Introduction
Prostate cancer is the most common malignancy in
males in Europe and North America, while being the second leading
cause of cancer-associated mortality in American men (1,2) as
well as the fifth most common cancer type in Korean men (3). Prostate cancer at the early stage
requires androgen for survival and proliferation. Androgen
activates androgen receptor (AR), which has an important role in
the development, maturation and function of the prostate gland as
well as the progression of prostate cancer (4). Thus, prostate cancer is initially
treated by hormone manipulation, specifically androgen-ablative and
anti-androgen therapy, to inhibit the development of metastasis
(5). However, these treatments
bear considerable cardiovascular side effects and most prostate
cancers progress rapidly to become hormone resistant (6). Therefore, novel therapies and
preventative methods are required to treat prostate cancer.
The mortality rate for prostate cancer in Asian men
is lower than that in men residing in Western countries. Several
studies have shown that the high consumption of soy products such
as whole soybeans, soy milk and bean curd (tofu) in Asian countries
may be associated with reduced risk of prostate cancer (7–10).
In support of this, dietary habits have been shown to be associated
with reduced risk of cancer (11–16).
Soybeans, an important component of traditional Asian diets, is a
rich source of isoflavones, saponin and plant protein (17). Soy milk, which is a product of
hulled soybeans, contains a variety of biologically active
substances and has become a popular substitute for dairy milk
(18). Soy milk may have
beneficial effects on bone health and may reduce obesity due to the
presence of isoflavones, saponin, calcium, vitamins and
phytosterols (18,19). Of note, soy milk consumption (more
than once a day) has been shown to reduce the risk of prostate
cancer in Californian men by 70% (20). Genistein (GEN), a major isoflavone
constituent of soybeans and soy products, has been reported to
inhibit the growth of androgen-dependent as well as
androgen-independent human prostate cancer cells (7,10).
Isoflavones are phytoestrogens with beneficial effects, the most
abundant of which are daidzein and GEN in soybeans, which also
contain glycitein (21). Soy
isoflavones are typically present in their glucoside form, which is
not bioactive. After ingestion of soybeans or soy milk, isoflavone
glucosides are hydrolyzed by intestinal mucosal as well as
bacterial β-glucosidases into aglycones, which are absorbed more
rapidly and in higher amounts (22,23).
The overall bioavailability of the active components of soy is
determined by gastrointestinal digestion, absorption and metabolism
(24). The isoflavones in soy
milk, including daidzein, genistein and glycitein, structurally
resemble 17β-estradiol (E2) (25)
and display differential binding affinities to the two estrogen
receptors (ERs), ERα and ERβ (26,27).
In particular, soy isoflavones bind to ERβ 20- to 30-fold more
strongly than E2 (7). ERβ is
highly expressed in LNCaP prostate cancer cells, whereas ERα
expression is relatively low or undetectable (6). According to several studies, ERβ may
have a role in the protection against malignant cell proliferation
and metastasis through mediation of estrogen activity in prostate
cancer (8,28).
Based on these previous studies, the present study
hypothesized that intake of soy milk may have inhibitory effects on
the growth of human prostate cancer cells, an effect exerted by
combined isoflavones in soy milk biologically activated during
digestion. To prove this hypothesis, a soy milk digestion extract
(SMD) was produced by simulated gastrointestinal digestion in
vitro to obtain aglycone isoflavones. The effects of SMD, GEN
and E2 with or without inhibitors of ER or AD on the proliferative
rate of LNCaP prostate cancer cells were assessed. Furthermore, the
effects of these treatments on the expression of ERβ, AR, prostate
cancer-specific antigen (PSA) and cell cycle regulatory genes were
assessed. The present in vitro study suggested that
ingestion of soy milk may prevent the development or progression of
prostate cancer in an ERβ-dependent manner.
Materials and methods
In vitro digestion and isoflavone
extraction
SMD was prepared in a manner similar to human
gastrointestinal digestion followed by extraction of aglycone
isoflavones, as previously described by Tate et al (29) with certain modifications. Soy milk
(300 ml) was obtained from Chung's Food (Cheongju, Chungcheongbuk,
Republic of Korea) and digested in vitro. The pH of the soy
milk was adjusted to 2.8 with 6 N HCl (Merck kGaA, Darmstadt,
Germany), followed by addition of 15 ml of a pepsin (Sigma-Aldrich,
St. Louis, MO, USA) suspension (4 g pepsin/100 ml 0.1N HCl; Merck
kGaA). The mixture was incubated with shaking at 37°C for 2 h,
after which the pH was adjusted to 5.7 with 5 M NaOH (Showa Denko,
Tokyo, Japan). Subsequently, 75 ml pancreatin-bile salt mixture
(0.2 g pancreatin and 1.2 g bile salts suspended in 100 ml 0.1 M
NaHCO3 (Sigma-Aldrich) was added to the mixture,
followed by incubation as above for an additional 2 h. Following
centrifugation at 1,096 × g for 20 min at room temperature, the
solvent of the supernatant was concentrated using a rotary vacuum
evaporator (BÜCHI Laboratechnik AG, Flawil, Switzerland) and
freeze-dried using a lyophilizer (FD5512, Ilshin Lab Co., Ltd.,
Dongducheon-si, Korea).
The freeze-dried supernatant was extracted to
determine the isoflavone concentration. The powder (0.2 g) was
extracted with 80% methanol (Merck kGaA) at 65°C in a water bath
with shaking (130 rpm) for 2 h. After cooling to room temperature,
3 ml of 2M NaOH (Showa Denko) was added followed by shaking for 10
min (300 rpm). Extracts were decanted into a 50-ml tube and
centrifuged at 3,500 rpm for 20 min at room temperature. The
supernatant was then concentrated using a rotary vacuum evaporator
(BÜCHI Labortechnik AG) and freeze-dried using a lyophilizer
(Ilshin Lab Co., Ltd.). The yield was ~3.34%.
Isoflavone analysis by high-performance
liquid chromatography (HPLC)
The isoflavone concentration of the SMD obtained was
analyzed by HPLC with ultraviolet detection (Agilent 1260 infinity;
Agilent Technologies, Palo Alto, CA, USA). SMD powder was extracted
in the same manner as above. Centrifuged supernatant was decanted
and diluted into a 10-ml volumetric flask containing 4 ml ultrapure
water (18.2 MΩ·cm) for HPLC analysis. After filtration using a
0.45-µm filter (17 mm nylon syringe filter; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) attached to a nylon syringe,
extracts were filled into HPLC tubes. A Spherex5 C18 column
(150×4.6 mm inner diameter; 5 µm; Phenomenex Co, Torrance,
CA, USA) was maintained at a temperature of 40°C. The mobile phase
was composed of solution A [100% methanol, acetic acid and
ultrapure water (44:5:1, v:v)] and solution B [100% methanol and
acetic acid (49:1, v:v)], and a constant A/B ratio of 9:1 was used.
Chromatographic separation was monitored at a flow rate of 1.5
ml/min. The concentrations of daidzein, glycitein, and GEN were
measured at 260 nm. The standard material of 12 types of isoflavone
isomer was used for isoflavone analysis. The stock solution
contained a mixture of 12 types of isomers. The stock solution
mixture was as follows: 0.2 mg/ml daidzin, 0.1 mg/ml glycitin, 0.2
mg/ml genistin, 0.2 mg/ml daidzein, 0.1 mg/ml glycitein, 0.2 mg/ml
geistein (all from Sigma-Aldrich); 0.02 mg/ml 6′-O-malonly daidzin,
0.02 mg/ml 6′-O-malonly glycitin, 0.02 mg/ml 6′-O-acetyl daidzin,
0.02 mg/ml 6′-O-malonly genistin, 0.02 mg/ml 6′-O-acetyl glycitin
and 0.02 mg/ml 6′-O-acetyl genistin (all from FUGICO Co., Ltd.,
Kitakyushu, Japan). The stock solution mixture was made into
working standard solution mixtures of 6 concentrations to generate
a standard curve (Table I)
 | Table IConcentration of working standard
solution for HPLC analysis |
Table I
Concentration of working standard
solution for HPLC analysis
| Isomer | Standards
(µg/ml)
|
|---|
| 1 | 2 | 3 | 4 | 5 | 6 |
|---|
| Daidzin | 4 | 8 | 16 | 32 | 64 | 128 |
| Glycitin | 2 | 4 | 8 | 16 | 32 | 64 |
| Genistin | 4 | 8 | 16 | 32 | 64 | 128 |
|
6′-O-malonyldaidzin | 0.4 | 0.8 | 1.6 | 3.2 | 6.4 | 12.8 |
|
6′-O-malonylglycitin | 0.4 | 0.8 | 1.6 | 3.2 | 6.4 | 12.8 |
|
6′-O-acetyldaidzin | 0.4 | 0.8 | 1.6 | 3.2 | 6.4 | 12.8 |
|
6′-O-malonylgenistin | 0.4 | 0.8 | 1.6 | 3.2 | 6.4 | 12.8 |
|
6′-O-acetylglycitin | 0.4 | 0.8 | 1.6 | 3.2 | 6.4 | 12.8 |
| Daidzein | 4 | 8 | 16 | 32 | 64 | 128 |
| Glycitein | 2 | 4 | 8 | 16 | 32 | 64 |
|
6′-O-acetylgenistin | 0.4 | 0.8 | 1.6 | 3.2 | 6.4 | 12.8 |
| Genistein | 4 | 8 | 16 | 32 | 64 | 128 |
Cell culture
The androgen-dependent human prostate cancer cell
line LNCaP (Korean Cell Line Bank, Seoul, Korea) was obtained from
Professor K.C. Choi (College of Veterinary Medicine, Chungbuk
National University, Cheongju, Chungbuk, Republic of Korea). The
cells were cultured in Dulbecco's Modified Eagle's Medium (DMEM;
Invitrogen; Thermo Fisher Scientific, Inc.) supplemented with 10%
fetal bovine serum (FBS; Invitrogen, Thermo Fisher Scientific,
Inc.), 10,000 U/ml penicillin G and 10,000 µg/ml
streptomycin (Invitrogen, Thermo Fisher Scientific, Inc.) and 1%
antifungal (4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid;
Invitrogen Thermo Fisher Scientific, Inc.) at 37°C in a humidified
atmosphere of 5% CO2 in air. Cells were detached with
0.25% trypsin-ethylenediaminetetraacetic acid (Invitrogen, Thermo
Fisher Scientific, Inc.). Phenol red-free DMEM (Invitrogen, Thermo
Fisher Scientific, Inc.) supplemented with 5 or 1%
charcoal-dextran-treated FBS (CD-FBS) was used to block the effects
of the estrogenic components of FBS and DMEM (12–15).
Cell proliferation assay
The inhibitory effects of SMD and its phytoestrogen
components were assessed using a
3-(4-5-dimethylthiazol-2-yl)-2,5-dyphenyltetrazolium bromide (MTT)
assay. LNCaP cells were plated at 4,000 cells per well in 96-well
plates in 100 µl DMEM with 10% FBS. After incubation for 24
h, the medium was replaced with phenol red-free DMEM with 1%
CD-FBS, followed by incubation for a further 48 h. Cells were
washed with phosphate-buffered saline (PBS) and treated with
vehicle [0.1% dimethyl sulfoxide (DMSO)], E2 (2.7×10−7
mg/ml), GEN (2.7×10−2 mg/ml), SMD (the lyophilized SMD
powder was collected and then 3 g was dissolved in 1 ml of 100%
methanol (Merck kGaA). The concentration of SMD was calculated by
determining the concentration of total aglycon according to HPLC
analysis, Table II, 0.79 mg/ml),
casodex (CDX; also known as bicalutamide, an anti-androgen used to
treat prostate cancer and hirsutism; 4.3×10−3 mg/ml), or
ER antagonist ICI 182,780 (6.1×10−6 mg/ml) in phenol
red-free DMEM with 5% CD-FBS medium for five days. All of the above
reagents were purchased from Sigma-Aldrich. To observe the effects
on ER, CDX was applied in combination with each reagent to block
the androgen receptor, while ICI 182,780 was used to block the ER.
Media were changed every other day. At the end of the incubation
period, 10 µl MTT solution (5 mg/ml; Sigma-Aldrich) was
added to each well, followed by incubation for 4 h at 37°C. The
supernatants were then replaced with 100 µl DMSO
(Sigma-Aldrich) to dissolve the formazan crystals. The optical
density of each well was measured at 540 nm using an ELISA reader
(SpectraMax® plus 384 absorbance microplate reader; cat.
no. MNRO7122, Molecular Devices, Sunnyvale, CA, USA) and was
expressed as a percentage of the vehicle-treated group to represent
the amount of viable cells.
| Table IIConcentrations of aglycone
isoflavones in SMD according to analytical high-performance liquid
chromatography. |
Table II
Concentrations of aglycone
isoflavones in SMD according to analytical high-performance liquid
chromatography.
| Isoflavone
isomer | Concentration in
SMD (mg/100 g) |
|---|
| Daidzin | 27.057±0.002 |
|
6′-O-malonyldaidzin | 18.616±0.026 |
|
6′-O-acetyldaidzin | 21.410±0.281 |
| Daidzein | 1.988±0.411 |
| Glycitin | 25.471±0.030 |
|
6′-O-malonylglycitin | 4.030±0.067 |
|
6′-O-acetylglycitin | 3.764±0.037 |
| Glycitein | 23.537±0.010 |
| Genistin | 36.565±0.003 |
|
6′-O-malonylgenistin | 1.216±0.080 |
|
6′-O-acetylgenistin | 0.784±0.010 |
| Genistein | 0.685±0.054 |
| Total
isoflavone | 165.123±0.629 |
| Total malonyl | 23.862±0.158 |
| Total acetyl | 25.958±0.291 |
| Total aglycon | 26.209±0.481 |
| Metabolized
daidzeina | 39.803±0.431 |
| Metabolized
glyciteina | 44.097±0.081 |
| Metabolized
genisteina | 24.787±0.045 |
Semi-quantitative reverse transcription
polymerase chain reaction (RT-PCR)
RT-PCR was performed to determine the expression of
selected target genes. LNCaP cells were cultured at a density of
3.5×106 cells per well in a 60-mm round dish. Each
reagent was applied at a concentration identical to that used in
the cell proliferation assay. Total RNA was extracted at 0 and 24 h
using TRIzol reagent (Invitrogen, Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions and as described
previously (12–15,30).
The concentration of total RNA was measured using a SpectraDrop
Micro-volume micro-plate (SpectraMax® plus384 absorbance
microplate reader) at 260/280 nm. Complementary DNA (cDNA) was
synthesized from total RNA using a power cDNA synthesis kit (iNtRON
Biotechnology, Inc., Sungnam, Kyeonggido, Republic of Korea) for
RT. cDNAs of ERβ, AR, PSA, cyclin D1, cyclin-dependent kinase
(CDK)4, p21 and GAPDH were amplified using specific forward and
reverse primer sets (Bioneer Corporation, Daejeon, Korea; Table III).
| Table IIIPrimer sequences and sizes for
semi-quantitative reverse-transcription polymerase chain
reaction. |
Table III
Primer sequences and sizes for
semi-quantitative reverse-transcription polymerase chain
reaction.
| Target gene | Sequences | Expected size |
|---|
| ERβ | Sense:
5′-TGTTACTGGTCCAGGTTCAAAGAGG-3′ | 200 bp |
| Antisense:
5′-AGCCACACTTCACCATTCCCAC-3′ |
| AR | Sense: 5′-CTC TCT
CAA GAG TTT GGA TGG CT-3′ | 342 bp |
| Antisense: 5′-CAC
TTG CAC AGA GAT GAT CTC TGC-3′ |
| PSA | Sense:
5′-TGCGCAAGTTCACCCTCA-3′ | 754 bp |
| Antisense:
5′-CCCTCTCCTTACTTCATCC-3′ |
| Cyclin D1 | Sense:
5′-TCTAAGATGAAGGAGACCATC-3′ | 354 bp |
| Antisense:
5′-CCCTCTCCTTACTTCATCC-3′ |
| CDK4 | Sense:
5′-TCGTGAGGTGGCTTTACTGA-3′ | 698 bp |
| Antisense:
5′-AGGCAGAGATTCGCTTGTGT-3′ |
| p21 | Sense:
5′-AGGCACCGAGGCACTCAGAG-3′ | 370 bp |
| Antisense:
5′-TGACAGGTCCACATGGTCTTCC-3′ |
| GAPDH | Sense:
5′-ATGTTCGTCATGGGTGTGAACCA-3′ | 351 bp |
| Antisense:
5′-TGGCAGGTTTTTCTAGACGGCAG-3′ |
Taq polymerase, 10X PCR buffer, deoxynucleotide
triphosphate mixture, MgCl2 solution (Applied
Biosystems, Thermo Fisher Scientific, Inc.) and cDNA template were
used for PCR (Veriti 96 well Thermal cycler, Applied Biosystems)
according to the manufacturer's instructions. PCR conditions were
as follows: Denaturation step at 95°C followed by 35 cycles of
denaturation at 95°C for 30 sec, annealing at 58°C for 30 sec, and
extension at 72°C for 30 sec, with a final extension at 72°C for 10
min. PCR products were run on a 2.0% agarose gel (Nusieve 3:1
Agarose; Lonza, Rockland, ME, USA) and gene bands were compared to
100-bp ladders (DAWINBIO, Hanam, Gyeonggi, Republic of Korea). Gels
were scanned and the density of the bands was quantified using gel
doc (Gel DocTM 2000 system, Bio-Rad Laboratories, Inc., Hercules,
CA, USA).
Statistical analysis
All data were analyzed with GraphPad Prism 5.0
software (GraphPad Inc., La Jolla, CA, USA). Values are expressed
as the mean ± standard deviation. Statistical analysis was
performed using Student's t-test or one-way analysis of variance,
followed by Dunnett's multiple comparisons tests. P<0.05 was
considered to indicate a statistically significant difference
between values.
Results
Concentration of isoflavone derivatives
in SMD
To determine the concentration of total isoflavones,
SMD was subjected to analytical HPLC. Twelve isoflavone isomers,
including aglycones, β-glucosides, 7′-O-acetylglucosides and
7′-O-malonylglucosides, were confirmed in SMD (Table II). The concentration of total
isoflavones in SMD was determined to be 165.123 mg/100 g,
indicating a high abundance of this compound type. The
concentrations of total 7′-O-acetylglucosides,
7′-O-malonylglucosides and aglycones were 23.862, 25.958 and
26.209 mg/100 g, respectively. Aglycones were most abundant among
all isoflavones in SMD. The content of daidzein, glycitein and GEN
was 1.988, 23.537, and 0.685 mg/100 g, respectively, with glycitein
being the most abundant among the total aglycones. Next, an SMD
solution with the same GEN content as that of the GEN solution used
(0.027 mg/ml) was prepared in order to compare their effects. These
results indicated that following digestion by bacterial
β-glucosidases in the body, soy milk may have enhanced biological
activities due to elevated aglycone contents. The expected aglycone
concentrations following conversion with β-glucosidases were based
on molecular weight and conversion coefficients (daidzin, 0.611;
glycitin, 0.637; genistin, 0.625; M-daidzin, 0.506; M-glycitin,
0.534; M-genistin, 0.625; A-daidzin, 0.555; A-glycitin, 0.582; and
A-genistin, 0.625).
SMD and its isoflavone components inhibit
the proliferation of prostate cancer cells
To determine the anti-proliferative effects of SMD
and its isoflavone component, an in vitro MTT assay was
performed using the LNCaP prostate cancer cell line (Fig. 1A). The cell viability was
significantly increased by 45±0.18% upon treatment with E2 compared
with vehicle treatment (0.1% DMSO). However, GEN and SMD reduced
the cell viability by 73.2±0.03 and 74.8±0.09%, respectively. These
results showed that E2 has estrogenic activity in LNCaP cells
expressing ERβ. Furthermore, GEN and SMD were demonstrated to
inhibit LNCaP cell growth.
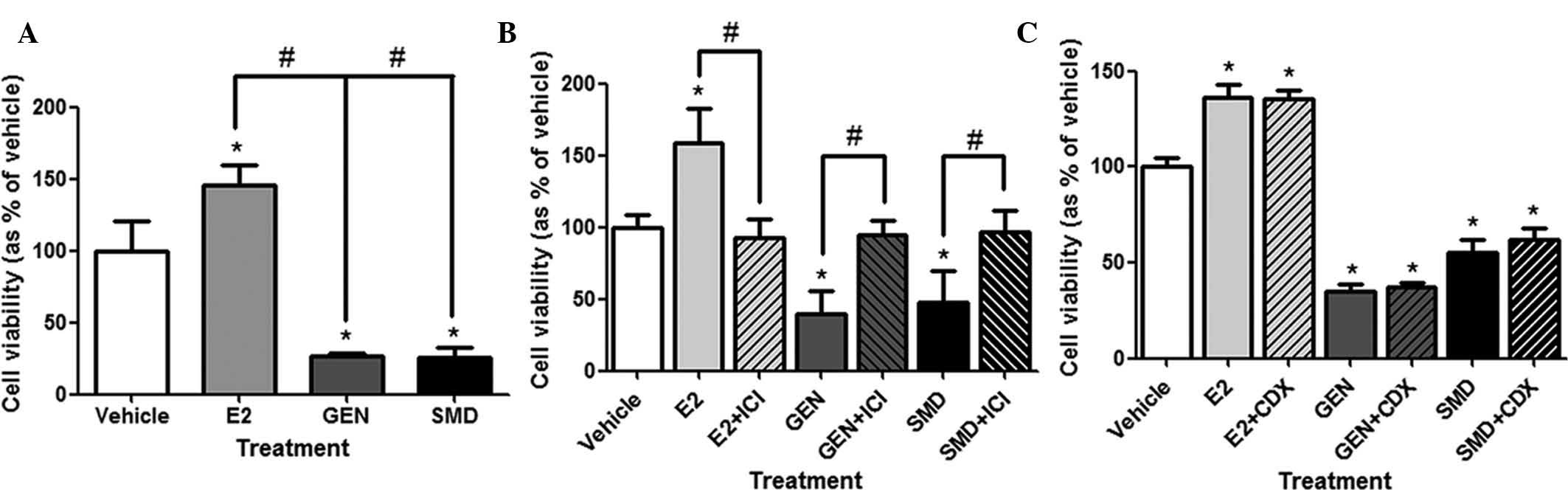 | Figure 1Effects of SMD and GEN in LNCaP cell
proliferation via ER signaling pathway. (A) Inhibitory effects of
phytoestrogens in SMD were analyzed using the MTT assay. Cells were
treated with vehicle (0.1% dimethyl sulfoxide), E2
(2.7×10−7 mg/ml), GEN (2.7×10−2 mg/ml) or SMD
(0.79 mg/ml) for five days. *P<0.05 vs. vehicle;
#P<0.05 vs. E2 group. (B) To evaluate the effects of
E2, GEN and SMD on cell growth in association with ER signaling,
the ER antagonist ICI 182,780 was used. LNCaP cells were treated
with vehicle, E2, GEN or SMD as above with optional co-treatment
with ICI 182,780 (6.1×10−6 mg/ml). (C) To determine
whether or not the effects of E2, GEN and SMD are associated with
the andogen receptor, cells were treated with CDX as above. LNCaP
cells were treated with vehicle, E2, GEN or SMD as above with
optional co-treatment with casodex (4.3×10−3 mg/ml).
Values are expressed as the mean ± standard deviation.
*P<0.05 vs. vehicle; #P<0.05 as indicated. ER,
estrogen receptor; SMD, soy milk digestion extract; E2,
17β-estradiol; GEN, genistein; CDX, carsodex (bicalutamide);
ICI, ICI 182,780. |
SMD inhibits prostate cancer cell growth
through ER
To determine whether the effects of E2, GEN and SMD
on LNCaP cell proliferation are mediated by ER signaling, the
typical ER antagonist ICI 182,780 was applied in combination with
E2, GEN or SMD (Fig. 1B). The
increases in cell viability by E2 treatment and the decreases in
cell viability by GEN and SMD treatment were completely abrogated
by co-treatment with ICI 182,780. This result indicated that the
cell proliferative effect of E2 as well as inhibitory effects of
GEN and SMD are mediated via ER signaling.
To determine whether the effects of SMD and its
isoflavone components are associated with the AR, cell
proliferation was assessed with co-treatment with the anti-androgen
CDX (Fig. 1C). E2-induced cell
proliferation as well as the anti-proliferative effects of GEN and
SMD were unaltered by CDX, suggesting that AR-associated signaling
is not involved.
SMD activates ERβ and deactivates PSA in
LNCaP cells
To determine the effects of SMD on the expression of
ERβ, AR and PSA in LNCaP cells, RT-PCR analysis was performed
(Fig. 2). The mRNA levels of ERβ
were significantly reduced by E2, while SMD and GEN significantly
increased ERβ expression compared with that in the DMSO-treated
group. The expression of AR was unaltered by ER, GEN and SMD,
indicating that they did not interact with the AR. The expression
of PSA, which is an gene upregulated by androgens, was
significantly reduced by GEN and SMD compared to that in the DMSO
group, whereas it was unaltered by E2. These results suggested that
E2-induced cell proliferation may be attributable to the inhibition
of ERβ gene expression, and that the anti-cancer activities of GEN
and SMD may be based on their ability to significantly enhance the
expression of ERβ and reduce the expression of PSA.
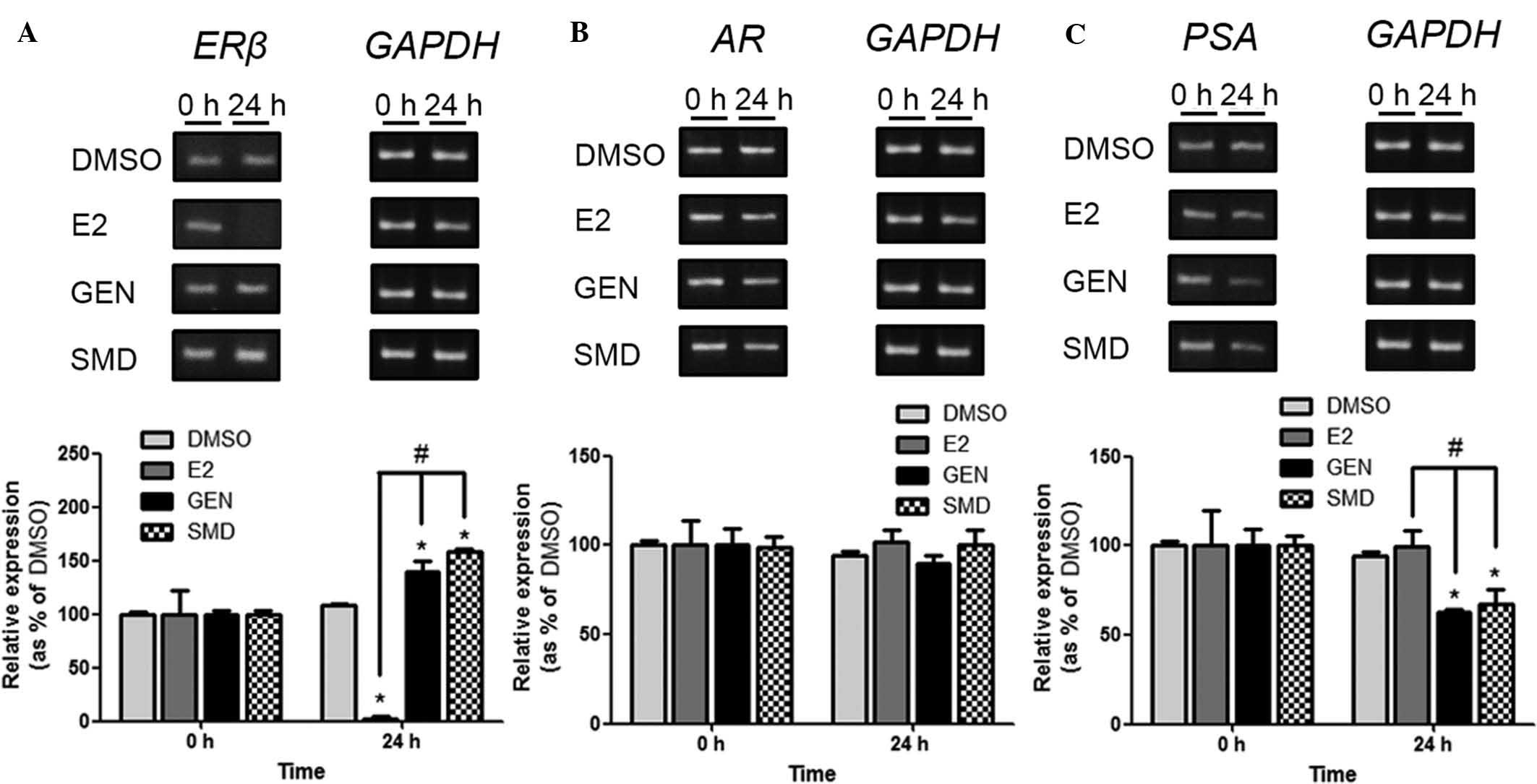 | Figure 2Effects of SMD on mRNA expression
levels of (A) ERβ, (B) AR and (C) PSA genes. LNCaP cells
(3.5×106/well) were treated with (0.1% DMSO), E2
(2.7×10−7 mg/ml), GEN (2.7×10−2 mg/ml) or SMD
(0.79 mg/ml) and mRNA expression levels of target genes were
determined by semi-quantitative reverse-transcription polymerase
chain reaction analysis. Values are expressed as the mean ±
standard deviation. *P<0.05 vs. DMSO group;
#P<0.05 vs. E2 group. ER, estrogen receptor; AR,
androgen receptor; PSA, prostate cancer-specific antigen; SMD, soy
milk digestion extract; E2, 17β-estradiol; GEN, genistein;
DMSO, dimethyl sulfoxide. |
SMD affects the mRNA expression of cell
cycle-regulatory genes
The effects of SMD, GEN and ERβ on the mRNA
expression levels of cell cycle-associated genes in LNCaP were then
assessed by semi-quantitative RT-PCR on total RNA samples isolated
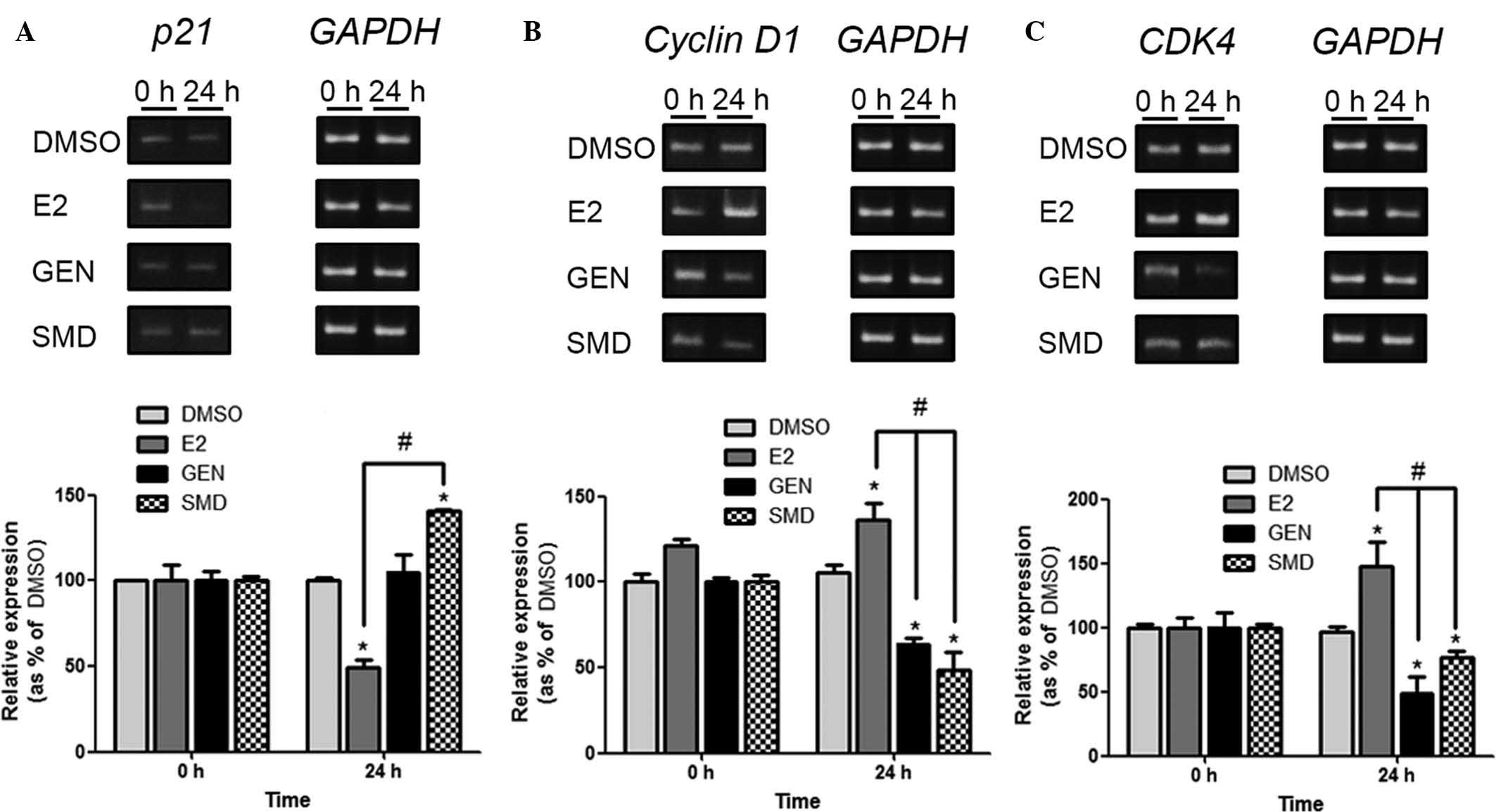
from cells at 0 and 24 h. As shown in Fig. 3, the gene expression of Cyclin D1
and CDK4 was significantly elevated by E2, while being
significantly reduced by GEN and SMD compared to that in the DMSO
group. The mRNA expression levels of p21 were significantly
elevated by SMD and reduced by E2, while remaining unaffected by
GEN compared to the DMSO group. These results indicated that the
gene expression of Cyclin D1, a gene inducing cell cycle
progression, was inhibited by the upregulated p21, a tumor
suppressor gene, upon SMD treatment. The anti-proliferative effects
of SMD on LNCaP prostate cancer cells are therefore likely to be
based on affecting these signaling mechanisms of cell cycle
regulatory genes.
Discussion
Soy milk, which is a product of whole soybeans,
contains high amounts of protein, iron and niacin, and is free from
cholesterol and lactose. In addition, soy-derived foods such as soy
milk have been indicated to exert effects against osteoporosis as
well as preventive effects against breast (31) and prostate cancers (23). Therefore, soy milk is an
alternative to cow's milk with additional health benefits (18). Soy milk and soybeans are the most
common and important dietary sources of isoflavones. Isoflavones, a
sub-group of phytoestrogens, are diphenolic compounds present in
plants, including soybeans, alfalfa, red clover and peanuts
(32). The major isoflavones in
soybeans are GEN, daidzein and glycitein. Soy isoflavones were
demonstrated to have biological activities, including anti-oxidant
and anti-inflammatory effects as well as anti-proliferative effects
against cancer cells (33,34). While soy isoflavones and their
metabolites have been indicated to be beneficial for the prevention
or treatment of prostate cancer, the mechanisms of action have
remained to be fully elucidated.
E2 (17β-estradiol) is a steroid hormone and exerts a
strong estrogenic effect. In addition, it was reported to be linked
to several types of cancer (12–15,25,26).
GEN, a major isoflavone, is phytoestrogen and xenoestrogen, which
influences multiple biochemical functions. E2 and GEN bind to the
estrogen receptor, but result in opposing effects. E2 promotes the
growth of cancer cells and GEN is reduces it (12,13).
In a previous study, the growth of LNCaP human prostate cancer
cells was significantly stimulated by E2 (35). In addition, E2 was shown to
contribute to the risk of prostate cancer, whereas the
phytochemical GEN derived from soy decreased the risk of prostate
cancer (10). For this reason, the
present study hypothesized that cell growth of prostate cancer
cells was promoted by E2 and thus was used to compare the effects
of isoflavones in the SMD. SMD contained a number of isoflavones,
including GEN, according to the results of HPLC analysis.
Therefore, it was hypothesized that GEN and SMD would have similar
effects.
The present study examined the mechanisms underlying
the preventative effects of SMD and its component GEN on prostate
cancer cell proliferation and the expression of ERβ, which is
associated with prostate cancer progression. The androgen-dependent
LNCaP human prostate cancer cell line, which expresses ERβ at high
levels, was adopted for evaluation of the tumor inhibitory effects
of SMD along with E2 and GEN. The ERβ gene is expressed in prostate
epithelial cells and has anti-proliferative and pro-apoptotic roles
in cellular homeostasis (36). Soy
isoflavones are structurally similar to E2 and are capable of
binding to ERs (32). Soy
isoflavones can be divided into three chemical groups: Aglycones
[daidzein (4′,7-dihydroxy-isoflavone), GEN
(4′,5,7-trihydroxyisoflavone) and glycitein
(4′,7-dihydroxy-6-methoxyisoflavone)], β-glycosides [daidzin, GEN
and glycitin], and further into 7′-O-acetylglucosides and
7′-O-malonylglucosides (37). Based on these facts, the present
study hypothesized that soy isoflavones contained in SMD may bind
to ERβ in LNCaP cells. Soy milk was digested and hydrolyzed using a
pepsin suspension and pancreatin-bile salt mixture for extraction
of aglycone isoflavones, which are bioavailable in
vitro.
The presence of 12 isoflavone isomers was confirmed
in SMD, including aglycone, and their concentration was determined.
The effects of E2, GEN and SMD on LNCaP cell proliferation and ER
signaling were evaluated using a cell proliferation assay and
RT-PCR. The results showed that the viability and growth of LNCaP
cells were promoted by E2, while being inhibited by SMD and GEN.
Co-treatment with the ER inhibitor ICI 182,780 inhibited the
effects of E2, GEN or SMD on the proliferation of LNCaP cells,
whereas co-treatment with AR inhibitor CDX had no obvious effects.
These results suggested that the effects of SMD are attributable to
ER, but not AR signaling. In support of this, expression of ERβ was
significantly elevated by SMD. mRNA expression of ERβ has been
shown to be induced by SMD, and the combination of GEN, daidzein,
and glycitein increases selectivity for ERβ binding. In addition,
expression of PSA, which can be upregulated by androgens, was
reduced by SMD, suggesting that SMD may potentially act as an
anti-androgen.
The cell cycle has two major checkpoints: The
G1/S checkpoint, which controls entry into S phase, and
the G2/M checkpoint (38). Regulation of the cell cycle is
closely linked to cancer cell proliferation and differentiation
(14). Soy isoflavones of SMD can
modulate the expression of genes that control the cell cycle and
cell survival via the ERβ pathway, which may have inhibitory
effects on G2 and M phases of the cell cycle (39). Cyclin D1 is a key regulator of cell
cycle progression. Cyclins bind to their corresponding CDKs and
form corresponding cyclin-CDK complexes. Cyclin D1 is known to
function as a regulatory sub-unit of CDK4 or CDK6, whose activity
is required for the G0/G1-S phase transition
(12,14,38,40).
In the present study, gene expression levels of Cyclin D1 and CDK4
were significantly elevated by E2 compared to those in the DMSO
group, whereas they were considerably decreased by SMD. The tumor
suppressor gene p21 directly regulates the G1/S phase
checkpoint by binding to cyclin-CDK complexes and leading to cell
cycle arrest in G1 phase. In the present study, the expression of
p21 was markedly increased by SMD. Therefore, SMD may potentially
regulate the cell cycle through suppression of the Cyclin D1-CDK4
complex by upregulating p21 (Fig.
4).
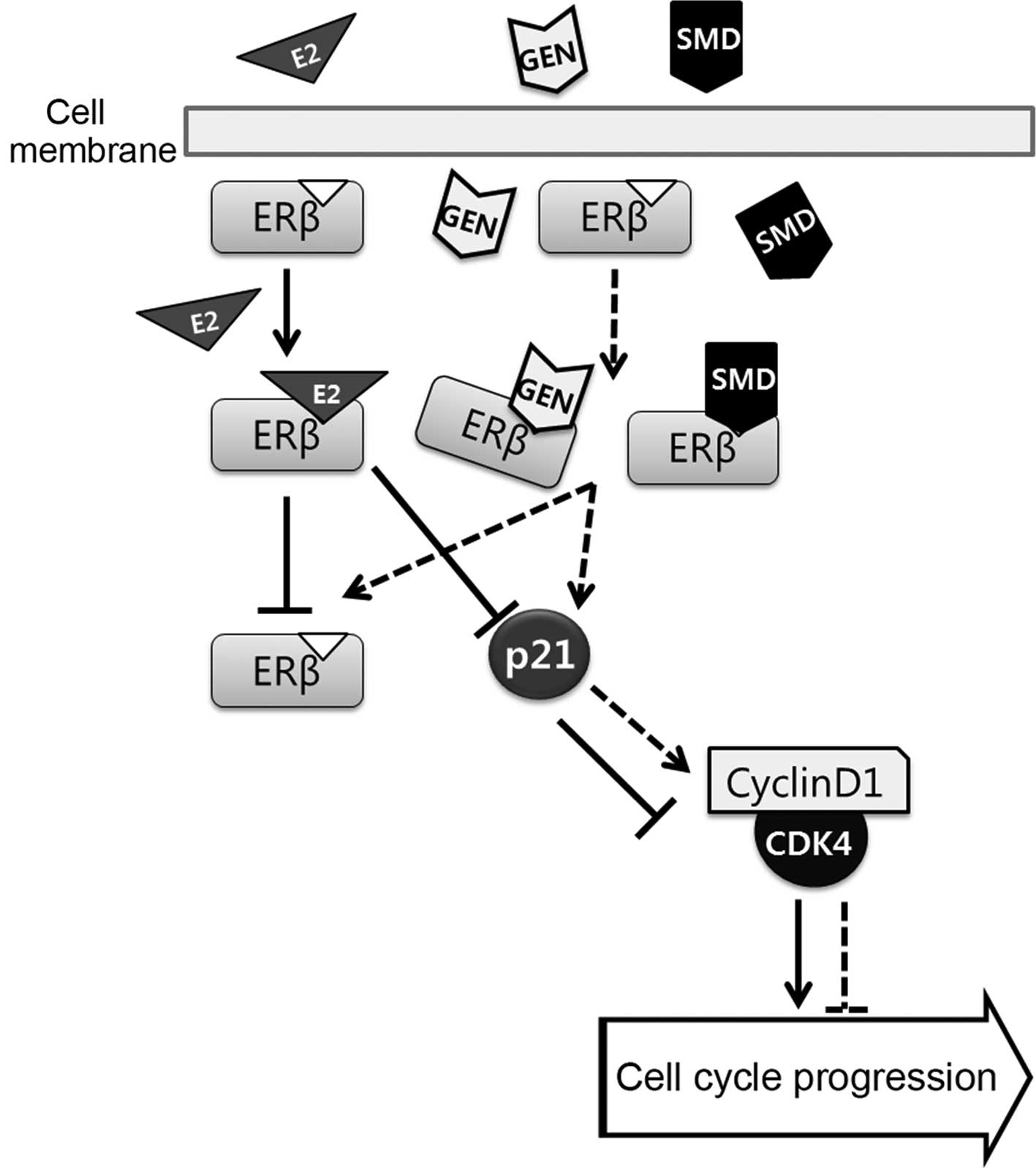 | Figure 4Role of SMD in the progression of
E2-dependent LNCaP prostate cancer cells. LNCaP cells were treated
with E2, GEN, and SMD. GEN, a phytoestrogen, and SMD, a combination
of soy isoflavones, bound to ERβ due to structural similarity to
E2. E2 reduced the translation of ERβ and p21, a tumor suppressor
gene, thus increasing the cyclin-CDK complex. The cell cycle was
promoted by E2 in LNCaP cells, while it was inhibited by GEN and
SMD through ERβ, which bound to GEN and SMD, and then activated p21
to suppress the cyclin-CDK complex. As a result, SMD inhibited the
growth of LNCaP cells by promoting the ERβ pathway and blocking
cell cycle progression. SMD, soy milk digestion extract; E2,
17β-estradiol; GEN, genistein; CDK, cyclin-dependent
kinase. |
In conclusion, the present study demonstrated that
aglycone isoflavones from digested soy milk display inhibitory
effects via ERβ signaling in LNCaP prostate cancer cells. SMD was
found to contain diverse aglycone isoflavones produced by digestive
processes and bacterial β-glucosidases, and inhibited the growth of
LNCaP cancer cells by promoting the expression of ERβ and
inhibiting the cell cycle. Therefore, soy milk may represent a
nutriceutical for the prevention and inhibition of prostate
cancer.
Acknowledgments
This research was supported by the Ministry of
Trade, Industry and Energy and the Korea Institute for Advancement
of Technology through promoting Regional specialized Industry. The
present study was supported by the High Value-added Food Technology
Development Program, Ministry of Agriculture, Food and Rural
Affairs (grant no. 113034-3).
References
|
1
|
Ogimoto I, Shibata A and Fukuda K: World
Cancer Research fund/american institute of cancer research 1997
recommendations: Applicability to digestive tract cancer in Japan.
Cancer Causes Control. 11:9–23. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Torfadottir JE, Steingrimsdottir L, Mucci
L, Aspelund T, Kasperzyk JL, Olafsson O, Fall K, Tryggvadottir L,
Harris TB, Launer L, et al: Milk intake in early life and risk of
advanced prostate cancer. Am J Epidemiol. 175:144–153. 2012.
View Article : Google Scholar :
|
|
3
|
Chung MS, Lee SH, Lee DH, Kim SJ, Kim CS,
Lee KS, Jung JI, Kim SW, Lee YS and Chung BH: Practice patterns of
Korean urologists for screening and managing prostate cancer
according to PSA level. Yonsei Med J. 53:1136–1141. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Castro B, Sánchez P, Torres JM, Preda O,
del Moral RG and Ortega E: Bisphenol A exposure during adulthood
alters expression of aromatase and 5α-reductase isozymes in rat
prostate. PLoS One. 8:e559052013. View Article : Google Scholar
|
|
5
|
Saad F and Asselah J: Chemotherapy for
prostate cancer: Clinical practice in Canada. Can Urol Assoc J.
7:S5–S10. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Weng C, Cai J, Wen J, Yuan H, Yang K,
Imperato-McGinley J and Zhu YS: Differential effects of estrogen
receptor ligands on regulation of dihydrotestosterone-induced cell
proliferation in endothelial and prostate cancer cells. Int J
Oncol. 42:327–337. 2013.
|
|
7
|
Basak S, Pookot D, Noonan EJ and Dahiya R:
Genistein down-regulates androgen receptor by modulating
HDAC6-Hsp90 chaperone function. Mol Cancer Ther. 7:3195–3202. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Matsumura K, Tanaka T, Kawashima H and
Nakatani T: Involvement of the estrogen receptor beta in
genistein-induced expression of p21 (waf1/cip1) in PC-3 prostate
cancer cells. Anticancer Res. 28:709–714. 2008.PubMed/NCBI
|
|
9
|
Messina MJ: Legumes and soybeans: Overview
of their nutritional profiles and health effects. Am J Clin Nutr.
70(Suppl 3): S439–S450. 1999.
|
|
10
|
Smith S, Sepkovic D, Bradlow HL and Auborn
KJ: 3,3′-Diindolylmethane and genistein decrease the adverse
effects of estrogen in LNCaP and PC-3 prostate cancer cells. J
Nutr. 138:2379–2385. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ahmad A, Biersack B, Li Y, Bao B, Kong D,
Ali S, Banerjee S and Sarkar FH: Perspectives on the role of
isoflavones in prostate cancer. AAPS J. 15:991–1000. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hwang KA, Kang NH, Yi BR, Lee HR, Park MA
and Choi KC: Genistein, a soy phytoestrogen, prevents the growth of
BG-1 ovarian cancer cells induced by 17β-estradiol or bisphenol A
via the inhibition of cell cycle progression. Int J Oncol.
42:733–740. 2013.
|
|
13
|
Hwang KA, Park MA, Kang NH, Yi BR, Hyun
SH, Jeung EB and Choi KC: Anticancer effect of genistein on BG-1
ovarian cancer growth induced by 17 β-estradiol or bisphenol A via
the suppression of the crosstalk between estrogen receptor α and
insulin-like growth factor-1 receptor signaling pathways. Toxicol
Appl Pharmacol. 272:637–646. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kang NH, Hwang KA, Kim TH, Hyun SH, Jeung
EB and Choi KC: Induced growth of BG-1 ovarian cancer cells by
17β-estradiol or various endocrine disrupting chemicals was
reversed by resveratrol via downregulation of cell cycle
progression. Mol Med Rep. 6:151–156. 2012.PubMed/NCBI
|
|
15
|
Kang NH, Hwang KA, Lee HR, Choi DW and
Choi KC: Resveratrol regulates the cell viability promoted by
17β-estradiol or bisphenol A via down-regulation of the cross-talk
between estrogen receptor α and insulin growth factor-1 receptor in
BG-1 ovarian cancer cells. Food Chem Toxicol. 59:373–379. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Seo YJ, Kim BS, Chun SY, Park YK, Kang KS
and Kwon TG: Apoptotic effects of genistein, biochanin-A and
apigenin on LNCaP and PC-3 cells by p21 through transcriptional
inhibition of polo-like kinase-1. J Korean Med Sci. 26:1489–1494.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu ZM, Ho SC, Chen YM and Woo J: A
six-month randomized controlled trial of whole soy and isoflavones
daidzein on body composition in equol-producing postmenopausal
women with prehypertension. J Obes. 3597632013.PubMed/NCBI
|
|
18
|
Matthews VL, Knutsen SF, Beeson WL and
Fraser GE: Soy milk and dairy consumption is independently
associated with ultrasound attenuation of the heel bone among
postmenopausal women: The adventist health study-2. Nutr Res.
31:766–775. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Keshavarz SA, Nourieh Z, Attar MJ and
Azadbakht L: Effect of soymilk consumption on waist circumference
and cardiovascular risks among overweight and obese female adults.
Int J Prev Med. 3:798–805. 2012.PubMed/NCBI
|
|
20
|
Jacobsen BK, Knutsen SF and Fraser GE:
Does high soy milk intake reduce prostate cancer incidence? The
adventist health study (United States). Cancer Causes Control.
9:553–557. 1998. View Article : Google Scholar
|
|
21
|
Sugiyama Y, Masumori N, Fukuta F, Yoneta
A, Hida T, Yamashita T, Minatoya M, Nagata Y, Mori M, Tsuji H, et
al: Influence of isoflavone intake and equol-producing intestinal
flora on prostate cancer risk. Asian Pac J Cancer Prev. 14:1–4.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cassidy A, Brown JE, Hawdon A, Faughnan
MS, King LJ, Millward J, Zimmer-Nechemias L, Wolfe B and Setchell
KD: Factors affecting the bioavailability of soy isoflavones in
humans after ingestion of physiologically relevant levels from
different soy foods. J Nutr. 136:45–51. 2006.
|
|
23
|
deVere White RW, Tsodikov A, Stapp EC,
Soares SE, Fujii H and Hackman RM: Effects of a high dose,
aglycone-rich soy extract on prostate-specific antigen and serum
isoflavone concentrations in men with localized prostate cancer.
Nutr Cancer. 62:1036–1043. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rodriguez-Roque MJ, Rojas-Graü MA,
Elez-Martinez P and Martin-Belloso O: Soymilk phenolic compounds,
isoflavones and antioxidant activity as affected by in vitro
gastrointestinal digestion. Food Chem. 136:206–212. 2013.
View Article : Google Scholar
|
|
25
|
Park MA, Hwang KA and Choi KC: Diverse
animal models to examine potential role (s) and mechanism of
endocrine disrupting chemicals on the tumor progression and
prevention: Do they have tumorigenic or anti-tumorigenic property?
Lab Anim Res. 27:265–273. 2011. View Article : Google Scholar
|
|
26
|
Lee HR, Kim TH and Choi KC: Functions and
physiological roles of two types of estrogen receptors, ERα and
ERβ, identified by estrogen receptor knockout mouse. Lab Anim Res.
28:71–76. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hwang KA, Park SH, Yi BR and Choi KC: Gene
alterations of ovarian cancer cells expressing estrogen receptors
by estrogen and bisphenol a using microarray analysis. Lab Anim
Res. 27:99–107. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cheng J, Lee EJ, Madison LD and Lazennec
G: Expression of estrogen receptor beta in prostate carcinoma cells
inhibits invasion and proliferation and triggers apoptosis. FEBS
Lett. 566:169–172. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tate PL, Bibb R and Larcom LL: Milk
stimulates growth of prostate cancer cells in culture. Nutr Cancer.
63:1361–1366. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kang NH, Hwang KA, Yi BR, Lee HJ, Jeung
EB, Kim SU and Choi KC: Human amniotic fluid-derived stem cells
expressing cytosine deaminase and thymidine kinase inhibits the
growth of breast cancer cells in cellular and xenograft mouse
models. Cancer Gene Ther. 19:412–419. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Takagi A, Kano M and Kaga C: Possibility
of breast cancer prevention: use of soy isoflavones and fermented
soy beverage produced using probiotics. Int J Mol Sci.
16:10907–10920. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Butkutė B, Lemežienė N, Dabkevičienė G, et
al: Source of variation of isoflavone concentrations in perennial
clover species. Pharmacogn Mag. 10(Suppl 1): S181–S188. 2014.
View Article : Google Scholar
|
|
33
|
Mahmoud AM, Yang W and Bosland MC: Soy
isoflavones and prostate cancer: A review of molecular mechanisms.
J Steroid Biochem Mol Biol. 140:116–132. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang Q, Ge X, Tian X, Zhang Y, Zhang J and
Zhang P: Soy isoflavone: The multipurpose phytochemical (Review).
Biomed Rep. 1:697–701. 2013.
|
|
35
|
Castagnetta LA, Miceli MD, Sorci CM,
Pfeffer U, Farruggio R, Oliveri G, Calabrò M and Carruba G: Growth
of LNCaP human prostate cancer cells is stimulated by estradiol via
its own receptor. Endocrinology. 136:2309–2319. 1995.PubMed/NCBI
|
|
36
|
Gao AM, Ke ZP, Shi F, Sun GC and Chen H:
Chrysin enhances sensitivity of BEL-7402/ADM cells to doxorubicin
by suppressing PI3K/Akt/Nrf2 and ERK/Nrf2 pathway. Chem Biol
Interact. 206:100–108. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chang Y and Choue R: Plasma
pharmacokinetics and urinary excretion of isoflavones after
ingestion of soy products with different aglycone/glucoside ratios
in South Korean women. Nutr Res Pract. 7:393–399. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Liu D, Liu J, Lin B, et al: Lewis y
regulate cell cycle related factors in ovarian carcinoma cell RMG-I
in Vitro via ERK and Akt signaling pathways. Int J Mol Sci.
13:828–839. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Li W, Winters A, Poteet E, Ryou MG, Lin S,
Hao S, Wu Z, Yuan F, Hatanpaa KJ, Simpkins JW and Yang SH:
Involvement of estrogen receptor β5 in the progression of glioma.
Brain Res. 1503:97–107. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Sherr CJ: The Pezcoller lecture: Cancer
cell cycles revisited. Cancer Res. 60:3689–3695. 2000.PubMed/NCBI
|