Introduction
Gliomas are solid tumors of the central nervous
system, which represent >30% of central nervous system tumors
and account for 70% of malignant primary brain tumors (1,2). It
is estimated that there are 20,000 newly diagnosed cases each year
in the USA (2). Histologically,
gliomas are divided into oligodendrogliomas, astrocytomas,
anaplastic astrocytomas, glioblastomas, and numerous other subtypes
(3). Malignant gliomas, including
anaplastic astrocytomas and glioblastoma multiforme are the most
common types of primary brain tumor, accounting for ~60% of gliomas
(4,5). The standard therapeutic strategies
for glioma treatment, including surgical resection, and adjuvant
radiotherapy and chemotherapy, have improved the 5-year survival
rates from 2 to 10%; however, the prognosis for patients diagnosed
at an advanced stage remains poor (6,7). It
has previously been demonstrated that dysregulated tumor suppressor
genes or oncogenes are closely associated with the development and
progression of glioma (8).
Therefore, more effective therapeutic strategies and targets for
the treatment of glioma should be developed, alongside an increased
understanding of the molecular pathogenesis of glioma.
Previous studies have indicated that microRNAs
(miRNAs) are important in numerous aspects of human cancer
(9–12). miRNAs are a class of non-coding RNA
molecules (length, ~18–25 nucleotides), which induce mRNA
degradation and suppress protein translation, predominantly by
binding to the seed sequences in the 3′-untranslated region of
mRNAs (13,14). miRNAs influence various
physiological and pathological processes, including proliferation,
differentiation, apoptosis, metabolism, migration and invasion
(11,15).
It has previously been estimated that miRNAs are
involved in almost every biological process by targeting ~30% of
protein-coding genes in the human genome (16). Since the first miRNA expression
profiles were obtained from a range of cancer types using
bead-based flow cytometry, hundreds of evolutionarily conserved
miRNAs have been identified in different species, including plants,
animals and viruses (10,17). There is also an increasing amount
of evidence suggesting that miRNAs may function as tumor
suppressors and oncogenes (18).
Downregulated miRNAs in cancer may normally function as tumor
suppressor genes and inhibit cancer by regulating oncogenes.
Conversely, upregulated miRNAs in cancer may function as oncogenes
by negatively regulating tumor suppressors (19). Therefore, the identification of
miRNA target genes is critical to understand the function of miRNAs
in cancer development and progression. It has also been suggested
that miRNAs may be a target for cancer therapy.
The expression levels of miR-133a have been reported
to be downregulated in numerous human malignancies (20–28);
however, to the best of our knowledge, the role of miR-133a in
human glioma has not yet been investigated. In the present study,
the expression levels and effects of miRNA-133a on human glioma
were examined. The results of the present study demonstrated that
miR-133a was downregulated in human glioma tumor tissues compared
with their normal adjacent tissues (NATs), and miR-133a was able to
suppress cell proliferation, migration and invasion by directly
targeting matrix metallopeptidase 9 (MMP9). These findings may have
therapeutic implications, and may be useful for the development of
novel treatments for human glioma.
Materials and methods
Clinical specimens
The present study was approved by the ethics
committee of The First Affiliated Hospital of Chongqing Medical
University (Chongqing, China) and informed consent was obtained
from all patients. The glioma and NAT specimens for reverse
transcription-quantitative polymerase chain reaction (RT-qPCR) were
obtained from 50 patients (32 male and 28 female; age range, 37–84
years) who had undergone surgery at Daping Hospital (Chongqing,
China) between January 2010 and December 2014. Of the 50 gliomas,
22 were classified as low-grade [World Health Organization (WHO) I
and WHO II] gliomas and 28 were classified as high-grade (WHO III
and WHO IV) gliomas (29). None of
the patients had received chemotherapy, immunotherapy or
radiotherapy prior to specimen collection. Tissues were snap-frozen
in liquid nitrogen and stored at −80°C.
Cell culture and transfection
The U251 and U87 human glioma cell lines were
obtained from the Shanghai Institute of Biochemistry and Cell
Biology (Shanghai, China). U251 and U87 cells were cultured in
Dulbecco's modified Eagle's medium (DMEM) supplemented with 10%
heat-inactivated fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) at 37°C in a 5% CO2
cell incubator.
Mature miR-133a mimics, negative control (NC) miRNA
mimics and the luciferase reporter plasmid were designed and
synthesized by Shanghai GenePharma Co., Ltd. (Shanghai, China). The
sequence of miR-133a mimics was 5′-UUU GGU CCC CUU CAA CCA GCUG-3′.
The sequence of NC mimics was 5′-UUC UCC GAA CGU GUC ACG UTT-3′.
Transient transfection was performed using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol.
RNA isolation and RT-qPCR analysis
The tissues were homogenized and the total RNA was
extracted from tissues with TRIzol® (Invitrogen; Thermo
Fisher Scientific, Inc.), according to the manufacturer's protocol.
The concentration and purity of all RNA samples were measured using
an ND-2000 spectrophotometer (NanoDrop Technologies; Thermo Fisher
Scientific, Inc., Wilmington, DE, USA). RT-qPCR analysis was
performed using One Step SYBR® PrimeScript™ miRNA RT-PCR
kit (Takara Bio, Inc., Otsu, Japan) in a CFX96™ Real-Time system
and a C1000™ Thermal Cycler (both Bio-Rad Laboratories, Inc.,
Hercules, CA, USA) according to the manufacturers' protocols. The
primers were obtained from Guangzhou RiboBio Co., Ltd. (Guangzhou,
China) Each reaction was performed in a final volume of 25
µl. The cycling conditions were as follows: 42°C for 5 min;
95°C for 10 sec; and 40 cycles of 95°C for 5 sec, 55°C for 30 sec
and 70°C for 30 sec. Each sample was analyzed in triplicate and the
data were normalized using the endogenous U6 small nuclear RNA. The
relative expression of miR-133a was analyzed by use of the
2−ΔΔCq method (30).
Cell proliferation assay
Cell proliferation was determined using Cell
Counting kit-8 (CCK-8) detection kit (Dojindo Molecular
Technologies, Inc., Kumamoto, Japan). The transfected cells (with
miR-133a mimics or NC) were seeded at a density of 3×103
cells/well into 96-well plates. Cell proliferation was assayed
every 24 h for 5 days. Briefly, 10 µl CCK-8 solution was
added to each well and incubated at 37°C for 2 h. Absorbance was
measured at 450 nm in an ELISA reader (Elx800; Bio-Rad
Laboratories, Inc.). The suppression rate of proliferation was
calculated using the following formula: Suppression rate = (1 -
ODmiR-133a/ODmiR-NC) × 100%. OD indicates
optical density. All the experiments were performed in
triplicate.
Cell migration and invasion assays
Migration and invasion of glioma cell lines were
assessed using Transwell chambers with an 8-µm pore
polycarbonate membrane (Costar; Corning Incorporated, Corning, NY,
USA). For the migration assay, 5×104 transfected cells
(with miR-133a mimics and NC) were harvested and suspended in 200
µl DMEM with 0.1% FBS. These cells were placed into the
upper chamber. A volume of 0.5 ml DMEM with 20% FBS was then added
to the lower chamber as a chemoattractant. For the invasion assay,
5×104 transfected cells were placed into the upper
chamber, which was coated with Matrigel (BD Biosciences, San Jose,
CA, USA). A volume of 0.5 ml DMEM with 20% FBS was then added to
the lower chamber as a chemoattractant. Cells were incubated for a
further 12 h for the migration assay and 24 h for the invasion
assay. In the two assays, the invaded cells on the lower surface
were fixed with 100% methanol (Shanghai Macklin Biochemical Co.,
Ltd., Shanghai, China), stained with 0.5% crystal violet (Beyotime
Institute of Biotechnology, Haimen, China), and were then counted
under an inverted microscope (CKX41; Olympus Corporation, Tokyo,
Japan) to calculate the relative numbers (magnification, x100).
Each experiment was repeated at least three times.
Western blotting
Mouse anti-human monoclonal primary antibodies
against MMP9 (dilution, 1:1,000; sc-21733) and β-actin (dilution,
1:1,000; sc-47778), which were obtained from Santa Cruz
Biotechnology, Inc. (Dallas, TX, USA), were used in the present
study. Following 72 h of transfection with miR-133a or NC, the
cells were washed twice with ice-cold phosphate-buffered saline and
lysed with radioimmunoprecipitation assay lysis buffer [50 mM
Tris-HCl (pH 7.4), 1% NP-40, 0.25% Na-deoxycholate, 150 mM NaCl, 1
mM EDTA, 1 mM phenylmethylsulfonyl fluoride, 1 µg/ml
aprotinin, 1 µg/ml leupeptin, 1 µg/ml pepstatin, 1 mM
Na3VO4 and 1 mM NaF]. The protein
concentration was measured using a bicinchoninic acid assay kit
(Beyotime Institute of Biotechnology). Equal quantities of protein
(20 µg) were separated by 10% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis and electrotransferred
to polyvinylidene difluoride membranes (Beyotime Institute of
Biotechnology). For western blotting, the membranes were blocked
with 5% skimmed milk and incubated with the primary antibodies at
dilutions specified by the manufacturer's protocols at 4°C
overnight. The membranes were washed with Tris-buffered saline with
0.5% Tween 20 (TBST; Beyotime Institute of Biotechnology) and then
incubated with a goat anti-mouse horseradish peroxidase-conjugated
secondary antibody (1:10,000; Santa Cruz Biotechnology, Inc.;
sc-2005) in TBST. The protein bands were developed with enhanced
chemiluminescence reagents (Pierce Biotechnology, Inc., Rockford,
IL, USA) and imaged with a FluorChem imaging system (version 4.1.0;
Alpha Innotec, San Leandro, CA, USA).
Luciferase assay
The U251 and U87 cells were transfected with 0.5
µg MMP9-3′-UTR-Wild type or MMP9-3′-UTR-Mutant, and 40 nmol
miR-133a mimics or NC in a 12-well plate using
Lipofectamine® 2000, according to the manufacturer's
protocol. The activities of the firefly and Renilla
luciferases in cell lysates were determined using the
Dual-Luciferase Reporter assay system (Promega Corporation,
Madison, WI, USA) 48 h post-transfection. The firefly luciferase
activity was normalized to the Renilla luciferase activity
for each transfected well. Each reporter plasmid was transfected at
least three times (on different days) and each sample was assayed
in triplicate.
Statistical analysis
Data are presented as the mean ± standard deviation.
Data were analyzed using Student's t-test and analysis of variance
in Stata 10.0 (StataCorp LP, College Station, TX, USA). P<0.05
was considered to indicate a statistically significant
difference.
Results
miR-133a is downregulated in glioma tumor
tissues compared with NATs
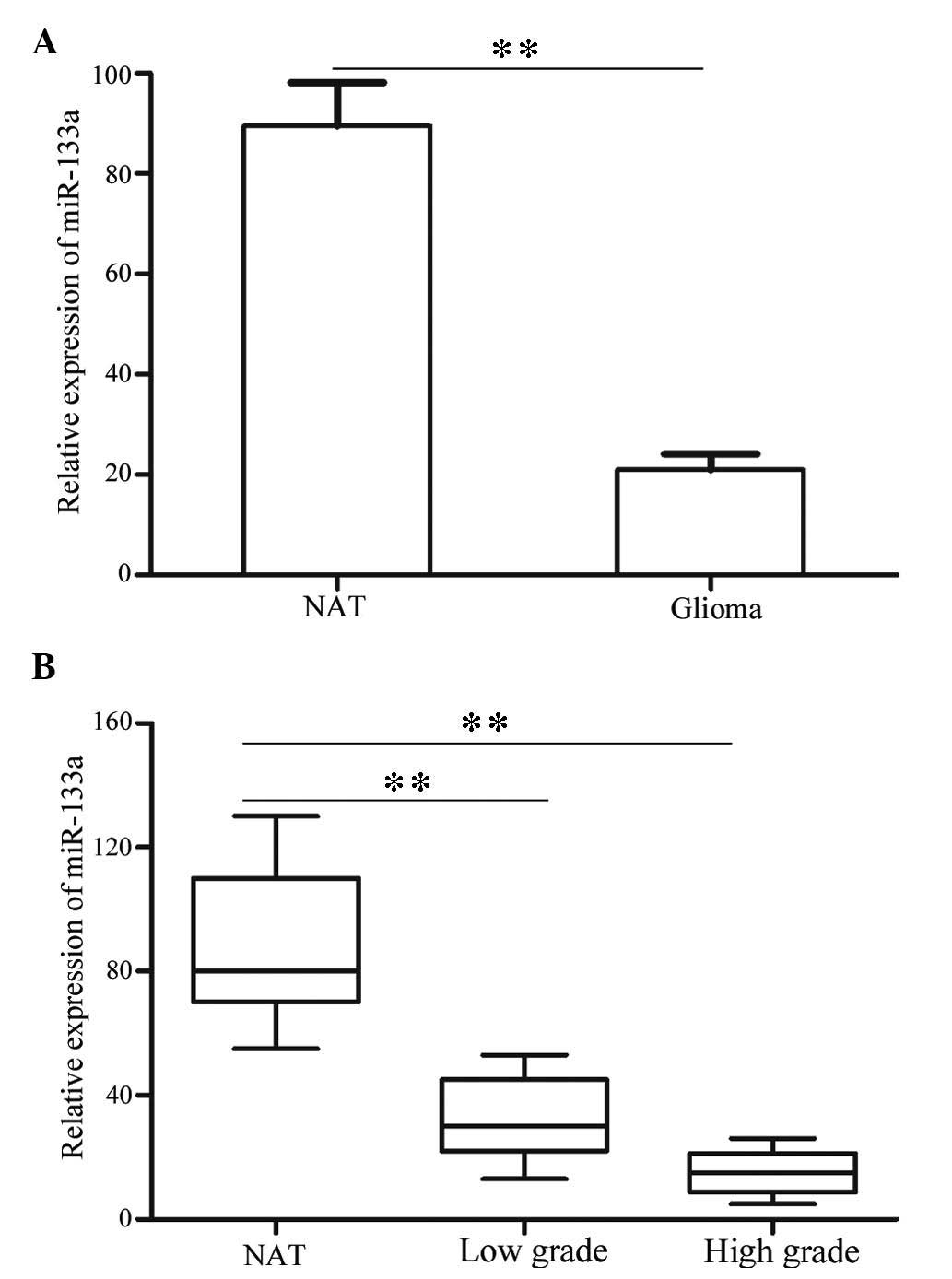
miR-133a expression levels were detected in all
glioma tissues and NATs using RT-qPCR. As presented in Fig. 1A, miR-133a was significantly
decreased in glioma tissues compared with NATs (P<0.01). These
results indicate that miR-133a may have an important role in
glioma.
Furthermore, the present study investigated whether
the expression levels of miR-133a were associated with tumor grade
and stage. The statistical analysis demonstrated that the
expression levels of miR-133a were significantly lower in
high-grade glioma (WHO III and WHO IV) compared with in low-grade
glioma (WHO I and WHO II; Fig. 1B;
P<0.01).
miR-133a suppresses cell proliferation in
U251 and U87 glioma cells
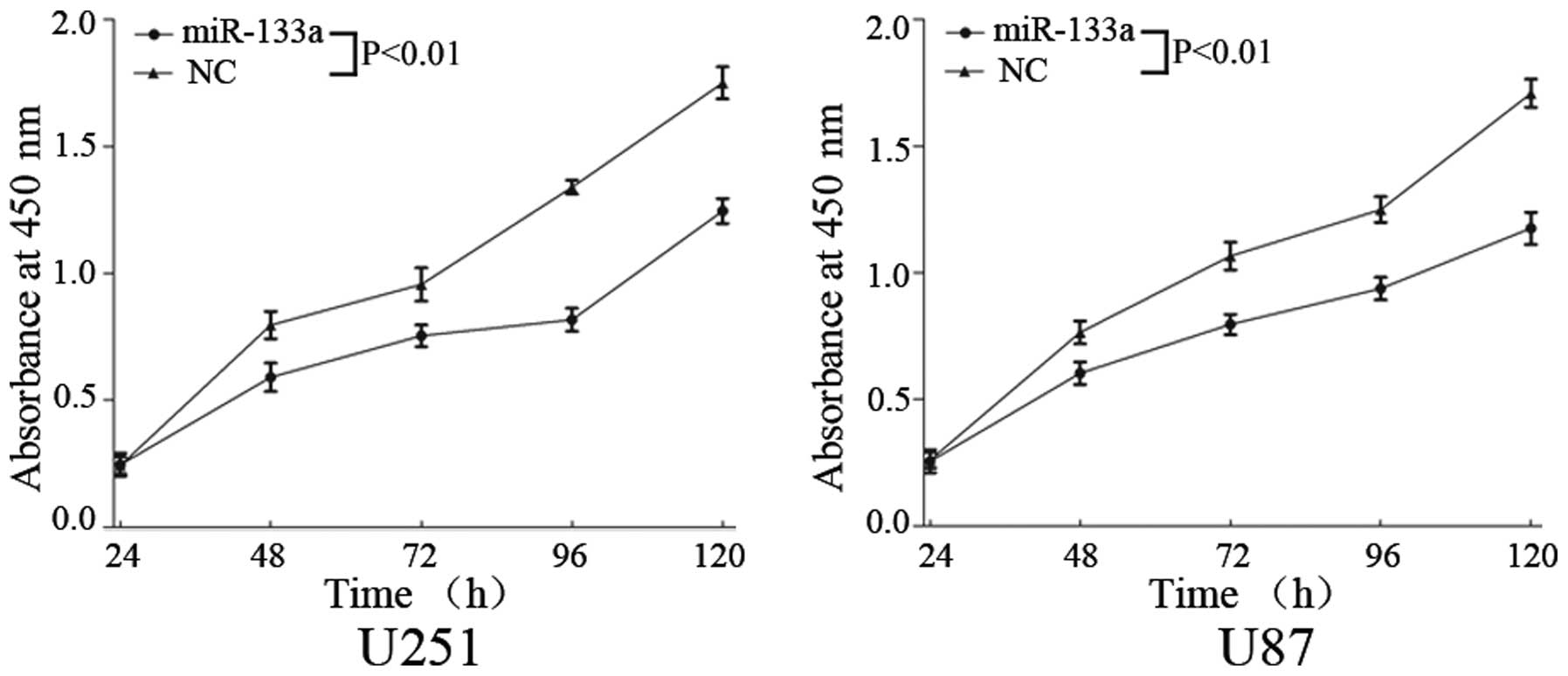
To investigate the effects of miR-133a on glioma
cell proliferation, the CCK-8 assay kit was used. Upregulation of
miR-133a markedly inhibited cell proliferation in glioma U251 and
U87 cells (Fig. 2; P<0.01).
These results indicate that miR-133a may function as a tumor
suppressor in human glioma.
miR-133a suppresses cell migration and
invasion in U251 and U87 glioma cells
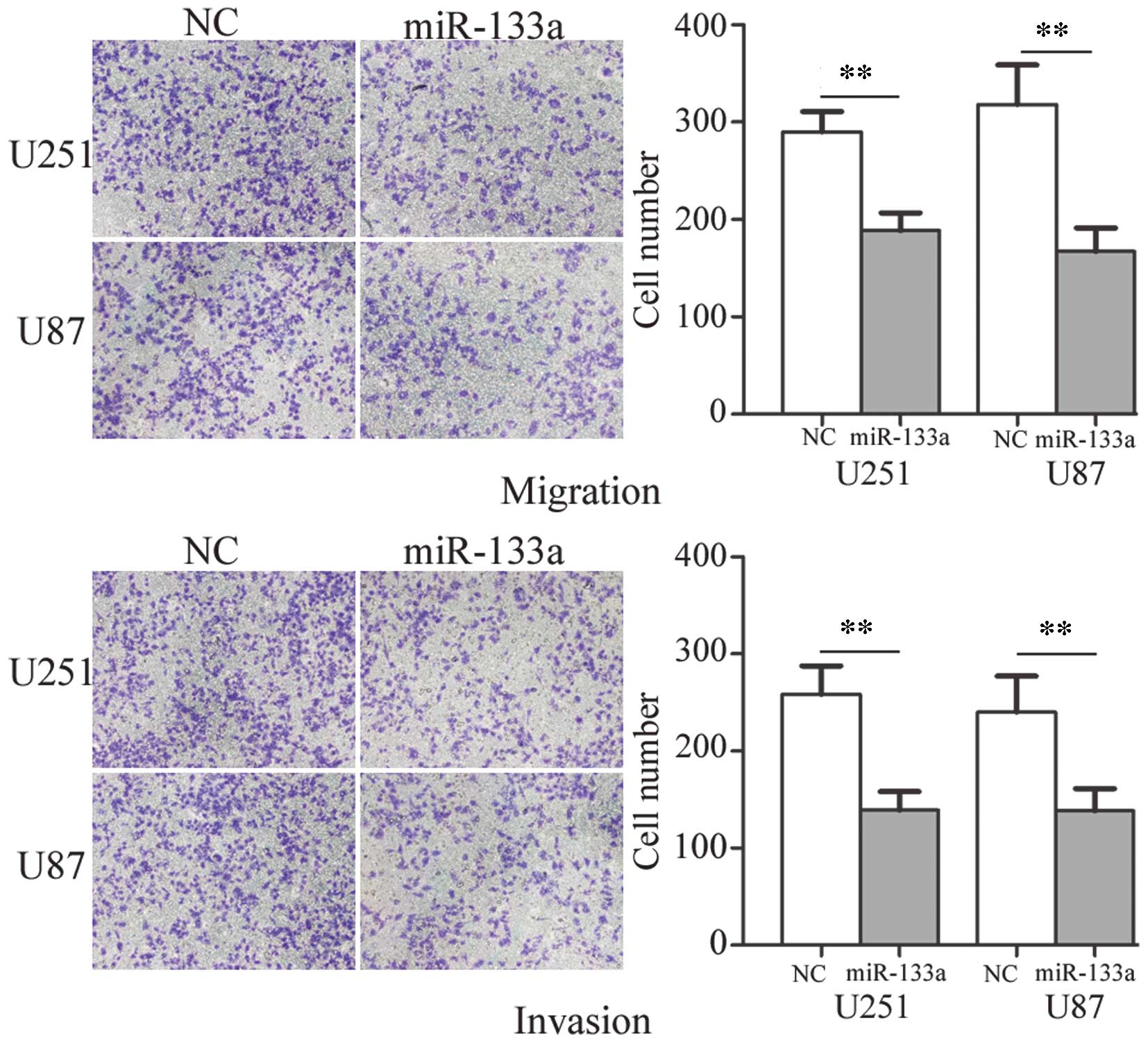
To determine the influence of miR-133a on tumor cell
migration and invasion, the Transwell assay was used. As shown in
Fig. 3, the migratory and invasive
ability of U251 and U87 glioma cells transfected with miR-133a was
markedly decreased compared with those transfected with NC
(P<0.01). These results indicate that miR-133a may decrease the
migration and invasion of glioma cells.
MMP9 is a direct target gene of miR-133a
in glioma cells
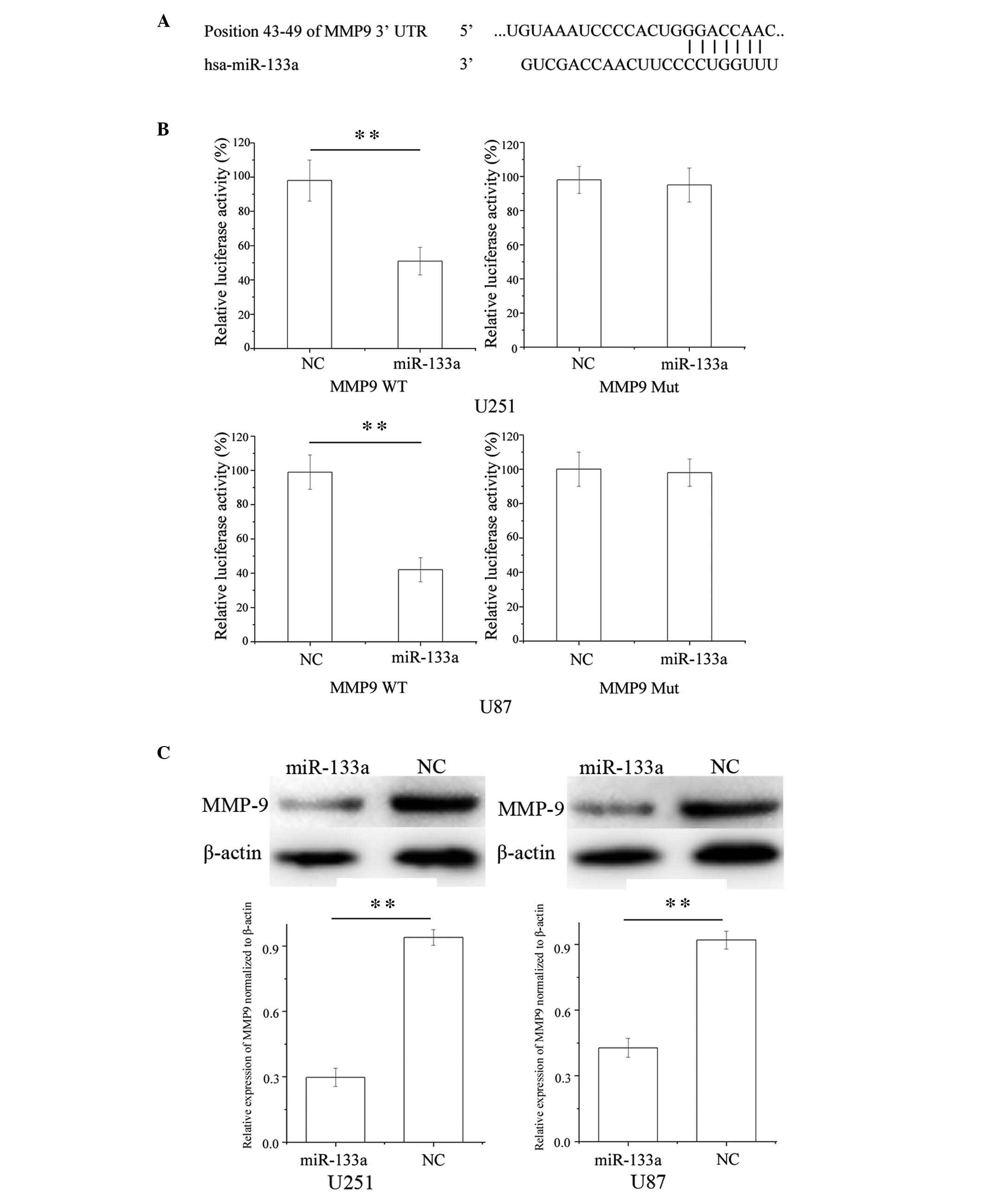
To identify the target of miR-133a in glioma cells,
TargetScan (http://www.targetscan.org) was used
and MMP9 was predicted to be a target of miR-133a (Fig. 4A). To verify whether miR-133a
directly targets MMP9, luciferase reporter assays were performed.
As presented in Fig. 4B, miR-133a
significantly inhibited the luciferase activity of wild-type MMP9
but not that of mutant MMP9 in U251 and U87 glioma cell lines
(P<0.01).
Furthermore, western blotting was performed to
investigate whether MMP9 was decreased following transfection of
U251 and U87 glioma cells with miR-133a. As presented in Fig. 4C, MMP9 was significantly
downregulated in U251 and U87 glioma cells post-transfection with
miR-133a (P<0.01). These results suggest that MMP9 may be a
direct target gene of miR-133a in the U251 and U87 glioma cell
lines.
Discussion
Previous studies have indicated that miRNAs are
aberrantly expressed in several types of human cancer, and are
widely involved in the regulation of cancer development (30,31).
However, the underlying molecular mechanisms by which miRNAs
regulate carcinogenesis and cancer progression remain to be
elucidated. Investigation into differentially expressed miRNAs in
cancer tissue samples has provided important information to aid
understanding of carcinogenesis (32). The present study is, to the best of
our knowledge, the first to demonstrate that miR-133a is
downregulated in human glioma, which led to the hypothesis that
miR-133a may exert a tumor-suppressive effect in human glioma
development and progression. By analyzing overexpression of
miR-133a in human glioma cell lines, the present study verified
that miR-133a reduced cell proliferation, migration and invasion,
also suggesting a tumor suppressive role of miR-133a. Therefore,
these results may have clinical implications in the future.
miR-133 is one of the most studied and best
characterized miRNAs. miR-133a and miR-133b are on chromosome 18 in
the same bicistronic unit (20).
miR-133a has been verified as a muscle-specific miRNA, which may
modulate myoblast differentiation and participate in heart and
myogenic diseases (33,34). It has been identified as
downregulated in numerous human malignancies, including prostate
cancer (21), bladder cancer
(20), renal cell carcinoma
(22), pancreatic ductal
adenocarcinoma (23), esophageal
squamous cell carcinoma (24),
ileal carcinoid tumors (25),
rhabdomyosarcoma (26), and
hepatocellular and lung carcinomas (27,28).
Recent studies have suggested that miR-133a inhibits cell
proliferation, migration and invasion in bladder and prostate
cancer by targeting epidermal growth factor receptor (EGFR) and its
downstream effector proteins (20,21).
Previous studies have reported the effect of ectopic miR-133a on
inhibition of cancer cell growth in lung squamous cell carcinoma,
maxillary sinus squamous cell carcinoma, tongue squamous cell
carcinoma, esophageal squamous cell carcinoma, renal cell carcinoma
and rhabdomyosarcoma (22,26,32,35–37).
It has been suggested that miR-133a may be important in these types
of cancer and may serve as a potential therapeutic target for their
treatment.
Identification of miR-133a target genes is critical
for understanding its role in tumorigenesis. Studies have
demonstrated that it may regulate oncogenic transcripts in human
cells, including EGFR (21),
moesin (38), Sp1 transcription
factor (39), fascin
actin-bundling protein 1 (40),
insulin like growth factor-1 (41), glutathione S-transferase pi 1
(42), LIM and SH3 protein 1
(43) and transgelin 2 (44). The present study hypothesized that
miR-133a may function as a tumor suppressor via downregulation of
MMP9 in glioma. miR-133a transfection resulted in decreased cell
viability, and reduced migration and invasion in human glioma
cells. These results suggested that it may be useful in the
development of novel molecular markers and therapeutic strategies
to inhibit metastasis in glioma.
MMP9, which is a member of the zinc-dependent
endopeptidases family, is a 92 kDa type IV collagenase and a key
component in the basement membrane (45–47).
Due to its extracellular matrix degrading characteristics, it has
been demonstrated that upregulation of MMP9 facilitates metastatic
spread of numerous human malignancies, and it appears to be one of
the most important molecules to directly promote cancer metastasis
(48). MMP9 has been shown to be
enhanced or activated by oncogenic proteins in several types of
human cancer, including prostate, breast, pancreatic and bladder
cancer (47,49–52).
MMP9 expression has also been detected in human glioma tissues,
where it may facilitate multiple biological events required for
glioma progression, including invasion, migration and dissemination
of glioma cells (53).
Furthermore, Yang et al (48) demonstrated that MMP9 expression is
associated with higher WHO grade, which is a key prognostic factor
for patients with glioma. Recent evidence has also suggested that
MMP9 has a distinct role in tumor angiogenesis, predominantly by
regulating the bioactivity of vascular endothelial growth factor,
which is considered the most promising factor in interfering with
tumor angiogenesis and, thus, a novel therapeutic target (54). In the present study, the results
suggested that miR-133a suppressed glioma cell migration and
invasion via downregulation of MMP9. Therefore, miR-133a may be
useful as a predictive value for early detection of tumor
metastasis, and as a therapeutic target for suppression of human
glioma invasion.
In conclusion, the present study is the first, to
the best of our knowledge, to demonstrate that miR-133a is
downregulated in human glioma, and contributes to cell
proliferation, migration and invasion by directly targeting MMP9 in
human glioma. The identification of candidate target genes of
miR-133a may provide further understanding regarding the potential
carcinogenic mechanisms in human glioma. The findings of the
present study have therapeutic implications and may be exploited
for further research into the treatment of human glioma. Future
investigations are required to address whether miR-133a may be
useful in cancer treatment, and may be a potential therapeutic
target for the treatment of glioma.
References
|
1
|
Xu Z, Zeng X, Tian D, Xu H, Cai Q, Wang J
and Chen Q: MicroRNA-383 inhibits anchorage-independent growth and
induces cell cycle arrest of glioma cells by targeting CCND1.
Biochem Biophys Res Commun. 453:833–838. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wen PY and Kesari S: Malignant gliomas in
adults. N Engl J Med. 359:492–507. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang C, Bao Z, Zhang W and Jiang T:
Progress on molecular biomarkers and classification of malignant
gliomas. Front Med. 7:150–156. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wu H, Liu Q, Cai T, Chen YD, Liao F and
Wang ZF: miR-136 modulates glioma cell sensitivity to temozolomide
by targeting astrocyte elevated gene-1. Diagn Pathol. 9:1732014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Low SY, Ho YK, Too HP, Yap CT and Ng WH:
MicroRNA as potential modulators in chemoresistant high-grade
gliomas. J Clin Neurosci. 21:395–400. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Stupp R, Hegi ME, Mason WP, van den Bent
MJ, Taphoorn MJ, Janzer RC, Ludwin SK, Allgeier A, Fisher B,
Belanger K, et al European Organisation for Research and Treatment
of Cancer Brain Tumour and Radiation Oncology Groups; National
Cancer Institute of Canada Clinical Trials Group: Effects of
radiotherapy with concomitant and adjuvant temozolomide versus
radiotherapy alone on survival in glioblastoma in a randomised
phase III study: 5-year analysis of the EORTC-NCIC trial. Lancet
Oncol. 10:459–466. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Stupp R, Mason WP, van den Bent MJ, Weller
M, Fisher B, Taphoorn MJ, Belanger K, Brandes AA, Marosi C, Bogdahn
U, et al European Organisation for Research and Treatment of Cancer
Brain Tumour and Radiation Oncology Groups; National Cancer
Institute of Canada Clinical Trials Group: Radiotherapy plus
concomitant and adjuvant temozolomide for glioblastoma. N Engl J
Med. 352:987–996. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Haapasalo J, Hyartt A, Salmi M, Nordfors
K, Lahtela SL, Kähkönen M, Helén P and Haapasalo H: Diagnosis and
prognosis of gliomas - current prospects of molecular diagnostics.
Duodecim. 130:893–901. 2014.In Finnish.
|
|
9
|
Croce CM and Calin GA: miRNAs, cancer, and
stem cell division. Cell. 122:6–7. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hwang HW and Mendell JT: MicroRNAs in cell
proliferation, cell death, and tumorigenesis. Br J Cancer.
94:776–780. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ryan BM, Robles AI and Harris CC: Genetic
variation in microRNA networks: The implications for cancer
research. Nat Rev Cancer. 10:389–402. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu C, Liang S, Xiao S, Lin Q, Chen X, Wu
Y and Fu J: MicroRNA-27b inhibits Spry2 expression and promotes
cell invasion in glioma U251 cells. Oncol Lett. 9:1393–1397.
2015.PubMed/NCBI
|
|
14
|
Wu D, Zhou Y, Pan H, Qu P and Zhou J:
MicroRNA-99a inhibits cell proliferation, colony formation ability,
migration and invasion by targeting fibroblast growth factor
receptor 3 in prostate cancer. Mol Med Rep. 11:1469–1475. 2015.
|
|
15
|
Kuhn AR, Schlauch K, Lao R, Halayko AJ,
Gerthoffer WT and Singer CA: MicroRNA expression in human airway
smooth muscle cells: Role of miR-25 in regulation of airway smooth
muscle phenotype. Am J Respir Cell Mol Biol. 42:506–513. 2010.
View Article : Google Scholar :
|
|
16
|
Baek D, Villén J, Shin C, Camargo FD, Gygi
SP and Bartel DP: The impact of microRNAs on protein output.
Nature. 455:64–71. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sun YC, Wang J, Guo CC, Sai K, Wang J,
Chen FR, Yang QY, Chen YS, Wang J, To TS, et al: miR-181b
sensitizes glioma cells to teniposide by targeting MDM2. BMC
Cancer. 14:6112014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ventura A and Jacks T: MicroRNAs and
cancer: Short RNAs go a long way. Cell. 136:586–591. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhou Y, Wu D, Tao J, Qu P, Zhou Z and Hou
J: MicroRNA-133 inhibits cell proliferation, migration and invasion
by targeting epidermal growth factor receptor and its downstream
effector proteins in bladder cancer. Scand J Urol. 47:423–432.
2013. View Article : Google Scholar
|
|
21
|
Tao J, Wu D, Xu B, Qian W, Li P, Lu Q, Yin
C and Zhang W: MicroRNA-133 inhibits cell proliferation, migration
and invasion in prostate cancer cells by targeting the epidermal
growth factor receptor. Oncol Rep. 27:1967–1975. 2012.PubMed/NCBI
|
|
22
|
Kawakami K, Enokida H, Chiyomaru T,
Tatarano S, Yoshino H, Kagara I, Gotanda T, Tachiwada T, Nishiyama
K, Nohata N, et al: The functional significance of miR-1 and
miR-133a in renal cell carcinoma. Eur J Cancer. 48:827–836. 2012.
View Article : Google Scholar
|
|
23
|
Szafranska AE, Davison TS, John J, Cannon
T, Sipos B, Maghnouj A, Labourier E and Hahn SA: MicroRNA
expression alterations are linked to tumorigenesis and
non-neoplastic processes in pancreatic ductal adenocarcinoma.
Oncogene. 26:4442–4452. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chiyomaru T, Enokida H, Tatarano S,
Kawahara K, Uchida Y, Nishiyama K, Fujimura L, Kikkawa N, Seki N
and Nakagawa M: miR-145 and miR-133a function as tumour suppressors
and directly regulate FSCN1 expression in bladder cancer. Br J
Cancer. 102:883–891. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ruebel K, Leontovich AA, Stilling GA,
Zhang S, Righi A, Jin L and Lloyd RV: MicroRNA expression in ileal
carcinoid tumors: Downregulation of microRNA-133a with tumor
progression. Mod Pathol. 23:367–375. 2010. View Article : Google Scholar :
|
|
26
|
Rao PK, Missiaglia E, Shields L, Hyde G,
Yuan B, Shepherd CJ, Shipley J and Lodish HF: Distinct roles for
miR-1 and miR-133a in the proliferation and differentiation of
rhabdomyosarcoma cells. FASEB J. 24:3427–3437. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Du L, Borkowski R, Zhao Z, Ma X, Yu X, Xie
XJ and Pertsemlidis A: A high-throughput screen identifies miRNA
inhibitors regulating lung cancer cell survival and response to
paclitaxel. RNA Biol. 10:1700–1713. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen X, Bo L, Zhao X and Chen Q:
MicroRNA-133a inhibits cell proliferation, colony formation
ability, migration and invasion by targeting matrix
metallopeptidase 9 in hepatocellular carcinoma. Mol Med Rep.
11:3900–3907. 2015.PubMed/NCBI
|
|
29
|
Louis DN, Ohgaki H, Wiestler OD, Cavenee
WK, Burger PC, Jouvet A, Scheithauer BW and Kleihues P: The 2007
WHO classification of tumors of the central nervous system. Acta
Neuropathologica. 114:97–109. 2007. View Article : Google Scholar
|
|
30
|
Tao J, Wu D, Li P, Xu B, Lu Q and Zhang W:
MicroRNA-18a, a member of the oncogenic miR-17-92 cluster, targets
Dicer and suppresses cell proliferation in bladder cancer T24
cells. Mol Med Rep. 5:167–172. 2012.
|
|
31
|
Nelson KM and Weiss GJ: MicroRNAs and
cancer: Past, present, and potential future. Mol Cancer Ther.
7:3655–3660. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kano M, Seki N, Kikkawa N, Fujimura L,
Hoshino I, Akutsu Y, Chiyomaru T, Enokida H, Nakagawa M and
Matsubara H: miR-145, miR-133a and miR-133b: Tumor-suppressive
miRNAs target FSCN1 in esophageal squamous cell carcinoma. Int J
Cancer. 127:2804–2814. 2010. View Article : Google Scholar
|
|
33
|
Rao PK, Kumar RM, Farkhondeh M,
Baskerville S and Lodish HF: Myogenic factors that regulate
expression of muscle-specific microRNAs. Proc Natl Acad Sci USA.
103:8721–8726. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bostjancic E, Zidar N, Stajer D and Glavac
D: MicroRNAs miR-1, miR-133a, miR-133b and miR-208 are dysregulated
in human myocardial infarction. Cardiology. 115:163–169. 2010.
View Article : Google Scholar
|
|
35
|
Moriya Y, Nohata N, Kinoshita T, Mutallip
M, Okamoto T, Yoshida S, Suzuki M, Yoshino I and Seki N: Tumor
suppressive microRNA-133a regulates novel molecular networks in
lung squamous cell carcinoma. J Hum Genet. 57:38–45. 2012.
View Article : Google Scholar
|
|
36
|
Nohata N, Hanazawa T, Kikkawa N, Sakurai
D, Sasaki K, Chiyomaru T, Kawakami K, Yoshino H, Enokida H,
Nakagawa M, et al: Identification of novel molecular targets
regulated by tumor suppressive miR-1/miR-133a in maxillary sinus
squamous cell carcinoma. Int J Oncol. 39:1099–1107. 2011.PubMed/NCBI
|
|
37
|
Wong TS, Liu XB, Chung-Wai Ho A, Po-Wing
Yuen A, Wai-Man Ng R and Ignace Wei W: Identification of pyruvate
kinase type M2 as potential oncoprotein in squamous cell carcinoma
of tongue through microRNA profiling. Int J Cancer. 123:251–257.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kinoshita T, Nohata N, Fuse M, Hanazawa T,
Kikkawa N, Fujimura L, Watanabe-Takano H, Yamada Y, Yoshino H,
Enokida H, et al: Tumor suppressive microRNA-133a regulates novel
targets: Moesin contributes to cancer cell proliferation and
invasion in head and neck squamous cell carcinoma. Biochem Biophys
Res Commun. 418:378–383. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Qiu T, Zhou X, Wang J, Du Y, Xu J, Huang
Z, Zhu W, Shu Y and Liu P: miR-145, miR-133a and miR-133b inhibit
proliferation, migration, invasion and cell cycle progression via
targeting transcription factor Sp1 in gastric cancer. FEBS Lett.
588:1168–1177. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Qin Y, Dang X, Li W and Ma Q: miR-133a
functions as a tumor suppressor and directly targets FSCN1 in
pancreatic cancer. Oncol Res. 21:353–363. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Guo J, Xia B, Meng F and Lou G: miR-133a
suppresses ovarian cancer cell proliferation by directly targeting
insulin-like growth factor 1 receptor. Tumour Biol. 35:1557–1564.
2014. View Article : Google Scholar
|
|
42
|
Uchida Y, Chiyomaru T, Enokida H, Kawakami
K, Tatarano S, Kawahara K, Nishiyama K, Seki N and Nakagawa M:
miR-133a induces apoptosis through direct regulation of GSTP1 in
bladder cancer cell lines. Urol Oncol. 31:115–123. 2013. View Article : Google Scholar
|
|
43
|
Chiyomaru T, Enokida H, Kawakami K,
Tatarano S, Uchida Y, Kawahara K, Nishiyama K, Seki N and Nakagawa
M: Functional role of LASP1 in cell viability and its regulation by
microRNAs in bladder cancer. Urol Oncol. 30:434–443. 2012.
View Article : Google Scholar
|
|
44
|
Li AY, Yang Q and Yang K: miR-133a
mediates the hypoxia-induced apoptosis by inhibiting TAGLN2
expression in cardiac myocytes. Mol Cell Biochem. 400:173–181.
2015. View Article : Google Scholar
|
|
45
|
Feng X, Miao G, Han Y and Xu Y: CARMA3 is
overexpressed in human glioma and promotes cell invasion through
MMP9 regulation in A172 cell line. Tumour Biol. 35:149–154. 2014.
View Article : Google Scholar
|
|
46
|
Yan Y, Liang H, Li T, Li M, Li R, Qin X
and Li S: The MMP-1, MMP-2, and MMP-9 gene polymorphisms and
susceptibility to bladder cancer: A meta-analysis. Tumour Biol.
35:3047–3052. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Roy R, Yang J and Moses MA: Matrix
metalloproteinases as novel biomarkers and potential therapeutic
targets in human cancer. J Clin Oncol. 27:5287–5297. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Yang X, Lv S, Liu Y, Li D, Shi R, Tang Z,
Fan J and Xu Z: The clinical utility of matrix metalloproteinase 9
in evaluating pathological grade and prognosis of glioma patients:
A meta-analysis. Mol Neurobiol. 52:38–44. 2015. View Article : Google Scholar
|
|
49
|
Stetler-Stevenson WG: Type IV collagenases
in tumor invasion and metastasis. Cancer Metastasis Rev. 9:289–303.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Choi JY, Jang YS, Min SY and Song JY:
Overexpression of MMP-9 and HIF-1α in breast cancer cells under
hypoxic conditions. J Breast Cancer. 14:88–95. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Aalinkeel R, Nair BB, Reynolds JL, Sykes
DE, Mahajan SD, Chadha KC and Schwartz SA: Overexpression of MMP-9
contributes to invasiveness of prostate cancer cell line LNCaP.
Immunol Invest. 40:447–464. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Kumar B, Koul S, Petersen J, Khandrika L,
Hwa JS, Meacham RB, Wilson S and Koul HK: p38 mitogen-activated
protein kinase-driven MAPKAPK2 regulates invasion of bladder cancer
by modulation of MMP-2 and MMP-9 activity. Cancer Res. 70:832–841.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Zhao J, Li G, Zhao Z, Wang J, Gao G and He
S: Matrix metalloproteinase-9 expression is increased in astrocytic
glioma and associated with prognosis of patients. Jpn J Clin Oncol.
42:1060–1065. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Du R, Lu KV, Petritsch C, Liu P, Ganss R,
Passegué E, Song H, Vandenberg S, Johnson RS, Werb Z and Bergers G:
HIF1alpha induces the recruitment of bone marrow-derived vascular
modulatory cells to regulate tumor angiogenesis and invasion.
Cancer Cell. 13:206–220. 2008. View Article : Google Scholar : PubMed/NCBI
|