Introduction
Loss of skeletal muscle mass, also known as atrophy,
may occur in normal aging-related conditions or in chronic
pathological conditions, including myopathy, denervation-associated
atrophy, cachexia and obesity (1,2).
Skeletal muscle atrophy is associated with increased fatigability
and metabolic health problems leading to a reduced quality of life,
which represents a major public health burden in several countries.
Therefore, great efforts have been made to identify therapeutic
tools to prevent or retard muscle atrophy. Muscle regeneration is a
coordinated process that involves proliferation and differentiation
of muscle progenitor cells. Skeletal myoblast differentiation is a
multistep process that is associated with cell cycle exit,
muscle-specific gene expression, and formation of multinucleated
myotubes via myoblast fusion (3).
Myogenesis is well-orchestrated by the myogenic basic
helix-loop-helix transcription factors, including MyoD, myogenin
and myogenic factor 5 (4). Mice
lacking MyoD exhibit delayed myogenesis in the limbs and branchial
arches (2). The activation of MyoD
is a key regulatory step for the induction of myoblast
differentiation. Notably, p38 mitogen-activated protein kinases
(MAPK) have a fundamental role in muscle differentiation via the
activation of chromatin remodeling proteins and myogenic
transcription factors, such as MyoD (5). p38 MAPK induces the
heterodimerization of MyoD with E proteins, thus resulting in
upregulation of muscle-specific genes, including myogenin and
myosin heavy chain (MHC) (6,7).
Various promyogenic cell surface signaling pathways, such as
Cdo-mediated cell adhesion signaling, activate p38 MAPK thereby
inducing myoblast differentiation (8).
Corydalis tuber, which is the rhizome of
Corydalis turtschaninovii, has been used to treat
inflammatory, hemorheological and allergic diseases in Korea and
China (9,10). It has previously been reported that
several alkaloid compounds can be isolated from Corydalis
tuber (10,11). Among these, dehydrocorydaline (DHC)
has been demonstrated to suppress the elevated mitochondrial
membrane potential in lipopolysaccharide-stimulated macrophages
(12), and to inhibit
proliferation of breast cancer cells by inducing apoptosis
(13). However, the effects of DHC
on myoblast differentiation have yet to be described.
In the present study, DHC, which is an isoquinoline
alkaloid, was selected in a screening of natural phytochemicals
purified from the Corydalis tuber (Papaveraceae) for the
activation of MyoD-responsive reporters and induction of MHC in
myoblasts. Subsequently, the effects of DHC on myoblast
differentiation and the underlying regulatory mechanisms were
investigated. Treatment of C2C12 myoblasts with DHC enhanced the
differentiation-linked activation of p38 MAPK and elevated the
interaction of MyoD with E proteins, thus resulting in promotion of
myoblast differentiation. In addition, DHC treatment rescued p38
MAPK activation and multinucleated myotube formation in
Cdo-depleted C2C12 cells. The present study is the first, to the
best of our knowledge, to report that phytochemical DHC promotes
MyoD-mediated myogenesis via activation of the p38 MAPK promyogenic
signaling pathway.
Materials and methods
Preparation of DHC from Corydalis
tuber
The Corydalis tuber was purchased from
Kyungdong Herb Medicine Market (Seoul, South Korea) and was
authenticated by Professor Dae-Keun Kim (Woosuk University, Jeonju,
South Korea). A voucher specimen (KHU070123) was reserved at the
Laboratory of Natural Products Chemistry (Kyung Hee University,
Yongin, South Korea). The powdered tuber (5 kg) was extracted with
80% aqueous MeOH (5.0 Lx2) at room temperature to give a dark
brownish extract (347 g). The methanol extract was then poured into
acidic water (pH 2.5; 2.0 L) and was washed twice with EtOAc (2.0
Lx2). The pH of the aqueous layer was increased to pH 12.0 using
20% NaOH solution, and was then extracted with EtOAc (2.0 Lx2) and
n-BuOH (1.5 Lx2) successively. The n-BuOH soluble
layer (CTB; 58 g) was subjected to SiO2 column
chromatography (70–230 mesh; 320 g) using CHCl3-EtOH
(10:1→5:1) and CHCl3-MeOH (3:1) as eluent to give eight
fractions (CTB1-CTB8). The second fraction (CTB2; 2.0 g) was
subjected to further SiO2 column chromatography (100 g)
and was eluted with CHCl3-MeOH (10:1) to provide pure
DHC (1.4 g). The structure of DHC was analyzed by nuclear magnetic
resonance (NMR; Varian Unity Inova AS-400 FT-NMR spectrometer;
Varian Medical Systems, Inc., Palo Alto, CA, USA) and mass
spectrometry (MS; JEOL JMSAX-700: JEOL Ltd., Tokyo, Japan).
Cell culture and expression vectors
The C2C12 myoblast cells, 10T1/2 embryonic
fibroblast cells and embryonic kidney 293T cells were cultured as
described previously (14). To
induce differentiation of C2C12 myoblasts, cells at near confluence
were removed from growth medium (GM), which consisted of Dulbecco's
modified Eagle's medium (DMEM) containing 15% fetal bovine serum
(FBS; Hyclone GE Healthcare Life Sciences, Logan, UT, USA), and
were cultured in differentiation medium (DM), which consisted of
DMEM containing 2% horse serum (HS; Hyclone; GE Healthcare Life
Sciences). Myotube formation was observed after 2 or 3 days of
differentiation. The efficiency of myotube formation was quantified
using a transient differentiation assay, as previously described
(14). To generate C2C12 cells
that transiently overexpressed short hairpin RNAs (shRNAs) against
Cdo, the cells were transfected with the indicated expression
vector using Lipofectamine® 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.) with 10T1/2 and 293T cells seeded at
3×105/10 cm cell culture plate containing DMEM, 10% FBS
and antibiotics, and the following day the cells were transfected
with 10 μg of MyoD construct or pcDNA vector as a control
for 2 days at 37°C. To produce C2C12 cell lines that stably
overexpress shRNA against Cdo, C2C12 cells were seeded at
1×105/10 cm cell culture plate containing DMEM, 15% FBS
and antibiotics. The cells were transfected for 1 day at 37°C with
10 μg Cdo construct and pSuper vectors served as a control.
Cultures were subsequently selected in puromycin-containing medium.
Puromycin-resistant cells were pooled and analyzed. Cdo shRNA was
purchased from Sigma-Aldrich (Merck Millipore, Darmstadt, Germany)
and its sequence is 5′-CAGCGTTGGTGCCGTTGTG-3′. pSuper was obtained
from Oligoengine (Seattle, WA, USA).
Western blot analysis and
immunoprecipitation
Western blot analysis was performed as described
previously (15). In C2C12
myoblasts, C2C12 cells were treated with indicated concentrations
of DHC and differentiation was induced for 2 or 3 days at 37°C.
10T1/2 and 293T cells were treated with 500 nM DHC. Following
incubation for 1 day at 37°C, cell lysates were subjected to
immunoprecipitation with E2A antibodies followed by immunoblotting
with MyoD antibodies. Briefly, cells were lysed in cell extraction
buffer [10 mM Tris-HCl (pH 8.0), 150 mM NaCl, 1 mM EDTA, 1% Triton
X-100] containing a complete protease inhibitor cocktail (Roche
Diagnostics, Indianapolis, IN, USA), and 30–50 μg proteins
were separated by 6–10% (6% for MHC, Cdo and pan-cadherin
antibodies, and 10% for all other antibodies) sodium dodecyl
sulfate-polyacrylamide gel electrophoresis. The primary antibodies
used were: Anti-MyoD (cat. no. sc-32758; mouse; 1:1,000),
anti-myogenin (cat. no. sc-576; rabbit; 1:1,000), anti-p38 MAPK
(cat. no. sc-728; anti-rabbit; 1:1,000), anti-E2A (cat. no. sc-763;
rabbit; 1:1,00) (all Santa Cruz Biotechnology, Inc., Dallas, TX,
USA), anti-phosphorylated (p)-p38 MAPK (p-p38; cat. no. 9211;
rabbit; 1:500, Cell Signaling Technology, Inc., Beverly, MA, USA),
anti-pan-Cadherin (cat. no. C3678; rabbit; Sigma-Aldrich; Merck
Millipore), anti-Cdo (cat. no. AF2429; mouse; R&D Systems,
Minneapolis, MN, USA) and anti-MHC (cat. no. MF20; mouse;
Developmental Studies Hybridoma Bank, Iowa City, IA, USA).
Horseradish peroxidase-conjugated secondary antibodies from Santa
Cruz Biotechnology, Inc. were diluted between 1:5,000 and 1:10,000
and used as follows: Goat anti-mouse IgG-HRP (cat. no. sc-2005);
goat anti-rabbit IgG-HRP (cat. no. sc-2004); and donkey anti-goat
IgG-HRP (cat. no. sc-2033). Primary antibodies were incubated
overnight at 4°C and secondary antibodies were incubated for 2–3 h
at room temperature. For the immunoprecipitation assay, 293T (3
×105/plate) or 10T1/2 cells (3 ×105/plate)
were transfected with 10 μg MyoD (based on the pcDNA vector,
accession number: NM-010866) using Lipofectamine 2000®
according to the manufacturer's protocol for 20 min at room
temperature and were treated with DHC. After 24 h of treatment of
C2C12, 10T1/2 and 293T cells with 500 nm DHC at 37°C, whole cell
extracts were incubated with anti-E2A and protein G agarose beads
(Roche Diagnostics) overnight at 4°C. Subsequently, the beads were
washed three times with extraction buffer, were resuspended in
extraction buffer, and the samples were analyzed by western
blotting.
Screening process
Phytochemicals purified from Corydalis tuber
were screened using MyoD luciferase assay and MHC western blotting.
For the MyoD luciferase assay, C2C12 cells were plated for 24 h in
24-well plates and transiently transfected with the 4RTK
reporter gene using Lipofectamine 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol.
Following incubation for 24 h, cells were treated with
phytochemicals (10 nM) from Corydalis tuber for an
additional 24 h. Luciferase activities were measured on a Berthold
luminometer, integrating light emission over 20 sec. Transfection
efficiencies were normalized by co-transfecting 50 ng of the
β-galactosidase plasmid, pCH110 (GE Healthcare Life Sciences). All
transfections were performed in duplicate a minimum of three times.
The second screening was conducted using MHC western blotting.
C2C12 cells were plated for 24 h in 6-well plates, and treated with
phytochemicals (10 nM) from Corydalis tuber in 2% horse
serum for an additional 48 h. The cells were lysed and western
blotting was performed. The primary antibody used was anti-MHC, and
quantification of the signal was performed using Image Gauge
software version 4.0 (Fujifilm, Tokyo, Japan).
Immunocytochemistry and confocal
microscopy
Immunostaining for MHC expression was performed as
described previously (14).
Briefly, C2C12 cells were treated with 125, 250 and 500 nM DHC for
3 days at 37°C, fixed with 4% paraformaldehyde, permeabilized with
0.1% Triton X-100 in phosphate-buffered saline (PBS), and were
blocked. Subsequently, the cells were stained with anti-MHC,
followed by incubation with an Alexa Fluor 594-conjugated secondary
antibody (cat. no. A11012; 1:500; Molecular Probes; Thermo Fisher
Scientific, Inc.). Primary antibodies were incubated overnight at
4°C and secondary antibodies were incubated for 2 h at room
temperature. Images were captured and processed using a Nikon
ECLIPSE TE-2000 U microscope and NIS-Elements F software (Nikon
Corporation, Tokyo, Japan). The quantitative differentiation assay
was performed at least three times. To observe the effects of DHC
on p38 phosphorylation in C2C12 cells, C2C12 cells were treated
with 500 nM DHC for 2 or 3 days at 37°C and were fixed with 4%
paraformaldehyde. Cultures were subsequently permeabilized with
0.1% Triton X-100 in PBS, blocked in 5% horse serum in 0.1% Triton
X-100 in PBS, and were incubated with anti-p-p38 followed by an
incubation with an Alexa Fluor 594-conjugated secondary antibody.
Nuclei were counterstained with 4′,6-diamidino-2-phenylindole
(DAPI; cat. no. sc-3598; Santa Cruz Biotechnology, Inc.). Images
were obtained using a Zeiss LSM-510 Meta confocal microscope (Carl
Zeiss AG, Oberkochen, Germany). Quantification of the p-p38
fluorescent signal was performed using Image Gauge software version
4.0.
Bromodeoxyuridine (BrdU) assay
C2C12 cells were cultured in 6-well plates and were
treated with dimethyl sulfoxide (DMSO) or 500 nM DHC in GM for 24 h
at 37°C. After 24 h of treatment, 10 μM BrdU (cat. no.
550891; 10 μg/ml; BD Biosciences, San Jose, CA, USA) was
added to the DMSO- or DHC-treated cells and was incubated for 30
min at 37°C. The cells were fixed and immunostained with anti-BrdU
(cat. no. sc-32323; mouse; 1:500; Santa Cruz Biotechnology, Inc.)
overnight at 4°C, followed by incubation for 2 h at room
temperature with a fluorescein isothiocyanate-conjugated secondary
antibody (cat. no. A11001; 1:500; Invitrogen; Thermo Fisher
Scientific, Inc.). Images were captured and processed using a Nikon
ECLIPSE TE-2000 U microscope and NIS-Elements F software. The
quantitative differentiation assay was performed at least three
times.
Statistical analysis
Data are presented as the mean ± standard deviation
from at least three independent experiments. Data were analyzed
using the Student's t-test. For comparisons between multiple
groups, statistical significance was determined using one-way
analysis of variance and the Scheffe test for post-hoc analysis.
Statistical analysis was conducted using SPSS 18.0 (SPSS, Inc.,
Chicago, IL, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results and Discussion
DHC enhances myogenic
differentiation
To identify effective activators of myoblast
differentiation, phytochemicals purified from Corydalis
tuber were screened, a luciferase assay was performed to measure
MyoD activity, and western blotting was conducted to detect the
expression levels of MHC. An active component was identified from
the alkaloidal fraction of Corydalis tuber (Fig. 1A). Its molecular weight was
determined to be 366 from electron ionization-MS spectra, and
1,2,4,5-tetrasubstituted benzene signals (7.18 and 6.94 ppm),
1,2,3,4-tetrasubstituted benzene signals (7.96 and 7.91 ppm), four
methoxy signals (4.28, 4.07, 4.00 and 3.95 ppm) and an allylmethyl
proton signal (2.98 ppm) were observed in 1H-NMR
spectrum. The same pattern of 13C-NMR spectroscopic data
also revealed the typical spectroscopic pattern of the isoquinoline
alkaloid. The structure of the active compound was finally
confirmed to be DHC by comparing the data with those reported in
the literature (16).
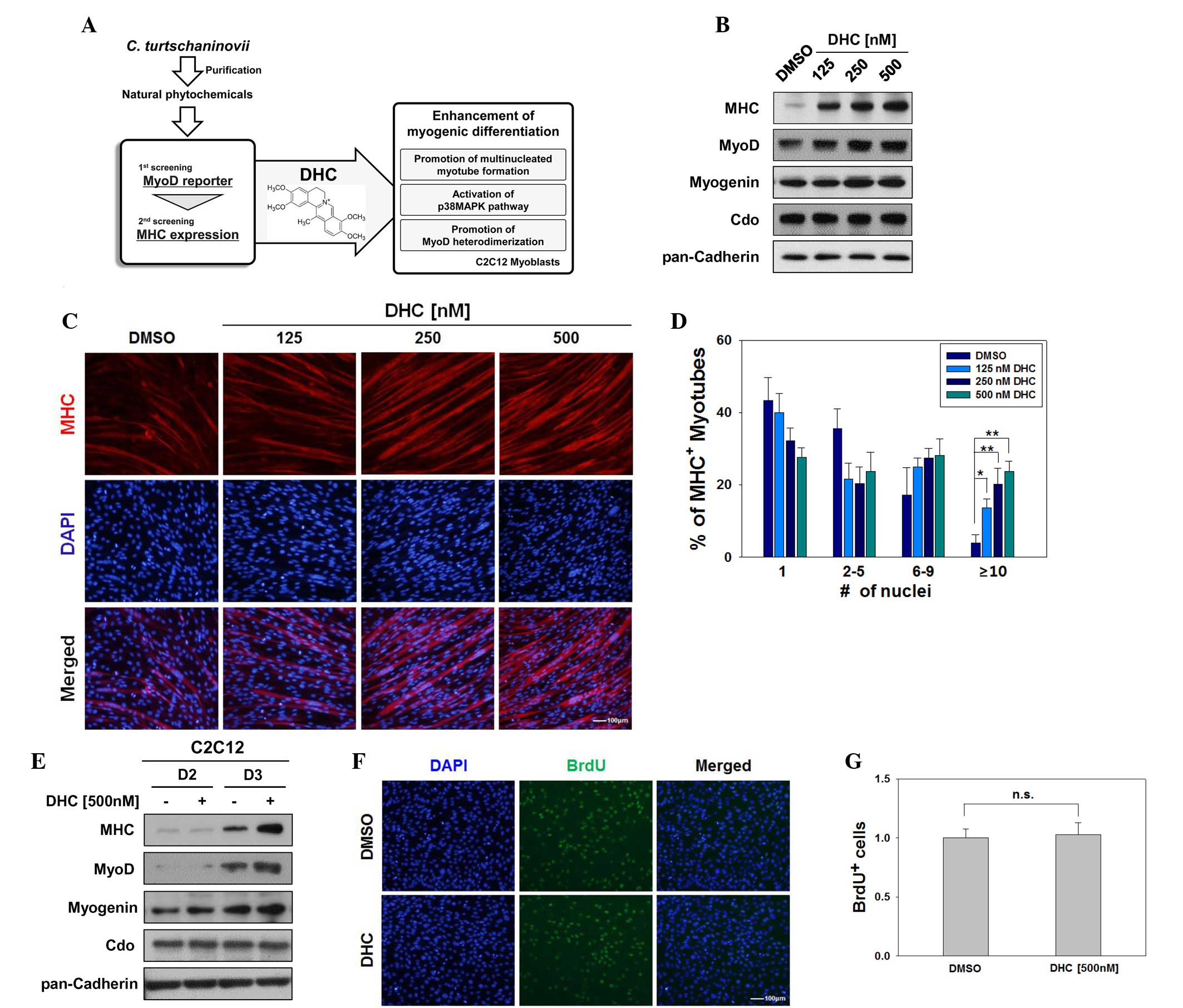 | Figure 1Dehydrocorydaline (DHC) enhances
myogenic differentiation. (A) Schematic diagram indicating the
experimental procedure for screening of natural compounds. (B)
C2C12 cells were treated with the indicated amounts of DHC or
dimethyl sulfoxide (DMSO) and were induced to differentiate for 2
days (D2). Lysates underwent western blotting with antibodies
against myosin heavy chain (MHC), MyoD, myogenin, Cdo, and
pan-Cadherin as a loading control. (C) C2C12 cells were treated
with DMSO or various concentrations of DHC and were induced to
differentiate for 3 days (D3). The cells were immunostained with
MHC antibodies followed by 4′,6-diamidino-2-phenylindole (DAPI)
staining to visualize nuclei. Scale bar, 100 μm. (D)
Quantification of myotube formation in the cells presented in (C).
Data are presented as the mean ± standard deviation (SD) of three
determinations. The experiment was repeated three times with
similar results. *P<0.05 and **P<0.01
vs. the DMSO-treated group. (E) C2C12 cells were treated with 500
nM DHC for D2 and D3. Lysates were immunoblotted with antibodies
against MHC, MyoD, myogenin, Cdo, and pan-Cadherin as a loading
control. (F) C2C12 cells were treated with DMSO or DHC for 1 day
and were labeled with bromodeoxyuridine (BrdU) for 30 min followed
by immunostaining with anti-BrdU and DAPI staining to visualize
nuclei. Scale bar, 100 μm. (G) Quantification of
BrdU-positive cells presented in (F). Data are presented as the
mean ± SD of three determinations. The experiment was repeated
three times with similar results. n.s., not significant. |
The present study initially aimed to confirm whether
DHC was able to enhance myoblast differentiation. C2C12 cells were
induced to differentiate for 2 days in the presence of varying
concentrations of DHC (125–500 nM). DHC-treated C2C12 cells
exhibited increased expression of muscle-specific proteins
including MHC, MyoD and myogenin in a dose-dependent manner
compared with in the control DMSO-treated cells (Fig. 1B). However, the expression levels
of the promyogenic receptor Cdo were not markedly altered. To
determine whether treatment with DHC enhanced the formation of
myotubes, DMSO- and DHC-treated C2C12 cells were induced to
differentiate for 3 days, and were fixed and immunostained with
anti-MHC followed by DAPI staining. As shown in Fig. 1C, DHC-treated C2C12 cells formed
larger myotubes with more nuclei per myotube compared with the
control cells. The MHC-positive cells were scored as follows:
Mononucleated, containing 2–5 nuclei, containing 6–9 nuclei, or
containing ≥10 nuclei (Fig. 1D).
In concordance with Fig. 1C, C2C12
cells treated with DHC exhibited a significant increase in myotubes
with ≥10 nuclei, relative to the vehicle-treated cells (Fig. 1D). Consistently, treatment with DHC
increased the protein expression levels of MHC, MyoD and myogenin
(Fig. 1E). In addition, DHC
enhanced the activity of ectopically expressed MyoD in non-muscle
cells, including 10T1/2 fibroblasts and 293T cells (data not
shown). To determine whether DHC affects myoblast proliferation, a
BrdU incorporation assay was conducted to assess proliferation. As
shown in Fig. 1F and G, treatment
with DHC had no obvious effect on the proliferative capacities of
C2C12 myoblasts compared with the control-treated cells. These data
suggest that DHC treatment promotes myoblast differentiation at the
morphological and biochemical level.
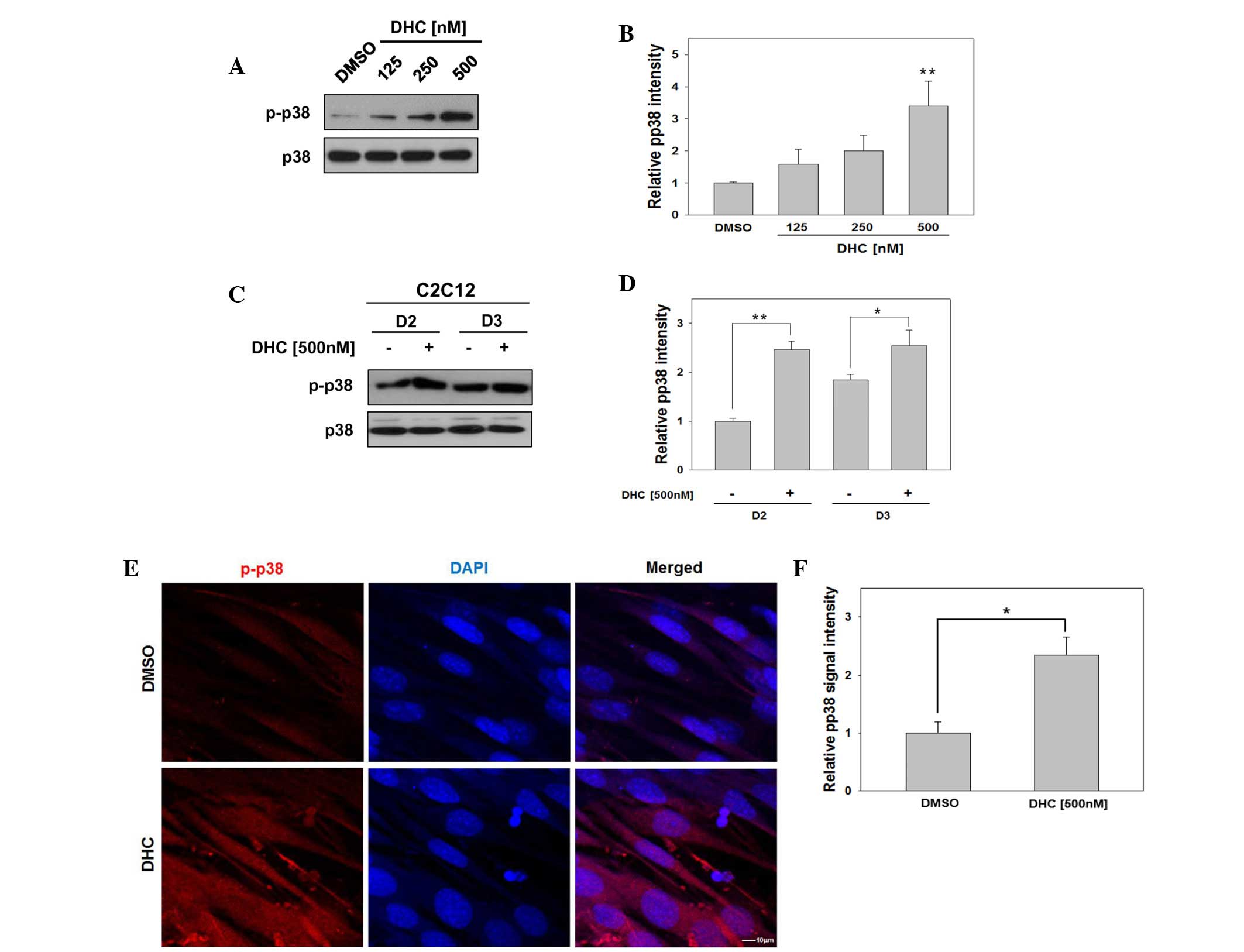
DHC activates p38 MAPK in myoblast
differentiation
Several signaling pathways activate p38 MAPK,
which in turn induces transcriptional activation of MyoD and
promotes myoblast differentiation (14,15,17,18).
To investigate whether DHC activates MyoD via p38 MAPK, C2C12 cells
were induced to differentiate in the presence of DMSO or three
concentrations of DHC, and p38 MAPK activation was analyzed.
Treatment of C2C12 cells with DHC upregulated the expression levels
of active p-p38 in a dose-dependent manner (Fig. 2A and B). In addition, DHC-treated
C2C12 cells exhibited strongly elevated levels of p-p38 at day 2,
compared with the vehicle-treated cells (Fig. 2C and D).
The present study aimed to confirm whether DHC
treatment enhanced phosphorylation of p38 by immunostaining and
confocal microscopy. C2C12 cells were induced to differentiate for
1 day, fixed and immunostained with antibodies against p-p38,
followed by DAPI staining to visualize nuclei. As presented in
Fig. 2E, DHC-treated C2C12 cells
displayed an increase in cells positive for p-p38 signals compared
with the control cells. Subsequently, p-p38 signal intensity was
quantified relative to that of DMSO-treated cells, which was set to
1.0. DHC-treated C2C12 cells displayed a 2.34-fold increase in
p-p38 signal intensity (Fig. 2F).
Previous studies have demonstrated that p38 MAPK is a key regulator
for MyoD activation to promote myoblast differentiation (19). These results indicate that DHC
promotes myoblast differentiation via activation of p38 MAPK and
MyoD.
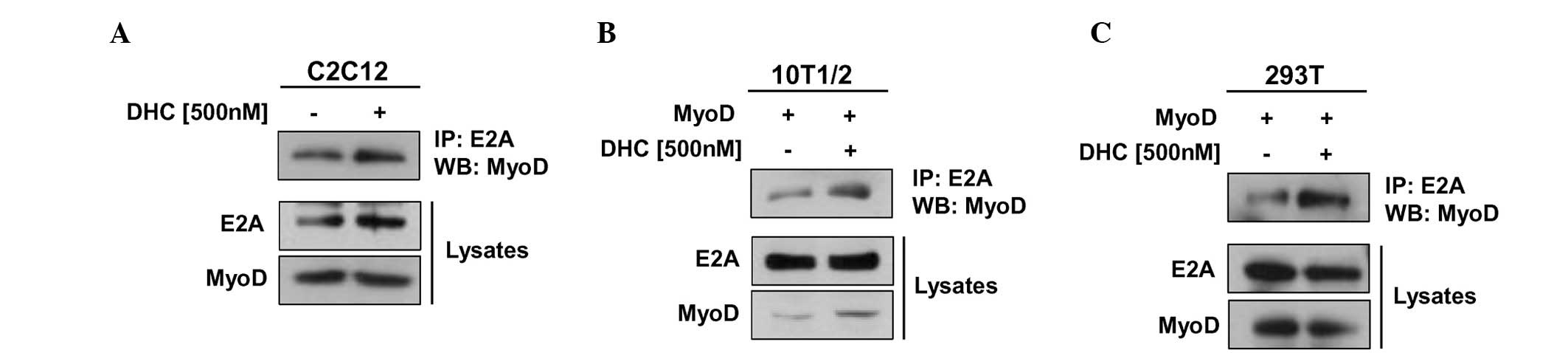
DHC enhances the heterodimerization of
MyoD with E proteins
The transcriptional activity of MyoD is regulated
through several regulatory pathways, including heterodimerization
of MyoD with E proteins, which is stimulated by p38 MAPK-mediated
phosphorylation of E proteins (4,20).
Since DHC treatment activated p38 MAPK and MyoD, the present study
investigated whether DHC regulates MyoD activity through enhancing
interactions between MyoD and an E protein (E2A) in myoblast
differentiation. C2C12 cells were treated with DHC, differentiated,
and were harvested at D2. Cell lysates were then subjected to
immunoprecipitation with an E2A antibody followed by immunoblotting
with MyoD and E2A antibodies. DHC-treated C2C12 cells exhibited
markedly increased precipitation of MyoD proteins with E2A compared
with the control cells (Fig. 3A).
In addition, total E2A levels appeared to be slightly increased by
DHC treatment. Control immunoprecipitation with immunoglobulin G
did not precipitate MyoD (data not shown). These results were
further confirmed using 10T1/2 embryonic fibroblasts transiently
transfected with MyoD expression vectors. Cells were treated with
DMSO or DHC and incubated for 24 h, followed by immunoprecipitation
analysis. As shown in Fig. 3B, DHC
treatment enhanced the interaction of MyoD with E2A in 10T1/2 mouse
embryonic fibroblasts. In the lysates, the protein levels of E2A
remained constant, whereas the protein levels of MyoD were
increased in DHC-treated cells. In agreement with these results,
DHC treatment also increased the binding of MyoD to E2A proteins in
MyoD-transfected 293T cells (Fig.
3C). Activation of MyoD in paired box
(Pax)3/Pax7/MyoD-expressing myoblasts has been shown to
sequentially induce alterations in muscle-specific gene expression,
thus leading to terminal differentiation of myoblasts into
multinucleated myotubes and myofibers (4). MyoD binds to the MyoD-responsive
element E-box in the promoter regions of downstream muscle target
genes, thereby inducing the transcription of these muscle-related
genes in collaboration with myocyte enhancer factor-2 (MEF2)
(21). In the transcriptional
regulation of MyoD, p38 MAPK has an important role in
heterodimerization of MyoD with E proteins (7). Notably, it has been reported that
tetrahydropalmatine, which is an alkaloid compound purified from
medicinal herbs, promotes transdifferentiation of 10T1/2
fibroblasts into myoblasts mediated by MyoD (11). Therefore, natural compounds,
including DHC, may elevate the transcriptional activity of MyoD in
non-muscle cells, and this property may be beneficial in
transdifferentiation of non-muscle cells into myoblasts. These data
suggest that DHC upregulates the interaction of MyoD with E2A via
activation of p38 MAPK.
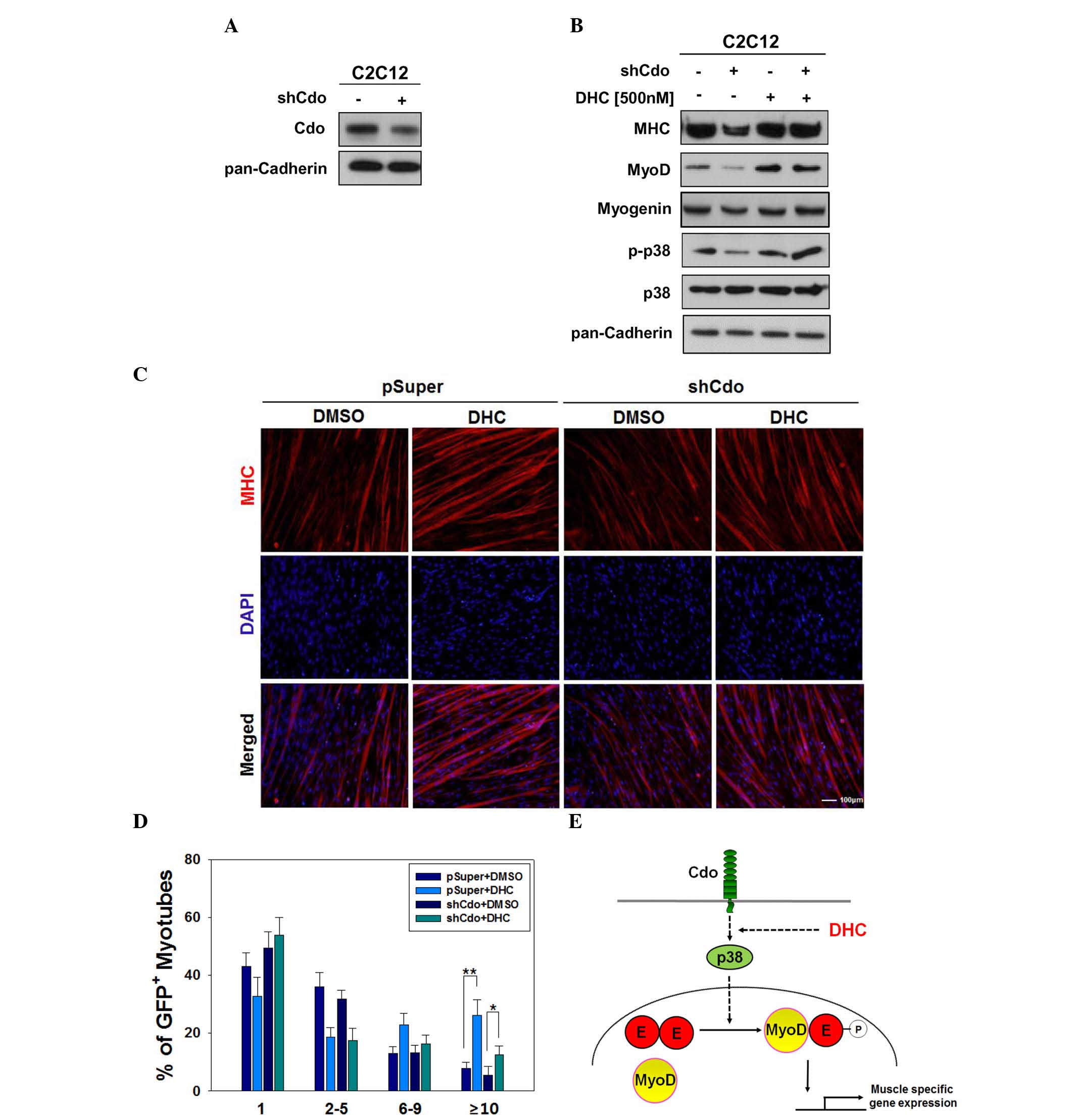
DHC rescues myogenic differentiation in
Cdo-depleted C2C12 cells
The promyogenic receptor Cdo has been reported to
act as a critical component that integrates cell contact-mediated
signals from the cell surface into the myogenic regulatory network
via p38 MAPK and MyoD activation (8). In a previous study, Cdo depletion or
deficiency in myoblasts resulted in reduced p38 MAPK and MyoD
activation, which in turn induced defective myoblast
differentiation (15). The results
of the present study suggested that DHC may increase myoblast
differentiation via activation of p38 MAPK. Therefore, experiments
were conducted to determine whether defective myoblast
differentiation induced by Cdo depletion can be rescued by DHC
treatment. C2C12 cells were transfected with pSuper and Cdo shRNA
expression vectors. C2C12/pSuper and C2C12/Cdo shRNA cells were
subsequently treated with DMSO or DHC, and were induced to
differentiate for 2 days, followed by western blot analysis. In
agreement with our previous study (11), Cdo-depleted cells (Fig. 4A) exhibited decreased levels of
MHC, MyoD and p-p38 compared with in the C2C12/pSuper control cells
(Fig. 4B). Treatment of C2C12/Cdo
shRNA cells with DHC restored the expression of muscle-specific
proteins to similar levels observed in DHC-treated C2C12/pSuper
cells (Fig. 4B). In addition,
DHC-treated C2C12/Cdo shRNA cells exhibited slightly higher levels
of p-p38 compared with in the DHC-treated C2C12/pSuper cells,
whereas total p38 protein expression remained constant.
The present study also investigated whether
defective myotube formation in Cdo-depleted cells can be improved
by DHC treatment. C2C12/pSuper and C2C12/Cdo shRNA cells were
treated with DMSO or DHC, and were induced to differentiate for 3
days, followed by immunostaining analysis for myotube formation.
C2C12/pSuper cells treated with DHC exhibited enhanced myotube
formation, and myotubes contained ≥10 nuclei, as compared with the
control-treated C2C12/pSuper cells. Vehicle-treated C2C12/Cdo shRNA
cells exhibited reduced myotube formation, which was markedly
improved following DHC treatment, almost to control cell levels
(Fig. 4C and D). Cdo is a
promyogenic receptor protein, which regulates myoblast
differentiation predominantly via p38 MAPK activation (18). During myoblast differentiation, Cdo
forms multiprotein complexes to activate the p38 MAPK pathway via
direct binding of the Cdo cytoplasmic tail to scaffold proteins
c-Jun NH2-terminal kinase-associated leucine zipper protein and
BCL2/adenovirus E1B 19 kDa-interacting protein 2 (17,18).
p38 MAPK subsequently regulates the activation of key muscle
transcription factors, including MyoD and MEF2C (22). The results of the present study
demonstrated that DHC ameliorated Cdo depletion-induced defective
differentiation via p38 MAPK activation, which is consistent with
the results of our previous study (18). Our previous study demonstrated that
reactivation of p38 MAPK via constitutively activated MKK6
(MKK6EE), which is an upstream kinase of p38 MAPK, restored the
differentiation capacity of Cdo-deficient myoblasts.
In conclusion, DHC isolated from Corydalis
tuber significantly promoted myoblast differentiation via the
activation of p38 MAPK and MyoD. DHC enhanced the expression of
muscle-specific proteins, including MHC, MyoD and myogenin.
Activation of the p38 MAPK pathway by DHC increased the
heterodimerization of MyoD and E proteins, resulting in MyoD
activation (Fig. 4E). These
findings provide evidence for the promyogenic effects of DHC, and a
mechanistic rationale for the potential pharmaceutical application
of DHC in improvement of muscle regeneration to treat muscle
atrophy.
Acknowledgments
The present study was supported by the Basic Science
Research Program through the National Research Foundation of Korea
(NRF) funded by the Ministry of Education, Science and Technology
(grant no. 2013R1A1A2010280), an NRF grant funded by the Ministry
of Science, ICT and Future Planning (grant no.
2015R1A2A1A15056117), and an NRF grant funded by the Korea
Government (MSIP) (grant no. NRF-2011-0030074).
References
|
1
|
Tedesco FS, Dellavalle A, Diaz-Manera J,
Messina G and Cossu G: Repairing skeletal muscle: Regenerative
potential of skeletal muscle stem cells. J Clin Invest. 120:11–19.
2010. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mangner N, Adams V, Sandri M, Hoellriegel
R, Hambrecht R, Schuler G and Gielen S: Muscle function and running
activity in mouse models of hereditary muscle dystrophy: Impact of
double knockout for dystrophin and the transcription factor MyoD.
Muscle Nerve. 45:544–551. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Horsley V and Pavlath GK: Forming a
multinucleated cell: Molecules that regulate myoblast fusion. Cells
Tissues Organs. 176:67–78. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Berkes CA and Tapscott SJ: MyoD and the
transcriptional control of myogenesis. Semin Cell Dev Biol.
16:585–595. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lluís F, Perdiguero E, Nebreda AR and
Muñoz-Cánoves P: Regulation of skeletal muscle gene expression by
p38 MAP kinases. Trends Cell Biol. 16:36–44. 2006. View Article : Google Scholar
|
|
6
|
Davis RL, Cheng PF, Lassar AB and
Weintraub H: The MyoD DNA binding domain contains a recognition
code for muscle-specific gene activation. Cell. 60:733–746. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lluís F, Ballestar E, Suelves M, Esteller
M and Muñoz-Cánoves P: E47 phosphorylation by p38 MAPK promotes
MyoD/E47 association and muscle-specific gene transcription. EMBO
J. 24:974–984. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Krauss RS, Cole F, Gaio U, Takaesu G,
Zhang W and Kang JS: Close encounters: Regulation of vertebrate
skeletal myogenesis by cell-cell contact. J Cell Sci.
118:2355–2362. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kubo M, Matsuda H, Tokuoka K, Ma S and
Shiomoto H: Anti-inflammatory activities of methanolic extract and
alkaloidal components from Corydalis tuber. Biol Pharm Bull.
17:262–265. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yun KJ, Shin JS, Choi JH, Back NI, Chung
HG and Lee KT: Quaternary alkaloid, pseudocoptisine isolated from
tubers of Corydalis turtschaninovi inhibits LPS-induced nitric
oxide, PGE(2) and pro-inflammatory cytokines production via the
down-regulation of NF-kappaB in RAW 264.7 murine macrophage cells.
Int Immunopharmacol. 9:1323–1331. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lee SJ, Yoo M, Go GY, Hwang J, Lee HG, Kim
YK, Seo DW, Baek NI, Ryu JH, Kang JS and Bae GU:
Tetrahydropalmatine promotes myoblast differentiation through
activation of p38MAPK and MyoD. Biochem Biophys Res Commun.
455:147–152. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ishiguro K, Ando T, Maeda O, Watanabe O
and Goto H: Dehydrocorydaline inhibits elevated mitochondrial
membrane potential in lipopolysaccharide-stimulated macrophages.
Int Immunopharmacol. 11:1362–1367. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Xu Z, Chen X, Fu S, Bao J, Dang Y, Huang
M, Chen L and Wang Y: Dehydrocorydaline inhibits breast cancer
cells proliferation by inducing apoptosis in MCF-7 cells. Am J Chin
Med. 40:177–185. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bae GU, Kim BG, Lee HJ, Oh JE, Lee SJ,
Zhang W, Krauss RS and Kang JS: Cdo binds Abl to promote
p38alpha/beta mitogen-activated protein kinase activity and
myogenic differentiation. Mol Cell Biol. 29:4130–4143. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tran P, Ho SM, Kim BG, Vuong TA, Leem YE,
Bae GU and Kang JS: TGF-β-activated kinase 1 (TAK1) and apoptosis
signal-regulating kinase 1 (ASK1) interact with the promyogenic
receptor Cdo to promote myogenic differentiation via activation of
p38MAPK pathway. J Biol Chem. 287:11602–11615. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Slavík J and Slavíková L: Alkaloids from
Corydalis cava (L). SCHW. et KOERTE. Collect Czech Chem Commun.
44:2261–2274. 1979. View Article : Google Scholar
|
|
17
|
Kang JS, Bae GU, Yi MJ, Yang YJ, Oh JE,
Takaesu G, Zhou YT, Low BC and Krauss RS: A Cdo-Bnip-2-Cdc42
signaling pathway regulates p38alpha/beta MAPK activity and
myogenic differentiation. J Cell Biol. 182:497–507. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Takaesu G, Kang JS, Bae GU, Yi MJ, Lee CM,
Reddy EP and Krauss RS: Activation of p38alpha/beta MAPK in
myogenesis via binding of the scaffold protein JLP to the cell
surface protein Cdo. J Cell Biol. 175:383–388. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Keren A, Tamir Y and Bengal E: The p38
MAPK signaling pathway: A major regulator of skeletal muscle
development. Mol Cell Endocrinol. 252:224–230. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Davis RL, Weintraub H and Lassar AB:
Expression of a single transfected cDNA converts fibroblasts to
myoblasts. Cell. 51:987–1000. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Puri PL and Sartorelli V: Regulation of
muscle regulatory factors by DNA-binding, interacting proteins, and
post-transcriptional modifications. J Cell Physiol. 185:155–173.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zetser A, Gredinger E and Bengal E: p38
mitogen-activated protein kinase pathway promotes skeletal muscle
differentiation. Participation of the Mef2c transcription factor. J
Biol Chem. 274:5193–5200. 1999. View Article : Google Scholar : PubMed/NCBI
|