Introduction
A cataract is defined as any type of opacity of the
eye lens that affects vision (1).
Cataracts are the main cause of blindness and visual impairment
worldwide, particularly in developing countries (2,3).
There are a number of risk factors for the development of
cataracts, including ultraviolet light exposure, diabetes, aging
and other environmental factors (4–6).
Despite the number of studies that have been conducted on
cataracts, their pathogenesis remains unknown. Thus, effective
non-surgical therapies are currently lacking, which is a
substantial burden to healthcare budget (7). Dysfunction of lens epithelial cells
(LECs) has been repeatedly proposed to serve a key role in cataract
formation, although this mechanism is unknown (8–10).
Laminins (LMs) are large-molecular-weight trimer
glycoproteins consisting of α-, β- and γ-subunits, which integrate
in almost all basement membranes (BMs), functioning as scaffolds
(11). It has been suggested that
LM trimers are assembled inside the cell, with extracellular
proteolytic processing of various subunits leading to the mature
LMs (12). In addition to their
function as scaffold of BMs, LMs can also interact with cell
surface receptors such as intergrins to control signaling events
that regulate cellular proliferation, migration and differentiation
(13,14).
Particular LM isoforms are present in specific
tissue and cell distributions during developmental and pathological
conditions, indicating that different LM isoforms have distinct
functional roles (12,14). Past studies have demonstrated that
LMα1, LMα5, LMβ1, LMβ2 and LMγ1 subunits exist in the developmental
human lens capsule (15,16). Another prior study demonstrated
that human adult lens capsule is composed of LMα1, LMα2, LMα3,
LMα4, LMα5, LMβ1, LMβ2 and LMγ1 subunits, and the predominant LM
trimer was LM521 (17).
Gene defects and the abnormal expression of LMs are
closely associated with various lens diseases (18–23).
LMβ2 mutations cause Pierson syndrome, which has various ocular
lesions, including abnormal lens shape, posterior lenticonus and
cataracts (19). It has been
demonstrated that development of LM-positive linear capsular
densities and occurrence lens epithelial degenerative alterations
are associated with aging (20).
Increased LM expression underneath lens capsules and abnormal LEC
migration were identified in FE65/FE65L1 double knockout lenses,
which may be involved in the formation of cortical cataracts
(21). Abnormal accumulation of
LMs in the lens capsule and epithelial-mesenchymal transition of
LECs was also observed in patients with anterior subcapsular
opacification and posterior capsule opacification (22,23).
Human primary LECs in long-term cultures could
synthesize a continuous sheet of capsule-like material (24). Theoretically, human LEC lines could
also produce anterior-capsule-like BM with rich LMs in
vitro, which could be a suitable disease model to study the
role of LMs in anterior lens capsule (ALC) in the pathogenesis of
lens diseases, particularly cataracts.
In the present study, the human LEC HLE B-3 line was
used to construct the BMs and further analyze the expression levels
of 11 LM subunits. Similarities in LM expression between HLE B-3
BMs and human ALCs were also evaluated. The results of the present
study may aid in elucidating the role of LMs in ALC and cataract
pathogenesis.
Materials and methods
HLE B-3 cell culture and preparation
of cell lysate and culture supernatant
The human LEC HLE B-3 line were obtained from the
American Type Culture Collection (Manassas, VA, USA) and cultured
in low-glucose DMEM (Hyclone; GE Healthcare Life Sciences, Logan,
UT, USA) containing 10% fetal bovine serum (FBS) (Gibco; Thermo
Fisher Scientific, Inc., Waltham, MA, USA), 100 U/ml penicillin and
100 µg/ml streptomycin (Beyotime Institute of Biotechnology,
Haimen, China) at 37°C in humidified atmosphere with 5%
CO2. Cells were seeded in 10-cm dishes and grown to 95%
confluence. Following discarding the culture supernatant, cells
were washed twice with PBS and lysed with cold
radioimmunoprecipitation assay (RIPA) buffer [50 mM Tris-HCl (pH
7.5), 150 mM NaCl, 1% Triton X-100 and 1% protease inhibitor
cocktail from Sigma-Aldrich; Merck KGaA, Darmstadt, Germany] at 4°C
for 30 min. Following centrifugation at 14,000 × g at 4°C for 20
min, the supernatant was harvested and kept as cell lysate. When
95% confluent, cells were washed twice with PBS and cultured in 5
ml low-glucose DMEM without FBS for 48 h. The cell supernatant was
harvested and centrifuged at 3,000 × g at 4°C for 5 min to remove
debris.
Immunocytochemical staining (ICC) of
HLE B-3 cells
For hematoxylin and eosin (H&E) staining, HLE
B-3 cells grown on glass slides were stained with hematoxylin and
eosin (5 and 2 min, respectively, at room temperature), and
examined under a light microscope (Olympus Corporation, Tokyo,
Japan; magnification, ×400). For ICC of LMs, HLE B-3 cells grown on
glass slides were incubated for 10 min at room temperature with 3%
hydrogen peroxide, and blocked with 10% normal goat serum for 20
min at room temperature, followed by incubation for 60 min at room
temperature with rabbit anti-LM antibody (1:200; cat. no. ab11575;
Abcam, Cambridge, UK). Subsequently, samples were incubated for 30
min at room temperture with peroxidase conjugated anti-rabbit
antibody (cat. no. PV-9000; OriGene Technologies, Inc., Beijing,
China) without dilution and color development was performed with
3,3′-diaminobenzidine (DAB). Staining was visualized with a light
microscope (Nikon TE300, Nikon Corporation, Tokyo, Japan;
magnification, ×400), and images were captured with an attached
digital camera and associated software (SPOT Basic™ image capture
software; cat. no. SPOT53BE; SPOT Imaging, a division of Diagnostic
Instruments, Inc., Sterling Heights, MI, USA).
BM preparation using HLE B-3
cells
When HLE B-3 cells reached 95% confluence, they were
washed with PBS and cultured in fresh low-glucose DMEM medium with
5% FBS; this day was recorded as day 0. At days 0, 3, 6, 9 and 12,
HLE B-3 BMs were harvested, as described previously (25,26).
Briefly, HLE B-3 cells were washed with PBS four times, after which
0.03% ammonia and 0.1% Triton-PBS was added to remove HLE B-3 cells
completely. Finally, HLE B-3 BM remained on the bottom of the dish,
which was washed with double distilled water 4 times and harvested
with 150 µl of 4X sample buffer [8% sodium dodecyl sulfate, 0.2 M
Tris-Cl (pH 6.8), 40% glycerol, 0.2% bromophenol blue and 0.1 M
dithiothreitol] at room temperature or 1 ml RIPA buffer on ice.
Human ALCs and protein lysates
Human ALCs were obtained from autopsies from 3
individuals (1 female and 2 male) aged 25–79 years old from
November to December of 2016 at the Eye Bank of Heilongjiang
Province (Harbin, China). Human ALCs were obtained by a single
ophthalmologist using central circular capsulorhexis method from
anterior surface of transparent lens (27). All experiments were performed with
the approval of the Internal Review Board of Harbin Medical
University and were conducted in accordance with Declaration of
Helsinki Principles. Informed consent was obtained from all
patients prior to mortality or from their families following
mortality for inclusion of autopsy data. Samples were harvested
within 4–24 h of mortality. Human ALCs were dissected from the
isolated transparent lenses and were lysed with cold RIPA buffer at
4°C overnight. Finally, the supernatant was kept as human ALC
lysate following centrifugation at 14,000 × g at 4°C for 20
min.
H&E staining and
immunohistochemistry (IHC) staining of human ALCs
Following careful dissection from transparent
lenses, excised human ALCs were frozen at −80°C. Frozen human ALC
tissues were transversely sectioned at 5-µm thickness, mounted on
glass slides, fixed with cold acetone for 15 min at 4°C, followed
by H&E staining or IHC with identical steps to ICC, as
aforementioned.
Bicinchoninic acid assay
Protein concentrations in the HLE B-3 cell lysate,
HLE B-3 cell culture supernatant, human ALC lysate and various HLE
B-3 BMs were measured using a Bicinchoninic Acid Assay kit
(Beyotime Institute of Biotechnology) according to the
manufacturer's protocol.
ELISA
The total LM levels were measured by using
commercially available LM ELISA kit in accordance with the
manufacturer's protocol (E-EL-H0128c; Elabscience, Houston, TX,
USA). The antibodies used in this kit were polyclonal antibodies
(pAbs) against all types of LMα, LMβ and LMγ subunits.
Antibodies
Goat pAbs against LMα5 and LMγ3 (1:200, cat nos.
sc-16592 and sc-133178; Santa Cruz Biotechnology, Inc., Dallas, TX,
USA), rabbit pAbs against LMα4, LMβ3 and LMγ2 (1:1,000, cat nos.
C13067, C13071 and C30224; Assay Biotechnology Company, San
Francisco, California), rabbit pAbs against LMγ1 (1:1,000, cat nos.
ab69632; Abcam, Cambridge, UK), mouse monoclonal antibodies (mAb)
against LMα4, LMα3, LMα2, LMα1, LMβ2 and LMβ1 (1:200, cat nos.
sc-130540, SC-13586, sc-55605, sc-74418, sc-133241 and sc-17763;
Santa Cruz Biotechnology, Inc.) were used in the present study.
Western blotting analysis
Five known LMα chains, three known LMβ chains and
three known LMγ chains in HLE B-3 cell lysate, HLE B-3 cell culture
supernatant and HLE B-3 BMs harvested at days 0, 3, 6, 9 and 12,
and human ALC lysate were analyzed by western blotting, as
described previously (28).
Briefly, antigen sources inclding HLE B-3 cell lysate, culture
supernatant or protein lysate of human ALCs were mixed with 2X
sample buffer, boiled for 2 min and separated by SDS-PAGE. The
separated proteins were transferred to a polyvinylidene difluoride
membrane (EMD Millipore, Billerica, MA, USA). Following blocking
with 5% skimmed milk in Tris-buffered saline containing 0.05%
Tween-20 (TBS-T) for 1 h at room temperature, membranes were
incubated with the aforementioned primary antibodies diluted in
solution 1 (Toyobo Life Science, Osaka, Japan) at 4°C overnight.
Following washes with TBS-T, membranes were incubated with
HRP-conjugated goat anti-mouse IgG (1:5,000; cat no. 31437; Thermo
Fisher Scientific, Inc.), HRP-conjugated rabbit anti-goat IgG
(1:5,000; cat no. 31433; Thermo Fisher Scientific, Inc.) or
HRP-conjugated goat anti-rabbit IgG (1:5,000; cat no. 31463; Thermo
Fisher Scientific, Inc.) diluted in solution 2 (Toyobo Life
Science) for 1 h at room temperature. Under identical experimental
conditions, normal rabbit IgG, normal mouse IgG or normal goat IgG
(all 1:200; cat nos. sc-2027, sc-2025 and sc-2028; Santa Cruz
Biotechnology, Inc.) were used as isotype controls. Finally, the
antibody-antigen complex was visualized using an Enhanced
Chemiluminescent kit (Beyotime Institute of Biotechnology).
Immunoprecipitation (IP)-western blot
analysis
All subsequent procedures were performed at 4°C,
unless otherwise stated. A total of 1 ml HLE B-3 cell culture
supernatant was incubated overnight on a shaker with protein G
agarose (Beyotime Institute of Biotechnology) and anti-LMα4 rabbit
pAb or anti-LMγ1 rabbit pAb. The same concentrations of normal
rabbit IgG were used as an isotype control for IP. Precipitates,
collected by centrifugation at 3,000 × g at 4°C for 2 min, were
washed three times with PBS containing 0.5% Triton X-100, and were
eluted from protein G agarose by boiling with 4X sample buffer for
2 min. Proteins in the supernatant were separated by SDS-PAGE and
detected by western blotting, as aforementioned.
Results
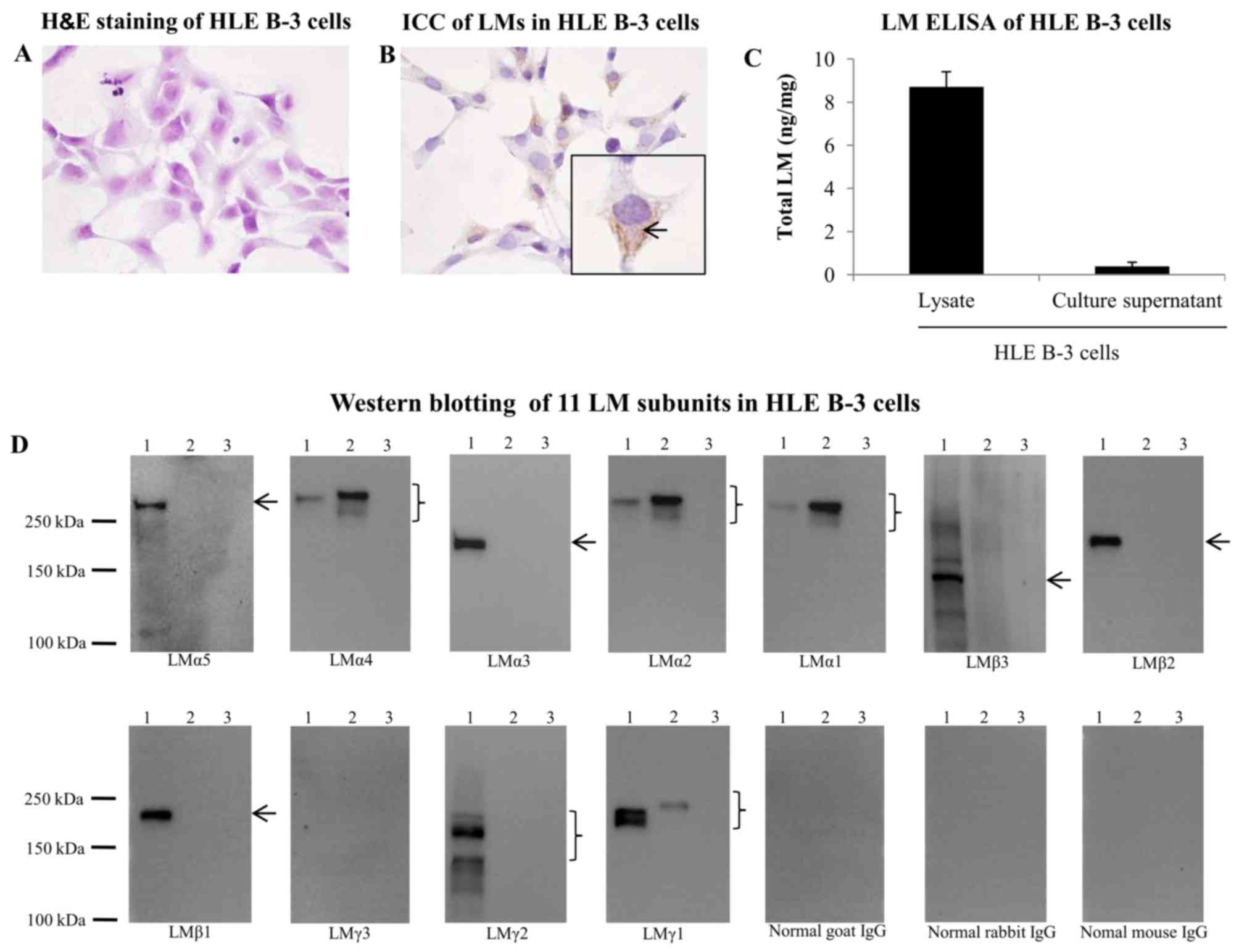
LMs in HLE B-3 cells
To investigate whether HLE B-3 cells were positive
for LMs, H&E staining and ICC for LMs were performed. As
depicted in Fig. 1A and B, LMs
were widely expressed in the cytoplasm of HLE B-3 cells. To
investigate the expression of LMs in HLE B-3 cell lysate and
culture supernatant, ELISA and western blot analysis was performed.
ELISA for the presence of LMs generated positive results in HLE B-3
cell lysate and culture supernatant (Fig. 1C). By western blotting, all 11 LM
subunits were probed in the HLE B-3 cell lysate and culture
supernatant. The HLE B-3 cell lysate was positive for LMα5, LMα4,
LMα3, LMα2, LMα1, LMβ3, LMβ2, LMβ1, LMγ2 and LMγ1 subunits, whereas
HLE B-3 cell culture supernatant was positive for LMα4, LMα2, LMα1
and LMγ1 subunits (Fig. 1D). The
10-fold concentrated HLE B-3 cell culture supernatant was also used
for detection of 11 LM subunits by western blotting, and the
results were identical to those obtained using the non-concentrated
supernatant (data not shown).
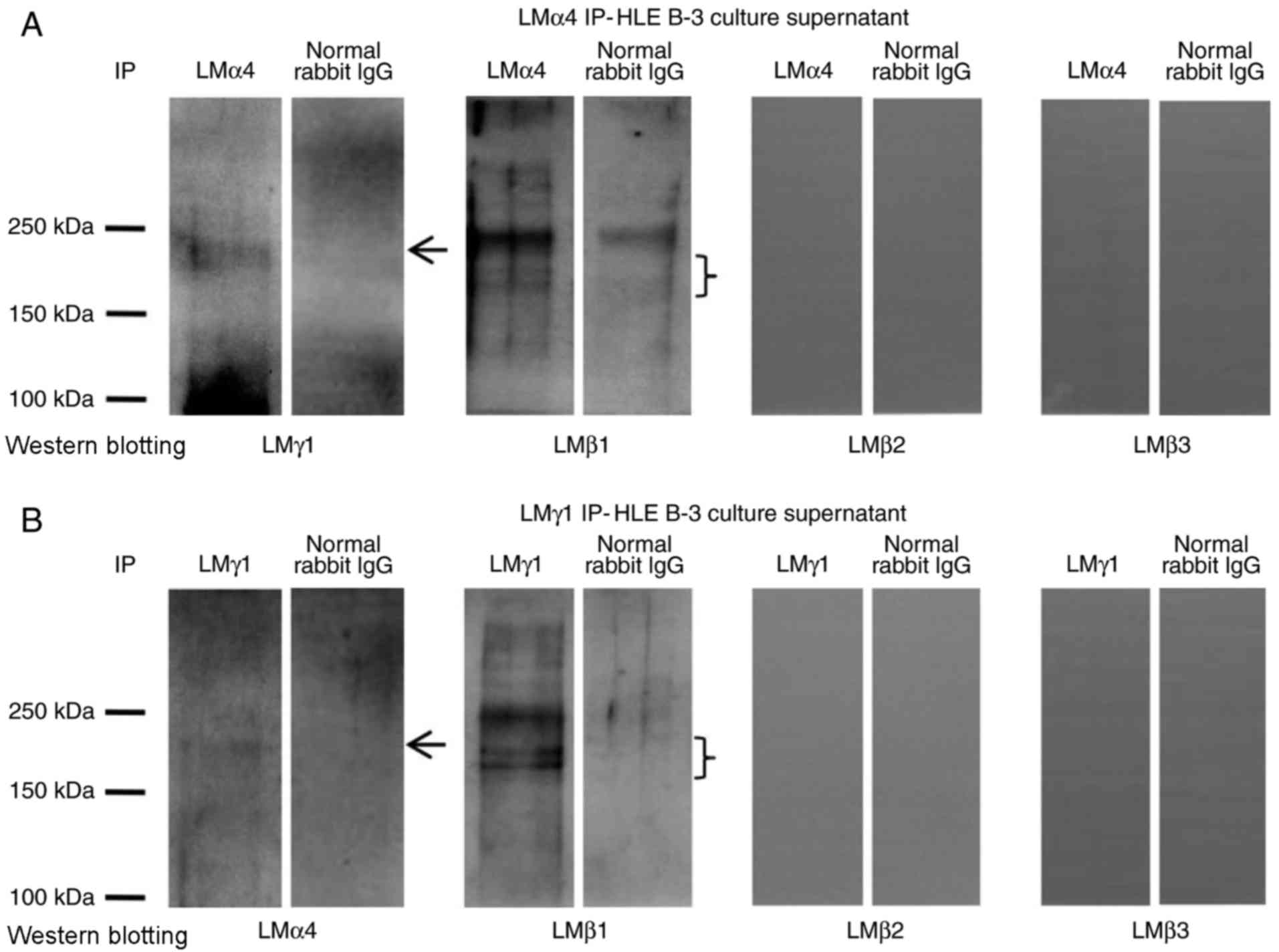
LM trimers in HLE B-3 cell culture
supernatant
To investigate what type of LM trimers appeared in
HLE B-3 cell culture supernatant, IP experiments with antibodies
against LMα1-LMα5 for IP, and antibodies against LMβ1-LMβ3 and
LMγ1-LMγ3 for western blotting were performed. The
immunoprecipitate obtained with antibodies for LMα1, LMα2, LMα3 and
LMα5 were negative in western blotting with antibodies for all LMβ
and LMγ subunits (data not shown). The immunoprecipitate obtained
with antibodies for LMα4 was positive in western blotting with
antibodies for LMβ1 and LMγ1 (Fig.
2A), indicating the presence of the LM411 trimer in the HLE B-3
cell culture supernatant. To confirm this result, the
immunoprecipitate obtained with antibodies for LMγ1 was also
analyzed by western blotting using antibodies against LMα4 and
LMβ1, which proved that LMγ1 immunoprecipitates contained LMα4 and
LMβ1 (Fig. 2B). These results
confirmed that the LM411 trimer existed in the HLE B-3 cell culture
supernatant.
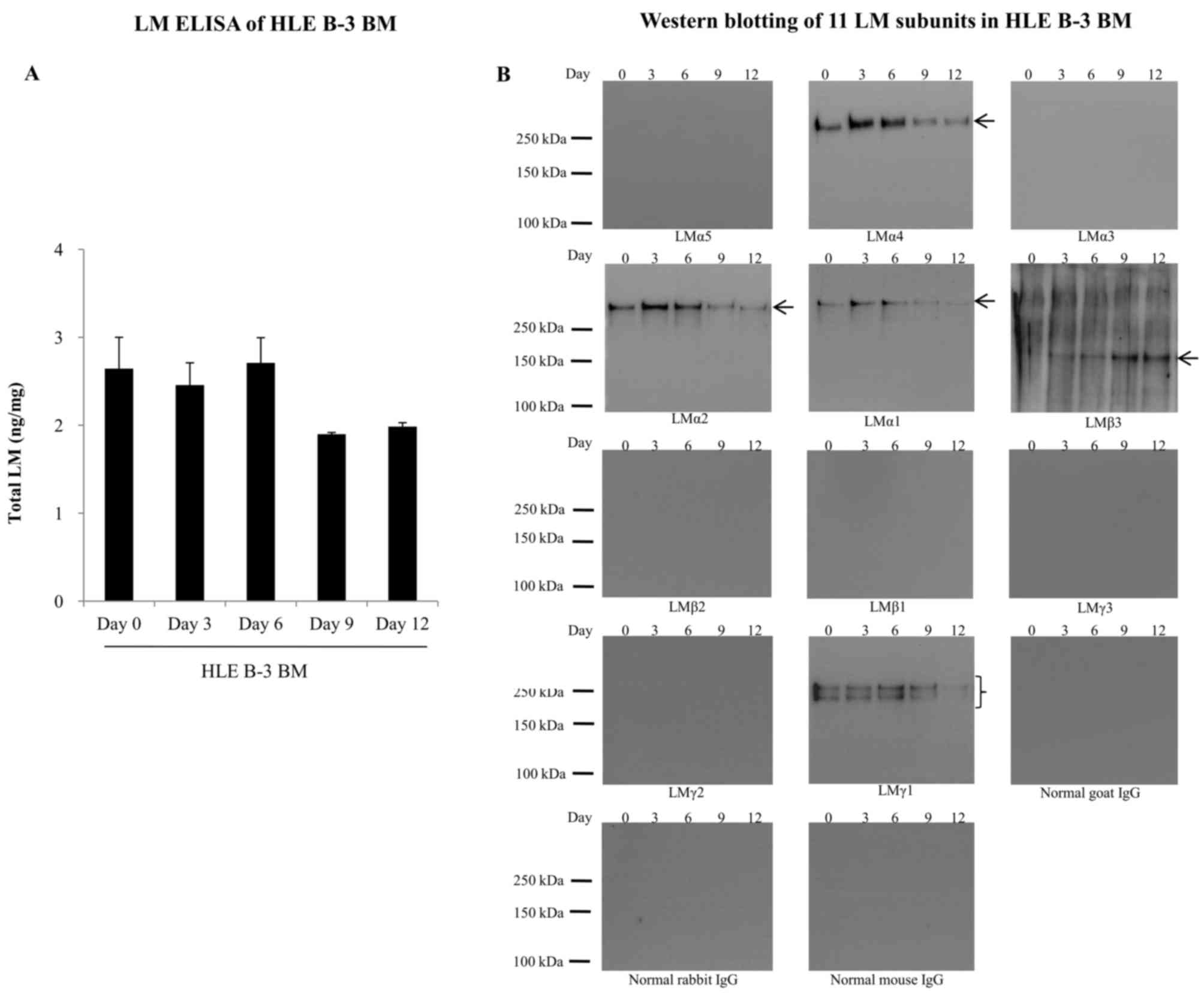
LMs in HLE B-3 BMs
HLE B-3 BMs were harvested at days 0, 3, 6, 9 and
12, and were analyzed for the expression of LMs by ELISA and
western blotting. In LM ELISA, total LM levels in HLE B-3 BMs were
similar at days 0, 3 and 6, but decreased to 70% at days 9 and 12
(Fig. 3A). Western blotting of HLE
B-3 BMs detected the presence of LMα4, LMα2, LMα1, LMβ3 and LMγ1
subunits (Fig. 3B). Expression
levels of LMα4, LMα2 and LMα1 in HLE B-3 BMs were relatively higher
at days 3 and 6. LMβ3 expression levels increased with culture
time. LMγ1 expression levels fluctuated with time irregularly, with
the lowest levels at day 12.
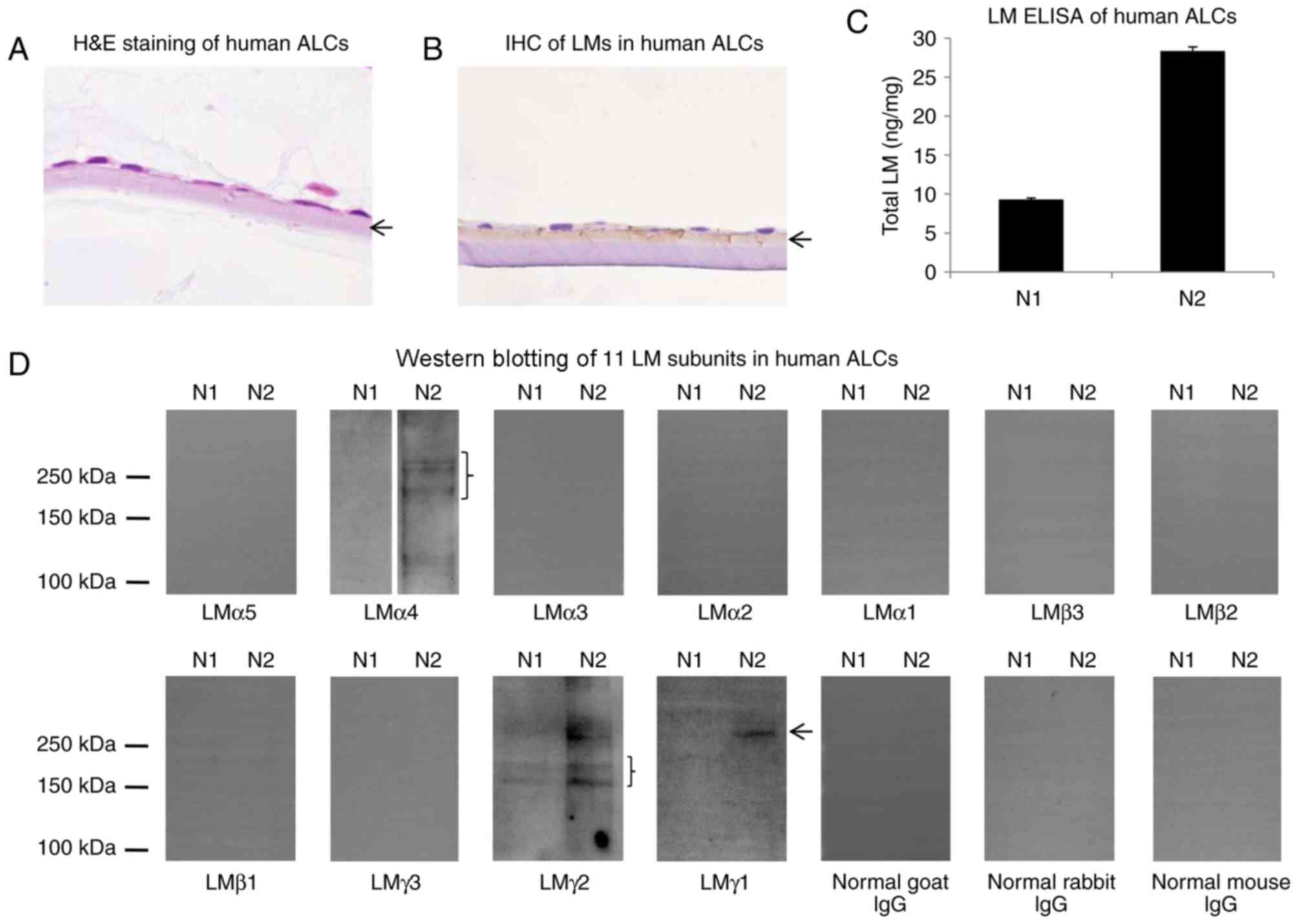
LMs in human ALCs
To investigate LM distribution in human ALCs,
H&E staining and IHC for LMs was performed. As depicted in
Fig. 4A and B, LMs were primarily
distributed in the ALC layer, close to the lens epithelial cells.
To evaluate LM expression levels in human ALC, extracts from 2
human ALCs were examined by ELISA and western blotting with
antibodies against all 11 LM subunits. In LM ELISA, total LM levels
in two human ALCs (N1 and N2 samples) differed by ~3-fold (Fig. 4C), which may have resulted from the
differences on age or gender. In western blotting, the N1 sample
was weakly positive for LMγ2 and negative for the remaining 10 LM
subunits, whereas the N2 sample was positive for LMα4, LMγ2 and
LMγ1 subunits and negative for the remaining eight LM subunits
(Fig. 4D).
Discussion
The present study provided novel information on the
LM composition in HLE B-3 cells, HLE B-3 BMs and human ALCs. The
LM411 trimer existed in the HLE B-3 cell culture supernatant.
Furthermore, the results of the present study clearly demonstrated
that the expression pattern of LM in ALC samples was similar to the
expression of BM in HLE B-3.
It has been reported that the LMγ1 subunit was
present in the HLE B-3 cell lysate (29). However, whether other LM subunits
exist in HLE B-3 cells remains unknown. The results of the present
study demonstrated that other than the LMγ1 subunit, nine other LM
subunits, including LMα5, LMα4, LMα3, LMα2, LMα1, LMβ3, LMβ2, LMβ1
and LMγ2 subunits, were also present in the HLE B-3 cell lysate.
Compared with that in the cell lysate, the total LM level in the
HLE B-3 cell culture supernatant was lower, and only 4 LM subunits,
LMα4, LMα2, LMα1 and LMγ1, were detected by western blot analysis.
The presence of the LM411 trimer in the HLE B-3 cell culture
supernatant indicated that LMβ1 should also be expressed in the HLE
B-3 cell culture supernatant. In addition, all LMs in the culture
supernatant of HLE B-3 might be involved in the formation of the
cell BM (30,31).
In the present study, it was also demonstrated that
HLE B-3 cells could synthesize BMs rich in LMs for a short period
(0–6 days following 95% confluence). HLE B-3 BMs contained LMα4,
LMα2, LMα1, LMβ3 and LMγ1 subunits, which were similar to LM
subunits in the HLE B-3 cell culture supernatant, except for LMβ3
and LMβ1. Considering the results of the various detection methods
for the LM subunits in HLE B-3 cell lysate, culture supernatant and
BMs, the LM411 trimer seems to be the major LM trimer in HLE B-3
BMs, although LMβ1 was not detected by western blot analysis.
Previous ICC studies have demonstrated that LMs are
a major component of LECs and ALCs (22,32),
results that are consistent with the findings of the present study.
Studies focusing on LM subunits in human ALCs are rare; the present
study provided evidence that 3 LM subunits (LMα4, LMγ2 and LMγ1)
could be detected in human ALCs by western blotting. In the present
study, HLE B-3 BM was positive for LMα4, LMα2, LMα1, LMβ3 and LMγ1
subunits, which contained two LM subunits (LMα4 and LMγ1) of the
three that were expressed in human ALCs. Therefore, HLE B-3 BM
constructed in vitro may represent a substitute for human
ALCs for future studies into LM. It is a limitation that only three
human ALCs were used in the present study. Further studies using
more human ALC samples for evaluation of LM expression levels are
necessary to confirm the results of the present study and their
similarities with the established BM model of HLE B-3.
The LM411 trimer may be the predominant LM trimer in
human ALCs. It is well known that LM411 can promote cell adhesion,
cell migration, angiogenesis, tumor invasion and the
differentiation of stem cells (33–35).
Therefore, LM411 may be involved in the progression of human LEC
development, migration and transformation into lens fiber cells,
and that abnormal expression of LM411 may contribute to the
pathogenesis of cataracts; however, this requires further
confirmation.
In previous studies, LECs were mainly used as an
in vitro cell model to study the possible pathogenesis of
cataracts, with the results indicating that dysfunction of LECs
might serve a key role in cataract formation (8–10).
To the best of our knowledge, the present study is the first to
report the construction of an in vitro ALC model using human
LEC cell lines. The in vitro ALC model is advantageous as it
allows for the study of the contribution of BM proteins, such as
LMs, on the pathogenesis of cataracts. Further studies are required
to confirm the present results, validate HLE B-3 BMs as an in
vitro ALC model and investigate the possible molecular pathways
involving LMs in the pathogenesis of cataracts in vitro and
in vivo.
In conclusion, an in vitro human ALC model
rich in LMs was successfully constructed using the HLE B-3 cell
line, which could be valuable for the study of the molecular
biological mechanisms of cataract development and for seeking novel
effective drugs for cataract treatment.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
Nature Science Foundation of China (no. 1470618), the Scientific
Research Foundation of First Affiliated Hospital of Haerbin Medical
University (no. 2017B013), the Major Program of Applied Technology
Research and Development Plan of Heilongjiang Province (no.
GY2016ZB0159), the Special Fund for the Doctoral Program of Higher
Education (no. 20132307120035), the Natural Science Foundation of
China (no. 81300728), the Natural Science Foundation of
Heilongjiang Province of China (no. QC2010113) and the Postdoctoral
Grant of Heilongjiang Province (no. LBH-Q12038).
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding author on reasonable
request.
Authors' contributions
The paper is an original study presenting novel work
that has not been published or accepted elsewhere. All authors have
seen the manuscript and approved to submit to your journal. Project
design: LX, TX, HT and LP; clinical samples and information
collection: YY, SY and TX; project guidance: QH and LX; experiment
and data analysis: YY, QH, JH, YH, HT, SL, WX, GH, ZH, LX and TX;
manuscript revision: HT.
Ethics approval and consent to
participate
All experiments were performed with the approval of
the Internal Review Board of Harbin Medical University and were
conducted in accordance with Declaration of Helsinki Principles.
Informed consent was obtained from all patients prior to mortality
or from their families following mortality for inclusion of autopsy
data.
Consent for publication
Written informed consent was obtained for all
patients.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ALC
|
anterior lens capsule
|
|
BM
|
basement membrane
|
|
H&E staining
|
hematoxylin and eosin staining
|
|
ICC
|
immunocytochemistry
|
|
IHC
|
immunohistochemical staining
|
|
IP
|
immunoprecipitation
|
|
LECs
|
lens epithelial cells
|
|
LM
|
laminin
|
|
mAb
|
monoclonal antibodies
|
|
pAb
|
polyclonal antibodies
|
References
|
1
|
Michael R and Bron AJ: The ageing lens and
cataract: A model of normal and pathological ageing. Philos Trans R
Soc Lond B Biol Sci. 366:1278–1292. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bourne RR, Stevens GA, White RA, Smith JL,
Flaxman SR, Price H, Jonas JB, Keeffe J, Leasher J, Naidoo K, et
al: Causes of vision loss worldwide, 1990–2010: A systematic
analysis. Lancet Glob Health. 1:e339–e349. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Khairallah M, Kahloun R, Bourne R, Limburg
H, Flaxman SR, Jonas JB, Keeffe J, Leasher J, Naidoo K, Pesudovs K,
et al: Number of people blind or visually impaired by cataract
worldwide and in world regions, 1990 to 2010. Invest Ophthalmol Vis
Sci. 56:6762–6279. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pollreisz A and Schmidt-Erfurth U:
Diabetic cataract-pathogenesis, epidemiology and treatment. J
Ophthalmol. 2010:6087512010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Roberts JE: Ultraviolet radiation as a
risk factor for cataract and macular degeneration. Eye Contact
Lens. 37:246–249. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Asbell PA, Dualan I, Mindel J, Brocks D,
Ahmad M and Epstein S: Age-related cataract. Lancet. 365:599–609.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rao GN, Khanna R and Payal A: The global
burden of cataract. Curr Opin Ophthalmol. 22:4–9. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bhat SP: The ocular lens epithelium.
Biosci Rep. 21:537–563. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li WC, Kuszak JR, Dunn K, Wang RR, Ma W,
Wang GM, Spector A, Leib M, Cotliar AM, Weiss M, et al: Lens
epithelial cell apoptosis appears to be a common cellular basis for
non-congenital cataract development in humans and animals. J Cell
Biol. 130:169–181. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Martinez G and de Iongh RU: The lens
epithelium in ocular health and disease. Int J Biochem Cell Biol.
42:1945–1963. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Schéele S, Nyström A, Durbeej M, Talts JF,
Ekblom M and Ekblom P: Laminin isoforms in development and disease.
J Mol Med (Berl). 85:825–836. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Durbeej M: Laminins. Cell Tissue Res.
339:259–268. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yurchenco PD and Patton BL: Developmental
and pathogenic mechanisms of basement membrane assembly. Curr Pharm
Des. 15:1277–1294. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Miner JH and Yurchenco PD: Laminin
functions in tissue morphogenesis. Annu Res Cell Dev Biol.
20:255–284. 2004. View Article : Google Scholar
|
|
15
|
Byström B, Virtanen I, Rousselle P,
Gullberg D and Pedrosa-Domellöf F: Distribution of laminins in the
developing human eye. Invest Ophthalmol Vis Sci. 47:777–785. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kohno T, Sorgente N, Ishibashi T,
Goodnight R and Ryan SJ: Immunofluorescent studies of fibronectin
and laminin in the human eye. Invest Ophthalmol Vis Sci.
28:506–514. 1987.PubMed/NCBI
|
|
17
|
Uechi G, Sun Z, Schreiber EM, Halfter W
and Balasubramani M: Proteomic view of basement membranes from
human retinal blood vessels, inner limiting membranes, and lens
capsules. J Proteome Res. Jul 17–2014.(Epub ahead of print).
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Danysh BP and Duncan MK: The lens capsule.
Exp Eye Res. 88:151–164. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zenker M, Tralau T, Lennert T, Pitz S,
Mark K, Madlon H, Dötsch J, Reis A, Müntefering H and Neumann LM:
Congenital nephrosis, mesangial sclerosis and distinct eye
abnormalities with microcoria: An autosomal recessive syndrome. Am
J Med Genet A. 130A:1–145. 2004. View Article : Google Scholar
|
|
20
|
Stunf S, Hvala A, Vidovič Valentinčič N,
Kraut A and Hawlina M: Ultrastructure of the anterior lens capsule
and epithelium in cataracts associated with uveitis. Ophthalmic
Res. 48:12–21. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Suh J, Moncaster JA, Wang L, Hafeez I,
Herz J, Tanzi RE, Goldstein LE and Guénette SY: FE65 and FE65L1
amyloid precursor protein-binding protein compound null mice
display adult-onset cataract and muscle weakness. FASEB J.
29:2628–2639. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Joo CK, Lee EH, Kim JC, Kim YH, Lee JH,
Kim JT, Chung KH and Kim J: Degeneration and transdifferentiation
of human lens epithelial cells in nuclear and anterior polar
cataracts. J Cataract Refract Surg. 25:652–658. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
de Iongh RU, Wederell E, Lovicu FJ and
McAvoy JW: Transforming growth factor-beta-induced
epithelial-mesenchymal transition in the lens: A model for cataract
formation. Cells Tissues Organs. 179:43–55. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Arita T, Murata Y, Lin LR, Tsuji T and
Reddy VN: Synthesis of lens capsule in long-term culture of human
lens epithelial cells. Invest Ophthalmol Vis Sci. 34:355–362.
1993.PubMed/NCBI
|
|
25
|
Hirako Y, Yonemoto Y, Yamauchi T,
Nishizawa Y, Kawamoto Y and Owaribe K: Isolation of a
hemidesmosome-rich fraction from a human squamous cell carcinoma
cell line. Exp Cell Res. 324:172–182. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li X, Qian H, Sogame R, Hirako Y, Tsuruta
D, Ishii N, Koga H, Tsuchisaka A, Jin Z, Tsubota K, et al: Integrin
β4 is a major target antigen in pure ocular mucous membrane
pemphigoid. Eur J Dermatol. 26:247–253. 2016.PubMed/NCBI
|
|
27
|
Andjelic S, Drašlar K, Hvala A and Hawlina
M: Anterior lens epithelium in cataract patients with retinitis
pigmentosa-scanning and transmission electron microscopy study.
Acta Ophthalmol. 95:e212–e220. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li X, Qian H, Takizawa M, Koga H,
Tsuchisaka A, Ishii N, Hayakawa T, Ohara K, Sitaru C, Zillikens D,
et al: N-linked glycosylation on laminin γ1 influences recognition
of anti-laminin γ1 pemphigoid autoantibodies. J Dermatol Sci.
77:125–129. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Patrick B, Li J, Jeyabal PV, Reddy PM,
Yang Y, Sharma R, Sinha M, Luxon B, Zimniak P, Awasthi S, et al:
Depletion of 4-hydroxynonenal in hGSTA4-transfected HLE B-3 cells
results in profound changes in gene expression. Biochem Biophys Res
Commun. 334:425–432. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sigle RO, Gil SG, Bhattacharya M, Ryan MC,
Yang TM, Brown TA, Boutaud A, Miyashita Y, Olerud J and Carter WG:
Globular domains 4/5 of the laminin alpha3 chain mediate deposition
of precursor laminin 5. J Cell Sci. 117:4481–4494. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Frank DE and Carter WG: Laminin 5
deposition regulates keratinocyte polarization and persistent
migration. J Cell Sci. 117:1351–1363. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang XH, Sun HM and Yuan JQ:
Extracellular matrix production of lens epithelial cells. J
Cataract Refract Surg. 27:1303–1309. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Gonzalez AM, Gonzales M, Herron GS,
Nagavarapu U, Hopkinson SB, Tsuruta D and Jones JC: Complex
interactions between the laminin alpha 4 subunit and integrins
regulate endothelial cell behavior in vitro and angiogenesis in
vivo. Proc Natl Acad Sci USA. 99:pp. 16075–16080. 2002; View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Patarroyo M, Tryggvason K and Virtanen I:
Laminin isoforms in tumor invasion, angiogenesis and metastasis.
Semin Cancer Biol. 12:197–207. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Qu H, Liu X, Ni Y, Jiang Y, Feng X, Xiao
J, Guo Y, Kong D, Li A, Li X, et al: Laminin 411 acts as a potent
inducer of umbilical cord mesenchymal stem cell differentiation
into insulin-producing cells. J Transl Med. 12:1352014. View Article : Google Scholar : PubMed/NCBI
|