Introduction
Cervical carcinoma is one of the most common
malignant tumors among females. Improving the survival rate of
patients with cervical carcinoma is an important aim of clinical
and basic research (1). With the
development of genetic engineering technology (particularly the
phage display technique), research and application of genetically
engineered antibodies in the diagnosis and treatment of diseases
has increased. Bispecific antibodies (BsAbs), prepared by chemical
coupling, hybridoma cell culture and gene engineering methods,
combine with associated antigens on the surface of tumor cells and
T cells and produce biological effects (2). BsAbs combine with target tumor cells,
as well as cytotoxic effector cells, resulting in efficient
targeted killing of tumor cells, and thus present a specific and
selective antibody-based treatment (3). Consequently BsAbs have become a
popular topic of tumor immunotherapy research. Single-chain BsAbs
(scBsAbs) link two different single-chain antibody (scAb) gene
fragments at the DNA level via an inter-chain linker and is
directly expressed as one scBsAb molecule. The scBsAb is of
interest due to qualities, such as weak antigenicity, strong
penetrability and few side effects (4).
Cytotoxic T cells are important in multiple immune
mechanisms associated with the prevention of tumors (5), and the activation and mediation of
the cytotoxic T cell killing effect on tumor cells is considered to
be a promising therapeutic strategy. Cluster of differentiation
(CD)3 is a type of membrane antigen located on the surface of
mature T cells, which forms complexes with the T-cell receptor, and
contributes to antigen recognition and intracellular signal
transduction (6). BsAbs, formed
using anti-CD3x antitumor monoclonal antibodies (mcAbs), may
trigger T cell major histocompatibility complex to kill and lyse
tumor cells. Numerous experiments have demonstrated that CD3-based
BsAbs are an efficient type of therapeutic agent for treating
tumors (7–9). In addition, the CD19 × CD13 scBsAb
constructed in a previous study is currently undergoing clinical
trials (10).
Currently, scBsAbs targeting different types of
tumor, such as ovarian cancer (11), prostate cancer (12) and lymphoma (13), have been constructed; however, to
the best of our knowledge, an scBsAb against cervical carcinoma has
not been constructed. The present study aimed to establish an
anti-human cervical carcinoma/human CD3 scBsAb and detect its
biological activity to provide the experimental and theoretical
basis for clinical application.
Materials and methods
Cell lines
The CSA125 anti-cervical cancer mcAb hybridoma cell
line, the Cs1213 cervical cancer cell line and the Jurkat T cell
line were established and preserved in our lab according to a
previous study (14). Cells were
cultured in RPMI-1640 medium (Borunlaite Science & Technology
Co., Ltd., Beijing, China) supplemented with penicillin (100 U/ml),
streptomycin (100 mg/l) from Anpei Chemical Science and Technology
Co., Ltd., (Nanjing, China) and 10% fetal calf serum (Beijing
Biodee Biotechnology Co., Ltd., Beijing, China). The current study
was approved by the ethics committee of the Experimental Animal
Center of Zhengzhou University (Zhengzhou, China), and written
informed consent was obtained prior to collection of patient
samples.
Plasmids and main reagents
The pMD18-T-anti-CD3 scFv plasmid was constructed
according to the methods of a previous study (15). Mouse monoclonal anti-human CD3
antibody (#BM0210) and an ELISA kit were purchased from Wuhan
Boster Biological Technology, Ltd. (Wuhan, China). Restriction
enzymes and DNA ligases were provided by Promega Corporation
(Madison, WI, USA) and DNA gel extraction, Mini Plasmid and MTT
kits, as well as E. coli Top10 competent cell and E.
coli BL21(DE3) bacterial strain were purchased from Tiangen
Biotech Co., Ltd. (Beijing, China).
Construction of anti-human cervical
carcinoma antibody single-chain Fv fragment (CSAs-1 scFv)
Total RNA was extracted. Briefly, 1×106
CSA125 anti-human cervical cancer hybridoma cells in the
logarithmic growth phase were collected and added into 1.5 ml
TRIzol lysate (Borunlaite Science & Technology Co., Ltd.).
Subsequently, 300 µl chloroform (Yilin Chemical Company,
Zhengzhou, China) was added, and the mixture was oscillated and
centrifuged at 4°C and 10,000 × g for 15 min. The solution was
transferred to a fresh Eppendorf (EP) tube (Topscien Instrument
Co., Ltd., Ningbo, China) and 0.5 ml Avantin (Yilin Chemical
Company) was added, the EP tube was left for 10 min at room
temperature and centrifuged at 4°C and 10,000 × g for a further 15
min. The precipitates were obtained and washed using 1 ml of 75%
alcohol. Centrifugation at 4°C and 1,500 × g was conducted to
remove the supernatant. The precipitates were dried, dissolved in
double-distilled (dd)H2O, processed with
diethylpyrocarbonate (DEPC; Borunlaite Science & Technology
Co., Ltd.) and stored at −70°C. Amplification of VL and VH gene
fragments was then performed as follows: i) Total RNA was collected
to synthesize the first chain cDNA as described above. The reaction
system, which contained 0.5 µl RNA, 1 µl oligo dT
joint primer, 1 µl AMV reverse transcriptase, 2 µl of
2.5 mmol/l dNTP, 2 µl of 10× RNA PCR buffer solution, 4
µl MgCl2 and 9.5 µl ddH2O, was
processed with DEPC. The reaction was conducted at 42°C for 30 min,
99°C for 5 min and 5°C for 5 min. ii) Amplification of the VH gene
fragment was conducted using the rapid amplification of cDNA ends
method with the following primers (synthesized by Sangon Biotech
Co., Ltd., Shanghai, China): Forward,
5′-GGTTCAGAAGTTCAACTAGTTGACATTGTGATGACCCAGTCTCCT-3′ (F1) and
reverse, 5′-GGCTCGAGTTTTATTTCCA-3′ (R1). iii) Amplification of the
VL gene fragment was conducted according to the conventional
polymerase chain reaction method using the following primers
(synthesized by Sangon Biotech Co., Ltd.): Forward,
5′-GCGAATTCCAGGTCCGCTTCAGCAGTCT-3′ (F2) and reverse,
5′-AGACCCACCACCAGCGCGCTTAAGTTCTGAGGAGACGGTGACTGAGG-3′ (R2). The
total reaction volume was 80 µl, containing 8 µl of
10× LA buffer solution, 1 µl LA Taq DNA polymerase (Takara
Biotechnology Co., Ltd., Dalian, China), 6 µl
MgCl2, 1 µl R2 primer and 1 µl F2 primer.
The PCR reaction procedure was as follows: 98°C for 10 sec, 55°C
for 30 sec and 72°C for 2 min for 30 cycles. The PCR products were
detected by 2% agarose gel electrophoresis and DNA extraction. The
CSAs-1 scFv was then constructed. Splicing overlap extension was
used to link VH to VL using a (Gly4Ser)3
linker (synthesized by Sangon Biotech Co., Ltd.). The VH, VL and
(Gly4Ser)3 linker (10 µl of each) were
mixed together, and 2.5 µl dNTP, 2 µl LA Taq DNA
polymerase, 2 µl of 10× PCR buffer solution was added.
ddH2O was then added to form a 50-µl solution.
The reaction was as follows: Initial denaturation at 94°C for 2
min, and 20 cycles of 94°C for 30 sec, 55°C for 30 sec and 72°C for
2 min, with a final extension at 72°C for 5 min. Then, 4 µl
F1 primer and 4 µl R2 primer were added and the reaction was
performed for 30 cycles under the above-mentioned conditions.
Agarose gel (0.015 g/ml; Borunlaite Science & Technology Co.,
Ltd.) electrophoresis was used for identification and separation.
Finally, the CSAs-1 scFv cloning vector was constructed. The CSAs-1
scFv gene was inserted into a pTHA90 vector (Biovector Co., Ltd.,
Beijing, China) and transformed into competent cell E. coli
Top10. The positive clone was screened for sequencing using Sanger
sequencing by Takara Biotechnology Co., Ltd.
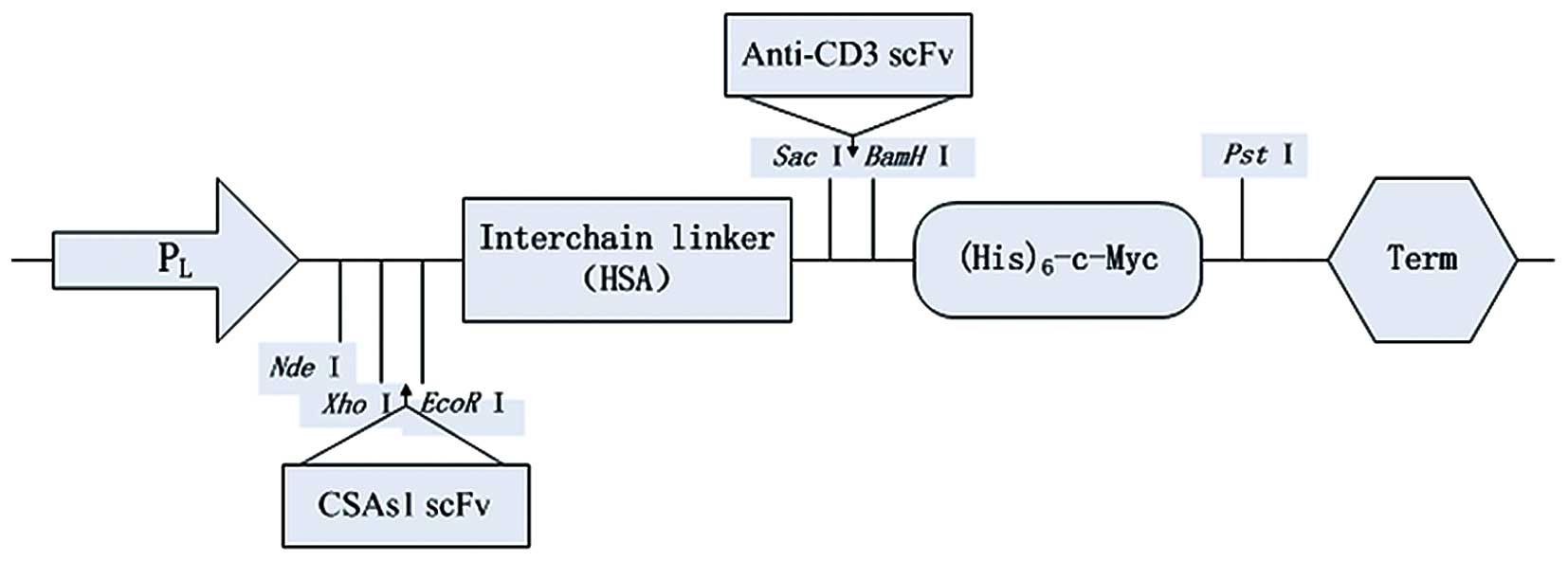
Construction of scBsAb universal
expression vector
The plasmid, pALM (Biovector Co., Ltd.) served as
the primary vector. To facilitate purification and activity
determination, the nucleotide sequence encoding
(His)6-c-Myc was synthesized and cloned between the
restriction enzyme sites, BamHI and PstI, of pALM.
The modified vector was designated, pALMm. The synthetic
inter-chain linker, human serum albumin (HSA;
N′-FQNALLVRYTKKVPQVSTPTLVEVS-C′; Seebio Biotech, Inc., Shanghai,
China) (16) was inserted between
the EcoRI and SacI sites of pALMm to construct the
scBsAb universal vector, pALMm-HSA via the sticky end connection
method, which was validated by sequencing. The primers used in
sequencing were as follows: 5′-TCTTACACATTCCAGCCCTG-3′ and
5′-TGTAAAACGACGGCCAGTGC-3′ (synthesized by Sangon Biotech Co.,
Ltd.). A schematic diagram of the constructed universal vector is
demonstrated in Fig. 1.
Construction and expression of anti-human
cervical carcinoma/anti-CD3 scBsAb
The anti-cervical carcinoma ScAb (CSAs-1) and
universal vector (pALMm-HSA) were digested by XhoI and
EcoRI, and ligated using T4 DNA ligase to form the
pALMm-CSAs-1-HSA plasmid. Similarly, anti-CD3 scFv and pALMm-HSA
were digested by SacI and BamHI and inserted into the
pALMm-HAS-anti-CD3 scFv plasmid. The pMD18-T-anti-CD3 scFv and
pALMm-CSAs-1 scFv-HSA plasmids were digested by SacI and
BamHI, respectively and anti-CD3 scFv was inserted into
pALMm-CSAs-1 scFv-HSA using a ligase to form the scBsAb expression
vector, pALMm-CSAs-1 scFv-HAS-anti-CD3 scFv. pALMm-CSAs-1-HSA,
pALMm-HSA-anti-CD3 scFv and pALMm-CSAs-1 scFv-HSA-anti-CD3 scFv
were transferred to the E. coli BL21(DE3) bacterial strain
and the positive clones were identified, which were then inoculated
with RM medium (Shanghai Seebio Biotech, Inc.) and cultivated at
30°C overnight. The following day, the bacterial solution was
transferred to RM medium (ratio, 1:10) and grown until the culture
reached an A600 of 0.6–0.8 at 37°C (measured with a 759S
UV-Visible Spectrophotometer; Lengguang Technology Co., Ltd.,
Shanghai, China). Tryptophan (Jinghai Amino Acid Co., Ltd., Wuxi,
China) was added to achieve a final concentration of 100
µg/ml and cultivated for a further 4 h. The pALMm-CSAs-1
scFv-HSA and pALMm-HSA-anti-CD3 scFv plasmids served as control
groups.
Purification and renaturation of
antibody
The bacteria were collected by centrifugation and
subsequently disrupted by ultrasonification with a JY92-II Sonifier
Cell Disrupter (Scientz Biotechnology Co., Ltd., Ningbo, China) at
a power of 400 W for 9 sec with intervals of 2 sec for a total of
30 min. Inclusion bodies were recycled through high-speed
centrifugation at 4,000 × g and purified through a C-terminal
(His)6 tag using Ni-NTA Agarose under denaturation
conditions. Purified antibodies were refolded by column
chromatography using Sephacryl S-200 HR (1.1×90 cm), which was
balanced by 3× the column volume of refolding buffer solution prior
to protein loading. The purified antibodies were loaded and passed
through the column. The refolding buffer solution was used to elute
the antibodies, which were collected and dialyzed with
phosphate-buffered saline (PBS), then stored at −80°C. The purified
and renatured proteins were detected by sodium dodecyl
sulfate-polyacrylamide gel electrophoresis analysis.
Determination of antigen binding activity
of scBsAb
The ELISA method was used to determine the antigen
binding activity of scBsAb. A Cs1213 (50 µg/ml) or Jurkat
(50 µg/ml) was diluted with coating buffer, then 0.5 ml was
added to each 96-well plate, which was then incubated at 4°C and
subsequently blocked with 5% skimmed milk at 37°C for 2 h;
renatured scBsAb samples at 1, 10, 20, 40 and 80 µg/ml
concentrations were then added. E. coli BL21(DE3) at the
same concentrations served as a negative control and the
corresponding scAb and mcAb served as positive controls, all of
which were incubated at 37°C for 2 h. Anti-c-Myc mcAb, 9E10 was
added and incubated at 37°C for 1 h. Horseradish peroxidase-labeled
immunoglobulin G (IgG) was added and incubated at 37°C for 1 h.
Finally, 3,3′-diaminobenzidine substrate (Sigma-Aldrich China,
Inc., Shanghai, China) was added for color development and
absorbance was measured at 490 nm using a Multiskan MK3 microplate
reader (Thermo Fisher Scientific, Inc., Waltham, MA, USA).
Blood pharmacokinetics assay
The iodogen method was used for radioiodination of
the antibodies. BALB/c mice were provided and raised by the Animal
Experimental Center of Zhengzhou University. The average daily feed
and water consumption values were 5 g and 6–7 ml per 100 g body
weight, respectively. Mice were caged at a temperature of 21±2°C,
with relative humidity of 30–70%, and a 12-h light/dark cycle.
BALB/c mice (n=36) were divided into four groups of nine mice per
group. Mice in each group were intravenously injected via the tail
with 125I-scBsAb, 125I-CSAs-1 scFv,
125I-anti-CD3 scFv and 125I-anti-CD3 mcAb
(0.2 ml/per mouse). Blood (10 µl) was collected from the
tail veins following these injections at 0, 5, 10, and 30 min, 1,
2, 3, 6, 12 and 24 h. A blood radioactivity count value of 1 min
was determined using a Hidex Automatic Gamma Counter (DL Naturegene
Life Sciences, Inc., Beijing, China) and a blood elimination curve
was drawn.
Rosette formation test of peripheral
blood lymphocytes (PBLs) and Cs1213 cells mediated by scBsAb
Peripheral blood mononuclear cells (PBMCs) and PBLs
were separated according to the standard method of Ficoll-Paque
density gradient centrifugation (17). Briefly, Ficoll lymphocyte
separation liquid was used to separate peripheral blood of three
healthy donors (3 ml fasting blood from each patient), who all
provided written informed consent. PBMCs were obtained following
the removal of platelets by centrifugation at 150 × g. Fresh PBLs
were obtained after the removal of macrophages and monocytes
subsequent to PBMC adherent growth for 2 h. RPMI-1640 complete
medium was added and the solution was placed in an incubator with
5% CO2 at 37°C. PBLs (1×106/ml) were
pre-stimulated with 20 µg/ml phytohemagglutinin and 100 U/ml
interleukin (IL)-2 (Shanghai Yuanye Biotechnology Co., Ltd.,
Shanghai, China) for 2 days. On the third day the cell
concentration was adjusted to 0.5×106/ml, cultivated in
complete medium containing 100 U/ml IL-2 and passaged every 2–3
days to maintain the same density of lymphocytes.
The rosette formation test was performed on the
following groups: i) scBsAb [scBsAb (40 µg/ml) + Cs1213 +
PBLs]; ii) CSAs-1 ScFv [CSAs-1 ScFv (20 µg/ml) + Cs1213 +
PBLs]; iii) Anti-CD3 ScFv [anti-CD3 ScFv (20 µg/ml) + Cs1213
+ PBLs]; iv) CSAs-1 ScFv + anti-CD3 ScFv [CSAs-1 ScFv (20
µg/ml) + Anti-CD3 ScFv (20 µg/ml) + Cs1213 + PBLs];
and v) medium (RPMI-1640 perfect medium + Cs1213 + PBLs). Cs1213
cells (1×105/ml) were collected and resuspended in PBS
and the antibodies at corresponding concentrations were added
according to the above groups until the final volume of each group
was 1 ml. The solutions were incubated at 4°C for 2 h and
centrifuged at 150 × g, and the supernatant was discarded. The
precipitates were washed with PBS three times and 1 ml fresh PBLs
(1×106/ml) was added and incubated at 4°C for a further
2 h. Rosette formation was observed under an IX73 inverted
microscope [Olympus Corporation (China) Co., Ltd., Beijing, China].
The number of Cs1213 cells (out of 200 Cs1213 cells) that combined
with at least three lymphocytes were counted to give the rosette
formation rate, and the number of lymphocytes that combined with
100 Cs1213 cells was counted to give the conjugate rate.
Determination of killing effect of
scBsAb-mediated PBLs on tumor cells
Cs1213 cells in the logarithmic growth phase were
seeded on 96-well plates (1×104/well). The plate was
incubated with 5% CO2 and cultivated overnight at 37°C
to grow Cs1213 adherent cells. The Cs1213 cells were designated as
the target cells and the PBLs were added at a ratio of 10:1
effector:target cells; in addition, antibodies at different
concentrations were added. The concentrations of scBsAbs were 1,
10, 20, 40 and 80 µg/ml and the corresponding controlled
concentrations of scAb were 0.5, 5, 10, 20 and 40 µg/ml. The
target Cs1213 cells and the RPMI-1640 complete medium group served
as controls. Following cultivation for 48–72 h, the effector cells
were washed away and MTT was added for further incubation for 4 h.
The supernatant was discarded and 100 µl dimethyl sulfoxide
was added. After standing at 37°C for 30 min, the absorbance at 570
nm was measured using a microplate reader and the killing rate was
calculated according to the following formula: Killing rate =
(Atarget−Aexperiment)/(Atarget−Acontrol)
× 100.
Results
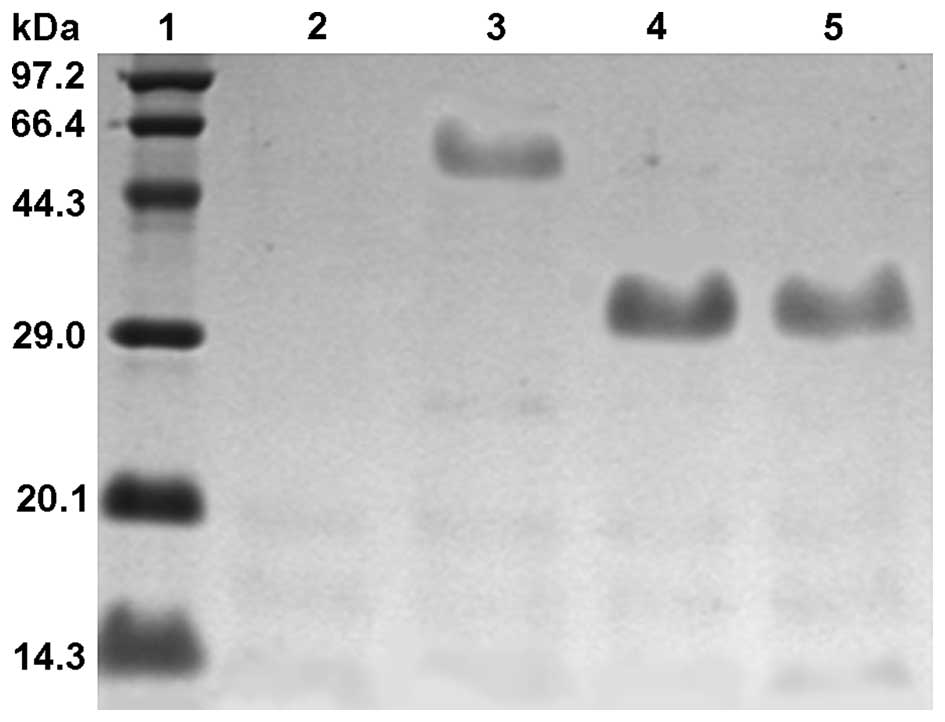
Purification and detection of
antibodies
scFvs and scBsAbs produced by induced expression
were purified by Ni-NTA metal affinity chromatography and
denaturation, and target proteins with >90% purity were
obtained. Following refolding by gel filtration chromatography, the
purified antibodies became active proteins. Detection of SDS-PAGE
demonstrated that the molecular weight of scBsAb was ~60 kDa,
compared with CSAs-1 scFv and anti-CD3 scFv, which were ~30 kDa
(Fig. 2), which was consistent
with the hypothesis.
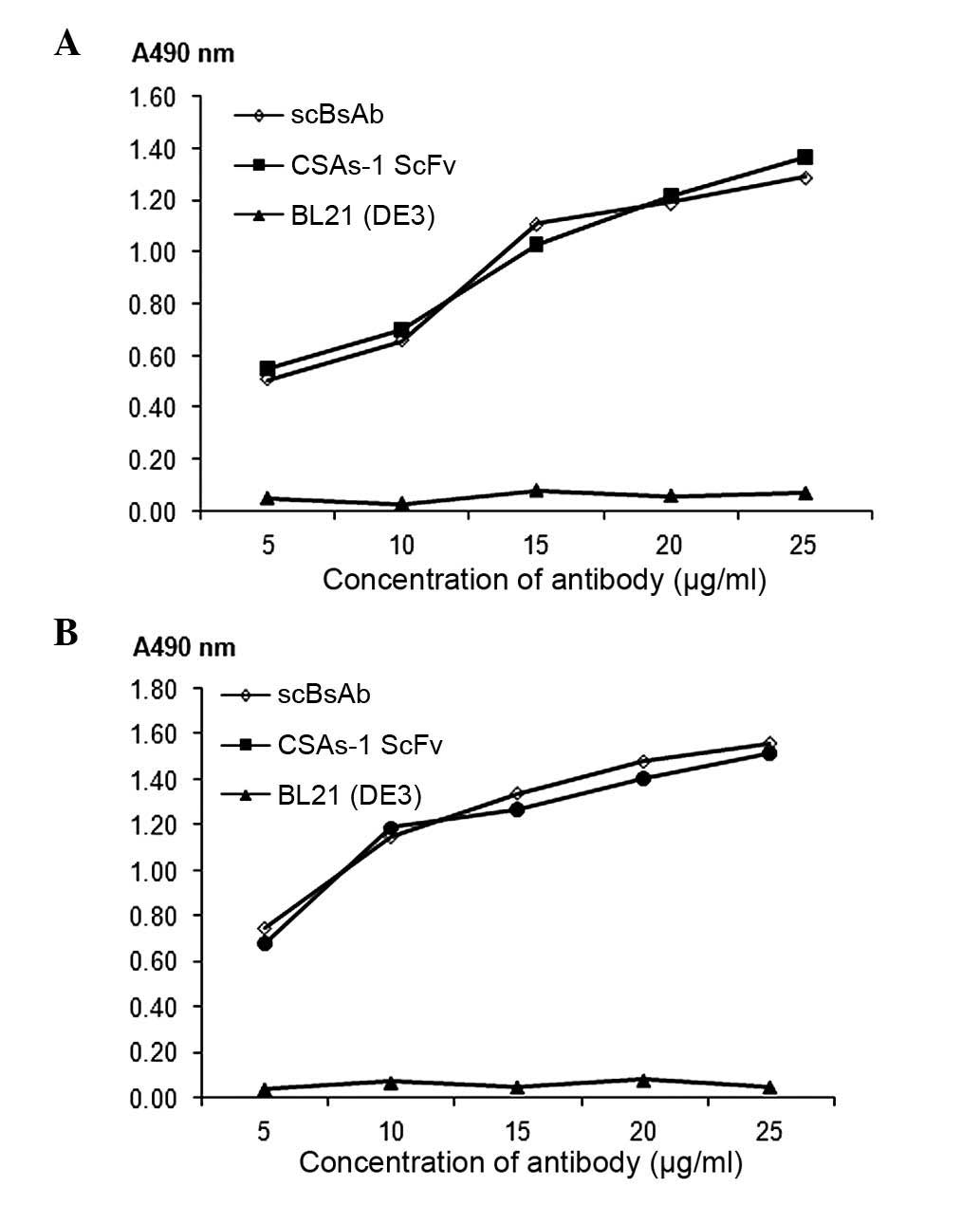
Antigen binding activity assay of
scBsAb
The ELISA assay indicated that there was no
significant difference between scBsAb and CSAs-1 ScFv in their
ability to bind to Cs1213 membrane antigens (P>0.05; Fig. 3A). Furthermore, there was no
significant difference between scBsAb and anti-CD3 scFv in their
ability to bind to Jurkat membrane antigens (P>0.05; Fig. 3B). No binding was observed between
E. Coli BL21(DE3) and the two antigens. This indicated that
the binding ability of scBsAb with the two antigens was comparable
to the original scFv. In addition, the A490 nm value increased with
the increase in antibody concentration within a range of 5~25
µg/ml.
Blood pharmacokinetics
The results of pharmacokinetics demonstrated that
the blood elimination curve of scBsAb was consistent with the two
compartment model, the distribution half-life
(T1/2α) was 17.8 min and the elimination
half-life (T1/2β) was 4.4 h. Compared with
complete mcAb, scBsAb demonstrated rapid distribution and quick
excretion (P<0.01). However, when compared with CSAs-1 scFv and
anti-CD3 scFv, scBsAb exhibited a prolonged retention time in the
body (P<0.01; Table I).
Additionally, the conjugate rate mediated by scBsAb was
significantly higher than that mediated by scFv (P<0.05) or
RPMI-1640 medium (P<0.01).
 | Table IAnalysis of pharmacokinetic
parameters. |
Table I
Analysis of pharmacokinetic
parameters.
| Antibody type |
T1/2α (min) |
T1/2β (h) |
|---|
| scBsAb | 17.8±0.12 | 4.4±0.05 |
| CSAs-1 scFv | 5.5±0.08a | 2.6±0.09a |
| Anti-CD3 scFv | 5.7±0.11a | 2.3±0.04a |
| Anti-CD3 mcAb | 60.3±0.10a | 18.1±0.11a |
Rosette formation detection
The number of rosettes formed by PBLs and Cs1213
cells following stimulation with scBsAb at 4°C for 2 h was
significantly higher than those that were stimulated with scFv
(P<0.05) or RPMI-1640 medium (P<0.01). However, there was no
significant difference between the scFv and the RPMI-1640 groups
(P>0.05; Table II). This
indicated that scBsAbs mediate the combination of CD3+ T
cells and Cs1213 cells.
| Table IIRosette formation and conjugate rates
induced by scBsAb. |
Table II
Rosette formation and conjugate rates
induced by scBsAb.
| Group | Rosette formation
rate (%) | Conjugate rate
(%) |
|---|
| scBsAb | 15.5±1.12 | 82.6±3.24 |
| RPMI-1640 | 10.9±1.45a | 54.8±1.89a |
| CSAs-1 scFv | 12.3±0.77b | 61.7±1.55b,c |
| Anti-CD3 scFv | 13.1±1.02b | 62.3±2.12b,c |
| CSAs-1 + anti-CD3
scFv | 12.6±1.14b | 51.9±1.87a |
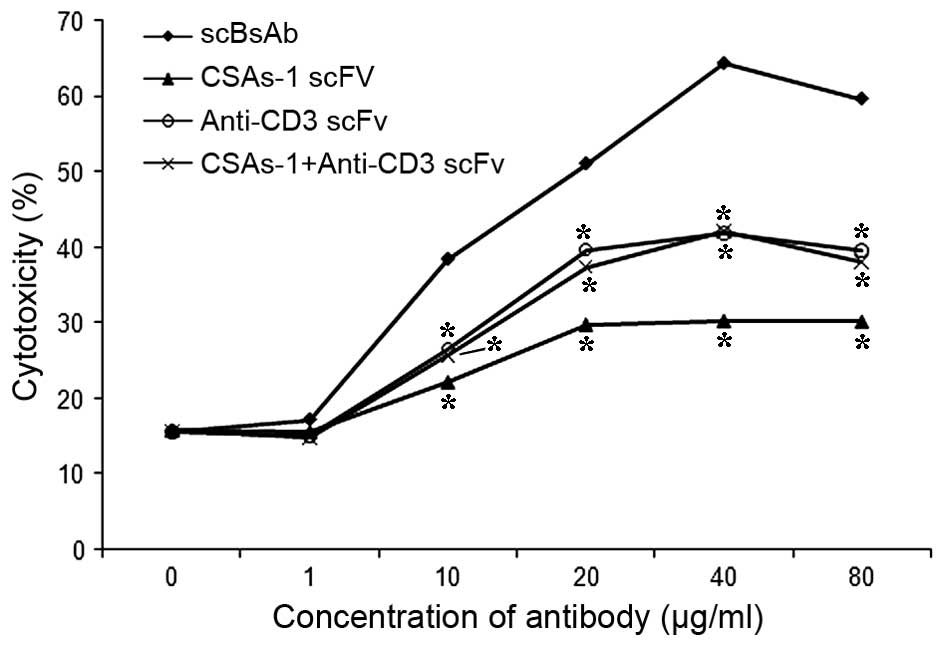
In vitro cytotoxic activity mediated by
scBsAbs
When the ratio of effector cells to target cells was
10:1 and the action time was 48 h, the strength of cytotoxic
activity increased in an antibody concentration-dependent manner
≤40 µg/ml. At a concentration of 40 µg/ml antibody,
the cytotoxic activity reached 64.5%. The killing rate of Cs1213 by
PBL in the scBsAb group was significantly higher than that in the
CSAs-1 scFv, anti-CD3 scFv and CSAs-1 scFv + anti-CD3 scFv groups
(P<0.05). Thus, the killing rate of target cells by PBLs was
associated with the dose of antibodies (Fig. 4).
Discussion
With the developments in biology, immunology and
molecular genetics, cancer biotherapy has become a key area of
research and BsAbs are a prominent focus (18). The aim of the present study was to
construct an scBsAb against cervical carcinoma. The hybridoma cell
strain, CSA125, which secretes highly specific anti-human cervical
carcinoma mcAb was used to construct the scAb gene, CSAs-1. The
modified plasmid, pLAM served as the universal expression vector,
and scBsAbs were constructed and successfully expressed. scBsAb was
demonstrated to combine with the Cs1213 cervical cancer and Jurkat
cell membrane antigens, and maintain the binding activities in a
similar manner to the original scAb. This indicated that scBsAb was
able to combine with the membrane antigens of tumor cells and
triggering molecules on effector cells to activate effector cells,
enabling them to accumulate on, and subsequently kill, the tumor
cells (19). Sharkey et al
(20) created CEA/HSG bispecific
antibodies combining PET and SPECT applied in the molecular imaging
of nude mice with colon cancer, which demonstrated that the
antibody had high target ability and was able to locate the tumor
accurately. Melanoma-associated chondroitin sulfate proteoglycan ×
CD3 bsAb constructed by Gary et al (21) produced CD4+ and
CD8+ T cells in order to kill melanoma cells. Therefore,
this may serve as a novel immunological method of treating
melanoma. Another study demonstrated that CD3×CD20 bispecific
antibodies could be applied in the oncotherapy following umbilical
cord blood stem cell transplantation (22). In the present study, the
constructed anti-human cervical carcinoma × anti-CD3 scBsAb
mediated the combination of PBLs with Cs1213 cells and activated
the specific killing effect of PBLs. This indicated that anti-human
cervical carcinoma × anti-CD3 scBsAb has potential clinical
application in the therapy of human cervical cancer.
Pharmacokinetics demonstrated that the retention
time of scBsAb was longer than scFv, although markedly shorter when
compared with the complete antibody, anti-CD3 mcAb. Compared with
complete IgG, small genetically engineered antibodies are able to
effectively permeate into tumors. However, due to their short
half-life, various genetically engineered antibodies have short
retention times and, therefore, low absolute intake by tumors.
Adams et al (23)
demonstrated that the tumor absorption peak of BsAb against
HER2/neu marked by 125I was 10.1%, and the
T1/2β was 6.42 h; markedly lower when
compared with a complete antibody. Thus, the differences in size
and affinity affect the pharmacokinetic features of BsAbs.
Increasing the molecular weight of the antibody reduces the
elimination rate whilst weakening the tumor infiltrability.
However, the molecular size is just one of the factors that
determines the metabolism in the body. Other factors include
conformation of the antibody, charge characteristics and
arrangement of domains, which may influence the pharmacokinetic
features of BsAb (24).
Certain studies demonstrated that sequences of
interchain linkers affect the antigen binding activity of the
antibody, as well as its stability within body (25). The development of a technique,
which uses HSA to prolong the half-life of polypeptides and protein
therapeutics, has become attractive. As a stable and natural
vector, HSA has been used successfully to improve certain metabolic
parameters of recombined molecules (26,27).
In conclusion, in the present study, the fragment of
HSA, which contained no Cys and numerous polar amino acids, was
regarded as an interchain linker. Consequently, it was identified
that the interchain linker, HSA was able to significantly prolong
the half-life of scBsAb in the body and enhance the therapeutic
effect of this small molecular antibody. The targeting of BsAbs
serves an important guiding function in the diagnosis and treatment
of disease. However, certain aspects remain unknown, including
which specific types of BsAbs produce the strongest targeting, the
longest residence time and the fewest side effects to normal
tissues and organs, and how the BsAbs being used from animal
experiment to clinical application.
References
|
1
|
Dillner J: Trends over time in the
incidence of cervical neoplasia in comparison to trends over time
in human papillomavirus infection. J Clin Virol. 19:7–23. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chames P and Baty D: Bispecific antibodies
for cancer therapy. Curr Opin Drug Discov Devel. 12:276–283.
2009.PubMed/NCBI
|
|
3
|
Urbanska K, Lynn RC, Stashwick C, Thakur
A, Lum LG and Powell DJ Jr: Targeted cancer immunotherapy via
combination of designer bispecific antibody and novel
gene-engineered T cells. J Transl Med. 12:3472014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dreier T, Baeuerle PA, Fichtner I, Grün M,
Schlereth B, Lorenczewski G, Kufer P, Lutterbüse R, Riethmüller G,
Gjorstrup P and Bargou RC: T cell costimulus-independent and very
efficacious inhibition of tumor growth in mice bearing subcutaneous
or leukemic human B cell lymphoma xenografts by a
CD19-/CD3-bispecific single-chain antibody construct. J Immunol.
170:4397–4402. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Makkouk A and Weiner GJ: Cancer
immunotherapy and breaking immune tolerance: New approaches to an
old challenge. Cancer Res. 75:5–10. 2015. View Article : Google Scholar
|
|
6
|
Cartellieri M, Arndt C, Feldmann A, von
Bonin M, Ewen EM, Koristka S, Michalk I, Stamova S, Berndt N, Gocht
A, et al: TCR/CD3 activation and co-stimulation combined in one T
cell retargeting system improve anti-tumor immunity.
Oncoimmunology. 2:e267702013. View Article : Google Scholar
|
|
7
|
Tang P, Li L, Zhou Y, Shen CC, Kang YH,
Yao YQ, Yi C, Gou LT and Yang JL: The preparation of VEGFR1/CD3
bispecific antibody and its specific cytotoxicity against
VEGFR1-positive breast cancer cells. Biotechnol Appl Biochem. Dec
12–2013.Epub ahead of print. PubMed/NCBI
|
|
8
|
English DP, Bellone S, Schwab CL, Roque
DM, Lopez S, Bortolomai I, Cocco E, Bonazzoli E, Chatterjee S,
Ratner E, et al: Solitomab, an epithelial cell adhesion
molecule/CD3 bispecific antibody (BiTE), is highly active against
primary chemotherapy-resistant ovarian cancer cell lines in vitro
and fresh tumor cells ex vivo. Cancer. 121:403–412. 2015.
View Article : Google Scholar
|
|
9
|
Cheng M, Ahmed M, Xu H and Cheung NK:
Structural design of disialoganglioside GD2 and CD3-bispecific
antibodies to redirect T cells for tumor therapy. Int J Cancer.
136:476–486. 2015. View Article : Google Scholar :
|
|
10
|
Löffler A, Kufer P, Lutterbüse R, Zettl F,
Daniel PT, Schwenkenbecher JM, Riethmüller G, Dörken B and Bargou
RC: A recombinant bispecific single-chain antibody, CD19 × CD3,
induces rapid and high lymphoma-directed cytotoxicity by
unstimulated T lymphocytes. Blood. 95:2098–2103. 2000.
|
|
11
|
Yang JZ, Zhang Z, Ma L, Yao XS, Zhou MQ,
Wang XB and Wang XN: Biologic properties of an anti-human ovarian
carcinoma/anti-human CD3 single chain bispecific antibody. Ai
Zheng. 24:787–791. 2005.In Chinese. PubMed/NCBI
|
|
12
|
Wang D, Wu GJ, Wang H, Wu WZ, Yang SL and
Tan JM: Comparison of biologic activity of two anti-PSA/anti-CD3
bispecific single-chain antibodies. Zhonghua Nan Ke Xue. 13:8–12.
2007.In Chinese. PubMed/NCBI
|
|
13
|
Zhou Y, Gou LT, Mu B, Liao WC, He J, Ma C,
Yao YQ and Yang JL: A fully human CD19/CD3 bi-specific antibody
triggers potent and specific cytotoxicity by unstimulated T
lymphocytes against non-Hodgkin's lymphoma. Biotechnol Lett.
34:1183–1191. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li X, Chen W, Yang YC and Si LS:
Establishment and characterization of a new squamous cell carcinoma
cell line CS1213 from the human uterine cervix. Zhonghua Fu Chan Ke
Za Zhi. 38:614–617. 2003.In Chinese.
|
|
15
|
Kipriyanov SM, Moldenhauer G, Martin AC,
Kupriyanova OA and Little M: Two amino acid mutations in an
anti-human CD3 single chain Fv antibody fragment that affect the
yield on bacterial secretion but not the affinity. Protein Eng.
10:445–453. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gruber M, Schodin BA, Wilson ER and Kranz
DM: Efficient tumor cell lysis mediated by a bispecific single
chain antibody expressed in Escherichia coli. J Immunol.
152:5368–5374. 1994.PubMed/NCBI
|
|
17
|
de Rock E and Taylor N: An easy method of
layering blood over Ficoll-Paque gradients. J Immunol Methods.
17:373–374. 1977. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rossi DL, Rossi EA, Cardillo TM,
Goldenberg DM and Chang CH: A new class of bispecific antibodies to
redirect T cells for cancer immunotherapy. MAbs. 6:381–391. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hoffmann SC, Wabnitz GH, Samstag Y,
Moldenhauer G and Ludwig T: Functional analysis of bispecific
antibody (EpCAMxCD3)-mediated T-lymphocyte and cancer cell
interaction by single-cell force spectroscopy. Int J Cancer.
128:2096–2104. 2011. View Article : Google Scholar
|
|
20
|
Sharkey RM, Karacay H, Vallabhajosula S,
McBride WJ, Rossi EA, Chang CH, Goldsmith SJ and Goldenberg DM:
Metastatic human colonic carcinoma: Molecular imaging with
pretargeted SPECT and PET in a mouse model. Radiology. 246:497–507.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gary R, Voelkl S, Palmisano R, Ullrich E,
Bosch JJ and Mackensen A: Antigen-specific transfer of functional
programmed death ligand 1 from human APCs onto CD8+ T cells via
trogocytosis. J Immunol. 188:744–752. 2012. View Article : Google Scholar
|
|
22
|
Stanglmaier M, Faltin M, Ruf P,
Bodenhausen A, Schröder P and Lindhofer H: Bi20 (fBTA05), a novel
trifunctional bispecific antibody (anti-CD20 × anti-CD3), mediates
efficient killing of B-cell lymphoma cells even with very low CD20
expression levels. Int J Cancer. 123:1181–1189. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Adams GP, Schier R, McCall AM, Crawford
RS, Wolf EJ, Weiner LM and Marks JD: Prolonged in vivo tumour
retention of a human diabody targeting the extracellular domain of
human HER2/neu. Br J Cancer. 77:1405–1412. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Asano R, Shimomura I, Konno S, Ito A,
Masakari Y, Orimo R, Taki S, Arai K, Ogata H, Okada M, et al:
Rearranging the domain order of a diabody-based IgG-like bispecific
antibody enhances its antitumor activity and improves its
degradation resistance and pharmacokinetics. MAbs. 6:1243–1254.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Le Gall F, Reusch U, Little M and
Kipriyanov SM: Effect of linker sequences between the antibody
variable domains on the formation, stability and biological
activity of a bispecific tandem diabody. Protein Eng Des Sel.
17:357–366. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kragh-Hansen U, Chuang VT and Otagiri M:
Practical aspects of the ligand-binding and enzymatic properties of
human serum albumin. Biol Pharm Bull. 25:695–704. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Muller D, Karle A, Meissburger B, Höfig I,
Stork R and Kontermann RE: Improved pharmacokinetics of recombinant
bispecific antibody molecules by fusion to human serum albumin. J
Biol Chem. 282:12650–12660. 2007. View Article : Google Scholar : PubMed/NCBI
|