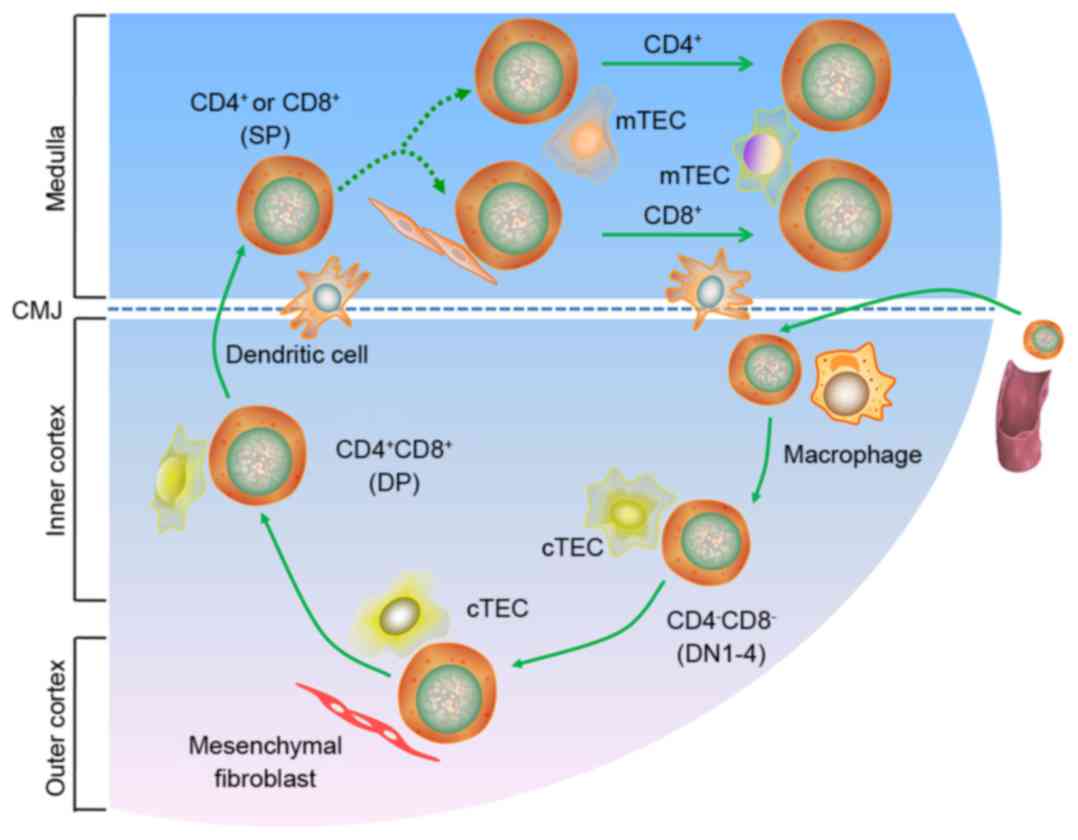
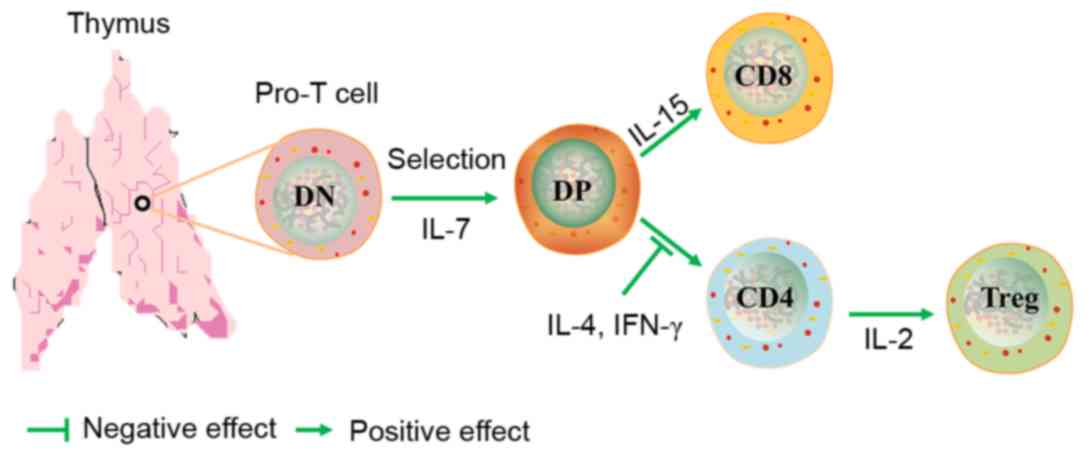
Introduction
The thymus is a bilobed organ located in the
superior mediastinum of the thorax, above the heart and behind the
sternum. It can be divided into two main subcompartments: The
cortex and the medulla. Each subcompartment contains numerous
subtypes of thymic epithelial cells (TECs), in addition to
dendritic cells, mesenchymal cells and endothelial cells (1–3). In
addition, the thymus establishes and maintains thymic
microenvironments, which are capable of supporting the efficient
development of T cells. The maintenance of these microenvironments
is dependent upon the specialized functions of thymic stromal
cells, and other major components of the thymic microenvironment
(4,5). During the development and maturation
of thymocytes from bone marrow-derived T cell progenitors, three
main events serve a critical role in each T cell bearing a unique T
cell receptor (TCR): The rearrangement and expression of TCRα and β
loci, which depends on their somatic assembly; positive selection
[the identification of cells that are able to recognize self-major
histocompatibility complex (MHC) in antigen presentation to T
cells]; and negative selection (the elimination of T cells that are
potentially autoreactive). T cells that survive the selection
processes eventually become mature cluster of differentiation
(CD)4+ or CD8+ single positive T cells
(Fig. 1). These processes ensure a
population of non-autoreactive peripheral T cells. T cell migration
is directed by several mediators, including chemokine receptors and
G protein-coupled receptors (GPCR), which are supported by guiding
stromal structures and by TECs, including cortical TECs and
medullary TECs (mTEC). The TECs form a three-dimensionally oriented
network, rather than the more ‘typical’ two-dimensional (2D)
epithelial structures (6,7). It is important to determine the
molecular mechanisms underlying the thymic regulation of T cell
development and of the proteins involved in T cell recognition.
However, to the best of our knowledge, the mechanisms underlying
these processes have not yet been fully explored.
Molecular mechanisms underlying regulatory T
cell generation in the thymus
In 1969, Nishizuka and Sakakura were the first to
present a mechanism for the generation of regulatory T
(Treg) cells in the thymus, based on a neonatal
thymectomy experiment (8).
Treg cells in the thymus are vital for the negative
regulation of immune-mediated inflammation, which features
prominently in autoimmune and autoinflammatory disorders, acute
allergies, cancer, chronic infections and commensal microbiota.
They are also important for the regulation of metabolic
inflammation for homeostasis and peripheral tolerance (9–11).
Recent studies have demonstrated that mice lacking the forkhead box
P3 (Foxp3) transcription factor experience overwhelming autoimmune
pathology, which they succumb to in a matter of weeks (12,13).
Although CD25 is not a specific marker expressed exclusively on
Treg cells, using specific anti-CD25 antibodies for the
depletion or inactivation of Treg cells, in combination
with immunostimulation, is an attractive treatment modality,
particularly in anti-tumour immunotherapy (14). The current understanding is that
Treg cell development occurs when the TCR avidity for
self-antigens lies between the TCR avidities that drive positive
and negative selection (15–19).
TCR engagement is also known to stimulate various downstream
signaling molecules and transcription factors. This stimulation
leads to an intricate web of downstream intracellular signaling
events. Proteins important in thymic Treg cell function
include phosphoinositide 3-kinase, protein kinase B (AKT),
mammalian target of rapamycin (mTOR), nuclear factor of activated T
cells, transcription factor activator protein 1 and nuclear
factor-κB (NF-κB). Numerous pathways contribute to Treg
cell development, including the TCR, AKT-mTOR and NF-κB pathways,
among others (20–22). Various types of antigen-presenting
cells (APCs) capture and present antigens to thymocytes through a
complex network of signaling pathways (Fig. 2). In addition, calcium signaling
appears to be involved in thymic Treg cell development
(23). Furthermore, increased
generation of the Foxp3 protein in developing thymic
Treg cells may have a positive role in Ca2+
signaling (24,25). However, calcium is also a powerful
negative regulator of Foxp3 in the AKT-mTOR pathway.
Phosphatidylinositol-4,5-bisphosphate 3-kinase/AKT signaling
regulates the phosphorylation and inhibition of forkhead box O
(FoxO) transcription factors. The FoxO transcription factors have
recently been reported to facilitate the expression of Foxp3 and
Treg cell development (26–28).
Although natural Treg (nTreg) and induced
Treg (iTreg) cells can enforce tolerance,
iTreg cells, such as those derived from commensal
bacteria in the gut, may have a particularly important role as they
increase antigen receptor diversity (29,30).
The mechanisms underlying the development and antigen specificities
of nTreg and iTreg cells are likely to
differ.
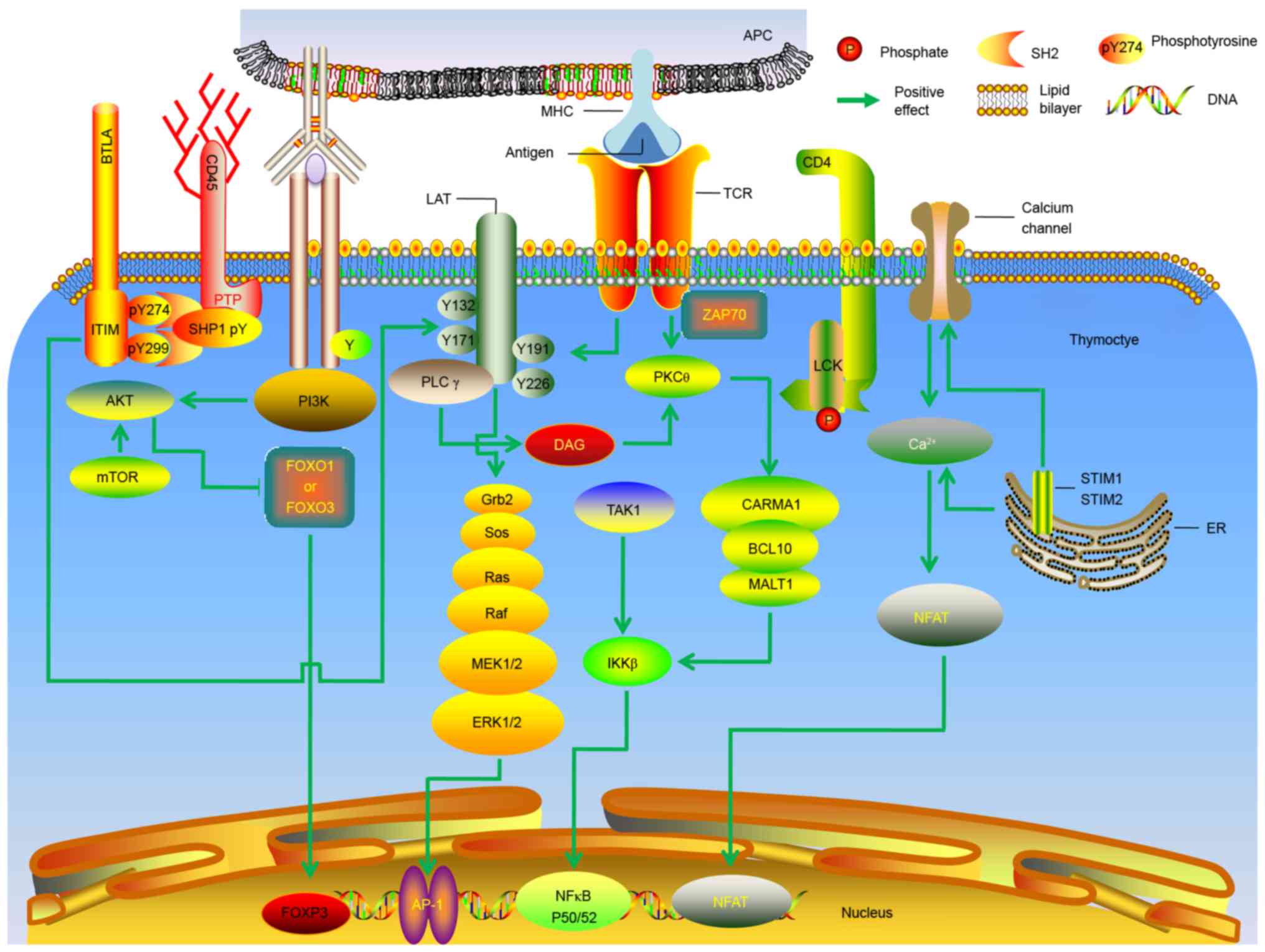 | Figure 2.Molecular mechanisms underlying the
generation of thymic regulatory T cells. Molecular signals
downstream of the TCR are presented. AP, activator protein; APC,
antigen-presenting cell; BCL, B cell lymphoma; BTLA, B and T
lymphocyte attenuator; Ca, calcium; CARMA, CARD-containing MAGUK
protein; CD, cluster of differentiation; DAG, diacylglycerol; ER,
endoplasmic reticulum; ERK, extracellular signal-regulated kinase;
IKKβ, inhibitor of nuclear factor κB; ITIM, immunoreceptor
tyrosine-based inhibition motif; MEK, mitogen-activated
extracellular signal-regulated kinase; MHC, major
histocompatibility complex; FoxO, forkhead box protein O; FOXP3,
forkhead box protein 3; NFAT, nuclear factor of activated T; Grb,
growth factor receptor-bound protein; LAT, linker for activation of
T cells; LCK, lymphocyte-specific protein tyrosine kinase p56;
MALT, mucosa-associated lymphoid tissue lymphoma translocation
protein; mTOR, mechanistic target of rapamycin; NF, nuclear factor;
PI3K, phosphatidylinositol-4,5-bisphosphate 3-kinase; PK, protein
kinase; PL, phospholipase; PTP, protein-tyrosine phosphatase; Ras,
rat sarcoma also known as p21; Raf, rapidly accelerated
fibrosarcoma; SHP, SH2-containing protein tyrosine phosphatase;
SOS, Son of Sevenless; STIM, stromal interaction molecule; TAK,
transforming growth factor beta-activated kinase; ZAP70,
ζ-associated protein of 70 kD. |
Over the past few years, substantial progress has
been made in understanding the developmental process of thymic
Treg cells and the molecular mechanism underlying their
regulation in the thymus. However, there remain numerous unanswered
questions. For example, the molecular differences between immature
CD4+ single positive (SP) thymocytes in the thymus and
naive peripheral T cells remain unknown. In addition, it remains to
be elucidated why Foxp3 expression occurs predominantly in
CD4+, and not CD8+, SP thymocytes, The
present review aimed to understand these molecular mechanisms and
how these molecular components are ‘wired’ into regulatory
signaling and transcriptional networks. Achieving this may aid in
the improvement of therapeutic strategies used to treat autoimmune
and inflammatory disorders.
Effects of cytokines on thymic function
Cytokines serve as molecular messengers between
immune cells, and have been reported to be of major importance to
thymic function. The effects of cytokine cascades on thymic
function are generally well understood. Almost all types of thymic
cells can produce cytokines, either spontaneously or following
stimulation with stimulating agents, including lipopolysaccharides,
phytohemagglutinin and ionomycin. The most important of the thymic
cell subsets are TECs, which are the principal source of cytokines
and chemokines required in early T cell development (31). The differential expression of major
cytokines produced by TECs can be divided into four branches:
Hemopoietins, proinflammatory cytokines, suppressor cytokines and
interleukin (IL)-6 and IL-7 cytokines (32,33).
Notably, cytokines and other growth factors serve important roles
in thymic function, regulating various cellular processes. However,
the functions of numerous cytokines in the thymus are not well
understood. Understanding the effects of intrathymic cytokines may
reveal some unknown aspects of thymic physiology.
The thymus produces hormones and cytokines that
regulate immune function. A previous study identified at least six
types of thymic cells (34). The
histological features of the thymus are broadly divided into the
central medulla and a peripheral cortex. Previous research has
demonstrated that cytokine secretion by T lymphocytes has a vital
role in mounting adaptive immune responses (35). In addition, the large number of
cytokines produced by the thymus maintains a fine balance between
thymocyte proliferation, maturation, activation, differentiation
and survival inhibition. Thymic cells also secrete the peptides
IL-1, IL-3, IL-4 and IL-6, and three major thymic hormones,
thymosins, thymopoietin and thymulin (36–39).
Thymic hormones serve a major role in preserving the functions of
the immune system, and cytokines have essential roles in the
control of immune responses.
Cytokines are small polypeptides that regulate cell
function and are predominantly secreted by immune cells. Numerous
cytokines responsible for the modulation of T cell differentiation
are produced by thymocytes and TECs. The ability of thymocytes to
produce cytokines is important in the regulation of thymic cytokine
production and the responses to their action (Fig. 3). Of these regulators, IL-7 serves
a particular role in thymocyte differentiation; IL-7 has been
reported to promote the rearrangement of TCR genes by enhancing the
production and activity of recombinases (40,41).
The thymic production of Treg cells requires IL-2, which
is also required during T cell development in the thymus and for
the maturation of Treg cells. Recent studies have
reported that IL-2 receptor is functionally active within the
thymus; it increases the number of CD4+Foxp3+
thymocytes and the expression of Foxp3 and CD25 to normal levels
(42–44). IL-4 is another cytokine produced by
T cells whose receptor contains a γ(c)-chain. It has previously
been demonstrated that IL-4 is synergistic with IL-2 in the
induction of thymocyte proliferation in fetal thymic organ culture.
In addition, IL-4 supports thymocytes through successive phases of
proliferation, acting alongside stimulatory agents (45,46).
Recently, research has been directed at the cytokine IL-10, which
is produced by Treg cells, and other chronically
stimulated T helper cells, B cells and APCs. IL-10 is important for
maintaining immune homeostasis at mucosal surfaces and also
contributes to immune suppression (47–49).
Interferon (IFN)-γ has numerous effects on TECs; it
activates TECs and increases surface expression of MHC classes I
and II, and other membrane proteins (50). Furthermore, IFN-γ stimulates the
secretion of IL-6 by TECs (51).
IFN-γ also supports thymocyte differentiation, through its action
on TEC functions. Tumor necrosis factor (TNF)-α has been reported
to have an important role in the regulation of thymocyte
production, inducing apoptosis and the proliferation of immature
CD3−CD4−CD8− T cells in the
presence of IL-7 (52).
Furthermore, TNF-α and IL-1 participate as cofactors in the
induction of CD4−CD8− thymocyte commitment
and differentiation (53). TNF-α
also stimulates the production of IL-6 and enhances the apoptosis
of CD4+CD8+ cells induced by glucocorticoids
(54,55).
Some molecules are multifunctional and serve
different functions in the cytokine system within the thymus than
they do in peripheral compartments of the immune system. For
example, some cytokines are pleiotropic in their biological
activities and exhibit different roles in these different systems.
The principal roles of thymic cytokines are in constitutive
processes, including thymocyte migration and development, and the
mediation of cell populations, but not inducible ones, such as
immune response/tolerance or inflammation, as in the periphery. The
synthesis of cytokines and the expression of their receptors in the
thymus is usually spontaneous, or is induced by cell-cell
interactions, unlike in the periphery. Information regarding the
production of cytokines in the thymus and the biological activity
of these cytokines is summarized in Table I.
 | Table I.Biological activity of cytokines
affects T cell-associated thymic function. |
Table I.
Biological activity of cytokines
affects T cell-associated thymic function.
| Author (year) | Name | Molecular weight
(kDa) | Cell producers | Biological
activity | (Refs.) |
|---|
| Coto et al
(1992); Dalloul et al (1991) | IFN-γ | 17.1 | Activated T cells;
Natural killer cells | Affects T-cell,
B-cell, and macrophage differentiation and maturation | (36,37) |
| Nitta and Suzuki
(2016); Galy et al (1990); Savino et al (2016) | IL-1 | 17.3; 17.5 | Thymic epithelial
cells; Macrophages; Monocytes | Can act as a growth
factor for thymocytes and promote thymocyte T cell activation,
proliferation and differentiation; Members of the IL-1 family of
receptors contain activators and suppressors of inflammation | (31,38,39) |
| Savino and Dardenne
(2000); Muegge et al (1993) | IL-2 | 13–17 | Activated T
cells | Promotes the
development of Treg cells within the inner thymus;
Promotes the activation of T cell proliferation, differentiation
and cytokine productio | (40,41) |
| Bayer et al
(2007); Varas et al (1997); Weist et al (2015);
Meilin et al (1997) | IL-4 | 18–19 | Activated T
cells | T cell growth
factor | (42–45) |
| Zlotnik et
al (1987); Shevach (2009); Barnes and Powrie (2009) | IL-6 | 19–28 | Monocytes;
Macrophages; Fibroblasts; T cells; Endothelial cells; B cells | Promotes the
development and maturation of thymocytes | (46–48) |
| Mittal and Roche
(2009); Patel et al (1995) | IL-7 | 20–28 | Stromal cells;
Keratinocytes Hepatocytes; Dendritic cells | Promotes
differentiation of CD8+ T cells in the thymus; Maintains
T cell proliferation | (49,50) |
| Meilin et al
(1995); Baseta and Stutman (2000) | IL-9 | 14–25 | Activated T
cells | T cell growth
factor | (51,52) |
| Zúñiga-Pflücker
et al (1995); Arzt et al (2000) | IL-12 | 75 | T cells | Maintains thymic
integrity and function | (53,54) |
| Cohen-Kaminsky
et al (1991); Wang et al (2016) | IL-17 | 34–52 | T lymphocytes | Activates CD4+
memory T lymphocytes; Produces Treg 17 cells | (55,56) |
| Shanley et
al (2009); Dooley and Liston (2012); Kappler et al
(1987) | IL-21 | 15–18 | Activated CD4 T
cells | Treg 17
cells Promotes CD4 T cell differentiation; Reduces the Th17
pathway; Costimulates activated natural killer and CD8 lymphocytes;
Desensitizes responding cells to the inhibitory effects of;
Treg cells act as a switch for immunoglobulin G
production in B cells | (57–59) |
| Xing and Hogquist
(2012); Roberts et al (1990) | IL-22 | 17–22 | T helper 17
cells | Promotes thymic
epithelial cells proliferation and survival; Affects T cell
development | (60,61) |
| Kisielow et
al (1988); Ramsdell and Fowlkes (1990); Howard et al
(1999) | TGF-β | 12.5 | Activated T cells;
Activated B cells | Inhibits the IL-1,
IL-2 and IL-7-dependent proliferation of thymocytes | (62,63,67) |
| Wang et al
(1994); Müller-Hermelink et al (1987); Gruver and Sempowski
(2008) | TNF-α | 17–26 | Monocytes;
Macrophages | Promotes T cells
and B cell proliferation | (68–70) |
| Boyd (1932); Gruver
et al (2007) | TSLP | 18.1 | Epithelial and
dendritic cells | Promotes T helper 2
differentiation of naïve CD4 T cells; Activates natural killer T
cells, basophils and other innate immune cells | (71,72) |
Regulation of molecular mechanisms in
stress-mediated thymic atrophy and involution
Stress is able to disrupt homeostasis of the immune
system, and various stressful conditions cause acute thymic
involution, including emotional distress, malnutrition and
pregnancy (56,57). Furthermore, numerous processes can
trigger thymic involution during pathological conditions, such as
bacterial and viral infections, inflammation, disease, clinical
cancer treatment and preparative regimens for bone marrow
transplants (58), as presented in
Fig. 4. Therefore, mechanisms must
exist to regulate these processes in various contexts. It is well
known that the thymus serves an important role in the body's immune
response. It provides the microenvironment essential for the
development of T cells from hematopoietic stem cells. The central
functions of the thymus are critical to immune tolerance in several
rodent and large animal models under normal or pathological
conditions. These functions act through various mechanisms, such as
clonal deletion or clonal anergy of self-reactive T cells,
elimination or control of self-reactive T cells, and anergy of
self-reactive T cells (59–63).
Recent mechanistic studies regarding central and peripheral T cell
tolerance have assisted in the design of novel, immunomodulating
therapeutic strategies for the treatment of autoimmune diseases,
and improve the prevention, detection and treatment of cancer and
associated diseases, as well as exert immunoregulatory effects in
transplantation outcomes using pharmacological or biological
interventions (64–66). Immunosenescence and immune atrophy,
which are associated with reduced immunity, are complex processes
that have yet to be fully understood. Numerous factors exert a
negative effect on thymopoiesis, acute stress-induced thymic
atrophy and on chronic thymic involution associated with aging.
These factors include starvation, environmental stressors,
bacterial infection, and irradiation or immunosuppressive therapies
(67–70).
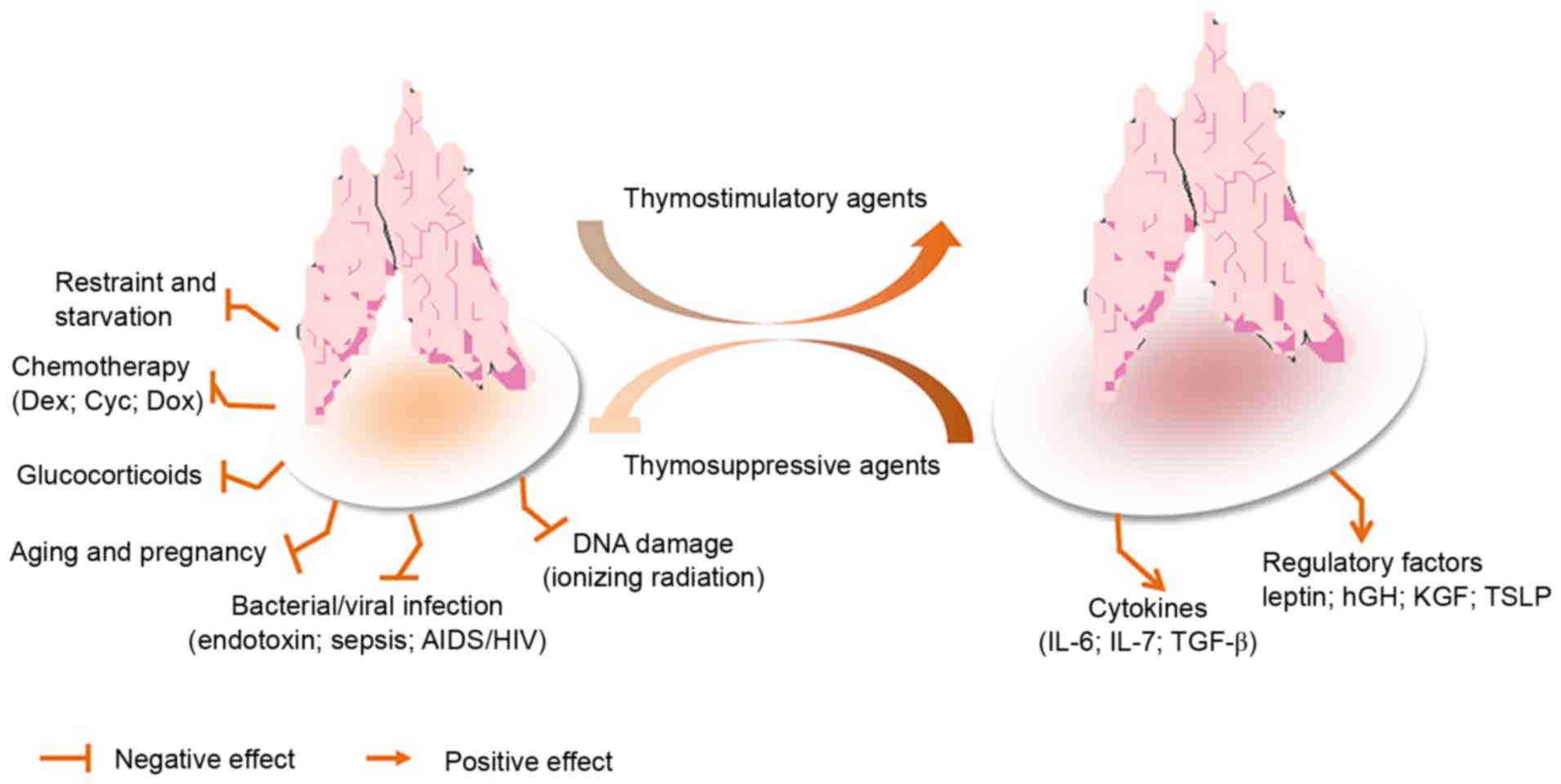 | Figure 4.Model of stress-induced thymic
atrophy, and thymosuppressive and thymostimulatory mediators. AIDS,
acquired immunodeficiency syndrome; Cyc, cyclophosphamide; Dex,
dexamethasone; Dox, doxorubicin; HIV, human immunodeficiency virus;
hGH, human growth hormone; IL, interleukin; KGF, keratinocyte
growth factor; TGF-β, transforming growth factor-β; TSLP, thymic
stromal lymphopoietin. |
The shrinkage of the thymus was reported >80
years ago by Boyd (71); however,
the underlying mechanisms are not well understood. Immunosenescence
is defined as deterioration in the immune system, which is
associated with aging (72–74),
and has attracted increasing interest in the scientific and
health-care sectors alike. Thymic atrophy has often been observed
due to the direct or indirect influences of drugs or the
environment on the thymus. However, one other major consideration
in thymic atrophy is a systemic rise in glucocorticoids and
inflammatory cytokines. Unfortunately, the thymus is acutely
sensitive to various stresses and injuries; therefore, it is often
considered as a ‘barometer of stress’ for the body. Prolonged
thymic atrophy in stress situations can contribute to peripheral T
cell deficiency or can inhibit immune reconstitution, thus
resulting in a decrease in thymopoiesis (75,76).
Therefore, mechanistic studies have increasingly focused on thymic
atrophy. A commonly used mouse model of endotoxemia-induced acute
thymic atrophy has been used to reveal the effects of acute stress
on thymopoiesis. For example, in a lipopolysaccharide (LPS)-induced
acute thymic atrophy model, microarray analysis revealed >11,000
probe sets with significant alterations (>1.4-fold), 1 day after
an LPS challenge. This finding has important implications regarding
how the direct intrathymic response to an endotoxin challenge
contributes to thymic involution during endotoxemia (77). In endotoxin-stressed mice, it has
previously been reported that leptin administration augments
thymopoiesis in LPS-treated leptin-deficient (ob/ob) mice,
but not in normal mice (78).
Furthermore, a recent study indicated that the number of thymocytes
and TECs was significantly decreased in LPS-treated neonatal thymic
involution (79).
Age-associated thymic involution must also be
considered. Aging is accompanied by a decline in the function and
development of the immune system. Understanding the aging process,
and how that process can be delayed or reversed, may allow us to
take action to adopt healthier lifestyles and live longer.
Age-associated thymic involution is characterized by progressive
diminution of novel T cell production (80). However, many previous findings are
contradictory. Some studies have reported the effects of aging on
the function of neutrophils, macrophages and natural killer cells,
whereas other studies have reported no association (81,82).
In addition, some studies have demonstrated that the systemic
administration of keratinocyte growth factor (KGF) enhances T cell
lymphopoiesis by stimulating TECs to secrete various cytokines that
then act on developing thymocytes in young and old mice (83,84).
Furthermore, a previous study was conducted on C57BL/6×DBA/2
recombinant inbred strains of mice to identify the genetic loci
influencing age-associared thymic involution, and demonstrated that
the strongest quantitative trait loci influencing the rate of
thymic involution in the recombinant-inbred mice were mapped to
chromosome (Chr) 9 (D9Mit20 at 62 cM) and Chr 10 (D10Mit61 at 32
cM) (85).
It is well known that stress on the immune system
leads to the suppression of immune cell functions, such as in T
cells, macrophages, dendritic cells and B cells, and the atrophy of
immune organs, predominantly the thymus and spleen. The thymus is
one of the central organs of the immune system, and is essential
for the development of the adaptive immune system. Insult,
infection, dysregulation of positive and negative selection,
suppression of cell adhesion, chemotaxis, cytotoxicity, increased
apoptosis or antigen presentation in the thymus, may all lead to
autoimmunity or immunosuppression (86,87).
Previous studies have suggested that exposure to immunosuppressive
agents, such as diethylstilbestrol, dexamethasone (DEX),
azathioprine, cyclophosphamide (Cyc),
2,3,7,8-tetrachlorodibenzo-p-dioxin or cyclosporin A may
induce immunotoxic effects resulting in hypocellularity, apoptosis
and atrophy in the thymus (88–92).
This provides evidence regarding the molecular mechanisms and
cellular targets involved in thymic atrophy-induced
immunosuppression. DEX is a synthetic glucocorticoid compound with
potent anti-inflammatory activity, which is associated with
clinically significant side effects that severely limit its
therapeutic use. In a previous study, DEX (20 mg/kg) was
administered to C57Bl/6 mice via intraperitoneal injection; the
thymuses were then harvested 5 days after treatment. Analysis of
the thymic tissues detected a depletion of
CD4+CD8+ double positive thymocytes, and
upregulation of IL-22 and IL-23 in wild-type mice (93). In another study, the
immunosuppressant cyclosporin A was reported to induce extensive
reductions in the autoimmune regulator tolerance-inducing MHC class
IIhigh mTECs (mTEChigh). The most distinctive
effects of Cyc and DEX exposure were extensive reductions in
thymocytes and stromal cells, and, as with cyclosporin A, severely
depleted tolerance-inducing mTEChigh (91).
Prediction of potential drug targets on the
thymus using proteomics
The thymus remains still largely uncharted territory
that invites further investigation. Understanding the role of the
thymus in T cell generation and homeostasis, and understanding
exactly how such systems work and what proteins are involved has
resulted in greater interest in thymus organogenesis. The
application of systems biology, combined with more traditional
methods, is essential to uncover and optimize the molecular
mechanisms underlying effects (drug-induced or otherwise) on the
thymus. These methods will allow the study of novel aspects of
thymic function and aid understanding regarding thymic function,
morphogenesis and development. This knowledge may then be used to
identify potential drug targets. In addition, these methods will
prove useful not only for studying gene and protein function in
thymus organogenesis, but also for clarifying the origin and
lineage relationship between cortical and medullary epithelial cell
types. Recently, modern approaches to chemical genomics,
metabolomics, genomics, transcriptomics, pharmacogenomics,
microbiomics and proteomics have proved to be useful in the
identification and characterization of molecular mechanisms
underlying all aspects of pharmacological sciences and
physiological processes, and in other areas (94,95).
Therefore, evidence suggests that proteomics may be effectively
used in the in-depth study of the thymus in different models and
pathological conditions.
Proteomics is the large-scale study of proteins, and
facilitates the systematic analysis of protein molecules in
complicated biological systems. Turiák et al (96) focused on the proteomic
characterization of thymocyte-derived microvesicles (MVs) and
apoptotic bodies in BALB/c mice; 195 and 142 proteins were
identified in MVs and apoptotic bodies, respectively. This previous
study also identified numerous molecules known to serve important
roles in the immune system, such as MHCI, MHCII, CD5 and CD97 in
MVs, and CD45 in both types of vesicles. Similarly, Billing et
al (97) used proteomic
profiling analysis to measure the non-genomic and concomitant
genomic effects of acute restraint stress on rat thymocytes. In
recent years, several methods have been developed for relative and
absolute quantitative proteomics. The most widely used quantitative
techniques include gel-based [2D gel electrophoresis, difference
gel electrophoresis (DIGE)] and liquid chromatography-mass
spectrometry (MS)-based methods (isotope-coded affinity tag, stable
isotope labeling with amino acids in cell culture, isobaric tags
for relative and absolute quantitation). MS-based proteomics
methods are typically divided into two categories: Label-free or
label-based approaches (98).
Proteomics research is applied to a wide range of biological
systems for the study of differentially expressed proteins,
particularly candidates for biomarker discovery and validation,
understanding disease processes and clinical proteomics (99). Notably, in a previous study
quantitative 2D-DIGE with matrix-assisted laser
desorption/ionization-time of flight (TOF)/TOF MS was used to
identify 108 proteins with differential subcellular localizations
in rat thymocytes; this may be the first study to determine the
rapid effects of stress-induced hypothalamus-pituitary-adrenal
activation at the proteome level in vivo (97). According to our current
understanding, doxorubicin (DOX) treatment leads to degeneration of
the thymus. Proteomics analysis is consistent with the notion that
DOX treatment in vivo leads to thymic senescence (100). Cyc has also been reported to
induce immunosuppression and thymic atrophy. Proteomic analysis
indicated that possible target-related processing was instigated
following Cyc-treatment in mice (101). Apoptosis serves an essential role
in the development and maturation of T lymphocytes during mammalian
thymus maturation. Experiments have indicated that several proteins
were differentially regulated in the cytosol of T cell precursors
by a signal from TCR, as identified using proteomic techniques
(102). Proteomics has been
widely used to study the experimentally induced acute phase
reaction, and to study numerous disease models associated with
cancer and inflammatory diseases (103,104). A previous study revealed the
cellular and molecular mechanisms using proteomic approaches
combined with bioinformatics analysis (105). Despite the increased use of
proteomics, knowledge of protein interactions and pathway networks
remains largely incomplete; however, data generated by quantitative
proteomics can still provide valuable insights (106).
Final remarks
More than 50 years ago, Miller (107) conducted seminal studies on the
immunological function of the thymus using neonatally thymectomized
mice. The importance of this primary lymphoid organ was quickly
established, as the thymus provides a unique microenvironment in
which T cells or T lymphocytes undergo development, differentiation
and clonal expansion during the physiological development of the
immune system. In recent years, there has been a marked interest in
the association between the immune system and the thymus,
generating results that confirmed that the thymus was endowed with
an immune function. The immune system has evolved to mount an
effective defense against pathogens and to minimize deleterious
immune-mediated inflammation caused by commensal microorganisms,
immune responses against self and environmental antigens, and
metabolic inflammatory disorders. It appears that Treg
cell-mediated suppression serves as a vital mechanism in the
negative regulation of immune-mediated inflammation, and features
prominently in autoimmune and autoinflammatory disorders, and
pathologies induced by fungi, parasites, allergies, acute and
chronic infections, cancer and metabolic inflammation.
Treg cells are considered important to researchers in
their efforts to increase the efficacy of vaccines for cancer,
acquired immune deficiency syndrome and autoimmune diseases. The
discovery that Foxp3 is the transcription factor that specifies the
Treg cell lineage has facilitated recent progress in
understanding the biology of Treg cells. These findings
may provide novel targets for subsequent drug development.
There is an increasingly in-depth understanding of
cytokines and their activities in biological pathways. Therefore,
an improved understanding regarding the cytokine network is
essential to determine the role of numerous key cytokines, and to
modulate thymic function. Cytokines, such as ILs, may be useful in
improving the functionality of the thymus and may be used to treat
immunodeficiency or autoimmune diseases. It has been reported that
cytokines, including IL-6, IL-7 receptor, IL-10 and IL-22, serve a
key regulatory role in T cell growth and differentiation processes
in the thymus. These cytokines may be mediated through various
regulatory mechanisms and signaling pathways to establish a
protective effect on the thymus. Understanding these pathways will
increase the understanding of the regulatory mechanism of the
thymus and the biology of Treg cells and secreted
cytokine function. Previous studies have analyzed the effects of
cytokine therapy as a complementary schedule to conventional
therapy with γ-globulin (108–110). The results suggested that the
treatment has a long-term positive effect on the immune response,
relative to other therapeutic interventions. A combination of
IFN-α2b, thymic factors, γ-globulin and granulocyte-macrophage
colony-stimulating factor may be a promising to treat common
variable immunodeficiency. In addition, a previous study reported
that cytokines not only serve an essential role during early T cell
development, but are also responsible for the development of other
thymic cells, such as thymic dendritic cells, generated from
precursors produced in bone marrow (32). At present, information on this
topic is limited. An essential difference between cytokine
production inside the thymus and in peripheral organs is the
different levels of dependence on cell activation, and possibly
cross talk, depending on the cytokine environment and
situation.
Studying protective mechanisms may provide novel
directions in research and the development of drugs for the
treatment of various stresses to the thymus, including
immunosenescence, immune atrophy and immunosuppression. There have
been reports of several small molecules having a protective effect
on the thymus, including leptin, KGF and IL-22. Studies have also
explored the molecular mechanisms involved, predominantly using
mice (70,111). In various chemical stress and
thymic atrophy models, these active molecules can enhance the
remodeling of the thymus, protecting the thymus from some
stressors, such as those involved in aging, as well as hunger,
radiation, hormones and immunosuppressants. Notably, researchers
have made great progress in examining the numerous mechanisms that
contribute to immune suppression and have provided a future
direction for research and a novel manner of developing
immune-modulating drugs (112).
It must be noted that there are differences between
immunosenescence, immune atrophy and immunosuppression; therefore,
these situations should be treated differently when developing
specific molecular signaling pathways and in targeted drug
development. The development of novel drugs, and signal
transduction research concerning these mechanisms, may benefit
patients that are immunocompromised, in a pathological state, or a
combination, to reduce the side effects of other drugs on the
thymus.
During the last decade, the development of
proteomics technology and protein targets for drug generation and
drug screening mechanisms has provided novel tools for biomedical
research (113,114). There have been several reports
regarding thymic molecular mechanisms using proteomics technology;
therefore, a more comprehensive analysis of protein alterations in
the thymus under various circumstances has been established. This,
combined with the related molecule-function databases, including
UniProt, the Kyoto Encyclopedia for Genes and Genomes and the Gene
Ontology database, has enabled protein network data analysis to
screen for known or predicted drug-protein or protein-protein
interactions in the thymus. A greater understanding of the mutual
regulation of protein molecules may allow the prediction of
possible molecular drug targets and drug development pathways.
Existing proteomics studies have provided some pathways for protein
regulation of signal transduction. These pathways are intricate
webs of downstream intracellular signaling events that ultimately
result in specific thymic immune response stresses. This
understanding may provide novel ways of treating immunological
diseases by targeting the stress protein molecules in the thymus,
and may be useful in improving the functionality of the thymus.
Collectively, these studies suggest that the markedly complex
action mechanisms underlying immunomodulatory effects in the thymus
are a promising therapeutic target for treatment of the immune
system.
Acknowledgements
This review was supported by the Young Scientists
Fund of the National Natural Science Foundation of China (grant no.
81403395), the Traditional Chinese Medicine Bureau of Guangdong
Province [grant no. (2014) 539] and the Specific Research Fund for
TCM Science and Technology of Guangdong Provincial Hospital of
Chinese Medicine (grant nos. YN2015QN09, YN2016QJ11 and
YN2015QN12).
References
|
1
|
Gordon J and Manley NR: Mechanisms of
thymus organogenesis and morphogenesis. Development. 138:3865–3878.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Blackburn CC and Manley NR: Developing a
new paradigm for thymus organogenesis. Nat Rev Immunol. 4:278–289.
2004. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Skogberg G, Lundberg V, Berglund M,
Gudmundsdottir J, Telemo E, Lindgren S and Ekwall O: Human thymic
epithelial primary cells produce exosomes carrying
tissue-restricted antigens. Immunol Cell Biol. 93:727–734. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Anderson G and Jenkinson EJ: Lymphostromal
interactions in thymic development and function. Nat Rev Immunol.
1:31–40. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Su M, Hu R, Jin J, Yan Y, Song Y, Sullivan
R and Lai L: Efficient in vitro generation of functional thymic
epithelial progenitors from human embryonic stem cells. Sci Rep.
5:98822015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fan Y, Tajima A, Goh SK, Geng X,
Gualtierotti G, Grupillo M, Coppola A, Bertera S, Rudert WA,
Banerjee I, et al: Bioengineering thymus organoids to restore
thymic function and induce donor-specific immune tolerance to
allografts. Mol Ther. 23:1262–1277. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
van Ewijk W, Wang B, Hollander G, Kawamoto
H, Spanopoulou E, Itoi M, Amagai T, Jiang YF, Germeraad WT, Chen WF
and Katsura Y: Thymic microenvironments, 3-D versus 2-D? Semin
Immunol. 11:57–64. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nishizuka Y and Sakakura T: Thymus and
reproduction: Sex-linked dysgenesia of the gonad after neonatal
thymectomy in mice. Science. 166:753–755. 1969. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Josefowicz SZ, Lu LF and Rudensky AY:
Regulatory T cells: Mechanisms of differentiation and function.
Annu Rev Immunol. 30:531–564. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hsieh CS, Lee HM and Lio CW: Selection of
regulatory T cells in the thymus. Nat Rev Immunol. 12:157–167.
2012.PubMed/NCBI
|
|
11
|
Wang YM, Ghali J, Zhang GY, Hu M, Wang Y,
Sawyer A, Zhou JJ, Hapudeniya DA, Wang Y, Cao Q, et al: Development
and function of Foxp3(+) regulatory T cells. Nephrology (Carlton).
21:81–85. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lahl K, Loddenkemper C, Drouin C, Freyer
J, Arnason J, Eberl G, Hamann A, Wagner H, Huehn J and Sparwasser
T: Selective depletion of Foxp3+ regulatory T cells induces a
scurfy-like disease. J Exp Med. 204:57–63. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kim JM, Rasmussen JP and Rudensky AY:
Regulatory T cells prevent catastrophic autoimmunity throughout the
lifespan of mice. Nat Immunol. 8:191–197. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Rosalia RA, Štěpánek I, Polláková V,
Šímová J, Bieblová J, Indrová M, Moravcová S, Přibylová H, Bontkes
HJ, Bubeník J, et al: Administration of anti-CD25 mAb leads to
impaired α-galactosylceramide-mediated induction of IFN-γ
production in a murine model. Immunobiology. 218:851–859. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wong J, Obst R, Correia-Neves M, Losyev G,
Mathis D and Benoist C: Adaptation of TCR repertoires to
self-peptides in regulatory and nonregulatory CD4+ T cells. J
Immunol. 178:7032–7041. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pacholczyk R, Ignatowicz H, Kraj P and
Ignatowicz L: Origin and T cell receptor diversity of
Foxp3+CD4+CD25+ T cells. Immunity. 25:249–259. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hsieh CS, Liang Y, Tyznik AJ, Self SG,
Liggitt D and Rudensky AY: Recognition of the peripheral self by
naturally arising CD25+ CD4+ T cell receptors. Immunity.
21:267–277. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Maloy KJ and Powrie F: Regulatory T cells
in the control of immune pathology. Nat Immunol. 2:816–822. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Klein L, Kyewski B, Allen PM and Hogquist
KA: Positive and negative selection of the T cell repertoire: What
thymocytes see (and don't see). Nat Rev Immunol. 14:377–391. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chapman NM and Chi H: mTOR links
environmental signals to T cell fate decisions. Front Immunol.
5:6862015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Akimzhanov AM and Boehning D: IP3R
function in cells of the immune system. WIREs Membr Transp Signal.
1:329–339. 2012. View
Article : Google Scholar
|
|
22
|
Sauer S, Bruno L, Hertweck A, Finlay D,
Leleu M, Spivakov M, Knight ZA, Cobb BS, Cantrell D, O'Connor E, et
al: T cell receptor signaling controls Foxp3 expression via PI3K,
Akt, and mTOR. Proc Natl Acad Sci USA. 105:7797–7802. 2008;
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Schwarz A, Schumacher M, Pfaff D,
Schumacher K, Jarius S, Balint B, Wiendl H, Haas J and Wildemann B:
Fine-tuning of regulatory T cell function: The role of calcium
signals and naive regulatory T cells for regulatory T cell
deficiency in multiple sclerosis. J Immunol. 190:4965–4970. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lin J, Yang L, Silva HM, Trzeciak A, Choi
Y, Schwab SR, Dustin ML and Lafaille JJ: Increased generation of
Foxp3(+) regulatory T cells by manipulating antigen presentation in
the thymus. Nat Commun. 7:105622016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Engel M, Sidwell T, Vasanthakumar A,
Grigoriadis G and Banerjee A: Thymic regulatory T cell development:
Role of signalling pathways and transcription factors. Clin Dev
Immunol. 2013:6175952013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ouyang W, Beckett O, Ma Q, Paik Jh,
DePinho RA and Li MO: Foxo proteins cooperatively control the
differentiation of Foxp3+ regulatory T cells. Nat Immunol.
11:618–627. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kerdiles YM, Stone EL, Beisner DL,
McGargill MA, Ch'en IL, Stockmann C, Katayama CD and Hedrick SM:
Foxo transcription factors control regulatory T cell development
and function. Immunity. 33:890–904. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Harada Y, Harada Y, Elly C, Ying G, Paik
JH, DePinho RA and Liu YC: Transcription factors Foxo3a and Foxo1
couple the E3 ligase Cbl-b to the induction of Foxp3 expression in
induced regulatory T cells. J Exp Med. 207:1381–1391. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Haribhai D, Williams JB, Jia S, Nickerson
D, Schmitt EG, Edwards B, Ziegelbauer J, Yassai M, Li SH, Relland
LM, et al: A requisite role for induced regulatory T cells in
tolerance based on expanding antigen receptor diversity. Immunity.
35:109–122. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Omenetti S and Pizarro TT: The Treg/Th17
axis: A dynamic balance regulated by the gut microbiome. Front
Immunol. 6:6392015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Nitta T and Suzuki H: Thymic stromal cell
subsets for T cell development. Cell Mol Life Sci. 73:1021–1037.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yarilin AA and Belyakov IM: Cytokines in
the thymus: Production and biological effects. Curr Med Chem.
11:447–464. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Shitara S, Hara T, Liang B, Wagatsuma K,
Zuklys S, Holländer GA, Nakase H, Chiba T, Tani-ichi S and Ikuta K:
IL-7 produced by thymic epithelial cells plays a major role in the
development of thymocytes and TCRγδ+ intraepithelial lymphocytes. J
Immunol. 190:6173–6179. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tian T, Zhang J, Gao L, Qian XP and Chen
WF: Heterogeneity within medullary-type
TCRalphabeta(+)CD3(+)CD4(−)CD8(+) thymocytes in normal mouse
thymus. Int Immunol. 13:313–320. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chemin K, Bohineust A, Dogniaux S, Tourret
M, Guégan S, Miro F and Hivroz C: Cytokine secretion by CD4+ T
cells at the immunological synapse requires Cdc42-dependent local
actin remodeling but not microtubule organizing center polarity. J
Immunol. 189:2159–2168. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Coto JA, Hadden EM, Sauro M, Zorn N and
Hadden JW: Interleukin 1 regulates secretion of zinc-thymulin by
human thymic epithelial cells and its action on T-lymphocyte
proliferation and nuclear protein kinase C. Proc Natl Acad Sci USA.
89:7752–7756. 1992; View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Dalloul A, Arock M, Fourcade C, Hatzfeld
A, Bertho JM, Debré P and Mossalayi MD: Human thymic epithelial
cells produce interleukin-3. Blood. 77:69–74. 1991.PubMed/NCBI
|
|
38
|
Galy AH, Dinarello CA, Kupper TS, Kameda A
and Hadden JW: Effects of cytokines on human thymic epithelial
cells in culture. II. Recombinant IL 1 stimulates thymic epithelial
cells to produce IL6 and GM-CSF. Cell Immunol. 129:161–175. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Savino W, Mendes-da-Cruz DA, Lepletier A
and Dardenne M: Hormonal control of T-cell development in health
and disease. Nat Rev Endocrinol. 12:77–89. 2016.PubMed/NCBI
|
|
40
|
Savino W and Dardenne M: Neuroendocrine
control of thymus physiology. Endocr Rev. 21:412–443. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Muegge K, Vila MP and Durum SK:
Interleukin-7: A cofactor for V(D)J rearrangement of the T cell
receptor beta gene. Science. 261:93–95. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Bayer AL, Yu A and Malek TR: Function of
the IL-2R for thymic and peripheral CD4+CD25+ Foxp3+ T regulatory
cells. J Immunol. 178:4062–4071. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Varas A, Vicente A, Romo T and Zapata AG:
Role of IL-2 in rat fetal thymocyte development. Int Immunol.
9:1589–1599. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Weist BM, Kurd N, Boussier J, Chan SW and
Robey EA: Thymic regulatory T cell niche size is dictated by
limiting IL-2 from antigen-bearing dendritic cells and feedback
competition. Nat Immunol. 16:635–641. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Meilin A, Sharabi Y and Shoham J: Analysis
of thymic stromal cell subpopulations grown in vitro on
extracellular matrix in defined medium-v. Proliferation regulating
activities in supernatants of human thymic epithelial cell
cultures. Int J Immunopharmacol. 19:39–47. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zlotnik A, Ransom J, Frank G, Fischer M
and Howard M: Interleukin 4 is a growth factor for activated
thymocytes: Possible role in T-cell ontogeny. Proc Natl Acad Sci
USA. 84:3856–3860. 1987; View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Shevach EM: Mechanisms of Foxp3+ T
regulatory cell-mediated suppression. Immunity. 30:636–645. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Barnes MJ and Powrie F: Regulatory T cells
reinforce intestinal homeostasis. Immunity. 31:401–411. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Mittal SK and Roche PA: Suppression of
antigen presentation by IL-10. Curr Opin Immunol. 34:22–27. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Patel DD, Whichard LP, Radcliff G, Denning
SM and Haynes BF: Characterization of human thymic epithelial cell
surface antigens: phenotypic similarity of thymic epithelial cells
to epidermal keratinocytes. J Clin Immunol. 15:80–92. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Meilin A, Shoham J, Schreiber L and
Sharabi Y: The role of thymocytes in regulating thymic epithelial
cell growth and function. Scand J Immunol. 42:185–190. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Baseta JG and Stutman O: TNF regulates
thymocyte production by apoptosis and proliferation of the triple
negative (CD3-CD4-CD8-) subset. J Immunol. 165:5621–5630. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Zúñiga-Pflücker JC, Jiang D and Lenardo
MJ: Requirement for TNF-alpha and IL-1 alpha in fetal thymocyte
commitment and differentiation. Science. 268:1906–1909. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Arzt E, Kovalovsky D, Igaz LM, Costas M,
Plazas P, Refojo D, Páez-Pereda M, Reul JM, Stalla G and Holsboer
F: Functional cross-talk among cytokines, T-cell receptor, and
glucocorticoid receptor transcriptional activity and action. Ann NY
Acad Sci. 917:672–677. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Cohen-Kaminsky S, Delattre RM, Devergne O,
Rouet P, Gimond D, Berrih-Aknin S and Galanaud P: Synergistic
induction of interleukin-6 production and gene expression in human
thymic epithelial cells by LPS and cytokines. Cell Immunol.
138:79–93. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Wang J, Zhuo Y, Yin L, Wang H, Jiang Y,
Liu X, Zhang M, Du F, Xia S and Shao Q: Doxycycline protects thymic
epithelial cells from mitomycin C-mediated apoptosis in vitro via
Trx2-NF-κB-Bcl-2/Bax axis. Cell Physiol Biochem. 38:449–460. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Shanley DP, Aw D, Manley NR and Palmer DB:
An evolutionary perspective on the mechanisms of immunosenescence.
Trends Immunol. 30:374–381. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Dooley J and Liston A: Molecular control
over thymic involution: From cytokines and microRNA to aging and
adipose tissue. Eur J Immunol. 42:1073–1079. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Kappler JW, Roehm N and Marrack P: T cell
tolerance by clonal elimination in the thymus. Cell. 49:273–280.
1987. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Xing Y and Hogquist KA: T-Cell tolerance:
Central and peripheral. Cold Spring Harb Perspect Biol. 4(pii):
a0069572012.PubMed/NCBI
|
|
61
|
Roberts JL, Sharrow SO and Singer A:
Clonal deletion and clonal anergy in the thymus induced by cellular
elements with different radiation sensitivities. J Exp Med.
171:935–940. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Kisielow P, Bluthmann H, Staerz UD,
Steinmetz M and von Boehmer H: Tolerance in T-cell-receptor
transgenic mice involves deletion of nonmature CD4+8+ thymocytes.
Nature. 333:742–746. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Ramsdell F and Fowlkes B: Clonal deletion
versus clonal anergy: The role of the thymus in inducing self
tolerance. Science. 248:1342–1348. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Nurieva R, Wang J and Sahoo A: T-cell
tolerance in cancer. Immunotherapy. 5:513–531. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Xing Y and Hogquist KA: T-cell tolerance:
Central and peripheral. Cold Spring Harb Perspect Biol. 4(pii):
a0069572012.PubMed/NCBI
|
|
66
|
Wood KJ and Sakaguchi S: Regulatory T
cells in transplantation tolerance. Nat Rev Immunol. 3:199–210.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Howard JK, Lord GM, Matarese G, Vendetti
S, Ghatei MA, Ritter MA, Lechler RI and Bloom SR: Leptin protects
mice from starvation-induced lymphoid atrophy and increases thymic
cellularity in ob/ob mice. J Clin Invest. 104:1051–1059. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Wang SD, Huang KJ, Lin YS and Lei HY:
Sepsis-induced apoptosis of the thymocytes in mice. J Immunol.
152:5014–5021. 1994.PubMed/NCBI
|
|
69
|
Müller-Hermelink HK, Sale GE, Borisch B
and Storb R: Pathology of the thymus after allogeneic bone marrow
transplantation in man. A histologic immunohistochemical study of
36 patients. Am J Pathol. 129:242–256. 1987.PubMed/NCBI
|
|
70
|
Gruver AL and Sempowski GD: Cytokines,
leptin, and stress-induced thymic atrophy. J Leukoc Biol.
84:915–923. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Boyd E: The weight of the thymus gland in
health and disease. Am J Dis Child. 43:1162–1214. 1932.
|
|
72
|
Gruver AL, Hudson LL and Sempowski GD:
Immunosenescence of ageing. J Pathol. 211:144–156. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Aw D, Silva AB and Palmer DB:
Immunosenescence: Emerging challenges for an ageing population.
Immunology. 120:435–446. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Fülöp T, Larbi A and Pawelec G: Human T
cell aging and the impact of persistent viral infections. Front
Immunol. 4:2712013. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Gruver AL, Ventevogel MS and Sempowski GD:
Leptin receptor is expressed in thymus medulla and leptin protects
against thymic remodeling during endotoxemia-induced thymus
involution. J Endocrinol. 203:75–85. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Haynes BF, Markert ML, Sempowski GD, Patel
DD and Hale LP: The role of the thymus in immune reconstitution in
aging, bone marrow transplantation, and HIV-1 infection. Annu Rev
Immunol. 18:529–560. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Billard MJ, Gruver AL and Sempowski GD:
Acute endotoxin-induced thymic atrophy is characterized by
intrathymic inflammatory and wound healing responses. PLoS One.
6:e179402011. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Hick RW, Gruver AL, Ventevogel MS, Haynes
BF and Sempowski GD: Leptin selectively augments thymopoiesis in
leptin deficiency and lipopolysaccharide-induced thymic atrophy. J
Immunol. 177:169–176. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Zhou YJ, Peng H, Chen Y and Liu YL:
Alterations of thymic epithelial cells in
lipopolysaccharide-induced neonatal thymus involution. Chin Med J
(Engl). 129:59–65. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Ann V Griffith, Venables T, Shi J, Farr A,
van Remmen H, Szweda L, Fallahi M, Rabinovitch P and Petrie HT:
Metabolic damage and premature thymus aging caused by stromal
catalase deficiency. Cell Rep. 12:1071–1079. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Dorshkind K, Montecino-Rodriguez E and
Signer RA: The ageing immune system: Is it ever too old to become
young again? Nat Rev Immunol. 9:57–62. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Gomez CR, Nomellini V, Faunce DE and
Kovacs EJ: Innate immunity and aging. Exp Gerontol. 43:718–728.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Min D, Panoskaltsis-Mortari A, Kuro-o M,
Holländer GA, Blazar BR and Weinberg KI: Sustained thymopoiesis and
improvement in functional immunity induced by exogenous KGF
administration in murine models of aging. Blood. 109:2529–2537.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Rossi SW, Jeker LT, Ueno T, Kuse S, Keller
MP, Zuklys S, Gudkov AV, Takahama Y, Krenger W, Blazar BR and
Holländer GA: Keratinocyte growth factor (KGF) enhances postnatal
T-cell development via enhancements in proliferation and function
of thymic epithelial cells. Blood. 109:3803–3811. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Hsu HC, Zhang HG, Li L, Yi N, Yang PA, Wu
Q, Zhou J, Sun S, Xu X, Yang X, et al: Age-related thymic
involution in C57BL/6J × DBA/2J recombinant-inbred mice maps to
mouse chromosomes 9 and 10. Genes Immun. 4:402–410. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Frawley R, White K Jr, Brown R, Musgrove
D, Walker N and Germolec D: Gene expression alterations in immune
system pathways in the thymus after exposure to immunosuppressive
chemicals. Environ Health Perspect. 119:371–376. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Boehm T and Swann JB: Thymus involution
and regeneration: Two sides of the same coin? Nat Rev Immunol.
13:831–838. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Bluth MH, Kohlhoff S, Norowitz KB,
Silverberg JI, Chice S, Nowakowski M, Durkin HG and Smith-Norowitz
TA: Immune responses in autoimmune hepatitis: Effect of prednisone
and azathioprine treatment: Case report. Int J Med Sci. 6:177–183.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Marchetti MC, Marco BD, Santini MC,
Bartoli A, Delfino DV and Riccardi C: Dexamethasone-induced
thymocytes apoptosis requires glucocorticoid receptor nuclear
translocation but not mitochondrial membrane potential transition.
Toxicol Lett. 139:175–180. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Gould KA, Shull JD and Gorski J: DES
action in the thymus: Inhibition of cell proliferation and genetic
variation. Mol Cell Endocrinol. 170:31–39. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Fletcher AL, Lowen TE, Sakkal S, Reiseger
JJ, Hammett MV, Seach N, Scott HS, Boyd RL and Chidgey AP: Ablation
and regeneration of tolerance-inducing medullary thymic epithelial
cells after cyclosporine, cyclophosphamide, and dexamethasone
treatment. J Immunol. 183:823–831. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Camacho IA, Singh N, Hegde VL, Nagarkatti
M and Nagarkatti PS: Treatment of mice with
2,3,7,8-tetrachlorodibenzo-p-dioxin leads to aryl hydrocarbon
receptor-dependent nuclear translocation of NF-kappaB and
expression of Fas ligand in thymic stromal cells and consequent
apoptosis in T cells. J Immunol. 175:90–103. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Dudakov JA, Hanash AM, Jenq RR, Young LF,
Ghosh A, Singer NV, West ML, Smith OM, Holland AM, Tsai JJ, et al:
Interleukin-22 drives endogenous thymic regeneration in mice.
Science. 336:91–95. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Larance M and Lamond AI: Multidimensional
proteomics for cell biology. Nat Rev Mol Cell Biol. 16:269–280.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Leung EL, Cao ZW, Jiang ZH, Zhou H and Liu
L: Network-based drug discovery by integrating systems biology and
computational technologies. Brief Bioinform. 14:491–505. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Turiák L, Misják P, Szabó TG, Aradi B,
Pálóczi K, Ozohanics O, Drahos L, Kittel A, Falus A, Buzás EI and
Vékey K: Proteomic characterization of thymocyte-derived
microvesicles and apoptotic bodies in BALB/c mice. J Proteomics.
74:2025–2033. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Billing AM, Revets D, Hoffmann C, Turner
JD, Vernocchi S and Muller CP: Proteomic profiling of rapid
non-genomic and concomitant genomic effects of acute restraint
stress on rat thymocytes. J Proteomics. 75:2064–2079. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Schulze WX and Usadel B: Quantitation in
mass-spectrometry-based proteomics. Annu Rev Plant Biol.
61:491–516. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Matt P, Fu Z, Fu Q and Van Eyk JE:
Biomarker discovery: Proteome fractionation and separation in
biological samples. Physiol Genomics. 33:12–17. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Sultana R, Di Domenico F, Tseng M, Cai J,
Noel T, Chelvarajan RL, Pierce WD, Cini C, Bondada S, St Clair DK
and Butterfield DA: Doxorubicin-induced thymus senescence. J
Proteome Res. 9:6232–6241. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Ma C, Yue QX, Guan SH, Wu WY, Yang M,
Jiang BH, Liu X and Guo DA: Proteomic analysis of possible
target-related proteins of cyclophosphamide in mice thymus. Food
Chem Toxicol. 47:1841–1847. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Kawakami T, Nagata T, Muraguchi A and
Nishimura T: Proteomic approach to apoptotic thymus maturation. J
Chromatogr B Analyt Technol Biomed Life Sci. 787:223–229. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Tyanova S, Albrechtsen R, Kronqvist P, Cox
J, Mann M and Geiger T: Proteomic maps of breast cancer subtypes.
Nat Commun. 7:102592016. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Chan PP, Wasinger VC and Leong RW: Current
application of proteomics in biomarker discovery for inflammatory
bowel disease. World J Gastrointest Pathophysiol. 7:27–37. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Peng F, Zhan X, Li MY, Fang F, Li G, Li C,
Zhang PF and Chen Z: Proteomic and bioinformatics analyses of mouse
liver microsomes. Int J Proteomics. 2012:8325692012. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Goh WW, Lee YH, Chung M and Wong L: How
advancement in biological network analysis methods empowers
proteomics. Proteomics. 12:550–563. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Miller JF: Immunological function of the
thymus. Lancet. 2:748–749. 1961. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Burns JC and Franco A: The
immunomodulatory effects of intravenous immunoglobulin therapy in
Kawasaki disease. Expert Rev Clin Immunol. 11:819–825. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Shankar-Hari M, Spencer J, Sewell WA,
Rowan KM and Singer M: Bench-to-bedside review: Immunoglobulin
therapy for sepsis - biological plausibility from a critical care
perspective. Crit Care. 16:2062012. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Gupta M, Noel GJ, Schaefer M, Friedman D,
Bussel J and Johann-Liang R: Cytokine modulation with immune
gamma-globulin in peripheral blood of normal children and its
implications in Kawasaki disease treatment. J Clin Immunol.
21:193–199. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Chaudhry MS, Velardi E, Malard F and van
den Brink MR: Immune reconstitution after allogeneic hematopoietic
stem cell transplantation: Time to T Up the thymus. J Immunol.
198:40–46. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Zhu YX, Kortuem KM and Stewart AK:
Molecular mechanism of action of immune-modulatory drugs
thalidomide, lenalidomide and pomalidomide in multiple myeloma.
Leuk Lymphoma. 54:683–687. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Ekins S, Gupta RR, Gifford E, Bunin BA and
Waller CL: Chemical space: Missing pieces in cheminformatics. Pharm
Res. 27:2035–2039. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Dobson CM: Chemical space and biology.
Nature. 432:824–828. 2004. View Article : Google Scholar : PubMed/NCBI
|