Introduction
Renal ischemia/reperfusion (IR) injury is a major
cause of acute kidney injury (AKI), the pathophysiology of which is
considered to be associated with renal tubular epithelial cell
injury (1). Vascular endothelial
cells may be involved in determining vascular permeability,
communicating with other cells and regulating vasomotion. In
addition, vascular endothelial cells may serve crucial roles in the
development of AKI. An injection of HUVECs can be implanted in the
vascular bed of the renal peritubular capillaries and significantly
improve renal function after I/R injury (2). It was hypothesized that HUVECs may
possess some characteristics or functions of renal peritubular
microvascular endothelial cells and thus HUVECs were used for in
vitro study. Delayed ischemic preconditioning (IPC) is a brief,
sublethal episode of ischemia that protects certain organs against
subsequent lethal ischemic insult and is thought to be an
endogenous mechanism of preserving organ function. The beneficial
effects of delayed IPC have been confirmed in the kidneys of rats
and mice (3–5); however, the role of renal vascular
endothelial cells in delayed IPC has not previously been
investigated.
microRNAs (miRNAs) are endogenous, short (18–22
nucleotides) RNA molecules that may be involved in the
physiological functions of the kidneys and in the pathological
processes of renal disease. Several miRNAs, including miR-200,
miR-21 and miR-133, have been previously demonstrated to be
associated with the protective effects of IPC on IR injury
(6,7). Our previous study demonstrated that
IPC significantly increased the expression of miR-21 in the mouse
kidney 24 h following IR. Knockdown of miR-21, combined with IPC,
significantly exacerbated subsequent renal IR injury (8). Other studies have demonstrated that
miR-21 is expressed in vascular endothelial cells (9,10),
and that programmed cell death 4 (PDCD4) is a proapoptotic target
gene of miR-21 (8). The present
study focused on vascular endothelial cells and hypothesized that
the protective role of miR-21 in renal delayed IPC may be
associated with reduced endothelial cell apoptosis by targeting
PDCD4.
Materials and methods
Mouse models of delayed renal IPC and
IR
A total of 60 male C57BL/6 mice (weight, 20–23 g;
age, 6–7 weeks) were housed in the Animal Center of Zhongshan
Hospital of Fudan University at 24–25°C, 5% CO2, free
access to food and water, and 16-h light/8-h dark cycle. The mice
were anesthetized intraperitoneally with 1% pentobarbital (50
mg/kg; Sigma-Aldrich, Merck KGaA, Darmstadt, Germany). Following
midline laparotomy, the bilateral renal pedicles were clamped for
15 min using micro-serrefine clips (Fine Science Tools, Inc.,
Foster City, CA, USA) to induce IPC. Mice were maintained at
35–37°C and the abdominal cavity was hydrated with saline-moistened
gauze. Mice in the IPC + IR group were subjected to 35 min
bilateral kidney ischemia 4 days post-IPC, followed by reperfusion
for 24 h. Mice in the Sham group underwent the same surgical
procedures, except that the renal pedicles were not clamped (no
IPC). Following treatment, the mice were anesthetized
intraperitoneally with 1% pentobarbital (50 mg/kg) at 24 h after
reperfusion, and then the blood samples were taken by cardiac
puncture and the kidneys collected; one kidney was snap-frozen in
liquid nitrogen for protein and RNA isolation followed by
transference to a −80°C freezer, and the other kidney was fixed for
histological analysis. Serum creatinine (SCr) was measured as
previously described by a Quantichrom creatinine Assay kit
(BioAssay Systems, Hayward, CA, USA) (8). The study was approved by the ethics
committee of Zhongshan Hospital of Fudan University (Shanghai,
China).
Histological analysis of renal injury
and immunohistochemical staining
Kidney tissues were fixed in 10% neutral-buffered
formalin at room temperature for 24 h and embedded in paraffin.
Tissues were sectioned (4 µm), deparaffinized and stained with
periodic acid-Schiff counterstained with alum hematoxylin.
Histopathological changes were examined in a blinded manner by
scoring tubular cell necrosis or swelling, interstitial
infiltration by multinucleated cells, tubular casts and brush
border loss; sections were scored according to the severity of
changes on a semi-quantitative scale: No injury (0); mild, <25%
(1); moderate, <50% (2); severe, <75% (3); and very severe, >75% (4). For immunohistochemistry, kidney
sections were deparaffinized in dimethylbenzene twice, dehydrated
in gradient ethanol and endogenous peroxidase activity was
eliminated by 3% H2O2 incubation at room
temperature for 30 min. The sections were blocked with 10% goat
serum (Sigma-Aldrich, Merck KGaA) for 20 min at room temperature
and incubated with monoclonal rat anti-mouse CD31 antibody (ab7388,
1:50; Abcam, Cambridge, MA, USA) overnight at 4°C. Antibody
dilution and washing steps were performed with PBS. The secondary
antibody incubation and staining was carried out by GTVision II
Immunohistochemistry Detection System/Mo&Rb (GK500611A; Gene
Tech Biotechnology Co., Ltd., Shanghai, China) according to the
manufacturer's protocol.
miR-21 in situ hybridization
(ISH)
ISH was performed on the formalin-fixed
paraffin-embedded kidney sections (4 µm) by microRNA ISH
Optimization kit 2 (Exiqon A/S, Vedbaek, Denmark) with 5′- and
3′-digoxigenin (DIG)-labeled miR-21 probes and U6 as a positive
control, according to the manufacturer's protocols, with minor
modifications. Briefly, tissue sections were deparaffinized in
xylene and rehydrated using an ethanol gradient. Sections were
treated with proteinase K (20 mg/ml) for 10 min at 37°C and
subsequently incubated with hybridization buffer at room
temperature for 30 min. The probes were diluted in hybridization
buffer (40 nM for miR-21 and 1 nM for U6) and preheated at 90°C for
5 min to linearize prior to adding to the slides (50 µl/tissue
section). Following incubation at 53°C for 2 h, slides were rinsed
twice (5 min each) in 5X saline sodium citrate (SSC), 1X SSC and
0.1X SSC at 53°C, followed by washes with PBS-Tween-20 (0.1%), and
blocking with sheep serum (2%; Jackson ImmunoResearch Laboratories,
Inc., West Grove, PA, USA) for 30 min at 37°C. Slides were
subsequently incubated with 1:800 anti-DIG alkaline phosphatase
antibody (11 093 274 910; Roche Applied Science, Rotkreuz,
Switzerland) for 1 h at room temperature, followed by incubation in
nitro blue tetrazolium/5-bromo-4-chloro-3-indolyl phosphate (11 697
471 001; Roche Applied Science) diluted in double-distilled
H2O for 2 h at 34°C. Slides were rinsed with alkaline
phosphatase stop solution to halt the color development, prior to
dehydration with 70, 96 and 99.99% ethanol in sequence (1 min
followed by immersing 10 times for each ethanol), clearing with
dimethylbenzene for 5 min and mounting in Eukitt®
(Sigma-Aldrich; Merck KGaA) at room temperature. The tissue was
visualized by microscope (Leica DM6000 B; Leica Microsystems GmbH,
Wetzlar, Germany) and images captured and analyzed by Leica
Application Suite software version 4.5 (Leica Microsystems
GmbH).
Assessment of renal vascular protein
leakage using Evans blue dye
Evans blue dye was used to assess vascular leakage
to evaluate renal microvascular permeability as previously
described (11). Mice were
injected intravenously with Evans blue dye (2 ml/kg; Sigma-Aldrich;
Merck KGaA) 30 min prior to sacrifice. Following sacrifice, mice
were perfused with PBS through the left ventricle to completely
eliminate blood. The kidneys were weighed and subsequently
homogenized in 1 ml pure formamide (Sigma-Aldrich; Merck KGaA) and
incubated at 55°C for 18 h. The supernatant was collected following
centrifugation (10,000 × g for 30 min at room temperature). The
amount of Evans blue dye in the supernatant was analyzed by
measuring absorbance at 620 nm. Results were calculated from a
standard curve of Evans blue dye and expressed as the concentration
of Evans blue dye (µg)/kidney weight (g).
Cell culture and cobalt (II) chloride
(CoCl2) treatment
HUVEC were obtained from American Type Culture
Collection (Manassas, VA, USA) and cultured in endothelial cell
medium with 5% fetal bovine serum (Sciencell Research Laboratories,
Inc., Carlsbad, CA, USA) at 37°C with 95% air and 5%
CO2. HUVECs at 60–70% confluence were exposed to 150 µM
CoCl2 for 24 h under 37°C to mimic hypoxia or placed in fresh
normal medium as a control as previously described (12). Following CoCl2 incubation, the
cells were harvested for RNA, protein extraction or flow
cytometry.
In vitro and in vivo use of locked
nucleic acid (LNA)-modified anti-miR-21 oligonucleotides
LNA-modified anti-miR-21 oligonucleotides (Exiqon
A/S) were used to suppress miR-21 expression in both in
vitro and in vivo experiments with the scrambled
oligonucleotides (Exiqon A/S) as the control. The sequence of the
anti-miR-21 and scrambled control are as follows: LNA anti miR-21
5′-FAM-TCAG TCT GAT AAG CT-3′; scrambled control: 5′-FAM-ACGT CTA
TAC GCC CA-3′. HUVECs at 80–90% confluence (1×106/well
in a 6 well plate) were transfected with anti-miR-21 and scrambled
control oligonucleotides (100 nM), using Lipofectamine®
2000 (Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
according to the manufacturer's protocols. The final concentration
of LNA™ microRNA inhibitors (LNA-oligos) was decided according to
the suggested concentration from LNA-oligos and
Lipofectamine® 2000 from the manufacturer combined with
the dose from a previous study (13). Briefly, LNA mix was incubated in
the Lipofectamine® 2000 mix for 20 min at room
temperature before pouring into dishes. The transfection medium was
replaced with the regular medium 4 h post-transfection. Following
20 h incubation, cells were treated with CoCl2. LNA-modified
anti-miR-21 and scrambled control oligonucleotides used for in
vivo suppression experiments were administered by intravenous
tail vein injection (10 mg/kg body weight) <1 h prior to
ischemia surgery (8).
TaqMan reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA from kidney tissue (1/4 kidney) and cells
with 80–90% confluence in 3.5 cm dishes was isolated using TRIzol
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. The RNA purity was determined according to
the ratio of A260/A280 (1.8–2.0). RT-qPCR of miR-21 were performed
with the TaqMan MicroRNA Reverse Transcription kit (Applied
Biosystems; Thermo Fisher Scientific, Inc.) for RT and then the
TaqMan(R) Universal PCR Master Mix (Applied Biosystems) for cDNA
amplification according to the manufacturer's protocol (14). U6 was used as internal control to
normalize miRNA expression. Ambion® (Thermo Fisher
Scientific) RT and PCR primers of miR-21 (has-miR-21; ID 000397)
and U6 (U6 snRNA ID 001973) are special for TaqMan®
MicroRNA Assays. The program of 16°C for 30 min, 42°C for 30 min
and 85°C for 5 min was used for RT. qPCR was conducted at 95°C for
10 min, followed by 40 cycles of 95°C for15 sec and 60°C for 1 min
in the Applied Biosystems® 7500 Real-Time PCR Systems
(Thermo Fisher Scientific, Inc.). The 7500 software version 2.0.6
was used for analysis. The expression level of miR-21 was analyzed
as described previously (8) and
the 2−∆∆Cq method was used for normalization (15).
Western blot analysis
The cells were lysed by cold RIPA lysis buffer (with
1 mM phenylmethylsulfonyl fluoride; KeyGen Biotech Co., Ltd.,
Nanjing, China). Half of one kidney was frozen and ground in liquid
nitrogen, then disrupted in cold lysis buffer. Supernatants were
collected after centrifuged. The BCA method of protein
quantification was used with the kit (KeyGen Biotech Co., Ltd.).
Protein samples (40 µg per lane) were separated by 10% SDS-PAGE and
transferred to a polyvinylidene difluoride membrane. The describe
membrane was blocked with 5% nonfat milk for 1 h at room
temperature, followed by incubation with primary antibodies against
PDCD4 (rabbit polyclonal; NBP1-76738; 1:1,000; Novus Biologicals,
LLC, Littleton, CO, USA) or β-actin (mouse monoclonal; 1:1,000;
Abcam) overnight at 4°C. The secondary antibodies included
horseradish peroxidase-conjugated anti-rabbit or anti-mouse
immunoglobulin G (sc-2372 for anti-rabbit and sc-2005 for
anti-mouse; 1:5,000; Santa Cruz Biotechnology, Inc.) and were used
for incubation for 1.5 h at room temperature. Protein bands were
developed with ECL Western Blotting Detection Reagents (Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocol.
The results were normalized to the protein levels of β-actin.
Flow cytometry analysis for
apoptosis
Apoptosis in cultured cells was measured by flow
cytometry with the Annexin V-FITC/PI Apoptosis Detection kit
(KeyGen Biotech Co., Ltd.) according to the manufacturer's
protocol. At 24 h following CoCl2 treatment with or without
LNA-oligos, the cells were harvested, washed with PBS and stained
with Annexin V-fluorescein isothiocyanate (FITC) and propidium
iodide (PI) for 10 min in a dark environment, and detection was
performed by BD FACSAria™ II Flow Cytometer (BD Biosciences, San
Jose, CA, USA). The data from flow cytometry was analyzed by FlowJo
software version 7.6.1 (FlowJo LLC, Ashland, OR, USA). Early
apoptotic cells exhibited Annexin V-FITC+/PI- staining patterns,
whereas late apoptotic cells exhibited Annexin V-FITC+/PI+ staining
patterns in the flow cytometry plots. All experiments were
performed in triplicate.
Statistical analysis
SPSS version 16.0 (SPSS, Inc., Chicago, IL, USA) was
used for statistical analysis. Data were analyzed using the
unpaired Student's t-test when comparing between 2 groups and
one-way analysis of variation with Tukey's multiple comparison when
comparing >2 groups. RT-qPCR data are presented as a percentage
of the control since data from multiple PCR plates were combined.
For these data, statistical analysis was performed on the original
data before conversion to percentage values. P<0.05 was
considered to indicate a statistically significant difference. Data
are presented as the mean ± standard error of the mean.
Results
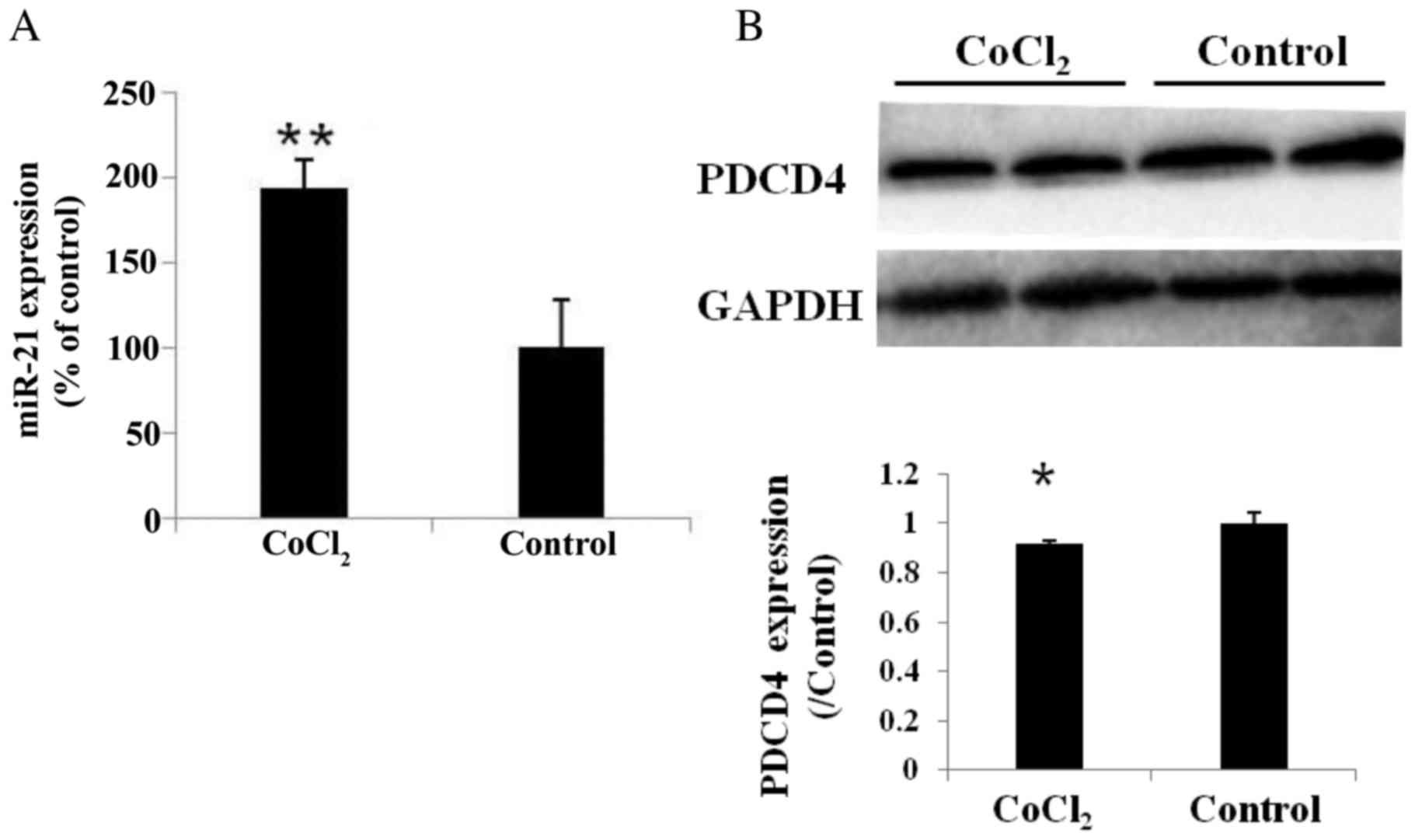
CoCl2 treatment affects the
expression of miR-21 and PDCD4 protein in HUVECs
Our previous study demonstrated that hypoxic
conditions were able to induce the expression of miR-21 in human
renal epithelial cells (13). The
present study used CoCl2 to mimic hypoxia in vitro, and
HUVECs treated with CoCl2 for 24 h exhibited a significant
upregulation of miR-21 expression compared with the control group
(193.44±16.32% of the control; P<0.01; Fig. 1A). In addition, the expression of
PDCD4 protein in the CoCl2 and control groups was investigated by
western blot analysis. PDCD4 protein expression was significantly
reduced compared with the control group (P<0.05; Fig. 1B).
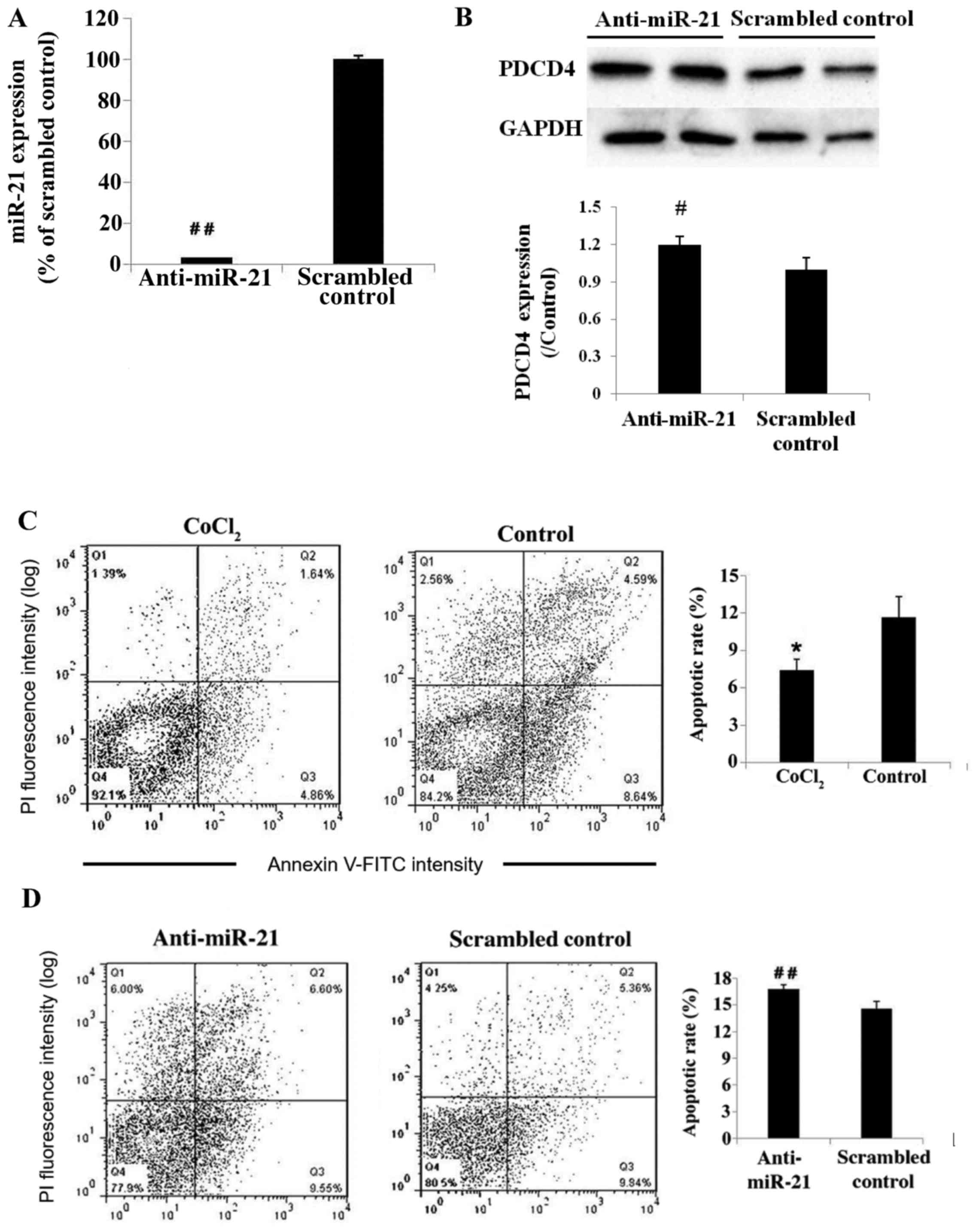
Knockdown of miR-21 increases
expression of PDCD4 and cell apoptosis in hypoxic conditions in
vitro
The CoCl2-induce increase in miR-21 expression was
significantly reduced in HUVECs transfected with LNA-modified
anti-miR-21 oligonucleotides, compared with scrambled control
(3.19% of scrambled control; P<0.01; Fig. 2A). PDCD4 protein expression
increased significantly in the anti-miR-21 group, compared with
scrambled control (P<0.05; Fig.
2B). In addition, the results demonstrated that treatment with
CoCl2 significantly attenuated apoptosis of HUVECs compared with
the untreated control group (P<0.05; Fig. 2C). However, CoCl2-treated HUVECs
transfected with anti-miR-21 exhibited an increase in the number of
apoptotic cells compared with hypoxic cells transfected with the
scrambled control (P<0.01; Fig.
2D).
miR-21 expression in renal vascular
endothelial cells contributes to the protective effects of renal
delayed IPC
Renal IPC for 15 min resulted in a significant
increase in the expression of miR-21 in the kidney 24 h following
the second IR, compared with mice in the Sham + IR group
(179.18±17.42% of the Sham + IR group; P<0.05; Fig. 3A). LNA-modified scrambled control
or anti-miR-21 oligonucleotides were administered to mice through
the tail vein prior to IPC surgery; treatment with anti-miR-21
significantly reduced the expression of miR-21 in the kidney 24 h
following the second IR compared with the scrambled control group
(2.43±0.71% of the scrambled control group; P<0.01; Fig. 3B). ISH for miR-21 expression was
examined in the cortex and medulla in the kidney under all
experimental conditions, and the results revealed that the
expression of miR-21 was increased in IPC + IR mice that were
co-treated with scrambled control, and suppressed with anti-miR-21
treatment (Fig. 3C). The result of
ISH in the scrambled control group shown that miR-21 could express
in the vascular endothelial cells (arrows in Fig. 3C) in addition to the tubular
epithelial cells (circles in Fig.
3C); however, this pattern of miR-21 expression was not
observed in either region in IPC + IR mice treated with
anti-miR-21. The expression of U6 positive control is shown in the
anti-scrambled control. To further identify the distribution of
miR-21 expression in vascular endothelial cells,
immunohistochemical staining of CD31 for vascular endothelial cells
and ISH for miR-21 in two serial sections was performed. As
demonstrated in Fig. 3D, the
positive expression area of CD31 and miR-21 are similar as
indicated by similar shaped arrows in the two images.
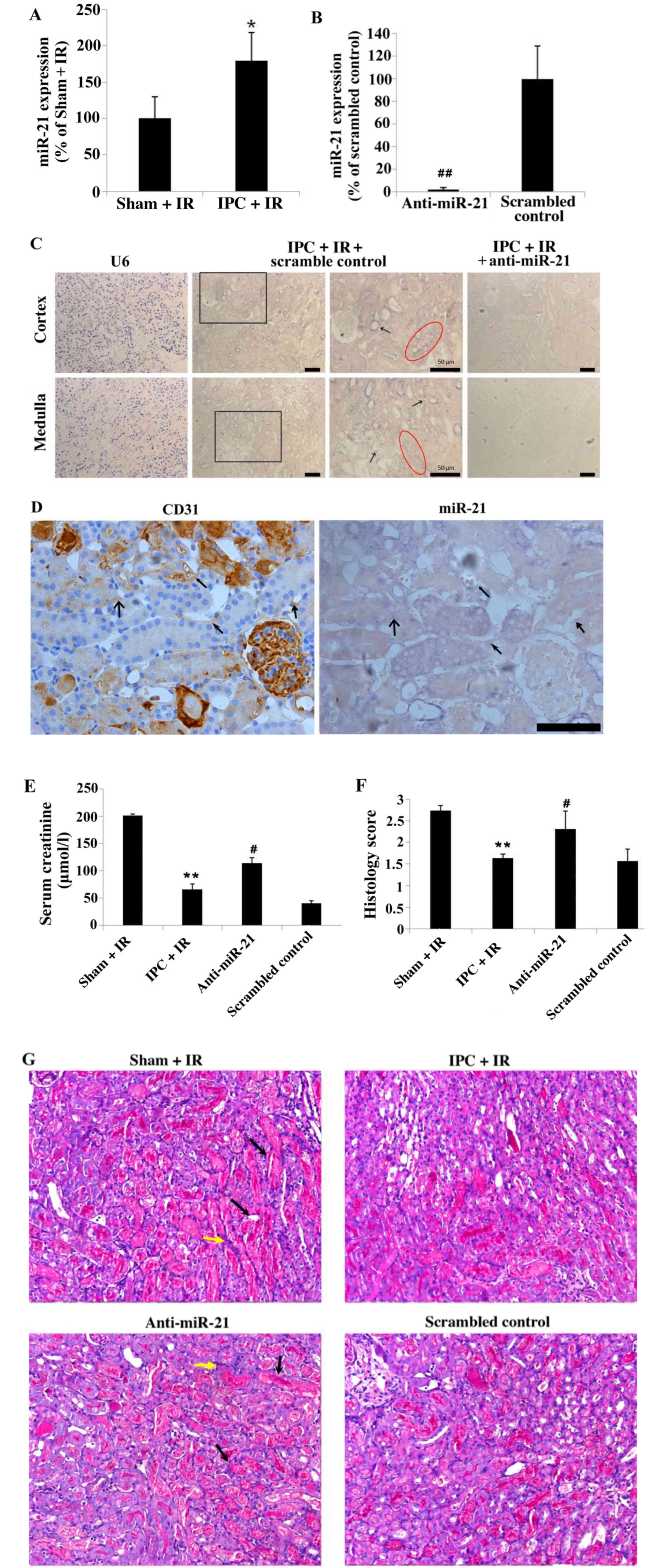 | Figure 3.Delayed IPC increased miR-21
expression in renal tubular and in endothelial cells, which was
inhibited by locked nucleic acid-modified anti-miR-21
oligonucleotide treatment. (A) miR-21 expression 24 h following IR
was increased in kidneys exposed to IPC compared with Sham + IR
mice. (B) Expression of miR-21 in the renal tissue 24 h following
IR was inhibited by treatment with anti-miR-21 administered at the
time of IPC, compared with mice treated with the scrambled control
oligonucleotides. (C) Representative images of U6 (positive
control) and miR-21 expression in mouse kidney sections by in
situ hybridization. The increase of miR-21 expression was
notable in vascular endothelial cells (arrows) of IPC + IR +
scrambled control-treated mice in addition to the renal tubular
epithelial cells (red circle). Magnification, ×20; scale bar, 50
µm. (D) miR-21 expression in vascular endothelial cells, which were
marked by CD31 staining and is indicated by the similar shaped
arrows in the two images. (E) Serum creatinine and (F) histology
score 24 h following IR were attenuated by delayed IPC, wherease
miR-21 knockdown in mice exposed to delayed IPC significantly
worsened renal IR injury. (G) Histopathological changes of mice
kidney sections from each group. Representative periodic
acid-Schiff-stained micrographs in the corticomedullary junction.
Magnification, ×20. Black arrow indicates cast formation and yellow
arrow indicates infiltration of inflammatory cells. Data are
presented as the mean ± standard error of the mean. n=6 per group;
*P<0.05 and **P<0.01 vs. Sham + IR group;
#P<0.05 and ##P<0.01 vs. scrambled
control group. IPC, ischemic preconditioning; IR,
ischemia/reperfusion; miR-21, microRNA-21; U6, U6 small nuclear
RNA. |
Delayed IPC significantly attenuated IR-induced
renal dysfunction, as demonstrated by the renal function and
histological damage observed (Fig.
3E-G). Compared with the Sham + IR group, IPC + IR mice
exhibited a significant decrease in SCr (P<0.01; Fig. 3E) and histology score (P<0.01;
Fig. 3F). However, treatment with
anti-miR-21 reversed the protective effect of IPC in IR mice: The
anti-miR-21 group had significantly increased SCr and histology
score compared with the scrambled control (P<0.05; Fig. 3E and F, respectively) and exhibited
characteristics of acute tubulointerstitial damage similar to that
observed in the Sham + IR group, including massive tubular
epithelial cell necrosis or swelling, tubular casts and
inflammatory cell infiltration (Fig.
3G).
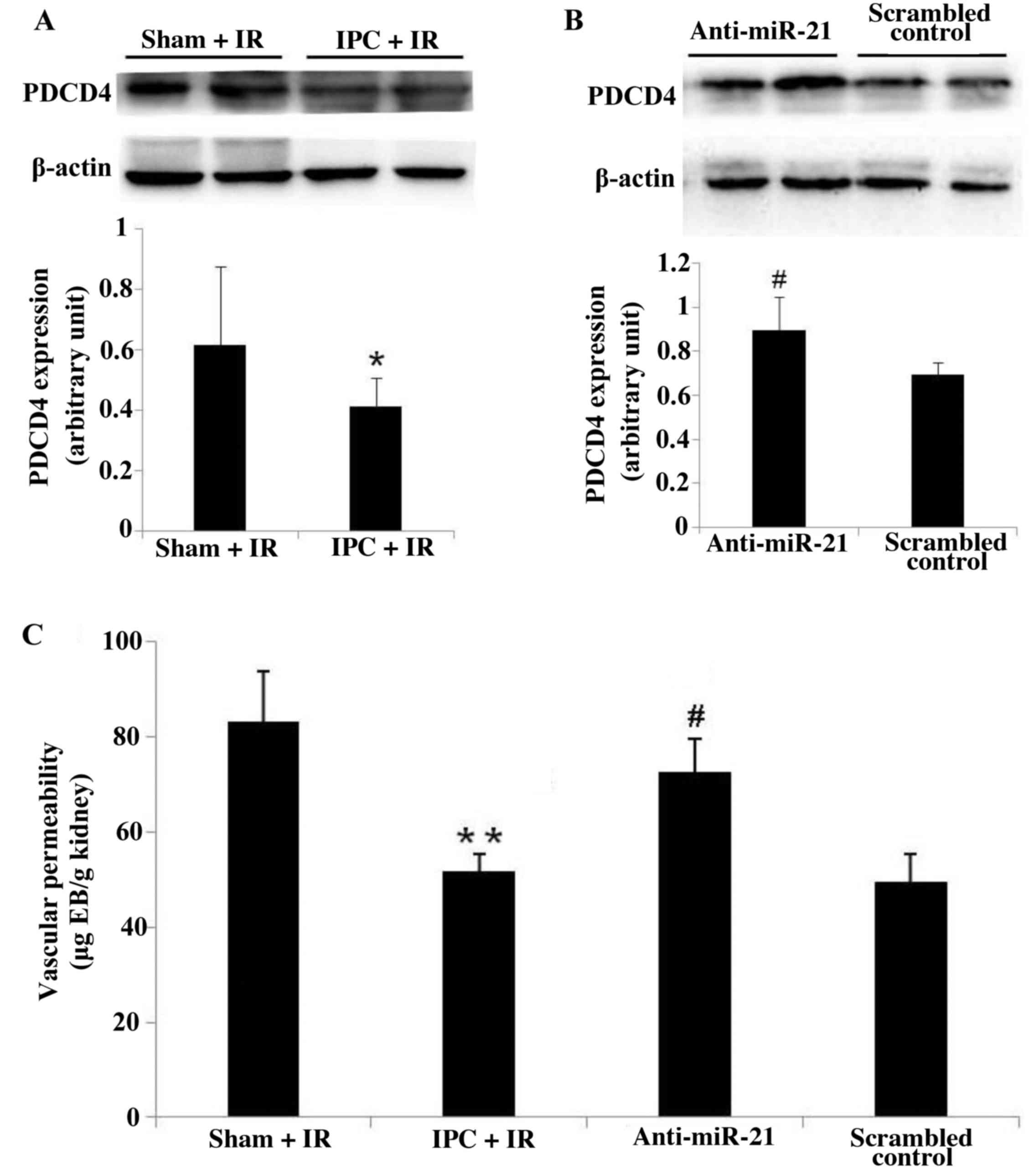
miR-21 knockdown may increase vascular
endothelial cell apoptosis by increasing PDCD4 expression
The expression of PDCD4 protein was examined in mice
from all groups. PDCD4 protein expression was significantly
downregulated in IPC + IR mice compared with Sham + IR mice
(P<0.05; Fig. 4A), whereas
PDCD4 protein expression was significantly upregulated by in IPC +
IR mice treated with anti-miR-21 compared with mice in the
scrambled control group (P<0.05; Fig. 4B), which was consistent with the
suggestion that PDCD4 may be targeted by miR-21 (16). Renal microvascular permeability was
measured by Evans blue dye leakage, and was assessed to indirectly
evaluate apoptosis of vascular endothelial cells. The amount of
Evans blue dye in the kidney of IPC + IR mice was significantly
lower compared with Sham + IR mice (P<0.01; Fig. 4C). In addition, IPC + IR mice that
received anti-miR-21 treatment exhibited a significant increase in
the amount of Evans blue dye in the kidney compared with IPC + IR
mice treated with scrambled control (P<0.05; Fig. 4C), which suggested an increase in
apoptosis of vascular endothelial cells in the kidney. The results
indicated that miR-21 may attenuate vascular endothelial cell
apoptosis by targeting PDCD4 and may serve a role in the protective
effects of delayed IPC.
Discussion
The present study, to the best of our knowledge, is
the first to focus on the involvement of vascular endothelial cells
in renal delayed IPC. Tubular epithelial cells had been considered
to the primarily target of renal ischemic insults (17), whereas the functional role of
endothelial cells has received less attention. The renal
microvascular endothelium has been reported to serve a vital role
in the process of renal IR injury (18). It has been previously demonstrated
that injury or apoptosis of the vascular endothelial cells may
contribute to functional impairment of peritubular
microcirculation, which triggers ischemic AKI (19–21).
The present study hypothesized that the loss of barrier function
may reflect endothelial cell apoptosis in the mouse kidney. A
previous study demonstrated that the activation of caspase-3
protein, a biomarker for endothelial cell apoptosis, was consistent
with increased microvascular permeability following ischemic
injury, although they did not observe apoptosis by TUNEL staining
(22). Owing to the rapid
clearance of apoptotic cells and the moderate sensitivity of the
TUNEL reaction in ischemic kidney tissue, the potential for
positive apoptosis of endothelial cell to be observed by TUNEL is
low. Therefore, the present study opted to indirectly evaluate
apoptosis injury of microvascular endothelial cells by
microvascular permeability assessment. Previous reports have
indicated that the improvement of endothelial cells function by
exogenous VEGF-121 or lecithinized superoxide dismutase, preserving
renal microvessel structure or increasing free radical scavenging,
may protect against IR injury and improve the prognosis of the AKI
(23,24). The present study hypothesized that
the protective mechanism of renal hypoxic/ischemic intolerance may
be associated with the antiapoptotic ability of renal vascular
endothelial cells. However, the increased permeability of
microvascular endothelial cells in this study may also have been
the result of tubular epithelial cell injury and the resultant
inflammation; which is a limitation of the present study.
miR-21 is an important antiapoptotic miRNA (25). Certain studies have suggested that
miR-21 expression may be increased in cells tolerant to hypoxia and
may be protective against hypoxia (13,26,27).
In tumor cells (ovarian or mammary carcinoma cell lines), miR-21
expression has been demonstrated to be induced through the Akt-2
pathway in hypoxic conditions and miR-21 expression promoted tumor
resistance (28). Our previous
study demonstrated that miR-21 expression was elevated by hypoxia
(2% O2 and CoCl2) treatment in human renal
epithelial cells (8). The present
study demonstrated that optimal hypoxia, that causing the least
cell injury, may have decreased vascular endothelial cell
apoptosis, consistent with the overexpression of miR-21 in HUVECs
in vitro. Knockdown of miR-21 in hypoxic vascular
endothelial cells led to a potential increase in apoptosis. PDCD4
was previously demonstrated to be a direct target of miR-21
(16), and miR-21 was reported to
inhibit apoptosis by downregulating PDCD4 expression in the heart
(29), cultured mouse tubular
epithelial cells (30) and in
mouse kidneys (8). Although the
present study confirmed the antiapoptotic mechanism of miR-21
targeting PDCD4 in HUVECs in vitro, it is likely that the
effect of miR21-PDCD4 pathway is partly from the endothelial cell
since this pathway was confirmed in the tubular epithelial cells
(30). In addition, phosphatase
and tensin homolog deleted on chromosome 10 (PTEN) (31), Fas ligand (27) and tropomyosin 1 (32) are inhibited by miR-21, which leads
to tolerance to cellular apoptotic signals. We aim to investigate
other miR-21 target genes in endothelial cells in the future.
Delayed IPC has been confirmed to be protective
against renal IR injury in the rodent kidney (8,33),
and miRNAs have been previously demonstrated to be involved in the
protective mechanism of IPC (8,34). A
protective effect of miR-21 was reported in cardiac IPC (29,34,35);
these studies reported that IPC increased miR-21 expression, which
was accompanied by reduced apoptosis in the border between the
infarcted and noninfarcted areas, and decreased infarct size.
Another study demonstrated that increased miR-21 expression was
most significant in the rat cerebral cortex 24 h post-IPC (14). Our previous study reported the
protective role of miR-21 in renal delayed IPC, which was
associated with decreased apoptosis of renal tubular epithelial
cells (8). It has also been
reported that high expression of miR-21 occurred in vascular
endothelial cells (36,37). In addition, miR-21 was demonstrated
to be significantly upregulated in vascular walls following balloon
injury (9). The present study
hypothesized that miR-21 expression in microvascular endothelial
cells may contribute to the protection of renal delayed IPC against
IR injury by attenuating injury of endothelial cells. The results
suggested that knockdown of miR-21 can increase the damage to
microvascular endothelial cells induced by renal IR injury 4 days
after 15 min ischemic pretreatment, which attenuated the protection
of delayed IPC. As multiple pathways have been implicated in the
protective mechanism of renal delayed IPC, including a series of
protective mediators and/or effectors, such as c-Jun N-terminal
kinase, hypoxia-inducible factor, inducible nitric oxide synthase
and heat shock protein 27 (4,5), it
is not unexpected that the knockdown of miR-21 had only a modest
effect on the preconditioning effects in the present study.
In conclusion, renal protection conferred by delayed
IPC may potentially be mediated by several mechanisms in different
types of renal cells. The present study demonstrated that delayed
IPC provided renal protection against IR injury by attenuating
damage of renal microvascular endothelial cells, which may be
partly attributed to miR-21.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (grant no. 81300570) and the Foundation
of Zhongshan Hospital grant.
References
|
1
|
Bonventre JV and Yang L: Cellular
pathophysiology of ischemic acute kidney injury. J Clin Invest.
121:4210–4221. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Brodsky SV, Yamamoto T, Tada T, Kim B,
Chen J, Kajiya F and Goligorsky MS: Endothelial dysfunction in
ischemic acute renal failure: Rescue by transplanted endothelial
cells. Am J Physiol Renal Physiol. 282:F1140–F1149. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jiang SH, Liu CF, Zhang XL, Xu XH, Zou JZ,
Fang Y and Ding XQ: Renal protection by delayed ischaemic
preconditioning is associated with inhibition of the inflammatory
response and NF-kappaB activation. Cell Biochem Funct. 25:335–343.
2007. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Joo JD, Kim M, D'Agati VD and Lee HT:
Ischemic preconditioning provides both acute and delayed protection
against renal ischemia and reperfusion injury in mice. J Am Soc
Nephrol. 17:3115–3123. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Park KM, Chen A and Bonventre JV:
Prevention of kidney ischemia/reperfusion-induced functional injury
and JNK, p38, and MAPK kinase activation by remote ischemic
pretreatment. J Biol Chem. 276:11870–11876. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Salloum FN, Yin C and Kukreja RC: Role of
microRNAs in cardiac preconditioning. J Cardiovasc Pharmacol.
56:581–588. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lee ST, Chu K, Jung KH, Yoon HJ, Jeon D,
Kang KM, Park KH, Bae EK, Kim M, Lee SK and Roh JK: MicroRNAs
induced during ischemic preconditioning. Stroke. 41:1646–1651.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xu X, Kriegel AJ, Liu Y, Usa K, Mladinov
D, Liu H, Fang Y, Ding X and Liang M: Delayed ischemic
preconditioning contributes to renal protection by upregulation of
miR-21. Kidney Int. 82:1167–1175. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ji R, Cheng Y, Yue J, Yang J, Liu X, Chen
H, Dean DB and Zhang C: MicroRNA expression signature and
antisense-mediated depletion reveal an essential role of microRNA
in vascular neointimal lesion formation. Circ Res. 100:1579–1588.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Guduric-Fuchs J, O'Connor A, Cullen A,
Harwood L, Medina RJ, O'Neill CL, Stitt AW, Curtis TM and Simpson
DA: Deep sequencing reveals predominant expression of miR-21
amongst the small non-coding RNAs in retinal microvascular
endothelial cells. J Cell Biochem. 113:2098–2111. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yasuda H, Yuen PS, Hu X, Zhou H and Star
RA: Simvastatin improves sepsis-induced mortality and acute kidney
injury via renal vascular effects. Kidney Int. 69:1535–1542. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen Y, Jiang S, Zou J, Zhong Y and Ding
X: Silencing HIF-1α aggravates growth inhibition and necrosis of
proximal renal tubular epithelial cell under hypoxia. Ren Fail.
38:1726–1734. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jiao X, Xu X, Fang Y, Zhang H, Liang M,
Teng J and Ding X: miR-21 contributes to renal protection by
targeting prolyl hydroxylase domain protein 2 in delayed ischaemic
preconditioning. Nephrology (Carlton). 22:366–373. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dharap A, Bowen K, Place R, Li LC and
Vemuganti R: Transient focal ischemia induces extensive temporal
changes in rat cerebral microRNAome. J Cereb Blood Flow Metab.
29:675–687. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Frankel LB, Christoffersen NR, Jacobsen A,
Lindow M, Krogh A and Lund AH: Programmed cell death 4 (PDCD4) is
an important functional target of the microRNA miR-21 in breast
cancer cells. J Biol Chem. 283:1026–1033. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Eltzschig HK and Eckle T: Ischemia and
reperfusion-from mechanism to translation. Nat Med. 17:1391–1401.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rabelink TJ, Wijewickrama DC and de Koning
EJ: Peritubular endothelium: The Achilles heel of the kidney?
Kidney Int. 72:926–930. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Basile DP, Donohoe D, Roethe K and Osborn
JL: Renal ischemic injury results in permanent damage to
peritubular capillaries and influences long-term function. Am J
Physiol Renal Physiol. 281:F887–F899. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yamamoto T, Tada T, Brodsky SV, Tanaka H,
Noiri E, Kajiya F and Goligorsky MS: Intravital videomicroscopy of
peritubular capillaries in renal ischemia. Am J Physiol Renal
Physiol. 282:F1150–F1155. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Snoeijs MG, Vink H, Voesten N, Christiaans
MH, Daemen JW, Peppelenbosch AG, Tordoir JH, Peutz-Kootstra CJ,
Buurman WA, Schurink GW and van Heurn LW: Acute ischemic injury to
the renal microvasculature in human kidney transplantation. Am J
Physiol Renal Physiol. 299:F1134–F1140. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Horbelt M, Lee SY, Mang HE, Knipe NL, Sado
Y, Kribben A and Sutton TA: Acute and chronic microvascular
alterations in a mouse model of ischemic acute kidney injury. Am J
Physiol Renal Physiol. 293:F688–F695. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Leonard EC, Friedrich JL and Basile DP:
VEGF-121 preserves renal microvessel structure and ameliorates
secondary renal disease following acute kidney injury. Am J Physiol
Renal Physiol. 295:F1648–F1657. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Koo DD, Welsh KI, West NE, Channon KM,
Penington AJ, Roake JA, Morris PJ and Fuggle SV: Endothelial cell
protection against ischemia/reperfusion injury by lecithinized
superoxide dismutase. Kidney Int. 60:786–796. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xu X, Kriegel AJ, Jiao X, Liu H, Bai X,
Olson J, Liang M and Ding X: miR-21 in ischemia/reperfusion injury:
A double-edged sword? Physiol Genomics. 46:789–797. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Qin Y, Yu Y, Dong H, Bian X, Guo X and
Dong S: MicroRNA 21 inhibits left ventricular remodeling in the
early phase of rat model with ischemia-reperfusion injury by
suppressing cell apoptosis. Int J Med Sci. 9:413–423. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Buller B, Liu X, Wang X, Zhang RL, Zhang
L, Hozeska-Solgot A, Chopp M and Zhang ZG: MicroRNA-21 protects
neurons from ischemic death. FEBS J. 277:4299–4307. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Polytarchou C, Iliopoulos D,
Hatziapostolou M, Kottakis F, Maroulakou I, Struhl K and Tsichlis
PN: Akt2 regulates all Akt isoforms and promotes resistance to
hypoxia through induction of miR-21 upon oxygen deprivation. Cancer
Res. 71:4720–4731. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cheng Y, Zhu P, Yang J, Liu X, Dong S,
Wang X, Chun B, Zhuang J and Zhang C: Ischaemic
preconditioning-regulated miR-21 protects heart against
ischaemia/reperfusion injury via anti-apoptosis through its target
PDCD4. Cardiovasc Res. 87:431–439. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Godwin JG, Ge X, Stephan K, Jurisch A,
Tullius SG and Iacomini J: Identification of a microRNA signature
of renal ischemia reperfusion injury. Proc Natl Acad Sci USA.
107:14339–14344. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jia P, Teng J, Zou J, Fang Y, Wu X, Liang
M and Ding X: Xenon protects against septic acute kidney injury via
miR-21 target signaling pathway. Crit Care Med. 43:e250–e259. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Qi L, Bart J, Tan LP, Platteel I, Sluis
Tv, Huitema S, Harms G, Fu L, Hollema H and Berg Av: Expression of
miR-21 and its targets (PTEN, PDCD4, TM1) in flat epithelial atypia
of the breast in relation to ductal carcinoma in situ and invasive
carcinoma. BMC Cancer. 9:1632009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jiang S, Chen Y, Zou J, Xu X, Zhang X, Liu
C, Fang Y and Ding X: Diverse effects of ischemic pretreatments on
the long-term renal damage induced by ischemia-reperfusion. Am J
Nephrol. 30:440–449. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yin C, Salloum FN and Kukreja RC: A novel
role of microRNA in late preconditioning: Upregulation of
endothelial nitric oxide synthase and heat shock protein 70. Circ
Res. 104:572–575. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Dong S, Cheng Y, Yang J, Li J, Liu X, Wang
X, Wang D, Krall TJ, Delphin ES and Zhang C: MicroRNA expression
signature and the role of microRNA-21 in the early phase of acute
myocardial infarction. J Biol Chem. 284:29514–29525. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Weber M, Baker MB, Moore JP and Searles
CD: MiR-21 is induced in endothelial cells by shear stress and
modulates apoptosis and eNOS activity. Biochem Biophys Res Commun.
393:643–648. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sabatel C, Malvaux L, Bovy N, Deroanne C,
Lambert V, Gonzalez ML, Colige A, Rakic JM, Noël A, Martial JA and
Struman I: MicroRNA-21 exhibits antiangiogenic function by
targeting RhoB expression in endothelial cells. PLoS One.
6:e169792011. View Article : Google Scholar : PubMed/NCBI
|