Introduction
Diabetes mellitus (DM) is a common endocrine
metabolic disorder all over the world (1–3). DM
is characterized by chronic inflammation, which can cause oxidative
stress and long-term chronic dysfunction (4–8).
Previous study has showed that increased ROS production could
contribute to the low antioxidative capacity of β-cells (5). Normally, cells could use the
superoxide dismutase (SOD), catalases (CAT), and peroxiredoxins to
combat ROS damage. Thus, oxidative stress could be reduced in
diabetic mice by increasing SOD production in previous description
(9,10). Specifically, the SOD and CAT
concentration are regulated by nuclear transcription factor-like 2
(Nrf2), resulting in the inhibition of oxidative stress and lipid
accumulation in DM (11).
Curcumin is extensively used in Asian countries, and
a wide range of pharmacological properties has been attributed to
it (12). Previous studies have
reported that curcumin has a number of beneficial effects and used
as an antioxidant and anti-inflammatory agent (13–17).
Shehzad et al noted the beneficial effects of curcumin on
obesity-related-metabolic syndrome (18). Given its beneficial effects, safety
and cost-effectiveness, curcumin could be used to treat diabetes
and complications (8,18). Moreover, previous studies have
reported that curcumin can increase oxygen consumption and the
activity of CAT, glutathione peroxidase (GSH-Px) and SOD, decrease
lipid levels and protein oxidation to protect against oxidative
stress in a high-fat diet induced diabetic rat (15,19–21).
GSH-Px, CAT, SOD and HO-1 are important parts of the antioxidant
response element (ARE), which is increased by activating Nrf2
(22,23). However, the network of curcumin
antioxidant effects resulting from the activation of the
Keap1-Nrf2-ARE signaling pathway associated with diabetes has not
been completely elucidated. Presented here is an investigation of
the capacity of curcumin to inhibit oxidative stress in a rat model
of type 1 diabetes by activating the Keap1 signaling pathway.
Materials and methods
Reagents
Streptozotocin and curcumin were purchased from
Sigma Company (USA). The kits of blood glucose, GSH-Px, SOD, CAT
and malondialdehyde (MDA) were purchased from Nanjing Jiancheng
Biologic Project Company (Nanjing, China). The kits of insulin and
glucagon radioimmunoassay were procured from North Institute of
Biological Technology (Beijing, China).
Animals and ethics statement
Seven-week-old male Sprague-Dawley (SD) rats (209±8
g) were obtained from SLAC Laboratory Animal Company (Shanghai,
China). After one-week of acclimatization to the laboratory
conditions, all male SD rats were weighed and randomly assigned.
They were fed on standard food pellets and tap water under
controlled environmental conditions (temperature: 23±2°C) and a 12
h light/dark cycle. All animal experimental procedures were
approved by Fujian Agriculture and Forestry University Animal Care
and Use Committee. All animal handling procedures were performed in
strict accordance with the care of laboratory animals of the Fujian
province Zoological Society.
Experimental design
The fifty-four SD rats were divided as follows:
group I (NC, n=18) was a control group, group II (DC,
n=18) was a diabetes control group, and group III (Diab-Cur,
n=18) was a normal diet plus 1.0% curcumin (weight ratio) group.
Considered the experiment of the power analysis, a large sample
size of 18 per group was used. All animals were fed for 21 days.
The rat model of diabetes was established by injecting
Streptozotocin (STZ) intraperitoneally at a dose of 80 mg/kg
(24), administered daily for
three days. The rats in group I were also injected
intraperitoneally with the buffer alone. After treatment, the
animals were fasted 12 h and then the fasting blood glucose levels
of all rats were measured. The rats were considered diabetic when
the fasting blood glucose levels exceeded 11.1 mmol/l. After 7
days, the rat health was worse than that before and the number of
rats that died was from the study (8/18 in Group II and 6/18 in
Group III). Thus, we were used the insulin 6 U/kg treatment once
every two day until the end of the experiment but the last time was
not used the insulin treatment in order to collected the samples.
In order to equal number in rats, the number of rats were excluded
from the study (8/18 in Group I and 2/12 in Group III) at the end
of experiment.
Blood and tissue collection
During the experiment, blood sample was collected
for biochemical measurement every three days starting on day 3. The
body weight and food and water intake of each rat were recorded
(once every day during the experiment). The animals were fasted 12
h on the last day, given mild ether anesthesia (1%, 2 min) and
killed by broken neck after collecting the blood sample. The plasma
was collected in heparin sodium vessels by centrifugation at 3,500
rpm for 15 min, and was subsequently stored at −70°C until assayed.
Tissue samples from heart, liver and leg muscle were immediately
collected, weighed, and stored at −80°C until analysis.
Blood analyzed
Blood glucose levels were determined by the glucose
oxidase method. The concentrations of MDA and the activities of SOD
(cat. no. A001-3), GSH-Px (cat. no. A006-1) and CAT (cat. no.
A007-2) were determined using assay kits by spectrophotometrically.
All produces were performed according to the manufacturer's
instruction.
The concentrations of insulin and glucagon were
measured using a radioimmunoassay kit (cat. no. 060514) according
to the manufacturer's instructions. The 125I-Ins (or
125I-Glu) and rabbit anti-Ins (or anti-Glu) antibody
were mixed with the samples, standards and controls and incubated
for 24 h. After centrifuging for 15 min, a separating medium was
removed and the sediment was saved for measurement on a
γ-counter.
RNA extraction, cDNA synthesis and
qPCR
Liver sample was collected by slaughter and
immediately frozen in liquid nitrogen on day 21. RNA was extracted
and isolated from liver tissue using TRIzol reagent (TaKaRa,
Dalian, China). The concentration of RNA was quantified in a
spectrophotometer (Eppendorf-Biotech, Hamburg, Germany) by
measuring the absorbance at 260 nm. mRNA expression was measured by
qRT-PCR according to our previous study (25) and performed using a MyiQ2 Real-time
PCR system (Bio-Rad, Hercules, USA). Primers designed using Primer
5.0 software and synthesized in the Sangon Biotech (Shanghai,
China) and listed in Table I.
 | Table I.Primers sequence and parameters. |
Table I.
Primers sequence and parameters.
| Primers | Genbank number | Primer sequence
(5′→3′) | Orientation | Product (bp) | Annealing
temperature (°C) |
|---|
| β-actin | NM_031144.3 |
CACCATGTACCCAGGCATTG | Forward | 229 | 59 |
|
|
|
ACAGTCCGCCTAGAAGCATT | Reverse |
|
|
| CAT | NM_012520.2 |
ACACTTTGACAGAGAGCGGA | Forward | 220 | 59 |
|
|
|
TTTCACTGCAAACCCACGAG | Reverse |
|
|
| GSH-Px | S50336.1 |
GACCGACCCCAAGTACATCA | Forward | 155 | 60 |
|
|
|
GCAGGGCTTCTATATCGGGT | Reverse |
|
|
| HO-1 | NM_012580.2 |
GATGGGTCCTCACACTCAGT | Forward | 201 | 59 |
|
|
|
AAGGAAGACACAGGAAGGGG | Reverse |
|
|
| NQO-1 | NM_017000.3 |
ACCTCTCTGTGGTTTAGGGC | Forward | 183 | 59 |
|
|
|
GGACCTGGGTGTGCTATGTA | Reverse |
|
|
| SOD1 | NM_017050.1 |
GCGTCATTCACTTCGAGCAG | Forward | 204 | 60 |
|
|
|
GGTCTCCAACATGCCTCTCT | Reverse |
|
|
SDS-PAGE and western blot
analysis
The protein was extracted from frozen liver tissue.
Cytoplasmic proteins and nuclear proteins were fractionted using a
CelLytic™ NuCLEAR™ Extraction kit
(Sigma-Aldrich Co. LLC, Beijing, China) (26). The protein concentration was
measured using the Bradford assay. Subsequently, 50 µg protein
samples were heated for 10 min at 98°C and subjected to
electrophoresis on a 10% SDS-PAGE gel to separate the proteins. The
separated proteins were transferred to nitrocellulose membranes
(Bio-Trace, USA) and blocked in 5% nonfat milk powder for 2 h at
20°C. After blocking, the protein was incubated overnight at 4°C
with anti-keap1 antibody (1:1,500, Abcam) and anti-Nrf2 antibody
(1:1,000, Abcam). After washing three times, the corresponding HRP
conjugated secondary antibodies were incubated at 4°C. The
anti-GADPH antibody (1:1,000, Abcam) and lamin B (1:1,000, Abcam)
was used served as loading controls of cytoplasmic and nuclear
fractions, respectively. Finally, blots were washed and detected by
enhanced chemiluminescence (ECL) using the LumiGlo substrate
(Pierce, USA) and Clarity Western ECL Substrate (BioRad, USA).
Statistical analysis
The statistical significance was analyzed by Tukey's
test model of ANOVA (SPSS-20.0 software; IBM Corp., Armonk, NY,
USA). P<0.05 was considered to indicate statistical
significance. The results were expressed as mean ± SE.
Results
Body weight, food and water intake,
blood glucose, and plasma insulin and glucogan concentration
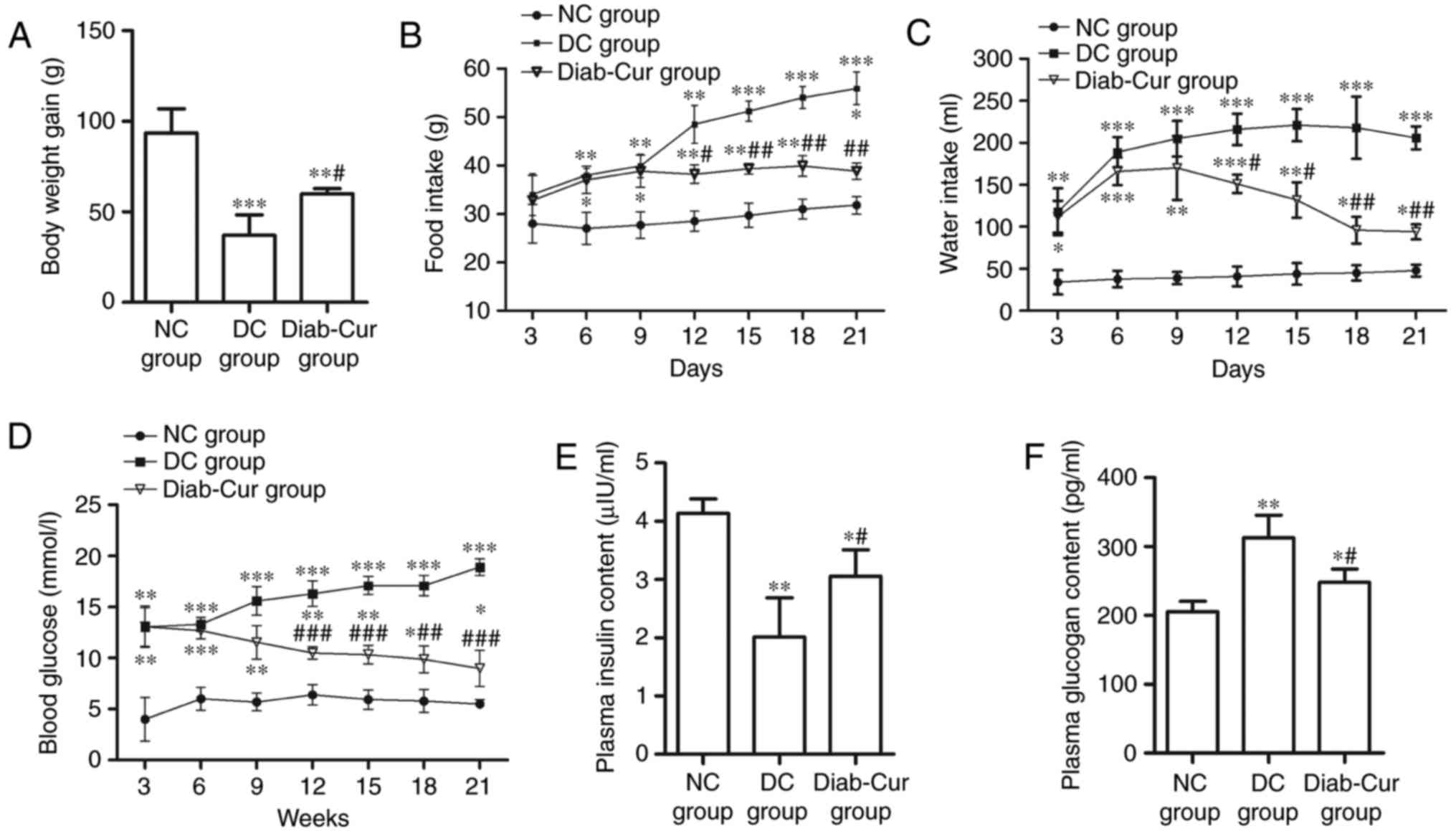
After 21 days of the experiment, the body weight
(BW) of the rats was significantly reduced in the Diab-Cur group
(P<0.05) and DC group (P<0.01) compared with the NC group.
However, the BW of rats was markedly increased in the Diab-Cur
group (P<0.05) compared to that in the DC group (Fig. 1A). The food and water intake was
distinctly increased in the Diab-Cur group compared with NC group
(Fig. 1B and C). After the 9th
day, curcumin treatment obviously reduced the food intake on days
12 (P<0.05), 15 (P<0.01), 18 (P<0.01), and 21 (P<0.01),
and reduced the water intake on days 12 (P<0.05), 15
(P<0.05), 18 (P<0.01), and 21 (P<0.01) compared with the
DC group.
The concentration of blood glucose was obviously
increased in the DC group and Diab-Cur group compared with the NC
group (Fig. 1D). After day 9, the
blood glucose level was significantly reduced on days 12
(P<0.001), 15 (P<0.001), 18 (P<0.01), and 21 (P<0.001)
in the Diab-Cur group. The plasma insulin concentration was
markedly decreased (P<0.05) in the DC and Diab-Cur groups
compared with the NC group, whereas the plasma insulin
concentration in the Diab-Cur group was obviously higher than those
in the DC group (P<0.05, Fig.
1E). Furthermore, the plasma glucagon levels were significantly
increased in the Diab-Cur group (P<0.05) and DC group
(P<0.01) compared with those in the NC group rats, but the
glucagon levels were significantly reduced (P<0.05) in the
Diab-Cur group compared with the NC group (Fig. 1F).
Tissue glycogen content
As shown in Table
II, the glycogen concentrations of hepatic, myocardial and
muscle tissue in the DC group were significantly decreased
(P<0.05) compared with the NC group, whereas the hepatic,
myocardial and muscular glycogen levels were significantly
increased in the Diab-Cur group (P<0.05). Although the levels of
hepatic and muscular glycogen were obviously decreased in the
Diab-Cur group compared with NC group rats (P<0.05), the
myocardial glycogen levels were significantly increased
(P<0.05).
| Table II.Concentrations of hepatic, myocardial
and muscular glycogen in type 1 diabetic rats. |
Table II.
Concentrations of hepatic, myocardial
and muscular glycogen in type 1 diabetic rats.
| Group | Hepatic (mg/g) | Myocardial
(mg/g) | Muscular
(mg/g) |
|---|
| NC |
11.02±2.42 |
0.75±0.18 |
0.86±0.39 |
| DC |
6.02±2.11b |
0.58±0.13a |
0.55±0.21a |
| Diab-Cur |
7.58±2.87a,d |
1.23±0.23a,d |
0.61±0.18a,c |
Plasma SOD, GSH-Px, CAT and MDA
level
As shown in Table
III, there was an obvious reduction of SOD activity in the DC
group compared with the NC group (P<0.01), but curcumin
treatment significantly increased SOD activity compared with the
levels in both the NC and DC groups (P<0.05 and P<0.01,
respectively). Compared to the NC group, the levels of GSH-Px, CAT
and MDA in the DC and Diab-Cur groups were significantly increased
(P<0.05). Compared with DC group, the levels of GSH-Px
(P<0.01), CAT (P<0.05) and MDA (P<0.01) were significantly
decreased in the Diab-Cur group.
| Table III.Plasma antioxidant enzyme levels of
superoxide dismutase, catalases and glutathione peroxidase and
malondialdehyde. |
Table III.
Plasma antioxidant enzyme levels of
superoxide dismutase, catalases and glutathione peroxidase and
malondialdehyde.
| Group | SOD (U/ml) | GSH-Px (U/ml) | CAT (U/ml) | MDA (nmol/ml) |
|---|
| NC |
26.01±4.47 |
1,732.40±73.20 |
19.03±3.04 |
6.03±1.03 |
| DC |
20.99±2.11b |
1,997.88±65.43c |
27.95±1.33b |
9.82±0.67c |
| Diab-Cur |
29.42±4.43a,e |
1,882.38±42.71a,d |
23.15±2.29a,d |
6.87±0.24a,e |
Antioxidase gene expression
As shown in Fig. 2,
the results showed that expression levels of the antioxidase genes
NAD (P)H quinone dehydrogenase 1 (NQO-1), CAT,
GSH-Px, and HO-1 were significantly upregulated in
the Diab-Cur group compared with those in the NC (P<0.05) and DC
groups (P<0.05). However, the antioxidase gene expression levels
of GSH-Px, HO-1 and NQO-1 were markedly
downregulated (P<0.05) in the DC group. Notably, the gene
expression of SOD1 was completely contrary to the expression
profiles of the other antioxidase genes.
 | Figure 2.Gene expression of several
antioxidant response element-regulated genes. RNA was extracted
from the liver, reverse-transcribed to cDNA and analyzed by reverse
transcription-quantitative polymerase chain reaction for gene
expression. The experiments used 18 samples from each group. Data
are presented as the mean ± standard error. *P<0.05, **P<0.01
and ***P<0.001 vs. NC group; #P<0.05,
##P<0.01 and ###P<0.001 vs. DC group.
NC, negative control; DC, diabetic control; Diab-Cur, diabetic with
1.0% curcumin treatment; HO-1, heme oxygenase-1; NQO-1, norvegicus
NAD(P)H quinone dehydrogenase 1; GSH-Px, glutathione peroxidase;
CAT, catalase; SOD1, superoxide dismutase 1. |
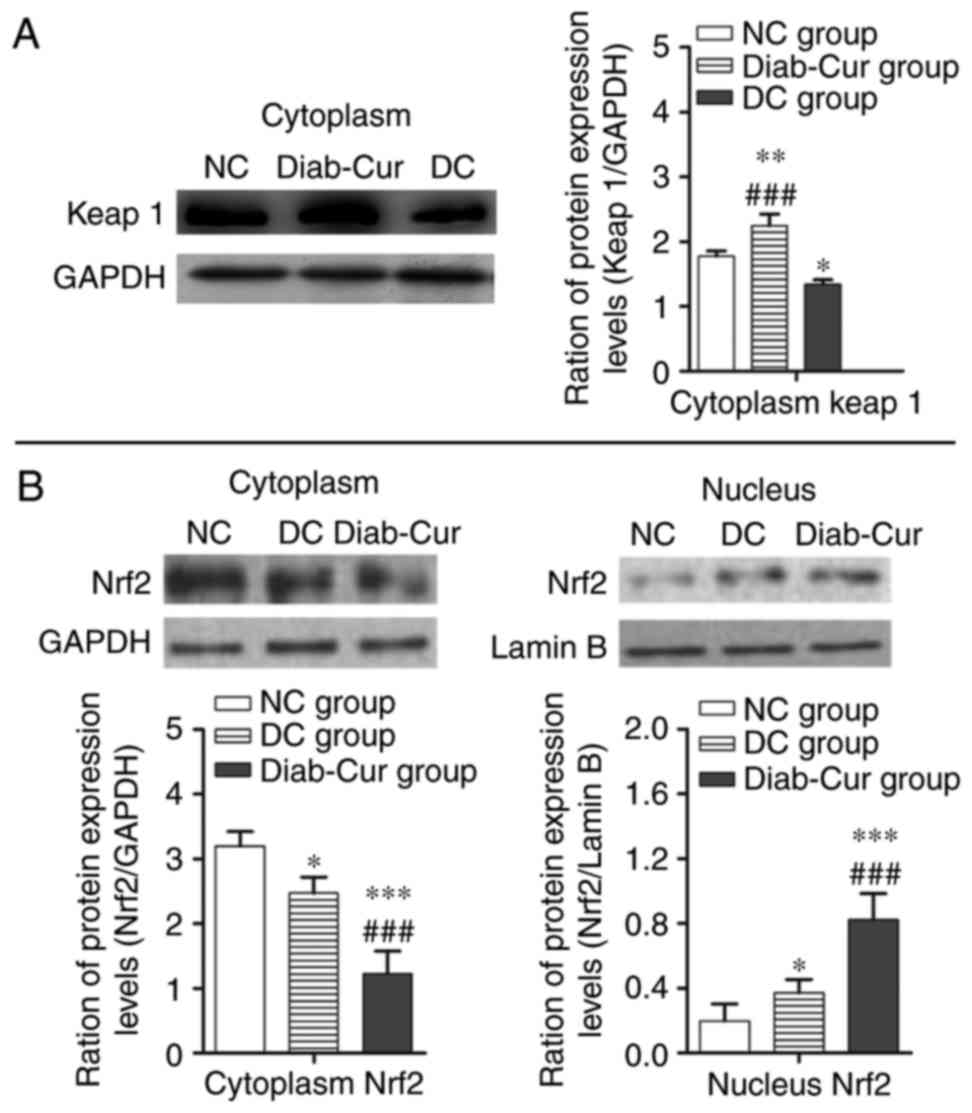
Protein expression
As illustrated in Fig.
3A, compared with the NC group, the protein expression level of
keap1 was significantly upregulated in the Diab-Cur group rats
(P<0.01), but downregulated in the DC group (P<0.05).
Compared with the DC group, protein expression of keap1 was
significantly upregulated in the Diab-Cur group (P<0.001). As
demonstrated in Fig. 3B, curcumin
treatment decreased cytosolic concentrations of Nrf2 while
increasing nuclear accumulation of Nrf2.
Discussion
STZ-induced type 1 diabetes models are characterized
by a loss of β-cells in the islets of Langerhans in the pancreas,
which leads to insulin deficiency and metabolic disease caused by
elevated blood sugar (7,8,27,28).
In this study, the results showed that BW was markedly decreased
and that the blood glucose levels and food and water intake were
clearly increased in rat with type 1 diabetes. Especially, the
blood glucose levels were elevated more than twofold in the DC
group compared with those of control group, whereas
curcumin-treated significantly decreased the concentrations of
blood glucose (Fig. 1D). Previous
studies have shown that curcumin has anti-diabetic activities and
that dietary supplements containing curcumin affect glycemic
control (12,29,30).
Moreover, curcumin supplementation to rat model in diabetes showed
that plasma insulin concentration was obviously increased and
plasma glucagon levels were markedly reduced (30). In this study, a decrease of 58% in
the level of plasma insulin and an increase of 52% in the level of
plasma glucagon when compared to the control group were observed
(Fig. 1E and F). But,
curcumin-treated rat model significantly increased plasma insulin
levels (Fig. 1E) and decreased
plasma glucagon levels (Fig. 1F).
At the same time, curcumin treated diabetic animals also showed a
significant increase in the hepatic and myocardial glycogen
concentrations (Table II). These
results imply that curcumin could reduce hyperglycemia.
However, the autoxidation of glucose could increase
due to free radicals production with chronic hyperglycemia during
the diabetes, which is involved in β-cell dysfunction (5,6,27).
β-cell dysfunction could lead to β-cell death, which depends on the
subsequent poly (ADP-ribose) synthetase activation and DNA
alkylation (5,6,31–33).
Moreover, decreases in the expression of antioxidant enzymes and
the range of minimum antioxidant competence of β-cells could lead
to free radical damage (28,34).
Previous results have demonstrated that the activity of the
antioxidant enzymes SOD, CAT and GSH-Px can prevent oxidative
stress (35). Importantly, SOD and
CAT play important roles in the detoxification of O2-
(35,36). GSH-Px can regulate the
intracellular redox system to defend cells against oxidative stress
(37). Additionally, through the
dismutation and generation of hydrogen peroxide, SOD destroys the
superoxide radical, whereas SOD activity is attenuated by catalase
or glutathione peroxidase (36,38,39).
Recently, reports have shown that pancreatic β-cells were not
damaged by oxidative stress under diabetic conditions by increasing
catalase activity (5,38). In the present study, plasma MDA
concentration, GSH-Px and CAT activity were significantly increased
and indicated that an impairment in antioxidant defenses may
increase the scavenging oxygen free radicals (36,40,41)
or enhance the antioxidant capacity (19,20,30).
In a previous study, activation of the Keap1-Nrf2-ARE pathway was
shown to reduce oxidative stress (42). Additional studies have demonstrated
that Nrf2 signaling, as activated by salvianolic acid, can be
effective against oxidative stress (22,43–45).
In the present study, we showed that Nrf2 signaling was activated
in curcumin-treated rats. First, the activities of the keap1-Nrf2
complex were reduced, which led to the uncoupling of keap1-Nrf2 in
curcumin-treated rats. Second, Nrf2 was stabilized and transported
into cell nuclei, leading to the transcription of several
ARE-regulated genes (HO-1, NQO-1, GSH-Px,
CAT and SOD1). Intriguingly, previous studies have
shown that upregulating the expression of Nrf2 protein in the
nuclear and increased the expression of HO1 and NQO-1
(46). In this study, the
expression of the Nrf2 protein and ARE-regulated genes was
upregulated in curcumin-treated rats compared to those in the DC
group. Importantly, the activation of Nrf2 by curcumin dramatically
relieved oxidative stress in the STZ-induced diabetic rat
model.
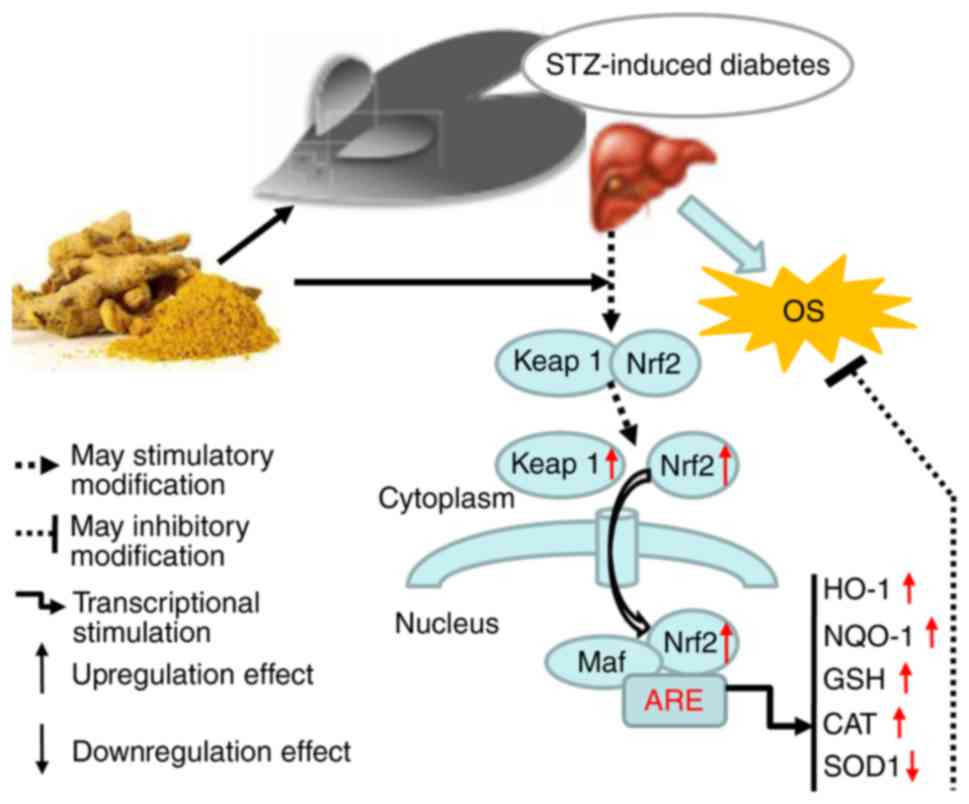
In conclusion, a diagram illustrating the proposed
mechanism of action of curcumin is shown in Fig. 4. We propose that curcumin may be
exerts protective effects in STZ-induced diabetes by activating
Keap1-Nrf2-ARE signaling through both a decrease in blood glucose
concentration and an increase in the transcription of several
antioxidant genes.
Acknowledgements
The present study was supported by the Initial
Funding of Jinshan College of Fujian Agriculture and Forestry
University (grant no. Z150701); the Natural Science Foundation of
Fujian Province (grant no. 2016J01091), and Scientific Research
Foundation for Young and Middle-aged Teachers of Fujian Province
(grant no. JAT160686).
References
|
1
|
Jiménez-Flores LM, López-Briones S,
Macías-Cervantes MH, Ramírez-Emiliano J and Pérez-Vázquez V: A
ppargamma, NF-κB and AMPK-dependent mechanism may be involved in
the beneficial effects of curcumin in the diabetic db/db mice
liver. Molecules. 19:8289–8302. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kume S, Koya D, Uzu T and Maegawa H: Role
of nutrient-sensing signals in the pathogenesis of diabetic
nephropathy. Biomed Res Int. 2014:3154942014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gao Q, Shen W, Qin W, Zheng C, Zhang M,
Zeng C, Wang S, Wang J, Zhu X and Liu Z: Treatment of db/db
diabetic mice with triptolide: A novel therapy for diabetic
nephropathy. Nephrol Dial Transplant. 25:3539–3547. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wang K: Molecular mechanisms of hepatic
apoptosis regulated by nuclear factors. Cell Signal. 27:729–738.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gerber PA and Rutter GA: The role of
oxidative stress and hypoxia in pancreatic beta-cell dysfunction in
diabetes mellitus. Antioxid Redox Signal. 26:501–518. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kajimoto Y and Kaneto H: Role of oxidative
stress in pancreatic beta-cell dysfunction. Ann N Y Acad Sci.
1011:168–176. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Maritim AC, Sanders RA and Watkins JB III:
Diabetes, oxidative stress, and antioxidants: A review. J Biochem
Mol Toxicol. 17:24–38. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Karunakaran U and Park KG: A systematic
review of oxidative stress and safety of antioxidants in diabetes:
Focus on islets and their defense. Diabetes Metab J. 37:106–112.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lieben L: Diabetic nephropathy: Lipid
toxicity drives renal disease. Nat Rev Nephrol. 13:1942017.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Flyvbjerg A: The role of the complement
system in diabetic nephropathy. Nat Rev Nephrol. 13:311–318. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nakai K, Fujii H, Kono K, Goto S, Kitazawa
R, Kitazawa S, Hirata M, Shinohara M, Fukagawa M and Nishi S:
Vitamin d activates the nrf2-keap1 antioxidant pathway and
ameliorates nephropathy in diabetic rats. Am J Hypertens.
27:586–595. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hsu CH and Cheng AL: Clinical studies with
curcumin. Adv Exp Med Biol. 595:471–480. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gupta SC, Patchva S, Koh W and Aggarwal
BB: Discovery of curcumin, a component of golden spice, and its
miraculous biological activities. Clin Exp Pharmacol Physiol.
39:283–299. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shishodia S: Molecular mechanisms of
curcumin action: Gene expression. Biofactors. 39:37–55. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sikora-Polaczek M, Bielak-Zmijewska A and
Sikora E: Molecular and cellular mechanisms of curcumin
action-beneficial effect on organism. Postepy Bioch. 57:74–84.
2011.
|
|
16
|
Joe B, Vijaykumar M and Lokesh BR:
Biological properties of curcumin-cellular and molecular mechanisms
of action. Crit Rev Food Sci Nutr. 44:97–111. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pescosolido N, Giannotti R, Plateroti AM,
Pascarella A and Nebbioso M: Curcumin: Therapeutical potential in
ophthalmology. Planta Med. 80:249–254. 2014.PubMed/NCBI
|
|
18
|
Shehzad A, Ha T, Subhan F and Lee YS: New
mechanisms and the anti-inflammatory role of curcumin in obesity
and obesity-related metabolic diseases. Eur J Nutr. 50:151–161.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Motterlini R, Foresti R, Bassi R and Green
CJ: Curcumin, an antioxidant and anti-inflammatory agent, induces
heme oxygenase-1 and protects endothelial cells against oxidative
stress. Free Radic Biol Med. 28:1303–1312. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Martínez-Morúa A, Soto-Urquieta MG,
Franco-Robles E, Zúñiga-Trujillo I, Campos-Cervantes A,
Pérez-Vázquez V and Ramírez-Emiliano J: Curcumin decreases
oxidative stress in mitochondria isolated from liver and kidneys of
high-fat diet-induced obese mice. J Asian Nat Prod Res. 15:905–915.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bhatia NK, Srivastava A, Katyal N, Jain N,
Khan MA, Kundu B and Deep S: Curcumin binds to the pre-fibrillar
aggregates of cu/zn superoxide dismutase (sod1) and alters its
amyloidogenic pathway resulting in reduced cytotoxicity. Biochim
Biophys Acta. 1854:426–436. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhou J, Qu XD, Li ZY, Wei J, Liu Q, Ma YH
and He JJ: Salvianolic acid b attenuates toxin-induced neuronal
damage via nrf2-dependent glial cells-mediated protective activity
in Parkinson's disease models. PLoS One. 9:e1016682014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zenkov NK, Menshchikova EB and Tkachev VO:
Keap1/nrf2/are redox-sensitive signaling system as a
pharmacological target. Biochemistry (Mosc). 78:19–36. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Golalipour MJ, Ghafari S, Kouri V and
Kestkar AA: Proliferation of the b-cells of pancreas in diabetic
rats treated with urtica dioica. Int J Morphol. 28:399–404.
2010. View Article : Google Scholar
|
|
25
|
Xie ZL, Ye PS, Zhang SK, Zhang YS and Shen
XZ: Endogenous lps alters liver GH/IGF system gene expression and
plasma lipoprotein lipase in goats. Physiol Res. 64:721–729.
2015.PubMed/NCBI
|
|
26
|
Deml B, Kariminejad A, Borujerdi RH,
Muheisen S, Reis LM and Semina EV: Mutations in mab21l2 result in
ocular coloboma, microcornea and cataracts. PLoS Genet.
11:e10050022015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Szkudelski T: The mechanism of alloxan and
streptozotocin action in B cells of the rat pancreas. Physiol Res.
50:537–546. 2001.PubMed/NCBI
|
|
28
|
Tiedge M, Lortz S, Drinkgern J and Lenzen
S: Relation between antioxidant enzyme gene expression and
antioxidative defense status of insulin-producing cells. Diabetes.
46:1733–1742. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yeh GY, Eisenberg DM, Kaptchuk TJ and
Phillips RS: Systematic review of herbs and dietary supplements for
glycemic control in diabetes. Diabetes Care. 26:1277–1294. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Rahimi HR, Mohammadpour AH, Dastani M,
Jaafari MR, Abnous K, Mobarhan Ghayour M and Oskuee Kazemi R: The
effect of nano-curcumin on HbA1c, fasting blood glucose and lipid
profile in diabetic subjects: A randomized clinical trial. Avicenna
J Phytomed. 6:567–577. 2016.PubMed/NCBI
|
|
31
|
Bennett RA and Pegg AE: Alkylation of DNA
in rat tissues following administration of streptozotocin. Cancer
Res. 41:2786–2790. 1981.PubMed/NCBI
|
|
32
|
Vikram A, Tripathi DN, Ramarao P and Jena
GB: Evaluation of streptozotocin genotoxicity in rats from
different ages using the micronucleus assay. Regul Toxicol
Pharmacol. 49:238–244. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Damasceno DC, Volpato GT, Sinzato YK, Lima
PH, Souza MS, Iessi IL, Kiss AC, Takaku M, Rudge MV and Calderon
IM: Genotoxicity and fetal abnormality in streptozotocin-induced
diabetic rats exposed to cigarette smoke prior to and during
pregnancy. Exp Clin Endocrinol Diabetes. 119:549–553. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lenzen S, Drinkgern J and Tiedge M: Low
antioxidant enzyme gene expression in pancreatic islets compared
with various other mouse tissues. Free Radic Biol Med. 20:463–466.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Pigeolet E, Corbisier P, Houbion A,
Lambert D, Michiels C, Raes M, Zachary MD and Remacle J:
Glutathione peroxidase, superoxide dismutase, and catalase
inactivation by peroxides and oxygen derived free radicals. Mech
Ageing Dev. 51:283–297. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Meghana K, Sanjeev G and Ramesh B:
Curcumin prevents streptozotocin-induced islet damage by scavenging
free radicals: A prophylactic and protective role. Eur J Pharmacol.
577:183–191. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Grant CM, MacIver FH and Dawes IW:
Glutathione is an essential metabolite required for resistance to
oxidative stress in the yeast saccharomyces cerevisiae. Curr Genet.
29:511–515. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Jin L, Xue HY, Jin LJ, Li SY and Xu YP:
Antioxidant and pancreas-protective effect of aucubin on rats with
streptozotocin-induced diabetes. Eur J Pharmacol. 582:162–167.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Liu Z, Li J, Zeng Z, Liu M and Wang M: The
antidiabetic effects of cysteinyl metformin, a newly synthesized
agent, in alloxan- and streptozocin-induced diabetic rats. Chem
Biol Interact. 173:68–75. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Afanasiev SA, Kondratieva DS, Rebrova TY,
Batalov RE and Popov SV: Coupling of the functional stability of
rat myocardium and activity of lipid peroxidation in combined
development of postinfarction remodeling and diabetes mellitus. J
Diabetes Res. 2016:25486892016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
He X, de Seymour JV, Sulek K, Qi H, Zhang
H, Han TL, Villas-Bôas SG and Baker PN: Maternal hair metabolome
analysis identifies a potential marker of lipid peroxidation in
gestational diabetes mellitus. Acta Diabetol. 53:119–122. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Shi L, Wu L, Chen Z, Yang J, Chen X, Yu F,
Zheng F and Lin X: Mir-141 activates nrf2-dependent antioxidant
pathway via down-regulating the expression of keap1 conferring the
resistance of hepatocellular carcinoma cells to 5-fluorouracil.
Cell Physiol Biochem. 35:2333–2348. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lin M, Zhai X, Wang G, Tian X, Gao D, Shi
L, Wu H, Fan Q, Peng J, Liu K and Yao J: Salvianolic acid b
protects against acetaminophen hepatotoxicity by inducing nrf2 and
phase II detoxification gene expression via activation of the pi3k
and pkc signaling pathways. J Pharmacol Sci. 127:203–210. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhang H, Liu YY, Jiang Q, Li KR, Zhao YX,
Cao C and Yao J: Salvianolic acid a protects RPE cells against
oxidative stress through activation of NRF2/HO-1 signaling. Free
Radic Biol Med. 69:219–228. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lee HJ, Seo M and Lee EJ: Salvianolic acid
B inhibits atherogenesis of vascular cells through induction of
NRF2-dependent heme oxygenase-1. Curr Med Chem. 21:3095–3106. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Suzuki T and Yamamoto M: Molecular basis
of the keap1-NRF2 system. Free Radic Biol Med. 88:93–100. 2015.
View Article : Google Scholar : PubMed/NCBI
|