Introduction
Hepatocellular carcinoma (HCC) is among the most
common types of malignant tumor. Various genetic and environmental
risk factors have been identified for HCC, including aflatoxin,
hepatitis, genetic factors (liver cancer of first-degree relatives)
and adverse environmental factors (chemical contamination of the
water environment) (1). A previous
study identified microRNAs (miRNAs/miRs), which are short
non-coding RNAs, as potential biomarkers and therapeutic targets in
HCC. An increasing amount of research has demonstrated that miRNAs
modulate gene expression in cancer and may promote or inhibit tumor
growth (2–4). miRNAs affect the stability and
transcriptional activity of their target mRNAs by binding to the
3′-untranslated regions (UTRs) of mRNA (5). miRNAs may be employed as biomarkers
and have an important role in cancer diagnosis and prognosis.
Therefore, miRNAs have the potential to be developed as targets for
anti-cancer drugs (6).
miR-19a forms part of the miR-17-92 cluster.
Research has revealed that its overexpression occurs in various
tumor types including liver (3,4),
lung (7), prostate (8) and breast cancer (9), gastric carcinoma (10), colorectal cancer (11) and osteosarcoma (12), and this overexpression may
stimulate tumor growth and inhibit tumor cell apoptosis. In the
present study, the downregulation of miR-19a prevented
proliferation, promoted cell apoptosis and inhibited cell invasion
in the SMMC-7721 HCC cell line. Additionally, reduced miR-19a
expression led to the upregulation of phosphatase and tensin
homolog (PTEN) and suppression of tumor growth, demonstrating that
PTEN may exhibit antioncogenic effects in HCC cells. Previous
investigation of the molecular biology of HCC has demonstrated that
the PTEN/Akt signaling pathway (13–17)
is closely associated with the proliferation, apoptosis and cell
cycle arrest of HCC cells (18–21).
It has also been reported that miR-19a may regulate the PTEN/Akt
signaling pathway and subsequently affect the biological behavior
of HCC cells (22,23). These discoveries are important for
the regulation of cancer cell growth and the development of
targeted cancer therapies.
Pterostilbene (Pter) is a derivative of resveratrol,
which is an established epigenetic modulator. It is extracted from
dietary compounds such as peanuts, grapes, berries and red wine,
and is associated with anti-inflammatory, antioxidative,
cardioprotective and antioncogenic activity (17,24).
Pter may modulate various molecular targets, including miR-19a, to
promote apoptosis, cell cycle arrest and cancer growth suppression
(25). The present study
demonstrated that the antioncogenic effect of Pter may be a result
of miR-19a downregulation, which subsequently leads to an increase
in PTEN expression.
Materials and methods
Reagents
Pter (purity, >99%; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) was dissolved in dimethyl sulfoxide (DMSO;
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany).
Cell culture
The SMMC-7721 HCC cell line (China Center for Type
Culture Collection, Wuhan University, Wuhan, China) was cultured
and maintained in RPMI-1640 (Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) containing 10% fetal bovine serum (FBS; Gibco;
Thermo Fisher Scientific, Inc.) with 100 U/ml penicillin and 100
U/ml streptomycin. Cells were incubated at 37°C with 5%
CO2.
Cell transfection
SMMC-7721 cells at a density of 60–70% confluence
were divided into the following five groups: Pter treatment group
(50 µM Pter), miR-19a inhibitor group (transfected with miR-19a
inhibitor 5′-UCAGUUUUGCAUAGAUUUGCACA-3′), Pter + miR19a inhibitor
group (50 µM Pter and transfected with miR-19a inhibitor), negative
control group (transfected with negative control
5′-UUGUACUACACAAAAGUACUG-3′) and the blank group (DMSO treatment)
at 37°C for 48 h. Lipofectamine 2000 (5 µl; Invitrogen; Thermo
Fisher Scientific, Inc.) was diluted to 100 µl in serum-free
RPMI-1640 medium. miR-19a inhibitor (20 pM) or negative control (20
pM) was also diluted to 50 µl in the serum-free RPMI-1640 medium at
room temperature for 5 min. Both of the dilutions were subsequently
mixed at room temperature for 20 min prior to transfer into culture
plates for cell transfection at 37°C in 5% CO2 for 6 h.
The transfected cells were subsequently cultivated in complete
medium (RPMI-1640 medium containing 10% FBS with 100 U/ml
penicillin and 100 U/ml streptomycin) for 48 h and then the
subsequent experiments including qPCR, western blotting, MTT and
cell cycle and apoptosis assay were performed.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Following treatment of cells (5×107
cells/well) with Ctrl (DMSO), 5, 25, 50 and 100 µM Pter at 37°C for
24 h, and the establishment of the 5 treatment groups described
above, the total RNA was extracted from SMMC-7721 cells using
TRIzol (Life Technologies; Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol. RNA concentration was
detected at an absorbance of A260 and A280.
cDNA was synthesized by using a PrimeScript RT reagent kit (Takara
Bio, Inc., Otsu, Japan). The PCR amplification primer sequences are
presented in Table I. qPCR was
performed by the MiniOpticon Real-Time PCR system (Bio-Rad
Laboratories, Inc., Hercules, CA, USA) using SYBR Premix Ex Taq
(Takara Bio, Inc.). Following initial denaturation at 95°C for 30
sec, amplifications were performed for 40 cycles at 95°C for 5 sec
and 60°C for 30 sec. miR-19a levels were compared to U6 as an
internal reference and PTEN was compared with β-actin. qPCR was
repeated 3 times. The relative expression of miRNA and mRNA was
calculated by the 2−ΔΔCq method (26).
 | Table I.Primer sequences for quantitative
polymerase chain reaction amplification. |
Table I.
Primer sequences for quantitative
polymerase chain reaction amplification.
|
| Primer
sequence |
|---|
|
|
|
|---|
| Target | Forward | Reverse |
|---|
| miR-19a |
5′-GCGTGTGCAAATCTATGCAA-3′ |
5′-GTGCAGGGTCCGAGGT-3′ |
| U6 |
5′-CTCGCTTCGGCAGCACA-3′ |
5′-AACGCTTCACGAATTTGCGT-3′ |
| PTEN |
5′-CCAAGCTTATGACAGCCATCATC-3′ |
5′-CGCGGATCCTCAGACTTTTGTAA-3′ |
| β-actin |
5′-GAATCAATGCAAGTTCGGTTCC-3′ |
5′-TCATCTCCGCTATTAGCTCCG-3′ |
Western blot analysis
Following treatment of cells (5×104
cells/well) with Ctrl (DMSO), 5, 25, 50 and 100 µM Pter at 37°C for
24 h, and establishment of the 5 treatment groups described above,
whole proteins were extracted by RIPA lysis buffer (Beyotime
Institute of Biotechnology, Shanghai, China) from SMMC-7721 cells
which was quantified by a BCA Protein Assay kit (Beyotime Institute
of Biotechnology) and separated at 20 µg/lane by 15% SDS-PAGE prior
to transfer onto polyvinylidene difluoride membranes (Bio-Rad
Laboratories, Inc.) using an Electrophoresis Transfer System
(Bio-Rad Laboratories, Inc.). The blots blocked with 5% fat-free
milk in a Tris-buffered saline with 5% Tween 20 (TBST) for 1 h at
room temperature, were probed with anti-PTEN (mouse monoclonal;
cat. no. ab79156; 1:500; Abcam, Cambridge, UK), anti-Akt (Rat
polyclonal; cat. no. ab8805; 1:500; Abcam), anti-phosphorylated
(p)-Akt (Rat polyclonal; cat. no. ab8933; 1:500; Abcam) and
anti-β-actin (Rat polyclonal; cat. no. ab8227; 1:2,000; Abcam)
primary antibodies overnight at 4°C, and subsequently treated with
the respective secondary antibodies (Rat, cat. no. ab218695;
1:4,000 or mouse cat. no. ab131368; 1:4,000 dilution; Abcam) for 1
h at 4°C. The proteins were visualized by using an enhanced
chemiluminescence detection kit (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). Densitometric analysis was performed to quantify
protein expression using ImageJ software version 1.49 (National
Institutes of Health, Bethesda, MD, USA).
MTT assay
Following treatment of cells (5×104
cells/well) with Ctrl (DMSO), 20, 40, 60, 80 and 100 µM Pter at
37°C for 24, 48 or 72 h, and establishment of the five treatment
groups described in the cell transfection section above, the
proliferation of SMMC-7721 was detected by MTT assays
(Sigma-Aldrich; Merck KGaA). To investigate cell viability, 0.5
mg/ml MTT solution was added into each well following seeding of
the treated cells for 24 h at a density of 3×105
cells/ml per well. Cells were subsequently incubated for 4 h at
37°C with 5% CO2. The supernatant liquid was removed and
150 µl DMSO was added to each well. A microplate reader (BioTek
Instruments, Inc., Winooski, VT, USA) was used to determine optical
density values at 570 nm after 24, 48 and 72 h. For the experiments
that consisted of five different treatment groups, measurements
were taken after 24 h.
Cell cycle and apoptosis analysis by
flow cytometry
Cell cycle and apoptosis was examined in the five
treatment groups by flow cytometry. Annexin V-fluorescein
isothiocyanate (10 µl) and propidium iodide (PI; 5 µl,
Sigma-Aldrich; Merck KGaA) were incubated with the cells
(5×105 cells/well) in each group in the dark at 4°C for
30 min. A flow cytometer (BD Biosciences, San Jose, CA, USA) was
used to calculate the percentage of apoptotic cells with FlowJo
version 10 software (FlowJo LLC, Ashland, OR, USA).
To analyze the cell cycle, cells (5×105
cells/well) were washed twice with PBS and fixed with 75% ethanol
overnight at 4°C. The cells were subsequently stained with 5 µl
PI/ribonuclease A (Sigma-Aldrich; Merck KGaA) at 4°C for 30 min in
the dark. Data was analyzed with a flow cytometer (BD Biosciences).
A total of 14,000 fluorescence signals for each PI-stained sample
were collected and calculated by ModFit LT version 3.2 software
(Verity Software House, Inc., Topsham, ME, USA).
Cell invasion
SMMC-7721 cell invasion ability was analyzed in
Transwell chambers (pore size, 8 µm; Corning Incorporated, Corning,
NY, USA) coated with Matrigel in serum-free medium. Following
incubation, the 5×104 cells/well were added into the
upper chamber at 37°C in a humidified 5% CO2 for 24 h.
Then the cells were removed from the upper chamber (medium
containing serum-free RPMI-1640) and the invading cells in the
lower chamber (medium containing 20% FBS) were fixed with 4%
paraformaldehyde at 37°C for 15 min, then stained with 0.1% crystal
violet at 37°C for 15 min and counted using a phase-contrast
microscope at ×200 magnification of each membrane in 5
representative fields.
Luciferase reporter gene assay
The binding sequence of PTEN 3′UTR and miR-19a was
predicted by TargetScanHuman version 7.1 (www.targetscan.org/vert_71/). The pGL3-Luciferase
vector with firefly and Renilla luciferase activity (Wuhan Jin
Kairui Biological Engineering Co., Ltd, Wuhan, China) was digested
with XbaI and the target sequence was inserted. The promoter
sequence of PTEN 3′UTR containing the binding site was synthesized
as the wild-type (WT) plasmid (PTEN-WT). Similarly, a mutated
binding site of PTEN was constructed as the mutant-type (MT)
plasmid (PTEN-MT). The cells (5×104 cells/well) were
transfected by using Lipofectamine 2000® (5 µl;
Invitrogen; Thermo Fisher Scientific, Inc.) at 37°C for 48 h. The
cells were subsequently co-transfected with the luciferase reporter
plasmid (20 pM) and the miR-19a inhibitor (20 pM) or negative
control (20 pM) prior to treatment with the Dual-Luciferase
Reporter Assay System (Promega Corporation, Madison, WI, USA),
according to the manufacturer's protocol. Reporter gene activity
was detected and normalized with the ratio of Renilla and firefly
luciferase activity.
Statistical analysis
Data are presented as the mean ± standard deviation
from three independent experiments. All statistical analyses were
performed with SPSS version 21.0 software (IBM Corp., Armonk, NY,
USA). And unpaired Student's t-tests were employed to analyze the
difference between two groups. P<0.05 was considered to indicate
a statistically significant difference.
Results
Pter and miR-19a inhibitor
downregulate miR-19a expression
Previous reports have indicated that the modulation
of PTEN-targeting miRNAs by Pter in prostate cancer led to the
overexpression of PTEN and downregulation of PI3K-Akt pathway
activity (17,27). Thus, the present study aimed to
confirm this hypothesis in HCC cells. qPCR was performed to measure
the relative expression of miR-19a in SMMC-7721 cells. The results
demonstrated that miR-19a expression was decreased by Pter
treatment in a dose-dependent manner in SMMC-7721 cells, indicating
that Pter may exhibit important antioncogenic activity (P<0.05;
Fig. 1A). In addition, miR-19a
expression was also significantly lower in the Pter, miR-19a
inhibitor and Pter + miR-19a inhibitor groups, compared with the
blank or negative control groups (P<0.05; Fig. 1B).
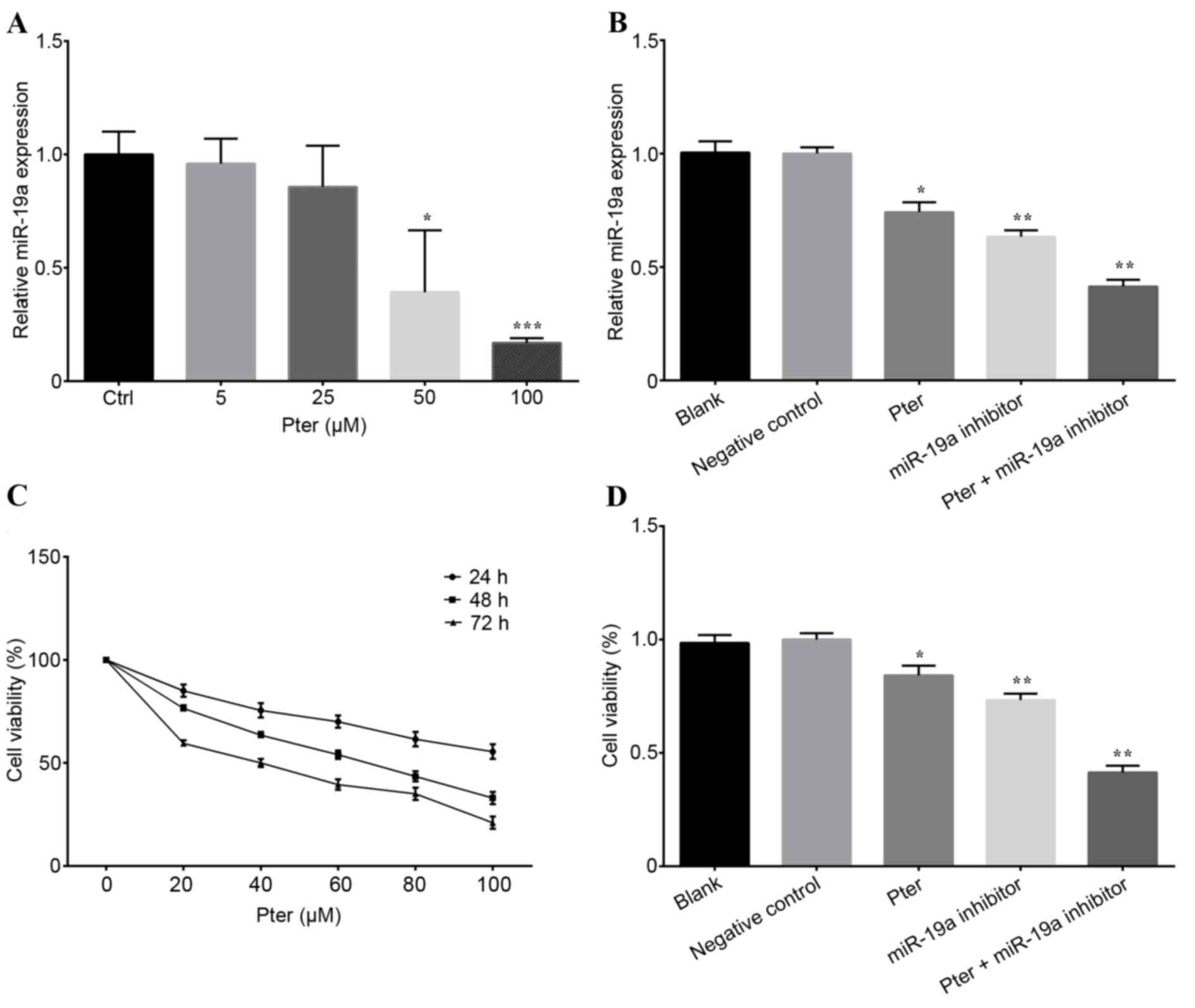 | Figure 1.Pter treatment and miR-19a inhibitor
transfection affected the miR-19a expression and cell viability.
(A) Cells were treated with Pter (0, 5, 25, 50 and 100 µM) for 24 h
and miR-19a expression was detected by RT-qPCR. (B) Relative
quantification of miR-19a expression was calculated by RT-qPCR for
SMMC-7721 cells treated with 50 µM Pter, miR-19a inhibitor, 50 µM
Pter + miR-19a inhibitor, negative control and blank. (C) An MTT
assay was performed to detect cell viability following treatment
with Pter at different concentrations (0, 20, 40, 60, 80 and 100
µM) for 24, 48 and 72 h. (D) Cells were treated with 50 µM Pter,
miR-19a inhibitor, 50 µM Pter + miR-19a inhibitor, negative control
and blank for 24 h. Cell viability was subsequently assessed by an
MTT assay. Each experiment was performed in triplicate. Data are
presented as the mean ± standard deviation. For part A, *P<0.05
and ***P<0.001 vs. Ctrl; for parts B and D, *P<0.05 and
**P<0.01 vs. blank or negative control group. The Pter-only
treatment group was compared with the blank control group, while
the miR-19a inhibitor and Pter + miR-19a inhibitor groups were
compared with the negative control group. Pter, pterostilbene; miR,
microRNA; RT-qPCR, reverse transcription-quantitative polymerase
chain reaction; Ctrl, control. |
Pter and miR-19a inhibitor treatment
represses cell proliferation
The MTT assay revealed that Pter repressed SMMC-7721
cell proliferation in a time- and dose-dependent manner (Fig. 1C). Furthermore, after 24 h, the
cell viability in the miR-19a inhibitor and Pter + miR-19a
inhibitor groups was markedly reduced compared with the blank or
negative control group (P<0.05; Fig. 1D). Additionally, a synergistic
effect was observed between Pter and the miR-19a-inhibitor. Both
Pter and miR-19a inhibitor treatments significantly repressed
SMMC-7721 cell proliferation (Fig.
1D). Pter exerted the most potent inhibitory effect on
SMMC-7721 cell proliferation.
Inhibition of miR-19a reduces cell
cycle progression, increases apoptosis and inhibits invasion
Flow cytometry was performed to analyze the cell
cycle of the cells treated with Pter. No statistically significant
differences were observed between the cells in the blank and
negative control groups in each phase (P>0.05; Fig. 2A and B). However, the Pter
treatment and miR-19a inhibitor groups demonstrated a significantly
higher number of cells in the S phase, and significantly fewer
cells in the G2/M phase, compared with the blank or negative
control groups (P<0.05; Fig. 2A and
B), indicating increased cell cycle arrest in S phase and
reduced cell mitosis following Pter treatment or miR-19a
inhibition. Furthermore, the apoptosis of cells was also analyzed;
the apoptotic rate of the blank and negative control groups were
not significantly different, but Pter treatment or miR-19a
inhibition significantly increased the apoptotic rate compared with
blank or negative control groups (P<0.05; Fig. 2C and D). Synergy between Pter
treatment and miR-19a inhibitor transfection was also observed
(Fig. 2C and D). Additionally,
cell invasion ability was significantly decreased by Pter and
miR-19a inhibitor, compared with blank or negative control groups
(Fig. 3A and B).
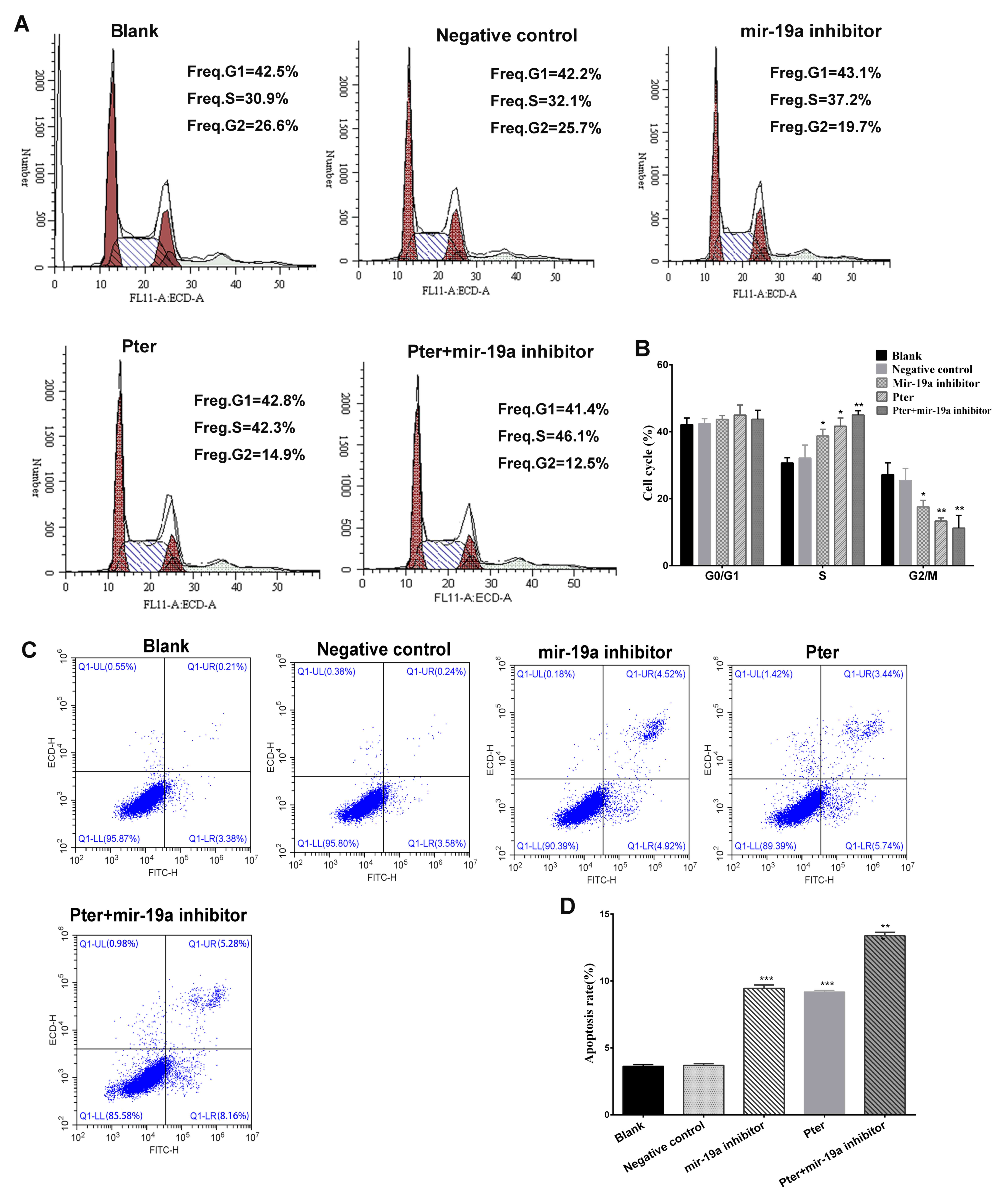 | Figure 2.Flow cytometry analysis of cell cycle
and apoptotic rate. (A) Cell cycle analysis of the Pter, miR-19a
inhibitor, Pter + miR-19a inhibitor, negative control and blank
control groups. (B) Quantification of the cell cycle analysis for
each treatment group. (C) Representative flow cytometry plots
indicating the apoptotic rate (the early and late stage apoptosis)
within Pter, miR-19a inhibitor, Pter + miR-19a inhibitor, negative
control and blank control groups. (D) Quantification of the
apoptotic rate for each treatment group. *P<0.05, **P<0.01
and ***P<0.001 vs. blank or negative control groups. The
Pter-only treatment group was compared with the blank control
group, while the miR-19a inhibitor and Pter + miR-19a inhibitor
groups were compared with the negative control group. Pter,
pterostilbene; miR, microRNA. |
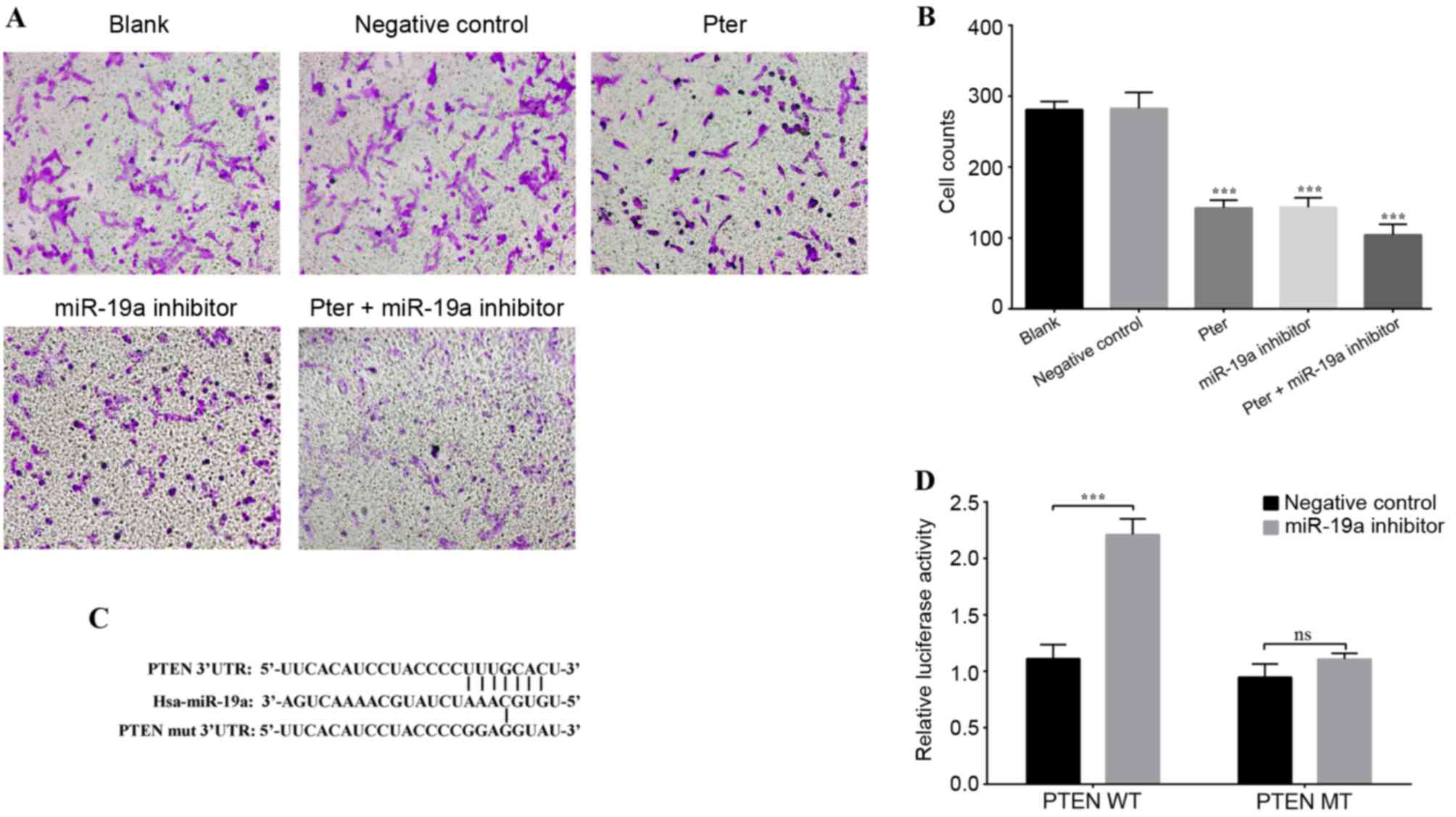 | Figure 3.Cell invasion and luciferase reporter
gene assays. (A) Representative images of the lower chamber of the
Transwell invasion assay in different treatment groups. (B) Cell
counts using a phase-contrast microscope revealed that Pter and
miR-19a inhibitor groups exhibited significantly reduced cell
invasion ability. This reduction was further enhanced in the Pter +
miR-19a group. (C) Binding sequence between miR-19a and WT PTEN
3′-UTR, and the MT binding site of the PTEN 3′-UTR. PTEN was
confirmed as a direct downstream target of miR-19a by
TargetScanHuman. (D) A luciferase reporter gene activity assay was
performed following co-transfection of PTEN 3′UTR MT/WT and miR-19a
inhibitor or negative control. miR-19a inhibited the luciferase
activity of the WT luciferase plasmid, while there was no
significant difference when the miR-19a inhibitor was
co-transfected with the MT luciferase plasmid. For part B,
***P<0.001 vs. blank or negative control groups; for part D,
***P<0.001, as indicated. In part B, the Pter-only treatment
group was compared with the blank control group, while the miR-19a
inhibitor and Pter + miR-19a inhibitor groups were compared with
the negative control group. Pter, pterostilbene; miR, microRNA; WT,
wild type; phosphatase and tensin homolog; UTR, untranslated
region; MT/mut, mutant type; ns, not significant. |
Targeted regulation of PTEN 3′UTR by
miR-19a
A luciferase reporter gene assay was performed to
verify whether PTEN may be a direct target gene of miR-19a. It was
demonstrated that the target sequence of miR-19a and PTEN 3′-UTR
matched (Fig. 3C). Therefore, the
pGL3-Luciferase vector was constructed and the PTEN 3′UTR WT
(AGCUUA) or MT (ACACGG) sequence was inserted downstream, and the
vector was subsequently co-transfected with the miR-19a inhibitor
or negative control. Following PTEN 3′-UTR-WT co-transfection with
miR-19a inhibitor, luciferase activity was significantly higher
compared with the negative control group; however, no statistically
significant difference was observed between the negative control
and miR-19a inhibitor group when co-transfected with PTEN 3′UTR-MT
(Fig. 3D). There results indicated
that the base sequence of miR-19a matched the PTEN mRNA 3′UTR and
that PTEN is a target site of miR-19a.
Pter downregulates PTEN mRNA and
protein expression through miR-19a
Initially, western blot analysis was performed to
determine PTEN protein expression following treatment with various
concentrations of Pter. The results demonstrated that Pter
increased PTEN protein expression in a dose-dependent manner
(Fig. 4A and B). Furthermore, to
investigate the association between miR-19a and PTEN expression
further, SMMC-7721 cells with low expression levels of miR-19a were
established by miR-19a inhibitor transfection. The Pter and miR-19a
inhibitor groups exhibited significantly increased expression of
PTEN mRNA compared with the blank or negative control groups,
respectively (P<0.05; Fig. 4C).
A synergistic effect was observed in the Pter + miR-19a inhibitor
group (P<0.05; Fig. 4C).
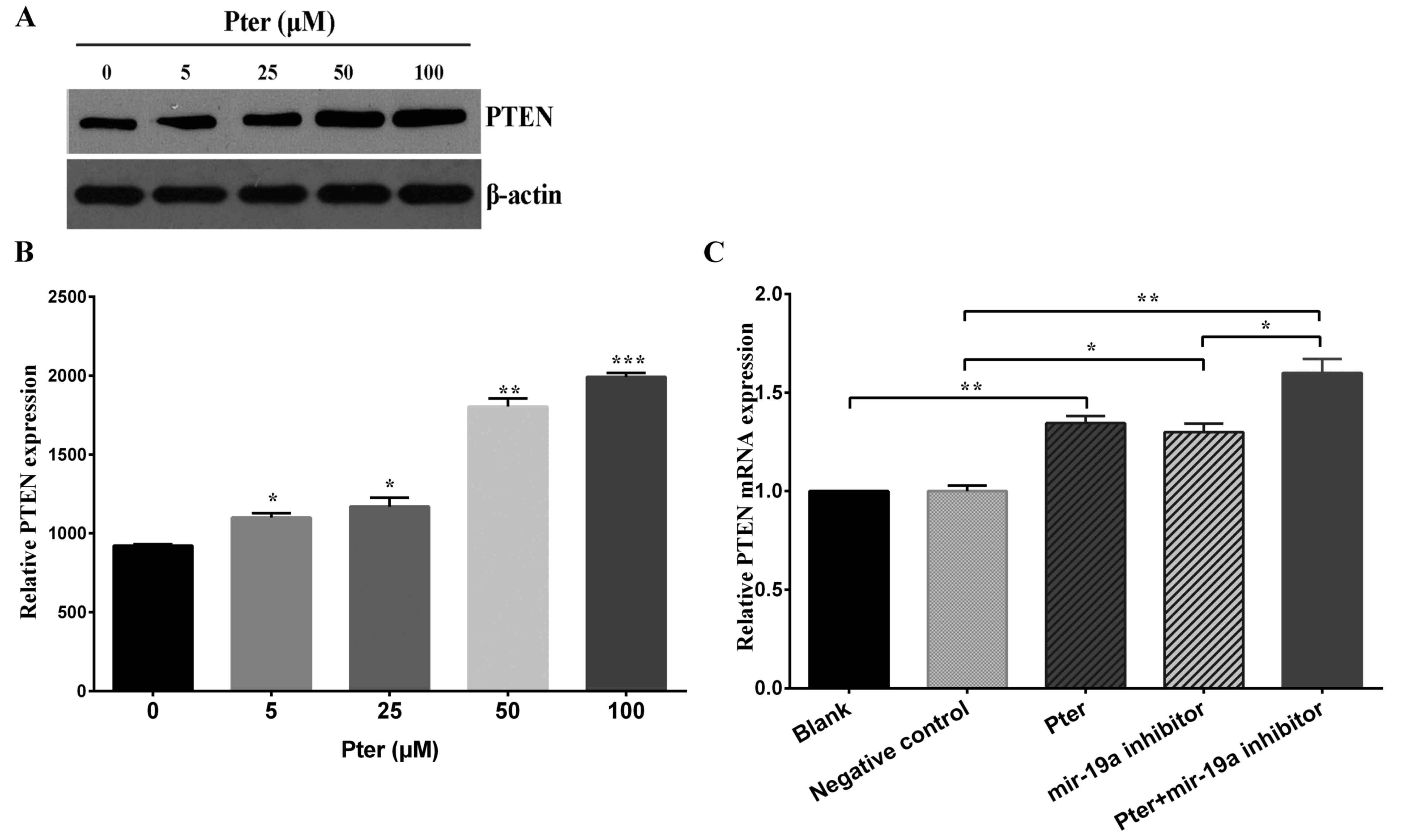 | Figure 4.Pter downregulated PTEN protein and
mRNA expression in SMMC-7721 cells. (A) SMMC-7721 cells were
treated with increasing concentrations (0, 5, 25, 50 and 100 µM) of
Pter for 24 h. The control group was treated with dimethyl
sulfoxide. PTEN protein expression was detected by western blot
analysis and representative bands are presented. (B) Western blot
analysis results were quantified with ImageJ analysis software. (C)
PTEN mRNA expression following treatment with 50 µM Pter, miR-19a
inhibitor, 50 µM Pter + miR-19a inhibitor, negative control and
blank. Cells were harvested and the relative expression of PTEN
mRNA was calculated by reverse transcription-quantitative
polymerase chain reaction. Data are presented as the mean +
standard deviation of the mean of three independent experiments.
For part B, *P<0.05, **P<0.01 and ***P<0.001 vs. 0 µM Pter
group; for part C, *P<0.05 and **P<0.01, as indicated. Pter,
pterostilbene; PTEN, phosphatase and tensin homolog; miR,
microRNA. |
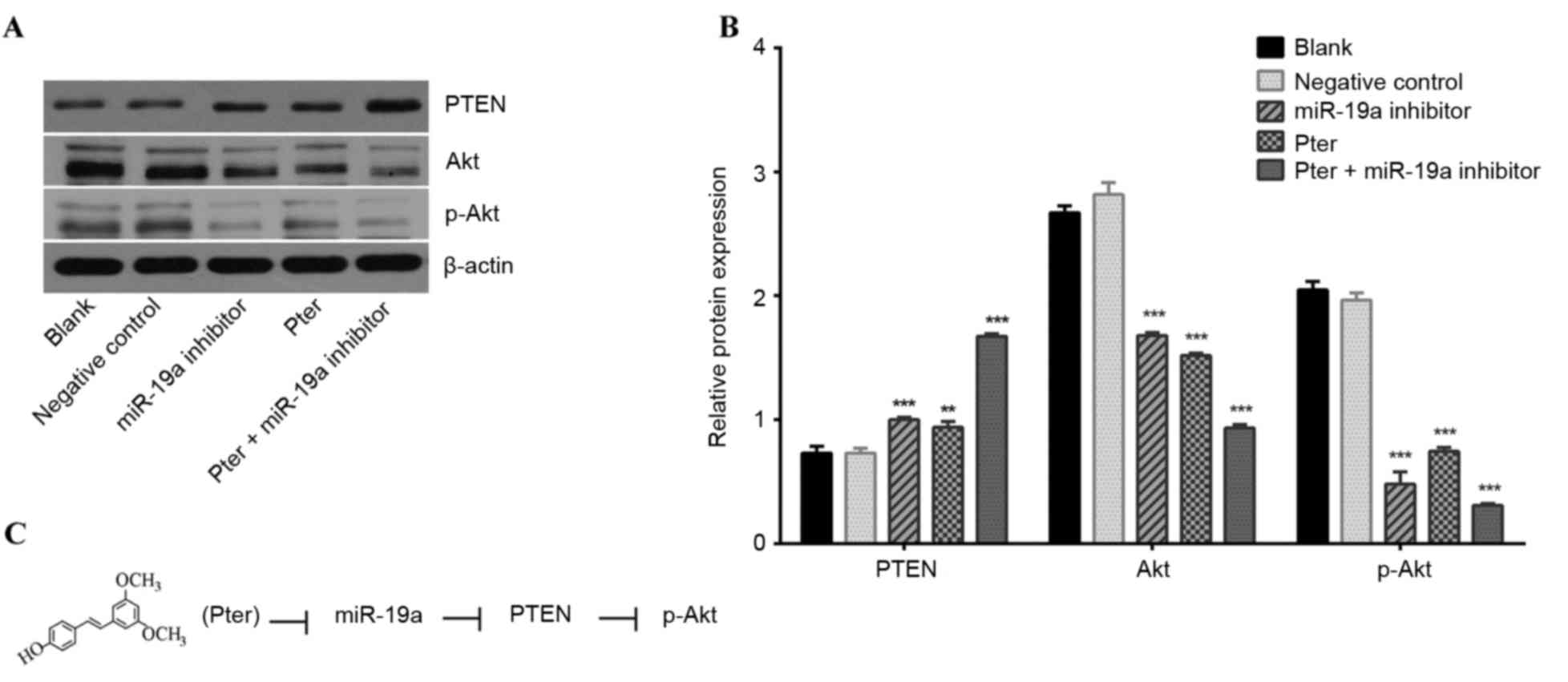
Additionally, reduced miR-19a expression induced a
marked upregulation of PTEN protein expression compared with the
negative control group (Fig. 5A and
B). These results indicated that Pter treatment may enhance
PTEN expression via the downregulation of miR-19a.
Pter treatment and miR-19a inhibitor
transfection mediates the PTEN/Akt signaling pathway
PTEN, Akt and p-Akt levels were detected by western
blot analysis. Previous research has indicated that PTEN is an
inhibitor of the PI3K/Akt pathway (28). The present study revealed a
negative association between PTEN and miR-19a expression, and a
positive association between PTEN and Akt expression, which
indicates an association between the PI3K/Akt pathway and miR-19a.
In the current study, the Pter and miR-19a inhibitor groups
exhibited significantly upregulated PTEN protein expression and
significantly reduced Akt and p-Akt levels (P<0.05; Fig. 5A and B). These results indicate
that miR-19a may mediate the downstream targets of the PTEN/Akt
signal pathway to affect the biological behavior of HCC (Fig. 5C).
Discussion
Previous research has demonstrated that miR-19a
overexpression occurs in various tumor types. Pter was reported to
inhibit the overexpression of certain oncogenic miRNAs (oncomiRs),
including miR-19a in prostate cancer cells (29). miR-19a has been previously reported
to regulate PTEN expression in various types of cancer, including
osteosarcoma (18), myeloma
(22), HCC (19) and breast cancer (23). Furthermore, the regulation of PTEN
by Pter was demonstrated to exert marked anticancer effects
(17,29). Therefore, in the present study, the
ability of Pter to increase PTEN expression through direct
downregulation of miR-19a in HCC was investigated and confirmed.
The potential molecular mechanism of this action is presented in
Fig. 5C.
An increasing number of studies have demonstrated
the anticancer properties of certain dietary agents, indicating
potential for their clinical application. The anticancer effects
exerted by several of these dietary compounds has been reported to
occur through the mediation of abnormal miRNA expression in
malignant cells (17,30,31).
Therefore, the present study aimed to confirm a direct association
between miR-19a and one of its targets, PTEN, as well as the effect
of the oncomiR on tumor cell function. The results of the current
study demonstrated that Pter may have an important role in
reversing the silencing of tumor suppressor PTEN expression by
miR-19a. Although several reports have revealed the oncogenic
potential of miR-19a in targeting the 3′UTR of PTEN mRNA in
malignant cells (15,16,32),
the present study focused on the ability of Pter to inhibit miR-19a
expression.
In the current study, low miR-19a expression,
established by transfection of a miR-19a inhibitor, induced a
significant increase in PTEN 3′UTR luciferase activity. Pter also
inhibited the expression of miR-19a in a dose-dependent manner,
indicating that Pter treatment may directly ameliorate miR-19a
overexpression. Overall, the results indicated that Pter
downregulated miR-19a expression, which led to increased PTEN
expression and subsequent PI3K/Akt signaling pathway
regulation.
PTEN has been reported to participate in processes
that are involved in basic cell function, including apoptosis,
proliferation and cell cycle arrest (33). Chang et al (34) identified that PTEN/p-Akt pathway
activation may have prognostic value in HCC. Furthermore, a lack of
PTEN was hypothesized to continuously activate signaling pathways
to promote uncontrollable cancer cell growth (35). In the present study, miR-19a
downregulation by Pter recovered PTEN expression and directly
affected the transcriptional activity of the Akt pathway.
The interaction between miR-19a and PTEN in HCC was
investigated further. It was demonstrated that PTEN was a target
gene of miR-19a through online TargetScan prediction. Increased
PTEN expression was previously detected in HCC cells transfected
with miR-19a inhibitor, which led to reduced cell growth and
invasion and an increase in the apoptotic rate (19,36).
Existing data indicates that miR-19a downregulation may reduce HCC
occurrence by increasing PTEN expression. Consistently, the present
study demonstrated that Pter treatment and miR-19a inhibitor
transfection significantly decreased miR-19a expression and
enhanced PTEN expression.
Wu et al (32) reported that malignant esophageal
cells exhibited excessive Akt expression, which was implicated in
uncontrollable cancer cell proliferation, and that PTEN repressed
the Akt activity. It was reported that miR-19a and the 3′UTR of
PTEN mRNA exhibit homology between their sequences and the
interaction of these sequences may lead to PTEN downregulation and
subsequent Akt upregulation (37).
In the present study, miR-19a inhibitor transfection led to
increased PTEN expression, which may be implicated in the cell
cycle arrest in the S phase, increased apoptosis, decreased
invasion and reduced viability that was observed in SMMC-7721 cells
following miR-19a inhibitor transfection and Pter treatment.
In conclusion, the results of the present study
demonstrated that Pter inhibited miR-19a expression, which
subsequently led to regulation of the PTEN/Akt pathway, reduced
cell viability, cell cycle arrest, apoptosis promotion and cell
invasion inhibition in HCC cells. The present study may provide a
foundation for future research, as miR-19a may have potential as a
molecular HCC marker and Pter may be developed as a therapy for
patients with HCC.
References
|
1
|
Schulze K, Imbeaud S, Letouzé E,
Alexandrov LB, Calderaro J, Rebouissou S, Couchy G, Meiller C,
Shinde J, Soysouvanh F, et al: Exome sequencing of hepatocellular
carcinomas identifies new mutational signatures and potential
therapeutic targets. Nat Genet. 47:505–511. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhang B, Pan X, Cobb GP and Anderson TA:
microRNAs as oncogenes and tumor suppressors. Dev Biol. 302:1–12.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Urtasun R, Elizalde M, Azkona M, Latasa
MU, García-Irigoyen O, Uriarte I, Fernández-Barrena MG, Vicent S,
Alonso MM, Muntané J, et al: Splicing regulator SLU7 preserves
survival of hepatocellular carcinoma cells and other solid tumors
via oncogenic miR-17-92 cluster expression. Oncogene. 35:4719–4729.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhu H, Han C and Wu T: MiR-17-92 cluster
promotes hepatocarcinogenesis. Carcinogenesis. 36:1213–1222. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Filipowicz W, Bhattacharyya SN and
Sonenberg N: Mechanisms of post-transcriptional regulation by
microRNAs: Are the answers in sight? Nat Rev Genet. 9:102–114.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sapre N and Selth LA: Circulating
MicroRNAs as biomarkers of prostate cancer: The state of play.
Prostate Cancer. 2013:5396802013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li L, Song W, Yan X, Li A, Zhang X, Li W,
Wen X, Zhou L, Yu D, Hu JF and Cui J: Friend leukemia virus
integration 1 promotes tumorigenesis of small cell lung cancer
cells by activating the miR-17-92 pathway. Oncotarget.
8:41975–41987. 2017.PubMed/NCBI
|
|
8
|
Dhar S, Hicks C and Levenson AS:
Resveratrol and prostate cancer: Promising role for microRNAs. Mol
Nutr Food Res. 55:1219–1229. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu F, Zhang F, Li X, Liu Q, Liu W, Song
P, Qiu Z, Dong Y and Xiang H: Prognostic role of miR-17-92 family
in human cancers: Evaluation of multiple prognostic outcomes.
Oncotarget. 8:69125–69138. 2017.PubMed/NCBI
|
|
10
|
Strickertsson JA, Rasmussen LJ and
Friis-Hansen L: Enterococcus faecalis infection and reactive oxygen
species Down-regulates the miR-17-92 cluster in gastric
adenocarcinoma cell culture. Genes (Basel). 5:726–738. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Knudsen KN, Nielsen BS, Lindebjerg J,
Hansen TF, Holst R and Sorensen FB: microRNA-17 Is the Most
Up-Regulated member of the miR-17-92 cluster during early colon
cancer evolution. PLoS One. 10:e01405032015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zou P, Ding J and Fu S: Elevated
expression of microRNA-19a predicts a poor prognosis in patients
with osteosarcoma. Pathol Res Pract. 213:194–198. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhou B, Wang J, Zheng G and Qiu Z:
Methylated urolithin A, the modified ellagitannin-derived
metabolite, suppresses cell viability of DU145 human prostate
cancer cells via targeting miR-21. Food Chem Toxicol. 97:375–384.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Song L, Liu S, Zhang L, Yao H, Gao F, Xu D
and Li Q: MiR-21 modulates radiosensitivity of cervical cancer
through inhibiting autophagy via the PTEN/Akt/HIF-1α feedback loop
and the Akt-mTOR signaling pathway. Tumour Biol. 37:12161–12168.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li C, Song L, Zhang Z, Bai XX, Cui MF and
Ma LJ: MicroRNA-21 promotes TGF-β1-induced epithelial-mesenchymal
transition in gastric cancer through up-regulating PTEN expression.
Oncotarget. 7:66989–67003. 2016.PubMed/NCBI
|
|
16
|
Fang H, Xie J, Zhang M, Zhao Z, Wan Y and
Yao Y: miRNA-21 promotes proliferation and invasion of
triple-negative breast cancer cells through targeting PTEN. Am J
Transl Res. 9:953–961. 2017.PubMed/NCBI
|
|
17
|
Dhar S, Kumar A, Rimando AM, Zhang X and
Levenson AS: Resveratrol and pterostilbene epigenetically restore
PTEN expression by targeting oncomiRs of the miR-17 family in
prostate cancer. Oncotarget. 6:27214–27226. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhao D, Chen Y, Chen S, Zheng C, Hu J and
Luo S: MiR-19a regulates the cell growth and apoptosis of
osteosarcoma stem cells by targeting PTEN. Tumour Biol.
39:10104283177053412017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yu G, Chen X, Chen S, Ye W, Hou K and
Liang M: MiR-19a, miR-122 and miR-223 are differentially regulated
by hepatitis B virus X protein and involve in cell proliferation in
hepatoma cells. J Transl Med. 14:1222016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ma Q, Peng Z, Wang L, Li Y, Wang K, Zheng
J, Liang Z and Liu T: miR-19a correlates with poor prognosis of
clear cell renal cell carcinoma patients via promoting cell
proliferation and suppressing PTEN/SMAD4 expression. Int J Oncol.
49:2589–2599. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu K, Liu S, Zhang W, Jia B, Tan L, Jin Z
and Liu Y: miR-494 promotes cell proliferation, migration and
invasion, and increased sorafenib resistance in hepatocellular
carcinoma by targeting PTEN. Oncol Rep. 34:1003–1010. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang X, Chen Y, Zhao P, Zang L, Zhang Z
and Wang X: MicroRNA-19a functions as an oncogene by regulating
PTEN/AKT/pAKT pathway in myeloma. Leuk Lymphoma. 58:932–940. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li X, Xie W, Xie C, Huang C, Zhu J, Liang
Z, Deng F, Zhu M, Zhu W, Wu R, et al: Curcumin modulates
miR-19/PTEN/AKT/p53 axis to suppress bisphenol A-induced MCF-7
breast cancer cell proliferation. Phytother Res. 28:1553–1560.
2014. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Aggarwal BB, Bhardwaj A, Aggarwal RS,
Seeram NP, Shishodia S and Takada Y: Role of resveratrol in
prevention and therapy of cancer: Preclinical and clinical studies.
Anticancer Res. 24:2783–2840. 2004.PubMed/NCBI
|
|
25
|
Wang G, Dai F, Yu K, Jia Z, Zhang A, Huang
Q, Kang C, Jiang H and Pu P: Resveratrol inhibits glioma cell
growth via targeting oncogenic microRNAs and multiple signaling
pathways. Int J Oncol. 46:1739–1747. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Dhar S, Kumar A, Li K, Tzivion G and
Levenson AS: Resveratrol regulates PTEN/Akt pathway through
inhibition of MTA1/HDAC unit of the NuRD complex in prostate
cancer. Biochim Biophys Acta. 1853:265–275. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Luo H, Yang Y, Duan J, Wu P, Jiang Q and
Xu C: PTEN-regulated AKT/FoxO3a/Bim signaling contributes to
reactive oxygen species-mediated apoptosis in selenite-treated
colorectal cancer cells. Cell Death Dis. 4:e4812013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kumar A, Rimando AM and Levenson AS:
Resveratrol and pterostilbene as a microRNA-mediated
chemopreventive and therapeutic strategy in prostate cancer. Ann N
Y Acad Sci. 1403:15–26. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li Y, Kong D, Wang Z and Sarkar FH:
Regulation of microRNAs by natural agents: An emerging field in
chemoprevention and chemotherapy research. Pharm Res. 27:1027–1041.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Milenkovic D, Jude B and Morand C: miRNA
as molecular target of polyphenols underlying their biological
effects. Free Radic Biol Med. 64:40–51. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wu YR, Qi HJ, Deng DF, Luo YY and Yang SL:
MicroRNA-21 promotes cell proliferation, migration, and resistance
to apoptosis through PTEN/PI3K/AKT signaling pathway in esophageal
cancer. Tumour Biol. 37:12061–12070. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang G, Li Y, Wang P, Liang H, Cui M, Zhu
M, Guo L, Su Q, Sun Y, McNutt MA and Yin Y: PTEN regulates RPA1 and
protects DNA replication forks. Cell Res. 25:1189–1204. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chang RM, Xu JF, Fang F, Yang H and Yang
LY: MicroRNA-130b promotes proliferation and EMT-induced metastasis
via PTEN/p-AKT/HIF-1α signaling. Tumour Biol. 37:10609–10619. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li P, Mao WM, Zheng ZG, Dong ZM and Ling
ZQ: Down-regulation of PTEN expression modulated by dysregulated
miR-21 contributes to the progression of esophageal cancer. Dig Dis
Sci. 58:3483–3493. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Meng F, Henson R, Wehbe-Janek H, Ghoshal
K, Jacob ST and Patel T: MicroRNA-21 regulates expression of the
PTEN tumor suppressor gene in human hepatocellular cancer.
Gastroenterology. 133:647–658. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yang H, Kong W, He L, Zhao JJ, O'Donnell
JD, Wang J, Wenham RM, Coppola D, Kruk PA, Nicosia SV and Cheng JQ:
MicroRNA expression profiling in human ovarian cancer: miR-214
induces cell survival and cisplatin resistance by targeting PTEN.
Cancer Res. 68:425–433. 2008. View Article : Google Scholar : PubMed/NCBI
|