Introduction
Hyperlipidemia is a lipid metabolism disorder, the
prevalence of which has markedly increased in recent years. This
disorder, which is caused by excessive consumption of food
containing high levels of fat and cholesterol, is closely
associated with hypertension, atherosclerosis (AS) and
cardiovascular diseases (CVD) (1–3). It
has previously been established that circulating low-density
lipoprotein cholesterol (LDL-c), total cholesterol (TC) and
triglycerides (TGs) are important risk factors in hypertension, AS
and CVD (4–6). Increased levels of LDL-c, TC and TG
in the blood weaken vessel walls and subsequently block blood flow,
which may lead to myocardial infarction and stroke.
Hypercholesterolemia may also be coupled with hepatic lipid
accumulation. Increased lipid content in the liver induces chronic
inflammation, which accelerates liver injury and may result in
cirrhosis, liver failure and cancer. Accordingly, downregulation of
increased LDL-c, TC and TG is required to prevent and treat these
vascular and hepatic diseases.
It is well established that amelioration of lipid
concentration in the blood prevents hypercholesterolemia and
hepatic lipid accumulation. Lipid metabolism in the liver is
controlled by fatty acid-synthesizing and energy expenditure
enzymes, with decreased energy expenditure enzymes and increased
fatty acid-synthesizing enzymes generally being observed in the
livers of obese animal models fed a high-fat diet (HFD) and/or a
high-cholesterol diet (HCD) (2,7–9).
Furthermore, the hepatic expression levels of LDL receptor (LDLR)
are associated with the concentration of circulating serum
cholesterol in experimental animals fed a HFD and/or HCD (10). Previous studies have demonstrated
that numerous candidates are able to decrease blood LDL-c, TC and
TG levels via the suppression of lipogenic factors and the
induction of lipolytic factors in obese animals, and several
candidates improved lipid components in the blood via regulation of
LDLR expression in hyperlipidemic rodents (2,7,9,11).
These results indicated that regulation of lipid metabolism enzymes
and LDLR are useful strategies for preventing liver fat
accumulation and hypercholesterolemia.
Deep sea water (DSW) is considered a potent material
that has food and medical applications. DSW contains abundant
minerals, including magnesium (Mg), calcium (Ca), potassium (K) and
zinc, which have important roles in cellular homeostasis and
physiological responses (12–24).
The beneficial effects of DSW on vascular diseases and metabolic
disorders have been well demonstrated and these effects are thought
to be associated with lipid metabolism (19,20).
Hwang et al (19)
demonstrated that DSW decreased body weight and improved lipid
components in ob/ob mice; in addition, the differentiation
of 3T3-L1 adipocytes was prevented by DSW (19,20).
Although the beneficial effects of DSW in lipid metabolism have
previously been investigated in several laboratories, the
preventative effects of DSW on liver fat accumulation and
hypercholesterolemia have not been fully investigated. Therefore,
the present study aimed to determine the effects of DSW on liver
fat accumulation and hypercholesterolemia in rats fed a HCD.
Materials and methods
Preparation of DSW
DSW was obtained from the Marine Deep Ocean Water
Application Research Center in the Korea Institute of Ocean Science
& Technology (Ansan, South Korea) from a depth of 500 m in the
East Sea (Goseong, South Korea). Saline and minerals in DSW were
removed and extracted by reverse osmosis filtration and
electrodialysis (16). Extracted
minerals were dissolved in desalinated DSW to generate hardness
4,000 (H4000) DSW containing 835.6 mg/l Mg, 279.9 mg/l Ca, 213.7
mg/l Na and 81.2 mg/l K (Mg:Ca concentration ratio, 3:1). H4000 DSW
was serially diluted with desalinated DSW to prepare DSW of various
hardness (400–2,000). The hardness of DSW was determined by the
following formula: Total hardness=Ca hardness [2.5 × Ca
concentration (mg/l)] + Mg hardness [4.1 × Mg concentration
(mg/l)].
Animals and treatment
Animal experiments were conducted following approval
by the Animal Use and Care Committee at Dongguk University
(approval IACUC-2013-001; Gyeongju, Korea). A total of 42 male
5-week old Sprague-Dawley rats (120–130 g) with a normal diet (ND;
5L57, containing no cholesterol) were obtained from Orient Bio Inc.
(Seongnam, Korea). The rats were housed under a 12 h light/dark
cycle at 25±2°C and a relative humidity of 50±5%. The rats received
the ND and tap water ad libitum for 1 week. Subsequently, rats
received a HCD (D12336, Research Diets, Inc., New Brunswick, NJ,
USA) with tap water or DSW of various hardness ad libitum for 6
weeks. The composition of the HCD is presented in Table I. Rats were randomly divided into 1
ND group and 6 experimental groups: Tap HCD, H0 HCD, H400 HCD, H800
HCD, H1500 HCD and H2000 HCD. Each group consisted of 6 rats. Body
weight, and food and water intake were measured every 2–3 days
during the experiment. After 6 weeks, animals were fasted for 24 h
and subsequently sacrificed with ether by inhalation, then blood
was collected to determine the lipid composition in each group.
 | Table I.Composition of high-cholesterol
diet. |
Table I.
Composition of high-cholesterol
diet.
| Ingredient | Amount (g/kg) |
|---|
| Casein | 75 |
| Soy protein | 130 |
| DL-methionine | 2 |
| Corn starch | 275 |
| Maltodextrin
10 | 150 |
| Sucrose | 30 |
| Cellulose | 90 |
| Soy bean | 50 |
| Cocoa butter | 75 |
| Coconut oil | 35 |
| Mineral mix | 35 |
| Calcium
carbonate | 5.5 |
| Sodium
chloride | 8 |
| Potassium
citrate | 10 |
| Vitamin mix
V10001 | 10 |
| Choline
bitartrate | 2 |
| Cholesterol | 12.5 |
| Sodium cholic
acid | 5 |
Analysis of blood lipid
components
TG, TC and high-density lipoprotein cholesterol
(HDL-c) in the blood were enzymatically analyzed using commercial
kits (AM157K, AM202K and AM203K respectively; Asan Pharmaceutical
Co., Ltd., Seoul, Korea) according to the manufacturer's protocols.
The LDL-c concentration was calculated using the Friedwald formula:
LDL-c concentration=TC concentration-HDL-c concentration-TG/2.
Evaluation of liver damage
indicators
Glutamate-oxaloacetate transaminase (GOT),
glutamate-pyruvate transferase (GPT) and alkaline phosphatase (ALP)
activity in the blood were assessed as indicators of liver damage.
GOT and GPT activities were measured using a commercial kit
(AM101K; Asan Pharmaceutical Co., Ltd.) based on the
Reitman-Frankel method (25),
whereas ALP activity was determined using a Kind-King method-based
commercial kit (AM105S; Asan Pharmaceutical Co., Ltd.) according to
the manufacturer's protocols.
Electron microscopic analysis
Livers were pre-fixed with 0.1 M PBS containing 2.5%
glutaraldehyde for 2 h at 4°C and subsequently washed with 0.1 M
PBS three times for 15 min. The tissues were subsequently
post-fixed by immersion in 2% osmium tetroxide solution for 2 h at
4°C followed by dehydration with ethanol. Tissues were embedded
with epon-812 resin, sectioned at 100 nm thickness using a Leica
Ultracut R (Leica Microsystems GmbH, Wetzlar, Germany) and
double-stained with uranyl acetate and lead nitrate. Finally,
tissues were visualized using a Hitachi H-7500 transmission
electron microscope (Hitachi, Ltd., Tokyo, Japan) at 80 kV.
Semi-quantitative and quantitative (q)
reverse transcription-polymerase chain reaction (RT-PCR)
The expression of fatty acid synthase (FAS),
carnitine palmitoyltransferase-1 (CPT-1), sterol regulatory element
binding protein-1c (SREBP-1c) and peroxisome proliferator-activated
receptor γ (PPARγ) was analyzed by semi-quantitative RT-PCR, and
qPCR was used to analyze the expression of LDLR. Livers were
rapidly frozen in liquid nitrogen and stored at −80°C. Total RNA in
individual liver samples was extracted using an easy-BLUE™ Total
RNA Extraction kit (17061; Intron Biotechnology, Inc., Seongnam,
Korea) according to the manufacturer's protocol. cDNA synthesis was
performed using PrimeScript™ 1st strand cDNA Synthesis kit (6110a;
Takara Bio, Inc., Otsu, Japan) according to the manufacturer's
protocols and amplification of PCR products for semi-quantitative
RT-PCR was performed with 2 µl of cDNA in Ex Taq DNA polymerase
mixture containing 2 mM MgCl2, 200 µM dNTP (Takara Bio,
Inc.) and 0.2 µM of each forward and reverse primer (Bioneer
Corporation, Daejeon, Korea) with a final reaction volume of 25 µl.
The PCR cycling conditions were as follows: 95°C for 10 min
(initial denaturation), 22–32 cycles at 95°C for 30 sec, 60°C for
30 sec, 72°C for 30 sec (amplification) and 72°C for 10 min (final
extension). All reactions were finished during the exponential
phases. PCR products and 100 bp ladder (WelGene Co., Daegu, Korea)
were subjected to agarose gel electrophoresis containing 0.5 µg/ml
ethidium bromide (Promega Corporation, Madison, WI, USA) and
observed using i-MAX Gel Image Analysis System with CoreBio MFC
software (CoreBio System Co., Ltd., Seoul, Korea). qPCR was
performed using a QGreen™ SYBR Green Master Mix kit (Cellsafe Co.
Ltd., Suwon, Korea) and the Eco Real-Time PCR system (Illumina,
Inc., San Diego, CA, USA). The PCR cycling conditions were as
follows: 95°C for 10 min followed by 45 cycles at 95°C for 10 sec,
60°C for 10 sec and 72°C for 30 sec. The relative intensity of the
target genes was calculated using Eco™ software version 3.1.7
(Illumina, Inc., San Diego, CA, USA) by the ΔΔCq method (26). GAPDH was used as an internal
control to normalize target gene expression. The PCR primer
sequences for target genes are presented in Table II.
| Table II.Sequences of primers used for
semi-quantitative RT-PCR and RT-qPCR. |
Table II.
Sequences of primers used for
semi-quantitative RT-PCR and RT-qPCR.
| A,
Semi-quantitative PCR |
|---|
|
|---|
| Gene | F/R primer | Primer
sequence | Annealing
temperature (°C) | Cycle number |
|---|
| FAS | F |
5′-CTGGACTCGCTCATGGGTG-3′ | 60 | 25 |
|
| R |
5′-CATTTCCTGAAGCTTCCGCAG-3′ |
|
|
| CPT-1 | F |
5′-AACCTTGGCTGCGGTAAGACTA-3′ | 60 | 22 |
|
| R |
5′-AGTGGGACATTCCTCTCTCAGG-3′ |
|
|
| SREBP-1c | F |
5′-GATGCCAACCAGATTCCCTAAG-3′ | 60 | 29 |
|
| R |
5′-TCAGTTGTTTCTTTGCCTTCCA-3′ |
|
|
| PPARγ | F |
5′-TTCAGTTTGGAGACTTCGGACC-3′ | 60 | 32 |
|
| R |
5′-TAGGCTCCTGCCAGATTACTCC-3′ |
|
|
| GAPDH | F |
5′-AACTTTGGCATCGTGGAAGG-3′ | 59 | 22 |
|
| R |
5′-TACATTGGGGGTAGGAACAC-3′ |
|
|
|
| B, RT-qPCR |
|
| Gene | F/R primer | Primer
sequence | Annealing
temperature (°C) | Cycle number |
|
| LDLR | F |
5′-CAGCTCTGTGTGAACCTGGA-3′ | 58 | 45 |
|
| R |
5′-TTCTTCAGGTTGGGGATCAG-3′ |
|
|
| GAPDH | F |
5′-AACTTTGGCATCGTGGAAGG-3′ | 58 | 45 |
|
| R |
5′-TACATTGGGGGTAGGAACAC-3′ |
|
|
Statistical analysis
Values were presented as the mean ± standard
deviation. Statistical analysis was performed using one-way
analysis of variance with SPSS software (version no. 22; SPSS,
Inc., Chicago, IL, USA) followed by Student's t-test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Changes in the lipid composition of
blood in response to DSW treatment
The present study monitored body weight, and food
and water (tap water or DSW) intake, in rats fed a HCD. No
significant differences in body weight (Fig. 1A) or food intake (Fig. 1B) were observed among the groups.
However, reduced total water intake was observed in DSW groups in a
hardness-dependent manner (Fig.
1C). In addition, blood lipid components were measured. Blood
TC and LDL-c in rats fed a HCD were increased ~3.4- and 29.9-fold,
respectively, and HDL-c was decreased ~4.8-fold compared with rats
fed a ND (data not shown). Despite the decreased total water intake
in DSW groups, significantly reduced levels of TC and LDL-c were
observed in the H800 (P<0.05) and H1500 (P<0.01) HCD groups
compared with the Tap HCD group (Fig.
2A and B). In addition, significantly increased HDL-c was
detected in response to DSW in the H800 and H1500 HCD groups
compared with in the Tap HCD group (P<0.05; Fig. 2C). However, no significant
alterations in TG were observed among the groups (Fig. 2D).
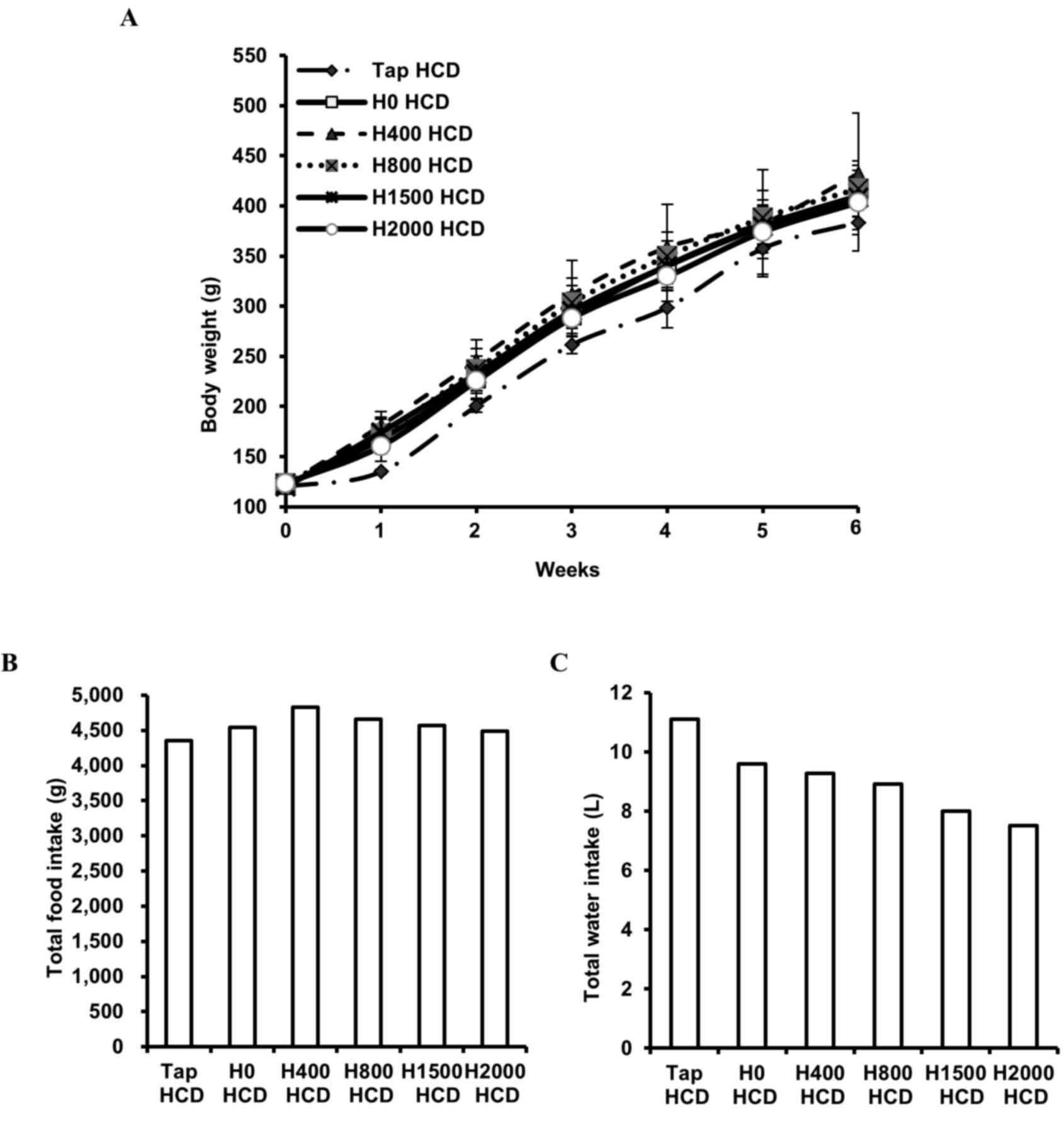 | Figure 1.Effects of DSW on body weight, and
total food and water intake, in rats fed a HCD. (A) Body weight of
each rat was measured every 2–3 days. Values are presented as the
mean ± standard deviation, n=6. (B) Total food intake and (C) total
tap water or DSW intake, was measured every 2–3 days. Values are
presented as the sum of the amount of food, and volume of water,
consumed in each group for 6 weeks. DSW, deep sea water; HCD,
high-cholesterol diet; Tap, tap water; H, hardness. |
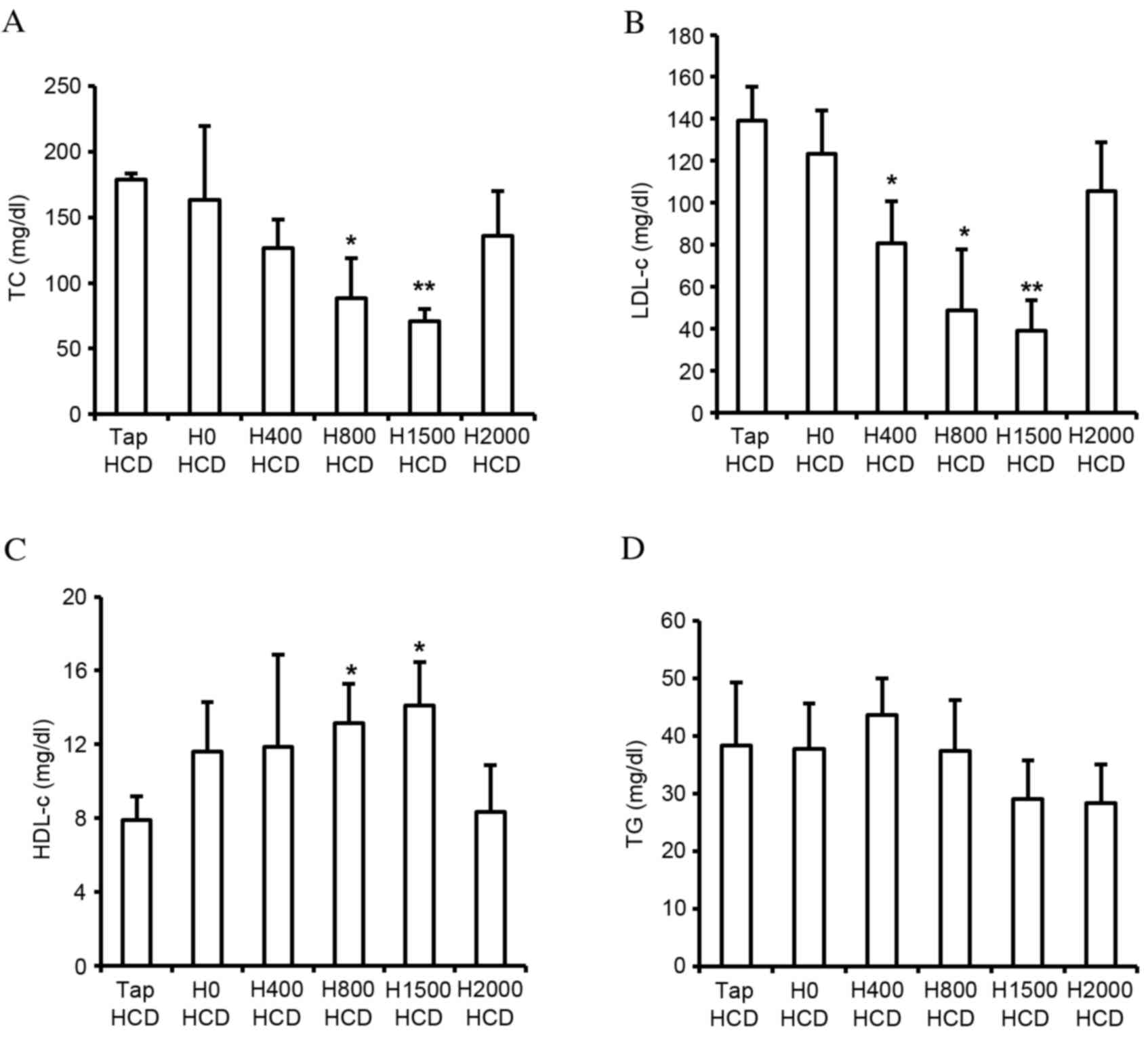 | Figure 2.Effects of DSW on levels of serum
lipid components. Serum (A) TC, (B) LDL-c, (C) HDL-c and (D) TG
concentrations were measured in rats fed a HCD with tap water or
DSW of various hardness for 6 weeks. Values are presented as the
mean ± standard deviation, n=6. *P<0.05 and **P<0.01 vs. the
Tap HCD group. DSW, deep sea water; TC, total cholesterol; LDL-c,
low-density lipoprotein cholesterol; HDL c, high-density
lipoprotein cholesterol; TG, triglyceride; HCD, high-cholesterol
diet; Tap, tap water; H, hardness. |
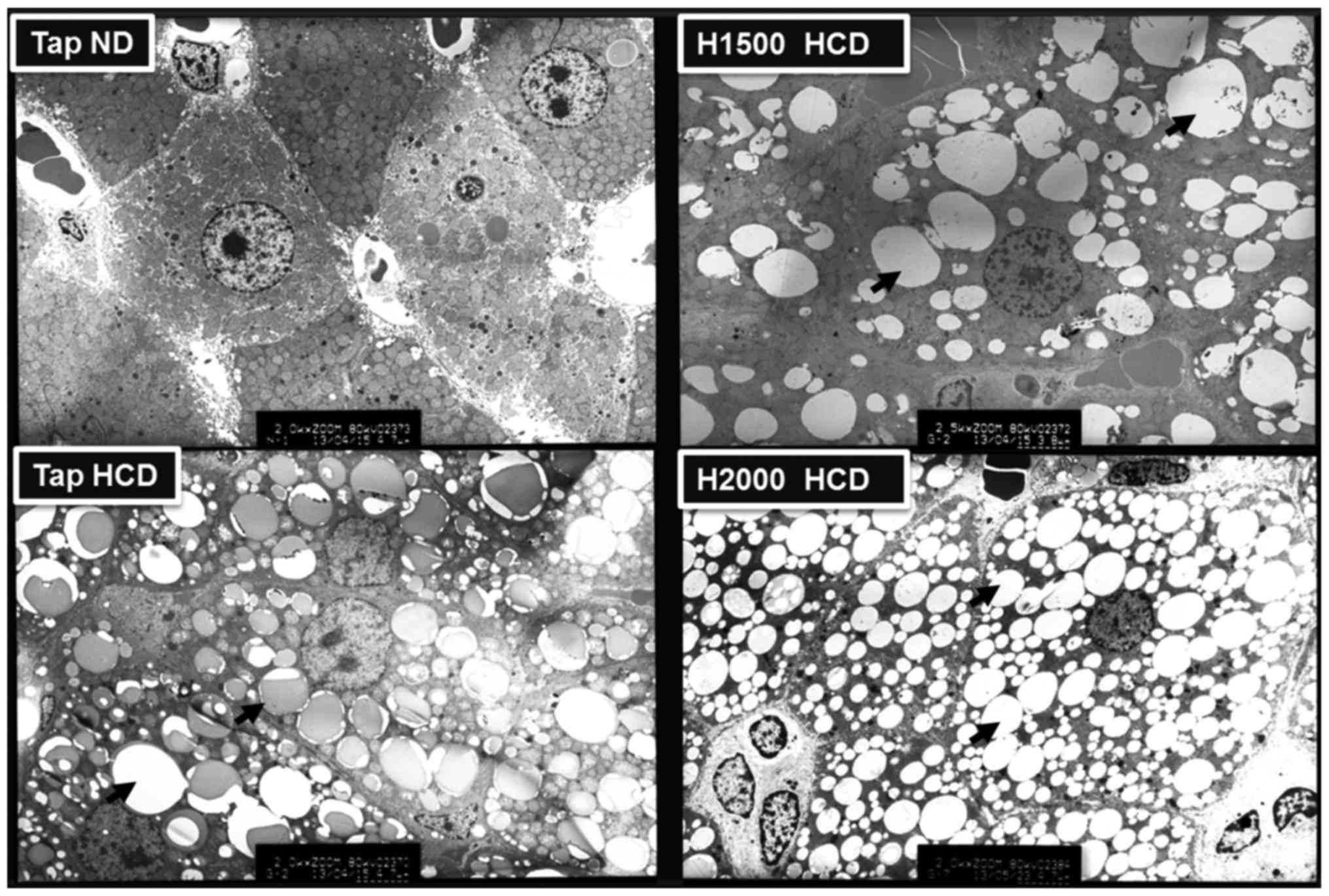
Suppression of hepatic lipid
accumulation
Metabolic diseases, including obesity, diabetes and
hypercholesterolemia, may be induced by a HCD and are associated
with hepatic lipid accumulation (27). Therefore, the present study
analyzed the distribution of lipid droplets in rat liver cells
using electron microscopy. The liver cells of rats fed a HCD
exhibited numerous lipid droplets and the number of lipid droplets
was visibly increased in HCD livers compared with ND-fed rat
livers. However, the H1500 DSW HCD group exhibited fewer liver cell
lipid droplets compared with the Tap HCD group (Fig. 3). Conversely, the H2000 group
exhibited increased numbers of lipid droplets in liver cells
compared with the H1500 DSW HCD group (Fig. 3). The results of electron
microscopy corresponded to the blood TC, LDL-c and HDL-c levels
observed in these groups.
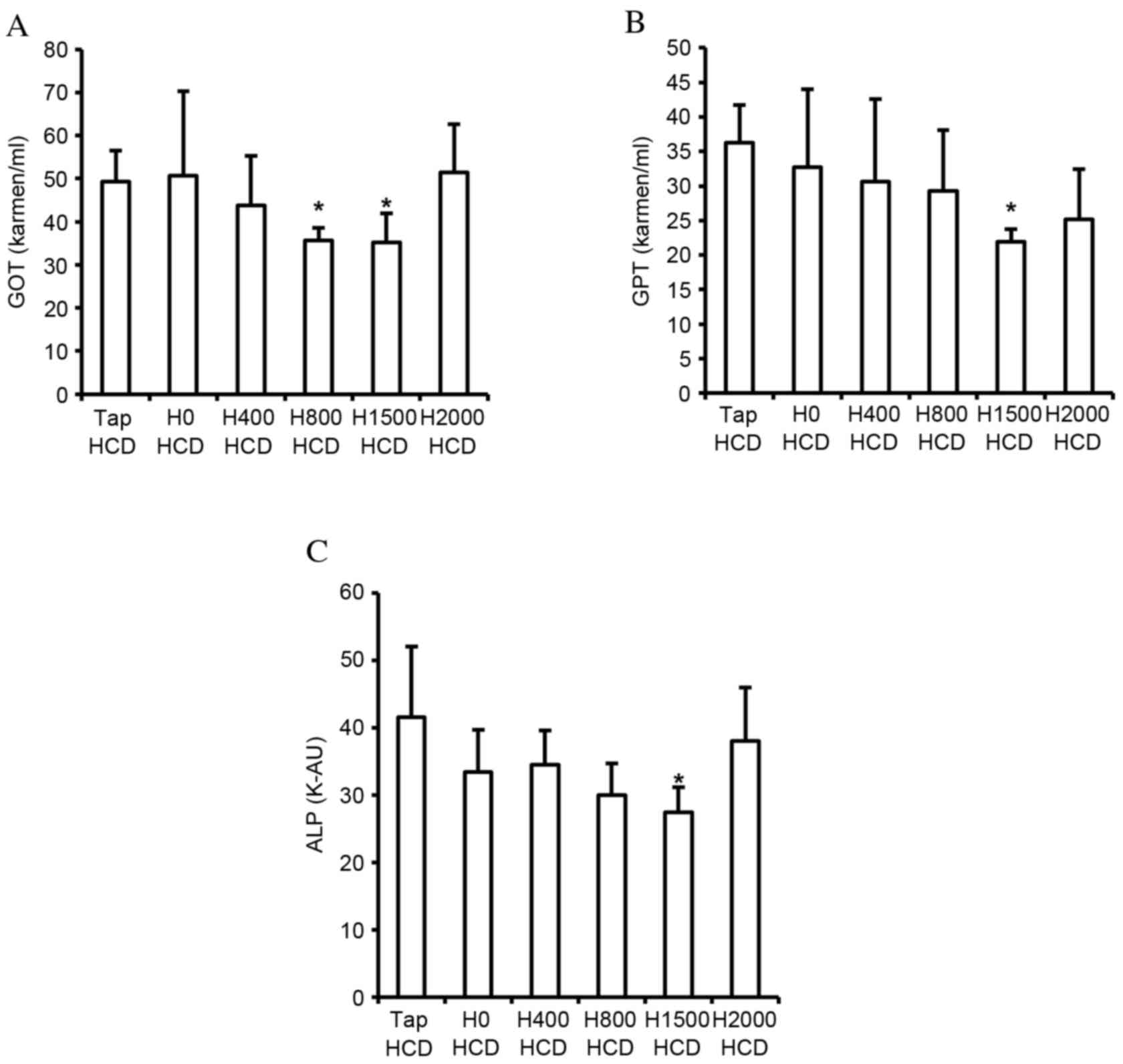
Alleviation of liver injury
indices
Lipid accumulation in the liver, and increased blood
TC and LDL-c concentration, are associated with liver injury. The
present study detected the suppressive effects of DSW on hepatic
lipid accumulation, and the elevation of blood TC and LDL-c
concentration. Therefore, the effects of DSW on liver injury
indices in the blood, including GOT, GPT and ALP, were assessed.
HCD-induced increased GOT, GPT and ALP levels in the blood were
significantly decreased by H1500 DSW compared with the Tap HCD
group (P<0.05; Fig. 4);
however, H2000 DSW did not significantly reduce levels compared
with the Tap HCD group (Fig. 4).
The decrease in GOT, GPT and ALP levels corresponded with the
decrease of hepatic lipid accumulation and blood TC and LDL-c
levels.
Regulation of lipid
metabolism-regulating gene expression in the liver
Lipid homeostasis in the liver is governed by the
balance of expression between fatty acid-synthesizing enzymes and
energy expenditure enzymes. Numerous studies have detected fat
accumulation in the livers of HFD- and/or HCD-fed rodents (7,8,28).
Furthermore, hepatic FAS, PPARγ and SREBP-1c expression in rodent
livers have previously been demonstrated to be significantly
increased by a HFD and/or HCD (8,11,16,28,29).
Therefore, the present study investigated the difference in the
expression of these genes between control and DSW groups in rats
fed a HCD. In addition, the expression of CPT-1, an energy
expenditure enzyme, was assessed. DSW groups exhibited
significantly reduced levels of FAS and SREBP-1c expression in
H800, H1500 and H2000 HCD groups compared with the Tap HCD group
(P<0.05; Fig. 5A and B).
However, no significant differences were observed in CPT-1 and
PPARγ expression (Fig. 5C and
D).
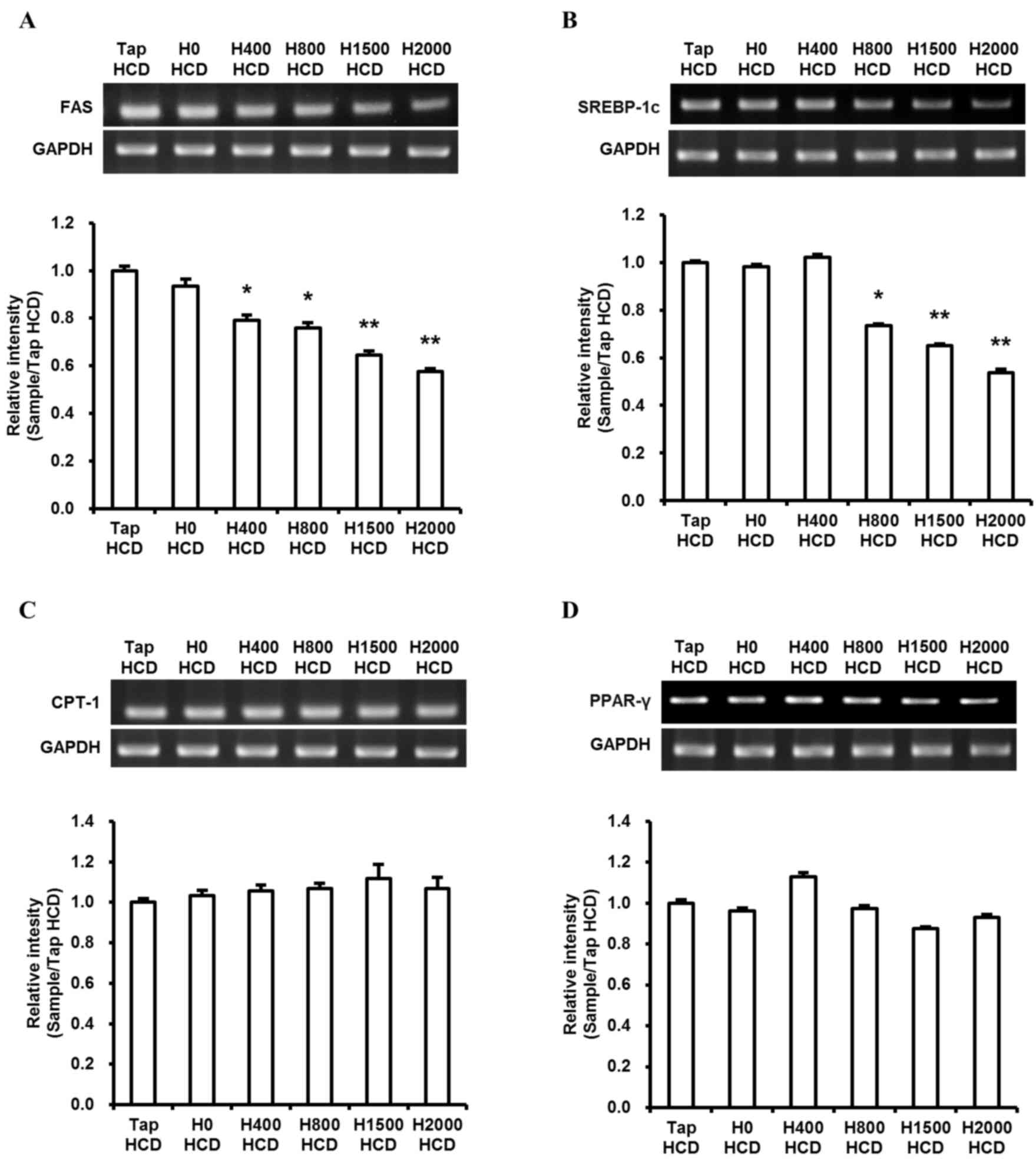 | Figure 5.Effects of DSW on hepatic lipid
metabolism-regulating gene expression. Levels of hepatic lipid
metabolism-regulating genes (A) FAS, (B) SREBP-1c, (C) CPT-1 and
(D) PPARγ were assessed by semi-quantitative RT-PCR and the
densities were normalized to GAPDH, which was used as an internal
control. To perform semi-quantitative RT-PCR, an equal amount of
six individual total RNA samples in each group were pooled. Values
are presented as the mean ± standard deviation, n=3. *P<0.05 and
**P<0.01 vs. the Tap HCD group. DSW, deep sea water; FAS, fatty
acid synthase; SREBP-1c, sterol regulatory element binding
protein-1c; CPT-1, carnitine palmitoyltransferase-1; PPARγ,
peroxisome proliferator-activated receptor γ; RT-PCR, reverse
transcription-polymerase chain reaction; Tap, tap water; HCD,
high-cholesterol diet; H, hardness. |
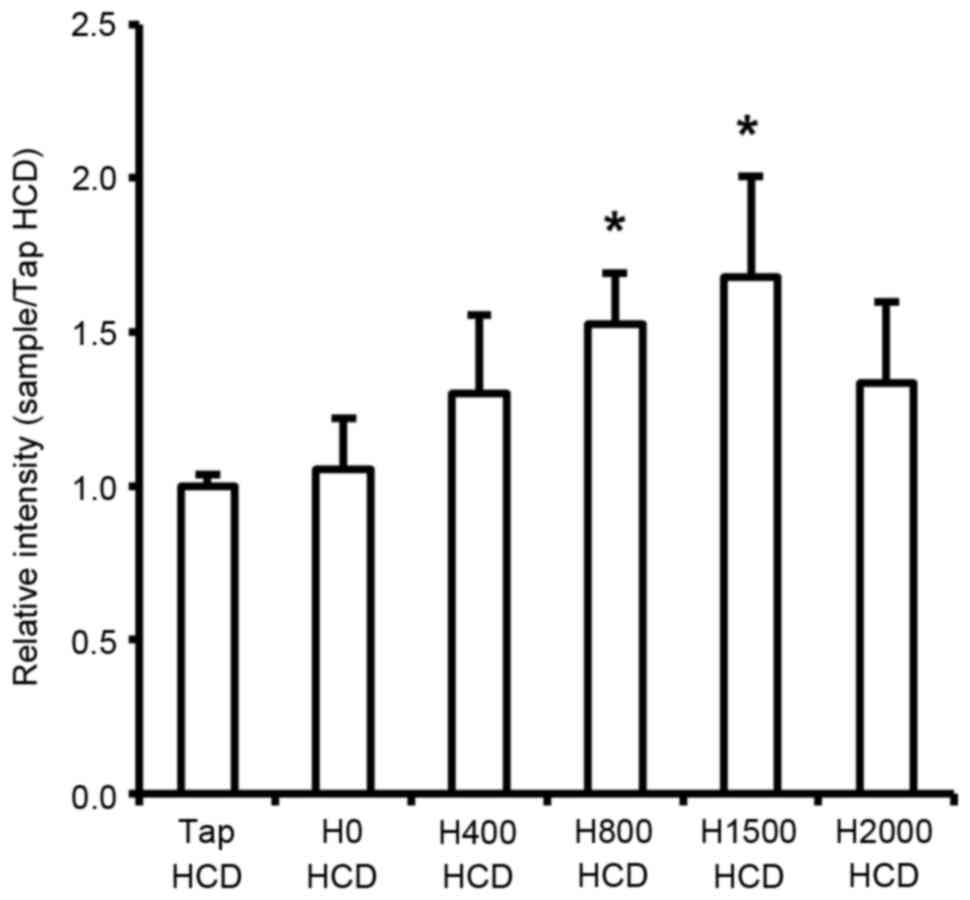
Regulation of hepatic LDLR gene
expression
The present study demonstrated that serum TC and
LDL-c levels were decreased in response to DSW in rats fed a HCD.
Circulating serum cholesterol is primarily absorbed in the liver
through hepatic LDLR-mediated endocytosis and is subsequently
metabolized (30,31). Consequently, serum cholesterol
levels should be associated with hepatic LDLR levels. Therefore,
the present study investigated mRNA expression of LDLR in the liver
of rats. The results revealed a significant increase in hepatic
LDLR mRNA in rats fed a HCD in response to DSW at H800 and H1500
compared with the Tap HCD group (P<0.05; Fig. 6). However, although H2000 DSW also
increased LDLR mRNA expression compared with in the Tap HCD group,
the increase was not statistically significant (Fig. 6).
Discussion
Several studies have demonstrated the importance of
minerals, including Mg and Ca, in lipid metabolism. For example,
increased Mg intake was demonstrated to prevent
hypercholesterolemia, lipid oxidation and oxidative damage
(32,33). Conversely, growth inhibition in
fetal mice was induced by altered lipid metabolism caused by
maternal Mg deficiency and low levels of Mg in blood were observed
in obese children from South India (34,35).
In addition, a high Ca intake was associated with low serum levels
of TC and LDL-c in humans (36).
The present study demonstrated that the blood lipid composition in
rats fed a HCD improved in response to DSW containing high levels
of Mg and Ca (concentration ratio Mg:Ca=3:1; Fig. 2). The results indicated that DSW
may reduce blood TC and LDL-c, and increase HDL-c, through
increased blood Mg and Ca levels. However, the TC, LDL-c and HDL-c
levels in rats treated with H2000 DSW, the highest hardness in this
experiment, were not significantly altered (Fig. 2A-C). These results demonstrated
that increased Mg levels in the blood may have an important role in
the reduction of harmful cholesterol; however, the beneficial
effects of excessive concentrations of Mg and Ca may be lower.
Increased levels of blood lipid components,
including TG, TC and LDL-c, induced by a HFD and/or HCD may lead to
liver fat accumulation. The hepatic accumulation of fat may be
prevented by lowering blood levels of these lipid components.
Previous studies (19,23,29)
have demonstrated that DSW attenuated hepatic lipid accumulation in
hamsters and mice. Furthermore, an increase of Mg and Ca in DSW led
to the alleviation of hepatic lipid accumulation and oxidation in a
concentration-dependent manner in hamsters fed HFDs (29). The results of the present study
demonstrated that H1500 DSW prevented lipid accumulation in the
liver; however, this decrease was not observed in the H2000 DSW
group (Fig. 3). Therefore,
although the association between hepatic lipid accumulation and
mineral (Mg and Ca) content is unclear, the results of the present
study indicated that the beneficial effects of DSW on hepatic lipid
accumulation may be determined by the concentration of Mg and Ca in
DSW.
Increased levels of liver injury indicators are
associated with liver fat accumulation and increased serum TC and
LDL-c. Chen et al (29)
detected decreased GOT and GPT in hamsters fed a HFD/HCD for 6
weeks following treatment with DSW drinking water. In addition, the
previous study demonstrated that the decrease was associated with a
reduction in TC and TG concentration. The results of the present
study are consistent with those of Chen et al (29; Fig. 4). High levels of GOT, GPT and ALP
have been observed in patients with liver diseases, including
hepatitis, cirrhosis, liver failure and liver cancer (37,38).
Therefore, the suppression of increases in GOT, GPT and ALP levels
may be important for the prevention of diet-induced hepatic
diseases.
PPARγ and SREBP-1c are transcriptional regulators of
lipid metabolism enzymes. Previous studies (28,39)
have demonstrated that hepatic PPARγ and SREBP-1c expression were
increased in rodents fed a HFD and/or HCD, and that suppression of
PPARγ and SREBP-1c gene expression reduced fat accumulation and
blood TC and LDL-c levels in livers of mice. Furthermore, decreased
lipid deposits in hepatocytes were observed when SREBP-1c silencing
was performed in vitro (40). In the present study, DSW suppressed
liver fat accumulation and reduced the HCD-induced increases in TC
and LDL-c levels in the blood and FAS and SREBP-1c transcription;
however, no effects on CPT-1 and PPARγ expression were observed
(Fig. 5). Although Chen et
al (29) demonstrated that
serum lipid component levels were improved in response to DSW
drinking water, no effects were observed on FAS and SREBP-1c
expression in response to DSW (29). The ratio of Mg:Ca in DSW drinking
water in Chen et al (29)
was 4-5:1; however, DSW in the present investigation was 3:1.
Therefore, the dissimilarity in the effects of DSW on FAS and
SREBP-1c expression may be caused by differences in the ratio of
Mg:Ca. The results of the present study indicated that DSW may
prevent lipid accumulation in the liver via suppression of FAS
expression regulated by SREBP-1c, without the induction of CPT-1
transcription, and may be more effective at preventing liver fat
accumulation and increases in TC and LDL-c levels.
Previous studies (31,41–43)
have demonstrated an association between decreasing serum
cholesterol and increasing LDLR expression in the liver in response
to various materials. The present study demonstrated that H800 and
H1500 DSW decreased serum LDL-c concentrations, and that this
decrease was accompanied by the induction of LDLR expression in
rats fed a HCD (Figs. 2B and
6). Therefore, the present study
indicated that the hypocholesterolemic effects of DSW may be
mediated by LDLR. However, although decreases in the expression
levels of FAS and SREBP-1c were observed (Fig. 5), H2000 DSW did not prevent liver
fat accumulation or improve serum lipid component levels (Figs. 2 and 3). In addition, LDLR expression was not
significantly increased by H2000 DSW compared with in the Tap HCD
group (Fig. 6). Although it is
unclear why H2000 DSW does not affect liver fat accumulation, serum
TG, TC and LDL-c levels, and hepatic LDLR expression, it may be
hypothesized that these effects may be associated with Mg and Ca
concentration. Consequently, the results of the present study
indicated that H1500 DSW may be most suitable for preventing liver
fat accumulation and hypercholesterolemia.
In conclusion, the present study assessed the
effects of DSW on HCD-induced hepatic lipid accumulation and
hypercholesterolemia in rats. The results demonstrated that DSW
decreased TG, TC, LDL-c, GOT, GPT and ALP levels in the blood, and
reduced lipid accumulation in the liver. Furthermore, the mRNA
expression levels of FAS and SREBP-1c were downregulated, whereas
the expression of LDLR was upregulated by DSW. Combined, these
results indicated that DSW may have the potential to prevent
hepatic lipid accumulation and may exert blood cholesterol-lowering
activity via the inhibition of fatty acid synthesis in the liver
and enhancement of LDL-c clearance in the blood, caused by
increased hepatic LDLR expression. The present study indicated that
DSW is a candidate for the prevention of hypercholesterolemia and
hepatic lipid accumulation.
Acknowledgements
This work was financially supported by the National
R&D Project ‘Development of New Application Technology For Deep
Sea Water Industry’ supported by the Ministry of Oceans and
Fisheries of the Republic of Korea.
References
|
1
|
Ma Y, Wang W, Zhang J, Lu Y, Wu W, Yan H
and Wang Y: Hyperlipidemia and atherosclerotic lesion development
in Ldlr-deficient mice on a long-term high-fat diet. PLoS One.
7:e358352012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhang X, Wu C, Wu H, Sheng L, Su Y, Zhang
X, Luan H, Sun G, Sun X, Tian Y, et al: Anti-hyperlipidemic effects
and potential mechanisms of action of the caffeoylquinic acid-rich
Pandanus tectorius fruit extract in hamsters fed a high fat-diet.
PLoS One. 8:e619222013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Daniels SR: Management of hyperlipidemia
in pediatrics. Curr Opin Cardiol. 27:92–97. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cholesterol Treatment Trialists' (CTT)
Collaboration. Baigent C, Blackwell L, Emberson J, Holland LE,
Reith C, Bhala N, Peto R, Barnes EH, Keech A, et al: Efficacy and
safety of more intensive lowering of LDL cholesterol: A
meta-analysis of data from 170,000 participants in 26 randomised
trials. Lancet. 376:1670–1681. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Berry JD, Dyer A, Cai X, Garside DB, Ning
H, Thomas A, Greenland P, Van Horn L, Tracy RP and Lloyd-Jones DM:
Lifetime risks of cardiovascular disease. N Engl J Med.
366:321–329. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Harchaoui KE, Visser ME, Kastelein JJ,
Stroes ES and Dallinga-Thie GM: Triglycerides and cardiovascular
risk. Curr Cardiol Rev. 5:216–222. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yang ZH, Miyahara H, Takeo J, Hatanaka A
and Katayama M: Pollock oil supplementation modulates
hyperlipidemia and ameliorates hepatic steatosis in mice fed a
high-fat diet. Lipids Health Dis. 10:1892011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yao Z, Liu XC and Gu YE: Schisandra
chinensis Baill, a Chinese medicinal herb, alleviates
high-fat-diet-inducing non-alcoholic steatohepatitis in rats. Afr J
Tradit Complement Altern Med. 11:222–227. 2013.PubMed/NCBI
|
|
9
|
Carrier B, Wen S, Zigouras S, Browne RW,
Li Z, Patel MS, Williamson DL and Rideout TC: Alpha-lipoic acid
reduces LDL-particle number and PCSK9 concentrations in high-fat
fed obese Zucker rats. PLoS One. 9:e908632014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Singh AB, Kan CF, Shende V, Dong B and Liu
J: A novel posttranscriptional mechanism for dietary
cholesterol-mediated suppression of liver LDL receptor expression.
J Lipid Res. 55:1397–1407. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kim H, Bartley GE, Arvik T, Lipson R, Nah
SY, Seo K and Yokoyama W: Dietary supplementation of chardonnay
grape seed flour reduces plasma cholesterol concentration, hepatic
steatosis, and abdominal fat content in high-fat diet-induced obese
hamsters. J Agric Food Chem. 62:1919–1925. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kim S, Chun SY, Lee DH, Lee KS and Nam KS:
Mineral-enriched deep-sea water inhibits the metastatic potential
of human breast cancer cell lines. Int J Oncol. 43:1691–1700.
2013.PubMed/NCBI
|
|
13
|
Katsuda S, Yasukawa T, Nakagawa K, Miyake
M, Yamasaki M, Katahira K, Mohri M, Shimizu T and Hazama A:
Deep-sea water improves cardiovascular hemodynamics in Kurosawa and
Kusanagi-Hypercholesterolemic (KHC) rabbits. Biol Pharm Bull.
31:38–44. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lee KS, Shin JS, Kwon YS, Moon DS and Nam
KS: Suppression of cancer progression and metastasis in HT-29 human
colorectal adenocarcinomas by deep sea water. Biotechnol Bioproc
Eng. 18:194–200. 2013. View Article : Google Scholar
|
|
15
|
Fu ZY, Yang FL, Hsu HW and Lu YF: Drinking
deep seawater decreases serum total and low-density
lipoprotein-cholesterol in hypercholesterolemic subjects. J Med
Food. 15:535–541. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ha BG, Shin EJ, Park JE and Shon YH:
Anti-diabetic effect of balanced deep-sea water and its mode of
action in high-fat diet induced diabetic mice. Mar Drugs.
11:4193–4212. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hataguchi Y, Tai H, Nakajima H and Kimata
H: Drinking deep-sea water restores mineral imbalance in atopic
eczema/dermatitis syndrome. Eur J Clin Nutr. 59:1093–1096. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hsu CL, Chang YY, Chiu CH, Yang KT, Wang
Y, Fu SG and Chen YC: Cardiovascular protection of deep-seawater
drinking water in high-fat/cholesterol fed hamsters. Food Chem.
127:1146–1152. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hwang HS, Kim HA, Lee SH and Yun JW:
Anti-obesity and antidiabetic effects of deep sea water on ob/ob
mice. Mar Biotechnol (NY). 11:531–539. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hwang HS, Kim SH, Yoo YG, Chu YS, Shon YH,
Nam KS and Yun JW: Inhibitory effect of deep-sea water on
differentiation of 3T3-L1 adipocytes. Mar Biotechnol (NY).
11:161–168. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li PC, Pan CH, Sheu MJ, Wu CC, Ma WF and
Wu CH: Deep sea water prevents balloon angioplasty-induced
hyperplasia through MMP-2: An in vitro and in vivo study. PLoS One.
9:e969272014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Miyamura M, Yoshioka S, Hamada A, Takuma
D, Yokota J, Kusunose M, Kyotani S, Kawakita H, Odani K, Tsutsui Y
and Nishioka Y: Difference between deep seawater and surface
seawater in the preventive effect of atherosclerosis. Biol Pharm
Bull. 27:1784–1787. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sheu MJ, Chou PY, Lin WH, Pan CH, Chien
YC, Chung YL, Liu FC and Wu CH: Deep sea water modulates blood
pressure and exhibits hypolipidemic effects via the AMPK-ACC
pathway: An in vivo study. Mar Drugs. 11:2183–2202. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yoshioka S, Hamada A, Cui T, Yokota J,
Yamamoto S, Kusunose M, Miyamura M, Kyotani S, Kaneda R, Tsutsui Y,
et al: Pharmacological activity of deep-sea water: Examination of
hyperlipemia prevention and medical treatment effect. Biol Pharm
Bull. 26:1552–1559. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Reitman S and Frankel S: A colorimetric
method for the determination of serum glutamic oxalacetic and
glutamic pyruvic transaminases. Am J Clin Pathol. 28:56–63. 1957.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Vuppalanchi R and Chalasani N:
Nonalcoholic fatty liver disease and nonalcoholic steatohepatitis:
Selected practical issues in their evaluation and management.
Hepatology. 49:306–317. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Inoue M, Ohtake T, Motomura W, Takahashi
N, Hosoki Y, Miyoshi S, Suzuki Y, Saito H, Kohgo Y and Okumura T:
Increased expression of PPARgamma in high fat diet-induced liver
steatosis in mice. Biochem Biophys Res Commun. 336:215–222. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen IS, Chang YY, Hsu CL, Lin HW, Chang
MH, Chen JW, Chen SS and Chen YC: Alleviative effects of
deep-seawater drinking water on hepatic lipid accumulation and
oxidation induced by a high-fat diet. J Chin Med Assoc. 76:95–101.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ma PT, Gil G, Südhof TC, Bilheimer DW,
Goldstein JL and Brown MS: Mevinolin, an inhibitor of cholesterol
synthesis, induces mRNA for low density lipoprotein receptor in
livers of hamsters and rabbits. Proc Natl Acad Sci USA.
83:8370–8374. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yasunobu Y, Hayashi K, Shingu T, Nomura K,
Ohkura Y, Tanaka K, Kuga Y, Nomura S, Ohtani H, Nishimura T, et al:
Reduction of plasma cholesterol levels and induction of hepatic LDL
receptor by cerivastatin sodium (CAS 143201-11-0, BAY w 6228), a
new inhibitor of 3-hydroxy-3-methylglutaryl coenzyme A reductase,
in dogs. Cardiovasc Drugs Ther. 11:567–574. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Abad C, Vargas FR, Zoltan T, Proverbio T,
Piñero S, Proverbio F and Marín R: Magnesium sulfate affords
protection against oxidative damage during severe preeclampsia.
Placenta. 36:179–185. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Olatunji LA and Soladoye AO: Increased
magnesium intake prevents hyperlipidemia and insulin resistance and
reduces lipid peroxidation in fructose-fed rats. Pathophysiology.
14:11–15. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gupta M, Solanki MH, Chatterjee PK, Xue X,
Roman A, Desai N, Rochelson B and Metz CN: Maternal magnesium
deficiency in mice leads to maternal metabolic dysfunction and
altered lipid metabolism with fetal growth restriction. Mol Med.
20:332–340. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Niranjan G, Anitha D, Srinivasan AR, Velu
VK, Venkatesh C, Babu MS, Ramesh R and Saha S: Association of
inflammatory sialoproteins, lipid peroxides and serum magnesium
levels with cardiometabolic risk factors in obese children of South
Indian population. Int J Biomed Sci. 10:118–123. 2014.PubMed/NCBI
|
|
36
|
Jacqmain M, Doucet E, Després JP, Bouchard
C and Tremblay A: Calcium intake, body composition, and
lipoprotein-lipid concentrations in adults. Am J Clin Nutr.
77:1448–1452. 2003.PubMed/NCBI
|
|
37
|
Miyake S: The mechanism of release of
hepatic enzymes in various liver diseases. II. Altered activity
ratios of GOT to GPT in serum and liver of patients with liver
diseases. Acta Med Okayama. 33:343–358. 1979.PubMed/NCBI
|
|
38
|
Cremers J, Drent M, Driessen A, Nieman F,
Wijnen P, Baughman R and Koek G: Liver-test abnormalities in
sarcoidosis. Eur J Gastroenterol Hepatol. 24:17–24. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Morán-Salvador E, López-Parra M,
García-Alonso V, Titos E, Martínez-Clemente M, González-Périz A,
López-Vicario C, Barak Y, Arroyo V and Clària J: Role for PPARγ in
obesity-induced hepatic steatosis as determined by hepatocyte- and
macrophage-specific conditional knockouts. FASEB J. 25:2538–2550.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Deng Q, Li X, Fu S, Yin L, Zhang Y, Wang
T, Wang J, Liu L, Yuan X, Sun G, et al: SREBP-1c gene silencing can
decrease lipid deposits in bovine hepatocytes cultured in vitro.
Cell Physiol Biochem. 33:1568–1578. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chang XX, Yan HM, Xu Q, Xia MF, Bian H,
Zhu TF and Gao X: The effects of berberine on hyperhomocysteinemia
and hyperlipidemia in rats fed with a long-term high-fat diet.
Lipids Health Dis. 11:862012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Benn T, Kim B, Park YK, Yang Y, Pham TX,
Ku CS, Farruggia C, Harness E, Smyth JA and Lee JY: Polyphenol-rich
blackcurrant extract exerts hypocholesterolaemic and hypoglycaemic
effects in mice fed a diet containing high fat and cholesterol. Br
J Nutr. 113:1697–1703. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhao Y, Peng L, Yang LC, Xu XD, Li WJ, Luo
XM and Jin X: Wedelolactone regulates lipid metabolism and improves
hepatic steatosis partly by AMPK Activation and Up-regulation of
expression of PPARα/LPL and LDLR. PLoS One. 10:e01327202015.
View Article : Google Scholar : PubMed/NCBI
|