Introduction
As of 2014, 39% of adults worldwide were overweight
and 13% were obese (1). Obesity is
a confirmed risk factor for hyperlipidemia, type-II diabetes,
hypertension and cardiovascular disease (2). Globally, 44% cases of diabetes, 23%
cases of ischemic heart disease and 7–41% cases of certain types of
cancer are attributable to excess weight and obesity (1). Therefore, reducing the rates of being
overweight and obese will have benefits for all. Recently, new
insights have indicated that microRNAs (miRNAs/miRs) serve a role
in obesity and may be suitable therapeutic targets (3). miRNAs are a class of small endogenous
non-coding RNAs that are involved in post-transcriptional gene
repression (4). Epigenetic
regulation of adipose functions mediated by miRNAs has emerged as
an important mechanism in the study of energy metabolism and
obesity in the last decade (5).
Numerous miRNAs have been demonstrated to be associated with
physiopathological disorders associated with obesity (6).
miR-1275, a 17 base pair (bp) intergenic microRNA,
lies on chromosome 6 (7). A
previous study demonstrated that miR-1275 was markedly
down-regulated during maturation of human preadipocytes (8); less expression of miR-1275 was also
observed in obese individuals compared with the non-obese ones
(8). miR-1275 inhibits human
visceral pre-adipocyte differentiation by silencing of ELK1, an
E-twenty-six-domain transcription factor associated with adipocyte
differentiation (2,8). These findings indicated that
down-regulation of miR-1275 is involved in adipogenesis and
obesity. Notably, lower levels of miR-1275 expression have been
reported in some other diseases, such as hepatocellular carcinoma
(9), breast cancer (10), coronary heart disease and some
infections (11). These studies
demonstrated that the down-regulation of miR-1275 is involved not
only in obesity, but also in cancers and infections. However,
little is known about the regulation of miR-1275 transcription.
Further study is needed to determine how miR-1275 expression is
down-regulated in obesity.
In obese individuals, adipose tissue is in a state
of chronic, low-grade inflammation (12). Increased tumor necrosis factor
(TNF)-α and interleukin (IL)-6 levels have been demonstrated to be
notable markers of chronic inflammation in white adipose tissue
(13). High levels of TNF-α and
IL-6 are associated with the regulation of microRNA transcription
in adipocytes (14). For instance,
the expression of miR-146b in adipose tissue is up-regulated by
TNF-α and IL-6 (15). It was
therefore hypothesized that miR-1275 expression in adipose tissue
may be regulated by TNF-α and IL-6. In the present study, a series
of experiments were performed and the results confirmed the above
hypothesis.
Materials and methods
Cell culture and treatment
Human visceral pre-adipocytes (ScienCell Research
Laboratories, San Diego, CA, USA) were maintained in preadipocyte
medium containing 5% fetal bovine serum, 1% preadipocyte growth
supplement, and 1% penicillin/streptomycin solution (PAM, cat. no.
7211; all ScienCell Research Laboratories) at 37°C in an incubator
under 5% CO2 and 95–100% humidity (16). To induce differentiation,
serum-free PAM (containing 50 nM insulin, 100 nM dexamethasone, 0.5
mM 3-isobutyl-1-methylxanthine, and 100 mM rosiglitazone) was added
to confluent human preadipocytes (day 0), and the medium was
replaced every 2 days for the next 4 days. Thereafter, the medium
was replaced with serum-free PAM containing 50 nM insulin and
replaced every 2 days until lipid droplets had accumulated in cells
(day 15) (17). Differentiated
adipocytes were used for experiments 15 days later, when >80% of
cells demonstrated morphological and biochemical properties of
adipocytes. After an overnight incubation in serum-free PAM, human
adipocytes were treated with a final concentration of 10 ng/ml
TNF-α (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) or 30 ng/ml
IL-6 (Sigma-Aldrich; Merck KGaA, Germany) for 0, 4, 8 or 24 h at
37°C in an incubator under 5% CO2 and 95–100%
humidity (18). Human embryonic
kidney 293T (HEK293T) cells were purchased from the American Type
Culture Collection (Manassas, VA, USA) and maintained in Dulbecco's
modified Eagle's medium supplemented with 10% fetal bovine serum
(Bovogen Biologicals, Keilor East, Victoria, Australia). HEK293T
cells were also incubated at 37°C in an incubator under 5%
CO2 and 95–100% humidity (17).
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total miRNA from human adipocytes was extracted
using an miRNeasyMiNi kit (Qiagen GmbH, Hilden, Germany). The
quality of microRNA was assessed using 1% agarose gel
electrophoresis and the concentration of microRNA was measured
using spectrophotometry at 260 nm. Equal quantities of microRNA
(200 ng) were used to synthesize cDNA using TaqMan microRNA Reverse
Transcriptase kit (Applied Biosystems; Thermo Fisher Scientific
Inc., Waltham, MA, USA) according to the manufacturer's protocol
(Table I). qPCR with SYBR (Power
SYBR™ Green PCR Master Mix; cat. no. 4368577; Thermo
Fisher Scientific, Inc.) was carried out using an Applied
Biosystems 7500 Sequence Detection system (ABI 7500 SDS; Thermo
Fisher Scientific, Inc.) following manufacturer's protocol
(Table II). Relative gene
expression was quantified using the 2−ΔΔCq method
(18). Total microRNA expression
was normalized to small nuclear (sn)RNA U6. Primer cat. numbers
are: 001973 for snRU6 and 002840 for miR-1275 (Applied Biosystems;
Thermo Fisher Scientific Inc.).
 | Table I.Temperature protocol for the microRNA
reverse transcription. |
Table I.
Temperature protocol for the microRNA
reverse transcription.
| Step | Temp, °C | Time, sec |
|---|
| Hold | 16 | 30 |
| Hold | 42 | 30 |
| Hold | 85 | 5 |
| Hold | 4 | ∞ |
| Table II.qPCR protocol implemented in the
present study. |
Table II.
qPCR protocol implemented in the
present study.
| Cycling
condition | Temp, °C | Time, sec |
|---|
| Initial
denaturation/hot start | 95 | 30 |
| Repeat steps 1–3
through 40 cycles |
|
|
| Step
1 | 95 | 5 |
| Step
2 | 58 | 10 |
| Step
3 | 72 | 15 |
Bioinformatic analysis
Hsa-miR-1275 precursor (pri-miR-1275) sequence was
obtained from Ensembl (http://www.asia.ensemble.org/index.html). The
precursor sequence was numbered from +1 to +80, and therefore 1,500
bp upstream of pri-miR-1275 was considered to be the predicted
promoter region. To identify the predicted binding sites of NF-κB,
the sequence of the predicted promoter was uploaded to Genomatrix
(http://www.genomatix.de), Jaspar (jaspar.genereg.net), and Promo_v3 (alggen.lsi.upc.es/cgi-bin/promo_v3/promo/promoinit.cgi?dirDB=TF_8.3),
and the intersection of putative binding sites was recorded
(19–21).
Plasmids and promoter reporter
constructs
The predicted promoter region of miR-1275 was
amplified by PCR with TaqDNA polymerase (cat. no. DR100A; Takara
Biotechnology Co., Ltd., Dalian, China). DNA was amplified at 95°C
for 40 sec for denaturation, 55°C for 35 sec for renaturation and
130 sec for elongation with 30 cycles. The site-specific mutation
vectors aimed at the binding site of NF-κB were constructed by
introducing point mutations with overlap-extension PCR. The
predicted binding sites of NF-κB are presented in Table III. PCR products were then cloned
into the dual-luciferase reporter plasmid PEZX-FR01 using
MluI/BamHI restriction sites (GeneCopoeia Inc.,
Rockville, MD, USA). The primer sequences used are listed in
Table IV.
| Table III.NF-κB binding sites predicted by
Genomatrix, Jaspar and Promo_v3. |
Table III.
NF-κB binding sites predicted by
Genomatrix, Jaspar and Promo_v3.
| Sequence | Position from | To | Core similarity on
Genomatrix | Score on
Jaspar | Dissimilarity on
Promo_v3 (%) |
|---|
| agaggTTCCcc | −1,490 | −1,380 |
|
6.27 | 3.01 |
|
atagggccTTCCtcc |
−947 |
−933 | 0.926 |
5.80 |
|
|
ctaggggtTTCCttca |
−806 |
−792 | 0.971 | 11.19 | 3.23 |
|
ctagggccTTCCttc |
−782 |
−768 | 0.926 |
4.35 |
|
|
ttgGGGAggccctgaa |
−288 |
−274 | 0.932 |
4.71 | 9.97 |
| Table IV.Primers used in the present
study. |
Table IV.
Primers used in the present
study.
| Primer | Sequences |
|---|
| Pro-1(−840 to
+1) | F:
5′-CAGAACATTTCTCTACTAGTACGCGTTCTTTGGCCAAAAGGACAAGGCAGGCA |
|
| AAAGTC-3′ |
|
|
R:5′-TTAGCATCGGCCATGGTGGCGGATCCAGTCCTTAGATGGTGATCTTGGTGAAGA |
|
| CCAAAG-3′ |
| Pro-3(−240 to
+1) |
F:5′-CAGAACATTTCTCTACTAGTACGCGTGGAGGGTCAGTTCCACTTCTTCTGTCTCCAC-3′ |
|
|
R:5′-TTAGCATCGGCCATGGTGGCGGATCCAGTCCTTAGATGGTGATCTTGGTGAAGACC-3′ |
| Mut1 |
F:5′-CAGAACATTTCTCTACTAGTACGCGTTCTT-3′ |
|
|
R:5′-TTAGCATCGGCCATGGTGGCGGATCCAGTCCTTAGATGGTGATCTTGGTGAAGACC |
|
| AAAGAC-3′ |
| Mut2 |
F:5′-CAGAACATTTCTCTACTAGTACGCGTTCTTTGGCCAAAAGGACAAGGCAGGCAA |
|
| AAGTCT-3′ |
|
| R:
5′-TTAGCATCGGCCATGGTGGCGGATCCAGTCCTTAGATGGTGATCTTGGTGAAGACC |
|
| AAAGAC-3′ |
| Mut3 | F:
5′-CAGAACATTTCTCTACTAGTACGCGTTCTT-3′ |
|
|
R:5′-TTAGCATCGGCCATGGTGGCGGATCCAGTCCTTAGATGGTGATCTTGGTGAAGACC |
|
| AAAGAC-3′ |
Dual-luciferase assays
HEK293T cells were cultured in 24-well plates at
density of 2×104 cells/well. When grown to a density of
60–70%, they were transfected with 2 µg/well reporter plasmids and
6-µl/well Lipofectamine (Invitrogen; Thermo Fisher Scientific Inc.)
for 6 h. Empty PEZX-FR01 plasmid served as a control. Following
this, cells were washed with serum-free medium and treated with the
NF-κB activator TNF-α (10 ng/ml) for 24 h. For the rescue assay,
HEK293T cells were treated with the NF-κB inhibitor
4-methyl-N1-(3-phenyl-propyl) -benzene-1,2-diamine (10 µM;
Calbiochem; Merck KGaA) for 1 h. Subsequently, cells were washed
with serum-free medium and treated with TNF-α (10 ng/ml) for 24 h.
After this, luciferase activity was analyzed using a dual
luciferase reporter assay (Promega Corporation, Madison, WI, USA)
with a GloMax 96 Microplate Luminometer E6501 (Promega Corporation)
(22). Firefly luciferase activity
was normalized to the Renilla luciferase activity.
Statistical analysis
The data were analyzed using SPSS version 17.0 (SPSS
Inc., Chicago, IL, USA). All data are presented as the mean ±
standard deviation. Statistical analysis was performed using
one-way analysis of variance followed by Dunnett's post hoc test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Expression of miR-1275 in human
adipocytes treated with TNF-α or IL-6
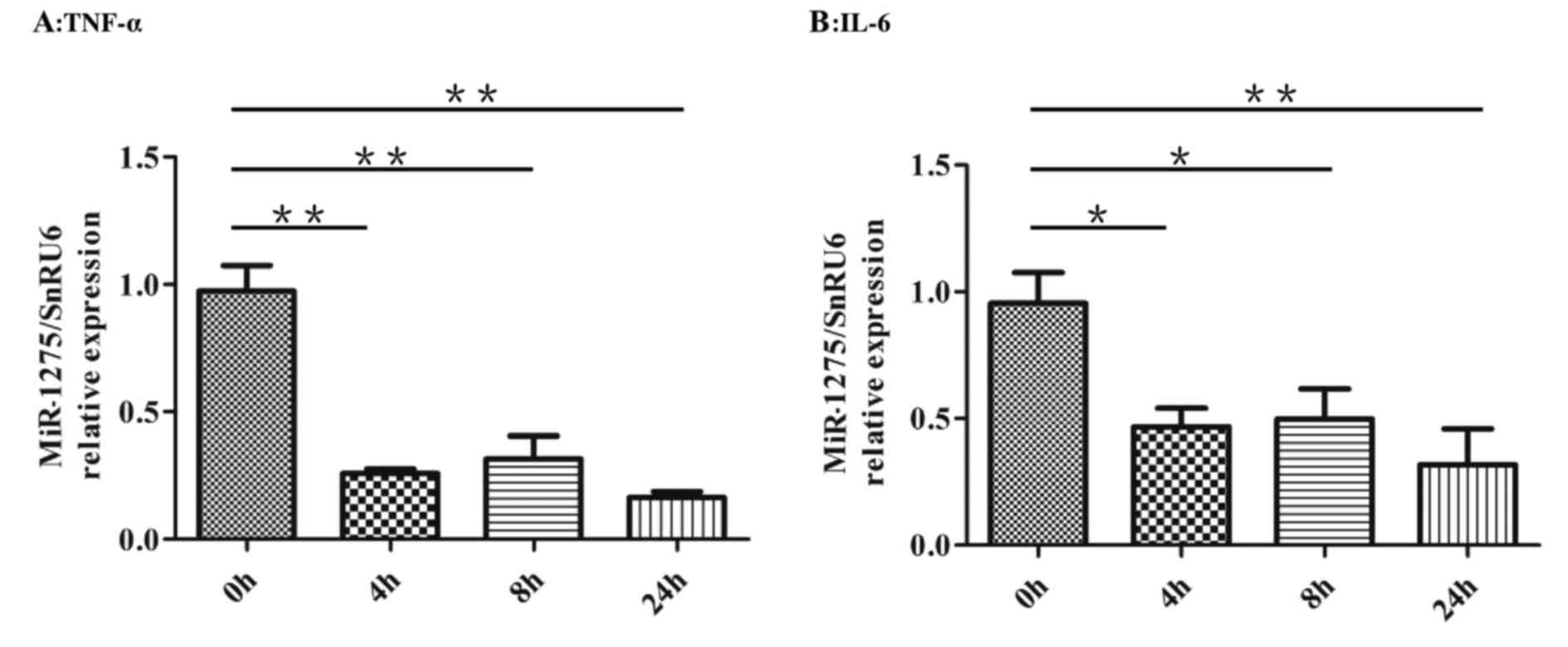
Human mature adipocytes were treated with 10 ng/ml
TNF-α or 30 ng/ml IL-6 for 24 h, and the expression of miR-1275 was
examined at different times (0, 4, 8 and 24 h). The expression of
miR-1275 was normalized to the expression of snRU6. After treatment
with TNF-α, miR-1275 relative expression was significantly lower
than in untreated control cells (Fig.
1A). A similar reduction in miR-1275 expression was also
observed in human adipocytes treated with IL-6 (Fig. 1B). These results indicated that
TNF-α and IL-6 could regulate the expression of miR-1275 in human
adipocytes.
Prediction of promoter regions of
miR-1275
Bioinformatic analysis was performed on the 1,500 bp
upstream sequence of pri-miR-1275 to further determine how TNF-α
and IL-6 regulate miR-1275 expression. Several binding sites of
NF-κB, the downstream transcription factor of TNF-α and IL-6, were
identified on the 1,500 bp sequence of the predicted promoter. This
suggested that TNF-α and IL-6 may regulate miR-1275 expression
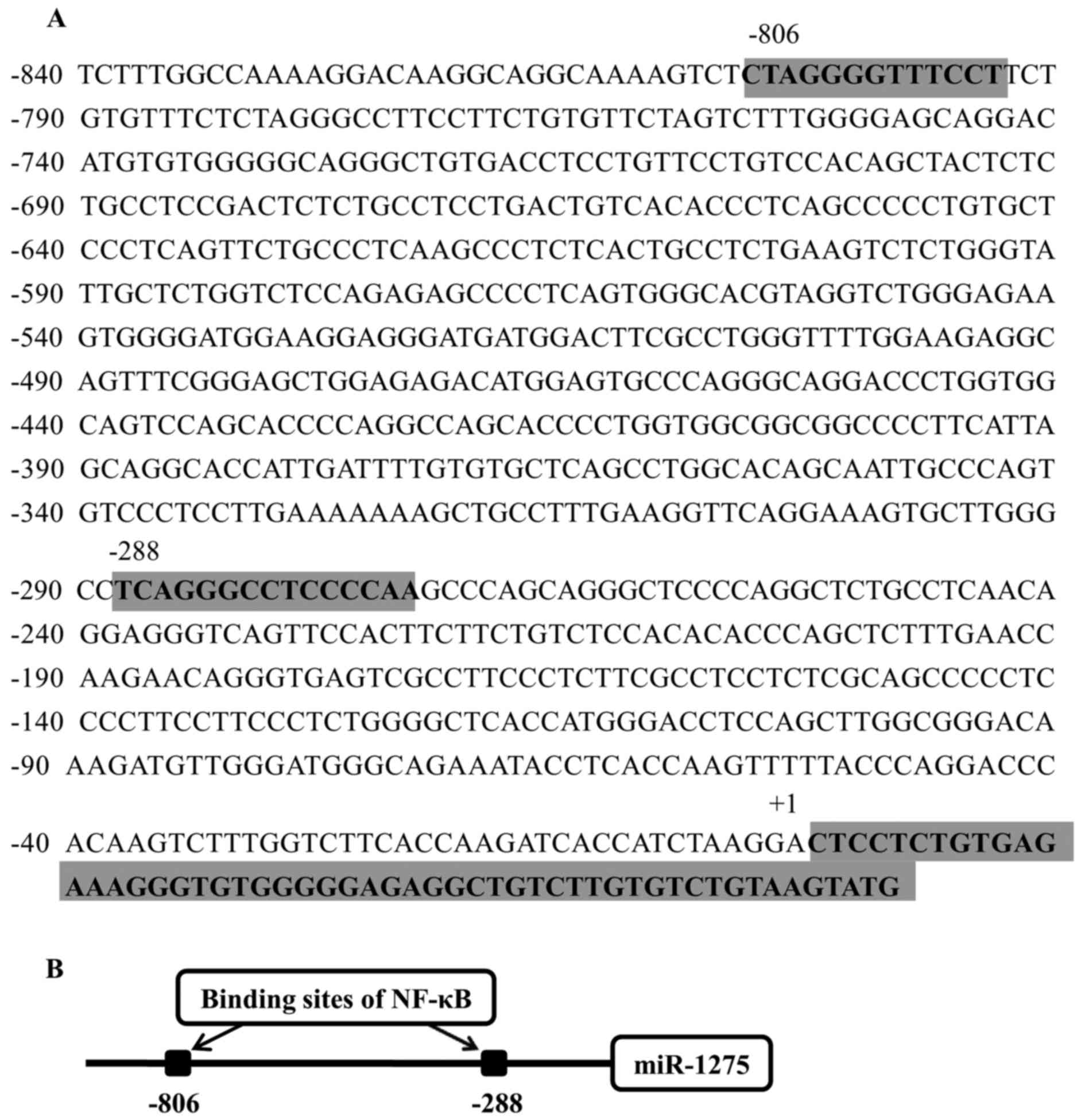
through NF-κB. To locate the strongest binding site of NF-κB, the
sequence of putative promoters was submitted to the following
websites: Genomatrix, Jaspar and Promo_v3. Two sites (from −806 to
−792 and from-288 to-274) were predicted by all three websites
(Table II). Therefore, these were
chosen to be the strongest binding sites of NF-κB (Fig. 2).
Confirmation of NF-κB binding sites on
the promoter region of miR-1275
To determine whether the two predicted binding sites
were functional, different promoters were made and their activity
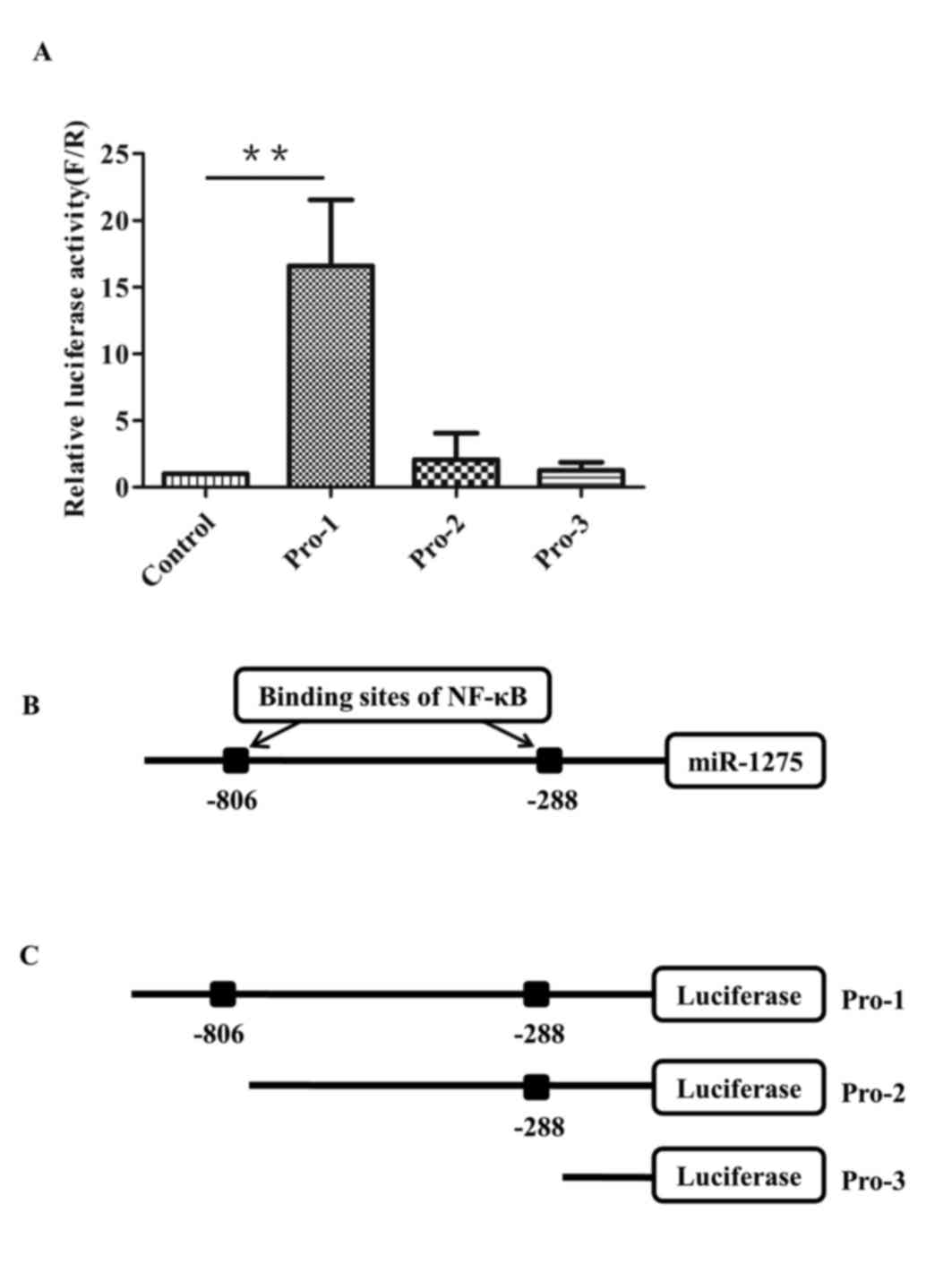
was examined in HEK293T cells. For the deletion assay (Fig. 2A), dual-luciferase reporter
plasmids PEZX-FR01 of Pro-1 (including both sites), Pro-2 (only
including the-288 site) and Pro-3 (including neither site) were
transfected into HEK293T cells. Pro-1 demonstrated >10 times
higher luciferase activity compared with the empty reporter
plasmids, whereas Pro-2 and Pro-3 showed similar luciferase
activity compared with the control (Fig. 3A). These results suggested that the
sequence from −840 to +1 was the promoter region of miR-1275, and
the −806 site was necessary for the promoter's activity (Fig. 3B and C).
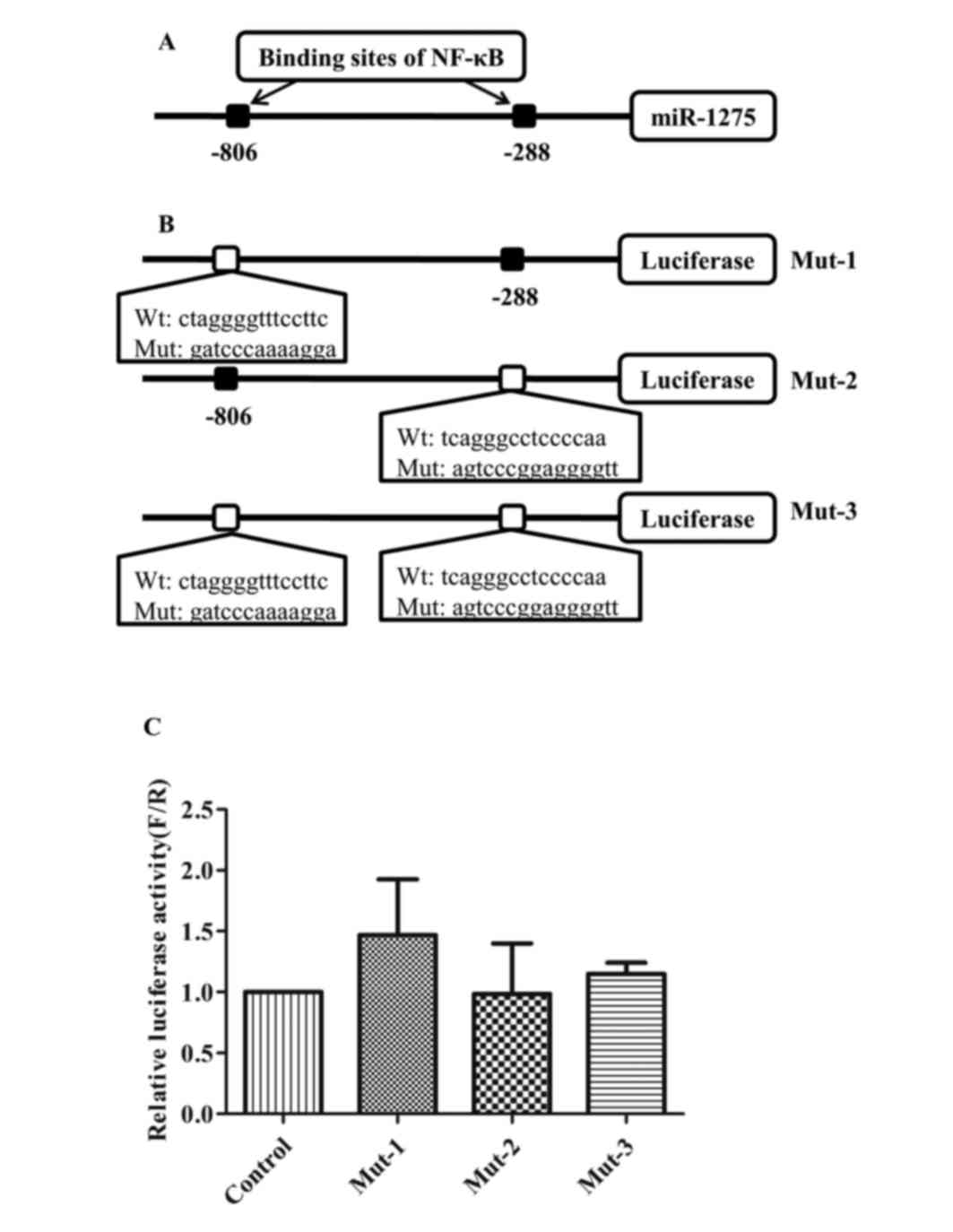
To confirm the role of −288 site, the core binding
sequences of the −806 and −288 sites were mutated (Fig. 4A and B). The luciferase activities
of Mut-1 (mutation on-806 site), Mut-2 (mutation on-288 site) and
Mut-3 (mutation on both-806 and-288 sites) demonstrated no
difference compared with the empty reporter plasmids (Fig. 4C). These results suggested that the
−288 site was also necessary for the promoter activity.
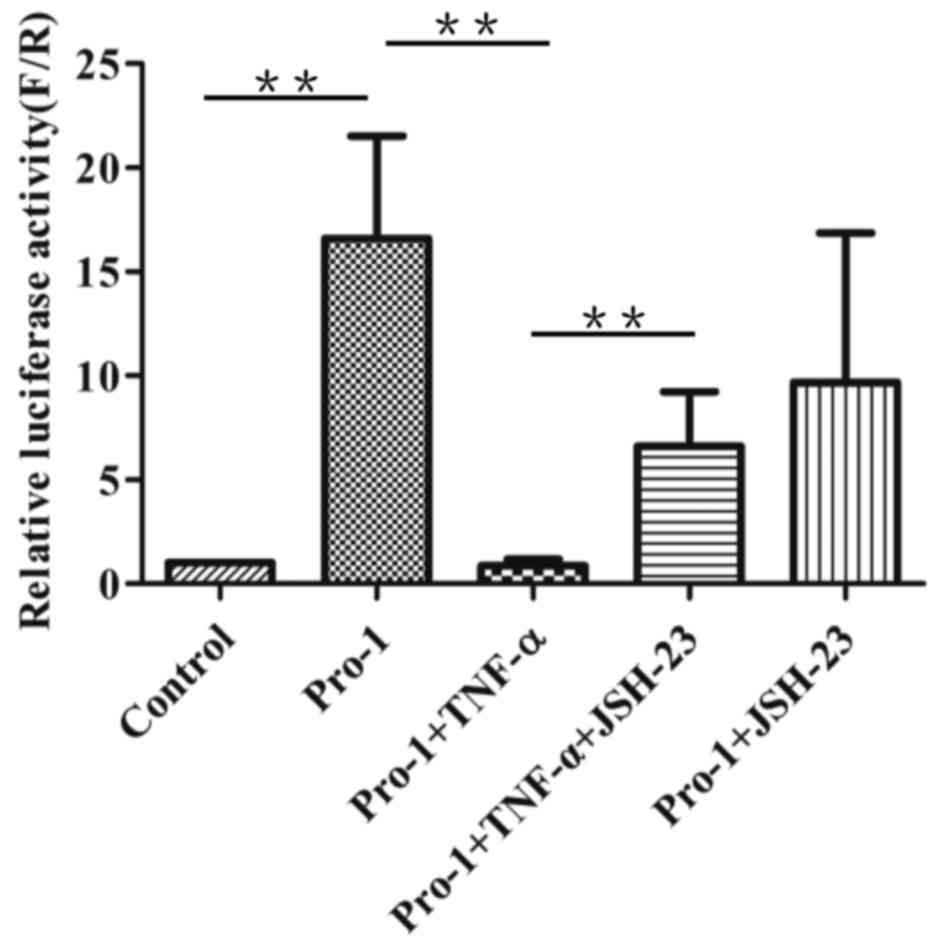
Effects of TNF-α and JSH-23 on
miR-1275 promoter activity
To confirm the above-mentioned results, a rescue
assay was performed. HEK293T cells were transfected by
Pro-1-PEZX-FR01 and then treated with JSH-23 before treatment with
TNF-α. As presented in Fig. 5,
cells treated with JSH-23 and TNF-α demonstrated significantly more
luciferase activity than cells treated with TNF-α alone. These
results confirmed that TNF-α decreased miR-1275 promoter activity
through NF-κB.
Discussion
A previous study has demonstrated that miR-1275 is
markedly down-regulated during differentiation and maturation of
human preadipocytes by miRNA expression microarrays (8). Reduced miR-1275 expression in obese
individuals compared with non-obese individuals has been observed.
Additionally, miR1275 has been reported to be dysregulated in
certain types of cancer and infection (23). Fawzy et al (7) reported that miR-1275 expression is
downregulated cirrhotic and cancerous liver tissue. However, the
exact mechanism is currently unclear.
In the present study, miR-1275 expression was
downregulated by >60% in response to TNF-α and by 50% in
response to IL-6 in mature adipocytes, which suggested that
inflammatory cytokines could serve an important role in regulating
miR-1275 expression in adipocytes. Obesity is characterized by
increased production of cytokines such as IL-6, IL-1, and TNF-α
(24). The role of miRNAs in
inflamed adipose tissues has been highlighted in several studies
(14). Shi et al (17) reported that IL-6 and TNF-α induce
obesity-associated inflammatory responses through transcriptional
regulation of miR-146b. Another study also demonstrated Mir-335 to
be involved in adipose tissue inflammation (25). The present study was consistent
with previous ones. It has confirmed that miR-1275 may be
downregulated by TNF-α and IL-6. The role of TNF-α and IL-6 in
regulating miR-1275, luciferase assays and rescue experiments were
implemented based on bioinformatics analysis.
NF-κB is a downstream transcription factor of TNF-α
and IL-6 (26). TNF-α activates
NF-κB by enhancing nuclear translocation of P65, which is the
active subunit of NF-κB (27,28).
Promoter analysis of the miR-146a gene revealed that NF-κB serves a
critical role in inducing its transcription by LPS, TNF-α, and IL-1
(29). miR-335 is also proved to
be up-regulated by TNF-α and IL-6 through the NF-κB signaling
pathway in adipocytes (25).
Previous studies have also demonstrated that let-7 and miR-365 are
under the control of NF-κB (30,31).
To determine whether NF-κB is involved in regulating miR-1275, a
1,500 bp upstream sequence of miR-1275 was uploaded to Jaspar, a
database that uses a transcription factor flexible models to
predict transcription factors of a given sequence (19). Notably, 23 sites were predicted as
binding sites of NF-κB. To confirm the results, the 1,500 bp
upstream sequence of miR-1275 was submitted to Genomatix and
Promo_v3 (20,21), and the intersection of binding
sites predicted by the three databases was taken (Table II. The region from −806 to-792
(here called site-806) and the region from-288 to-274 (here called
site-288) were predicted by all three databases.
A total of six different sequences, termed Pro-1,
Pro-2, Pro-3, Mut-1, Mut-2 and Mut-3, were designed to assess the
promoter activity of sites-806 and-288. Pro-1 (which has both
sites) demonstrated >10 times higher luciferase activity
compared with empty reporter plasmids, whereas Pro-2 and Pro-3
demonstrated equal luciferase activity compared with the control.
Luciferase activities of Mut-1 (mutation at site −806), Mut-2
(mutation at site −288) and Mut-3 (mutations at both sites)
demonstrated no difference compared with the empty reporter
plasmids. These results illustrated that both sites are necessary
for promoter activity. These data also indicated that NF-κB was
involved in the transcription of miR-1275 by binding to sites-806
and-288. The next step was to prove whether NF-κB participates in
the regulation of miR-1275 by TNF-α.
HEK293T cells were transfected with Pro-1-PEZX-FR01
and were then treated with JSH-23 prior to TNF-α. TNF-α
dramatically decreased promoter activity of Pro-1, while cells
treated with JSH-23 and TNF-α demonstrated significantly higher
luciferase activity compared with cells treated with TNF-α alone.
This meant that JSH-23 could partially reverse the down-regulation
of the promoter activity of Pro-1 caused by TNF-α. JSH-23 is an
NF-κB inhibitor and acts by inhibiting NF-κB/P65 nuclear
translocation (32,33). Notably, a potential mechanism could
be the following: In response to TNF-α, with the nuclear
translocation of NF-Κb/P65, NF-κB is activated via NF-Κb/P65
nuclear translocation which then binds to sites-806 and-288 of the
miR-1275 promoter region. This leads to inhibiting miR-1275
transcription. Other studies have illustrated a similar association
between NF-κB activation and miRNA downregulation. Sasaki and
Vageli (34) have reported lower
levels of miR-34a, −451a and-375 in response to NF-κB activation;
miR-1908 was also down-regulated when NF-κB was bound to its
promoter region (22).
In conclusion, the present results demonstrated that
miR-1275 expression in mature human adipocytes can be
down-regulated by TNF-α and IL-6. NF-κB was proven to be involved
in the regulation of miR-1275 transcription by binding to its
promoter region. In response to TNF-α, NF-κB was bound to promoter
region of miR-1275 and inhibited its transcription. This may
partially explain the down-regulation of miR-1275 in mature human
adipocytes. More studies are required to explore whether miR-1275
could be a therapeutic target for obesity.
Acknowledgements
The present study was supported by the Nanjing
Medical University (grant no. 2013NJMU0027), the Education
Department of Jiangsu Province (grant nos. 13KJB20014 and
JX10617801), and the National Natural Science Foundation of China
(grant no. 81300706). The authors would like to thank LetPub
(http://www.letpub.com) for its linguistic
assistance during the preparation of this manuscript.
References
|
1
|
World Health Organization, . Obesity and
Overweight Fact Sheet. June. 2016http://www.who.int/mediacentre/factsheets/fs311/en/3–August.
2016
|
|
2
|
Sandfort V, Lai S, Ahlman MA, Mallek M,
Liu S, Sibley CT, Turkbey EB, Lima JA and Bluemke DA: Obesity is
associated with progression of atherosclerosis during statin
treatment. J Am Heart Assoc. 5(pii): e0036212016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Peng Y, Yu S, Li H, Xiang H, Peng J and
Jiang S: MicroRNAs: Emerging roles in adipogenesis and obesity.
Cell Signal. 26:1888–1896. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Abente EJ, Subramanian M, Ramachandran V
and Najafi-Shoushtari SH: MicroRNAs in obesity-associated
disorders. Arch Biochem Biophys. 589:108–119. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yang L, Shi CM, Chen L, Pang LX, Xu GF, Gu
N, Zhu LJ, Guo XR, Ni YH and Ji CB: The biological effects of
hsa-miR-1908 in human adipocytes. Mol Biol Rep. 42:927–935. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fawzy IO, Hamza MT, Hosny KA, Esmat G, El
Tayebi HM and Abdelaziz AI: miR-1275: A single microRNA that
targets the three IGF2-mRNA-binding proteins hindering tumor growth
in hepatocellular carcinoma. FEBS Lett. 589:2257–2265. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pang L, You L, Ji C, Shi C, Chen L, Yang
L, Huang F, Zhou Y, Zhang J, Chen X and Guo X: MiR-1275 inhibits
adipogenesis via ELK1 and its expression decreases in obese
subjects. J Mol Endocrinol. 57:33–43. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Das K, Saikolappan S and Dhandayuthapani
S: Differential expression of miRNAs by macrophages infected with
virulent and avirulent Mycobacterium tuberculosis. Tuberculosis
(Edinb). 93 Suppl:S47–S50. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Katsushima K, Shinjo K, Natsume A, Ohka F,
Fujii M, Osada H, Sekido Y and Kondo Y: Contribution of
microRNA-1275 to Claudin11 protein suppression via a
polycomb-mediated silencing mechanism in human glioma stem-like
cells. J Biol Chem. 287:27396–27406. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Akamatsu S, Hayes CN, Tsuge M, Miki D,
Akiyama R, Abe H, Ochi H, Hiraga N, Imamura M, Takahashi S, et al:
Differences in serum microRNA profiles in hepatitis B and C virus
infection. J Infect. 70:273–287. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dandona P, Aljada A and Bandyopadhyay A:
Inflammation: The link between insulin resistance, obesity and
diabetes. Trends Immunol. 25:4–7. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hotamisligil GS, Arner P, Caro JF,
Atkinson RL and Spiegelman BM: Increased adipose tissue expression
of tumor necrosis factor-alpha in human obesity and insulin
resistance. J Clin Invest. 95:2409–2415. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Karkeni E, Astier J, Tourniaire F, El Abed
M, Romier B, Gouranton E, Wan L, Borel P, Salles J, Walrand S, et
al: Obesity-associated inflammation induces microRNA-155 expression
in adipocytes and adipose tissue: Outcome on adipocyte function. J
Clin Endocrinol Metab. 101:1615–1626. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen L, Dai YM, Ji CB, Yang L, Shi CM, Xu
GF, Pang LX, Huang FY, Zhang CM and Guo XR: MiR-146b is a regulator
of human visceral preadipocyte proliferation and differentiation
and its expression is altered in human obesity. Mol Cell
Endocrinol. 393:65–74. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xu LL, Shi CM, Xu GF, Chen L, Zhu LL, Zhu
L, Guo XR, Xu MY and Ji CB: TNF-alpha, IL-6 and leptin increase the
expression of miR-378, an adipogenesis-related microRNA in human
adipocytes. Cell Biochem Biophys. 70:771–776. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shi C, Zhu L, Chen X, Gu N, Chen L, Zhu L,
Yang L, Pang L, Guo X, Ji C and Zhang C: IL-6 and TNF-α induced
obesity-related inflammatory response through transcriptional
regulation of miR-146b. J Interferon Cytokine Res. 34:342–348.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mathelier A, Fornes O, Arenillas DJ, Chen
CY, Denay G, Lee J, Shi W, Shyr C, Tan G, Worsley-Hunt R, et al:
JASPAR 2016: A major expansion and update of the open-access
database of transcription factor binding profiles. Nucleic Acids
Res. 44:D110–D115. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kaur S, Anjali G, Bhardwaj P, Taneja J and
Singh R: Data in support of FSH induction of IRS-2 in human
granulosa cells: Mapping the transcription factor binding sites in
human IRS-2 promoter. Data Brief. 6:162–167. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Farré D, Roset R, Huerta M, Adsuara JE,
Roselló L, Albà MM and Messeguer X: Identification of patterns in
biological sequences at the ALGGEN server: PROMO and MALGEN.
Nucleic Acids Res. 31:3651–3653. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kuang Q, Li J, You L, Shi C, Ji C, Guo X,
Xu M and Ni Y: Identification and characterization of NF-kappaB
binding sites in human miR-1908 promoter. Biomed Pharmacother.
74:158–163. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Parafioriti A, Bason C, Armiraglio E,
Calciano L, Daolio PA, Berardocco M, Di Bernardo A, Colosimo A,
Luksch R and Berardi AC: Ewing's sarcoma: An analysis of miRNA
expression profiles and target genes in paraffin-embedded primary
tumor tissue. Int J Mol Sci. 17(pii): E6562016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hotamisligil GS: Inflammation and
metabolic disorders. Nature. 444:860–867. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhu L, Chen L, Shi CM, Xu GF, Xu LL, Zhu
LL, Guo XR, Ni Y, Cui Y and Ji C: MiR-335, an adipogenesis-related
microRNA, is involved in adipose tissue inflammation. Cell Biochem
Biophys. 68:283–290. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ozes ON, Mayo LD, Gustin JA, Pfeffer SR,
Pfeffer LM and Donner DB: NF-kappa B activation by tumour necrosis
factor requires the Akt serine-threonine kinase. Nature. 401:82–85.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang D and Baldwin AS Jr: Activation of
nuclear factor-kappaB-dependent transcription by tumor necrosis
factor-alpha is mediated through phosphorylation of RelA/p65 on
serine 529. J Biol Chem. 273:29411–29416. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen LF and Greene WC: Shaping the nuclear
action of NF-kappa B. Nat Rev Mol Cell Bio. 5:392–401. 2004.
View Article : Google Scholar
|
|
29
|
Taganov KD, Boldin MP, Chang KJ and
Baltimore D: NF-kappaB-dependent induction of microRNA miR-146, an
inhibitor targeted to signaling proteins of innate immune
responses. Proc Natl Acad Sci USA. 103:12481–12486. 2006;
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Iliopoulos D, Hirsch HA and Struhl K: An
epigenetic switch involving NF-kappaB, Lin28, Let-7 MicroRNA and
IL6 links inflammation to cell transformation. Cell. 139:693–706.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xu Z, Xiao SB, Xu P, Xie Q, Cao L, Wang D,
Luo R, Zhong Y, Chen HC and Fang LR: miR-365, a novel negative
regulator of interleukin-6 gene expression, is cooperatively
regulated by Sp1 and NF-kappaB. J Biol Chem. 286:21401–21412. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Borghi A, Verstrepen L and Beyaert R:
TRAF2 multitasking in TNF receptor-induced signaling to NF-κB, MAP
kinases and cell death. Biochem Pharmacol. 116:1–10. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Vranková S, Barta A, Klimentová J,
Dovinová I, Líšková S, Dobešová Z, Pecháňová O, Kuneš J and Zicha
J: The regulatory role of nuclear factor Kappa B in the heart of
hereditary hypertriglyceridemic rat. Oxid Med Cell Longev.
2016:98140382016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sasaki CT and Vageli DP: miR-21, miR-155,
miR-192, and miR-375 deregulations related to NF-kappaB activation
in gastroduodenal fluid-induced early preneoplastic lesions of
laryngeal mucosa in vivo. Neoplasia. 18:329–338. 2016. View Article : Google Scholar : PubMed/NCBI
|