wei T, Werngren O, Cheung L,
Fisher RM, Grandér D, Corcoran M and Ehrenborg E: MicroRNA-9
regulates the expression of peroxisome proliferator-activated
receptor δ in human monocytes during the inflammatory response. Int
J Mol Med. 31:1003–1010. 2013.PubMed/NCBI
|
|
34
|
Bazzoni F, Rossato M, Fabbri M, Gaudiosi
D, Mirolo M, Mori L, Tamassia N, Mantovani A, Cassatella MA and
Locati M: Induction and regulatory function of miR-9 in human
monocytes and neutrophils exposed to proinflammatory signals. Proc
Natl Acad Sci USA. 106:5282–5287. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Delaloy C and Gao FB: MicroRNA-9
multitasking near organizing centers. Nat Neurosci. 11:625–626.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Coolen M and Bally-Cuif L: MicroRNAs in
brain development and physiology. Curr Opin Neurobiol. 19:461–470.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wan HY, Guo LM, Liu T, Liu M, Li X and
Tang H: Regulation of the transcription factor NF-kappaB1 by
microRNA-9 in human gastric adenocarcinoma. Mol Cancer. 9:162010.
View Article : Google Scholar : PubMed/NCBI
|