Introduction
Neuropathic pain (NP) is defined as a chronic pain
state caused by damage or abnormal function of the peripheral or
central nervous system (CNS) (1,2).
This pain may result from a wide variety of pathological processes
including inflammation, nerve trauma, nerve compression, genetic
abnormalities and metabolic disorders (3). Previous studies indicate that the
expression of potassium-chloride cotransporter 2 (KCC2) is
decreased in models of inflammation pain and NP (4,5).
Coull et al (6) first
proposed that KCC2 has a role in the production and development of
NP. Two isoforms of the cation-Cl (−) cotransporter family are
expressed in neurons and modulate neurotransmission, the KCC and
sodium potassium-chloride-co transporters (NKCC), mediate the
electroneutral movements of K+ and Cl-ions across cell membranes in
a coupled manner (7). KCC2
involves in the control of numerous neuronal processes, including
migration, dendritic outgrowth, the formation of synaptic
connections, and exerts a great influence on the establishment of
neuronal [Cl-]i (6,8). It maintains a low concentration of
intracellular Cl-, which is essential for inhibitory synaptic
transmission by γ-aminobutyric acid (GABA) in the mature nervous
system (9). Previous work has
demonstrated that GABA-ergic interneurons are important in spinal
nociceptive processing and nociceptive attenuation (10,11).
GABAA receptors, which are the major synaptic targets for the
neurotransmitter GABA, are located in primary afferent terminals
and interneurons in laminae I–IV of the spinal cord dorsal horn
(12,13). The GABAA receptors are ligand-gated
chloride channels in the CNS and mediating inhibitory
neurotransmission, consist of subunits such as α1-6, β1-3, γ1-3, δ,
ε, π, θ and ρ1–3 (14,15), the most common subtype is composed
of α1, β2 and γ2 subunits (16).
The location of γ2 subunit mRNA-containing neurons differs from
those containing α- or β-subunits. Earlier work has indicated that
the distribution of strongly labeled γ2 subunit nuclei partially
coincided with that of glutamate decarboxylase, suggesting that the
GABAA receptor γ2 subunit may be involved in the mechanism of
GABA-ergic transmission (17).
Electro-acupuncture (EA) is known to relieve both
peripheral NP and acute or chronic inflammatory pain via electrical
stimulation (18,19). However, little is understood about
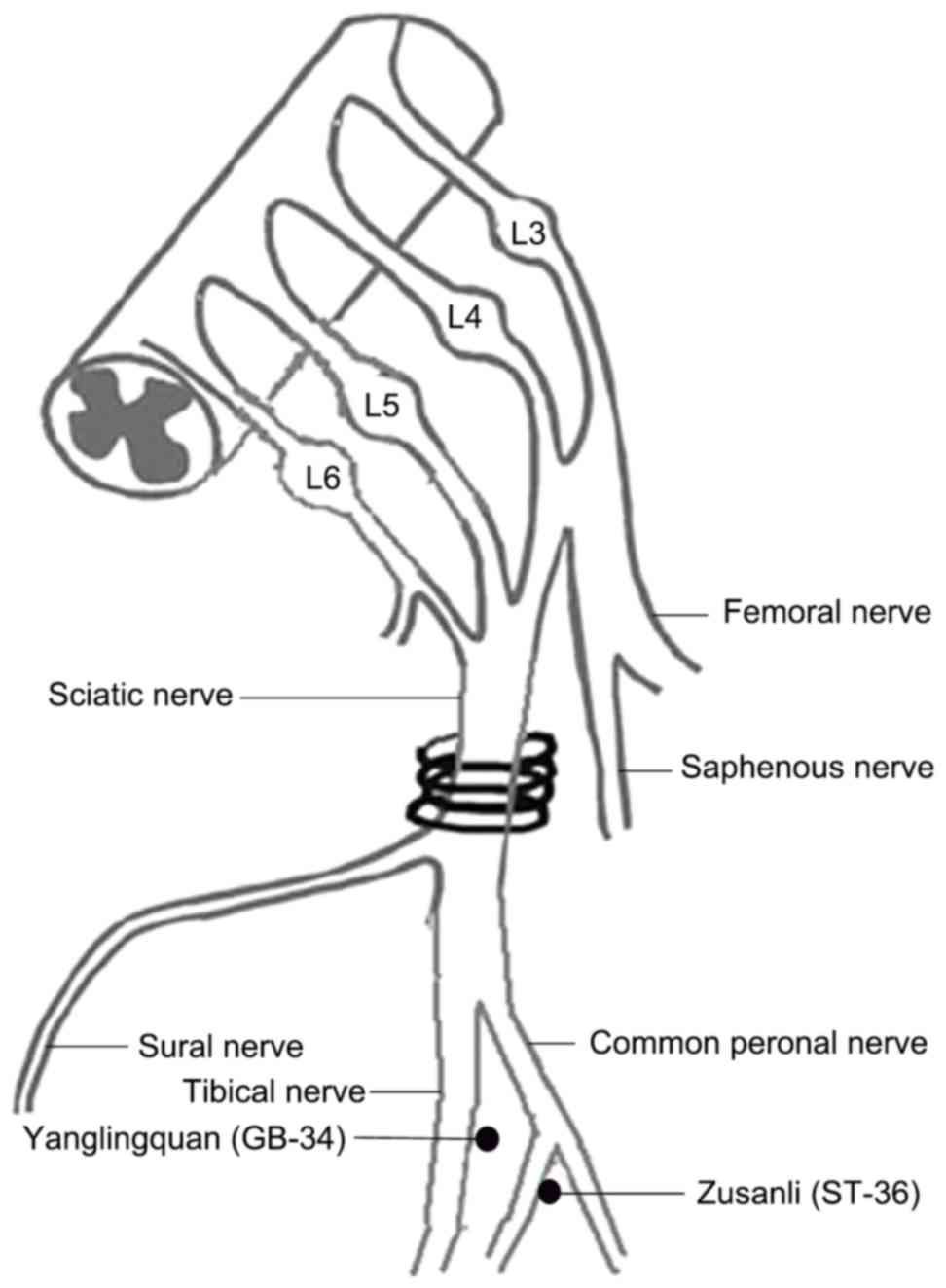
its mechanism of action. In the present work, the acupoints Zusanli
(ST-36) and Yanglingquan (GB-34) were applied following the
production of NP by chronic constriction injury (CCI).
The current study investigated the application of EA
on ST-36 and GB-34 acupoints and its effect on modulating
mechanical allodynia and thermal hyperalgesia in rats with NP. In
addition, the relationship between EA and the KCC2-GABAA receptor
signaling pathway in CCI rats was investigated.
Materials and methods
Experimental animals
Male Sprague-Dawley rats (180–200 g) were purchased
from Wenzhou Medical University (Wenzhou, China). All experiments
were maintained and cared for according to the guidelines of the
National Institutes of Health Guide for the Care and Use of
Laboratory Animals (NIH Publications no. 80–23, revised 1978). The
experimental protocol was approved by the Institutional Animal Care
and Use Committee of Wenzhou Medical University (Wenzhou, China).
All efforts were made to minimize the animals' pain and discomfort
during the experiments. A total of 60 male Sprague-Dawley rats were
housed in a climate-controlled room with a constant temperature
(22±2°C) and humidity (55±5%) in a 12-h light/dark cycle with food
and water available ad libitum. Rats were randomly divided
into four groups: Normal group, sham-CCI group, CCI group and CCI
plus EA group. All experiments were conducted with the
experimenters blinded to treatment conditions.
CCI surgery
CCI to the sciatic nerve of the right hind limb in
rats was performed based on previous description (2). Briefly, animals were anaesthetized
with 4% chloral hydrate (10 ml/kg; i.p.). The sciatic nerve of the
right hind limb was exposed at the middle of the thigh by blunt
dissection. To prevent the interruption of blood circulation
through the epineural vasculature, four chromic gut ligatures were
loosely tied (4.0 silk) around the nerve with spacing at ~1 mm. In
the sham-CCI group rats, the right sciatic nerve was exposed for
2–3 min, but was not ligated. Following surgery, the skin was
closed with a single suture, and the animals were allowed to
recover for 7 days.
Mechanical withdrawal threshold
(MWT)
Mechanical allodynia and thermal hyperalgesia are
reproducible and sensitive behavioral readouts of neuropathic pain.
Behavioral testing was conducted prior to surgery and on days 3, 5,
7, 10, 12 and 14 following surgery. Animals were allowed to
acclimate to elevated cages (20×14×16 cm) with a wire mesh bottom.
MWT was measured by assessing hind paw sensitivity to innocuous
mechanical stimulation. Ascending mechanical pressure (0–70 g) was
applied to the plantar aspect of right hind paw by the 2392
Electronic von Frey Anesthesiometer (IITC Life Science, Woodland
Hills, CA, USA). Lifting, licking the paw and running away were all
considered as positive responses. The maximum applied pressure was
recorded. The MWT of each animal was the average of six
measurements taken at 5 min intervals.
Thermal withdrawal latency (TWL)
In this assay, rats were placed in a transparent,
square, bottomless acrylic box (17×11.5×14 cm) and allowed to adapt
for 20 min. Responses to thermal stimulation were evaluated using a
37370 plantar test apparatus (UgoBasile SRL, Milan, Italy) as a
source of radiant heat. A beam of focused light set at 60°C was
directed towards the plantar surface of the hind paw, and the
maximum latency time was recorded. The time to purposeful
withdrawal of the foot from the beam of light was measured. A
cut-off time was set at 40 sec to prevent tissue damage. Every hind
paw was tested alternately at 5 min intervals. The results obtained
for each rat were expressed in sec (sec) as the mean of five
withdrawal latencies. Finally, the average value was used for
statistical analysis.
EA stimulation
EA treatment was given between 9:00 and 11:00 a.m.
every day and lasted for 7 days. The rats were maintained in an
immobilization apparatus designed by the authors' laboratory
(patent application number: 201110021482.5, State Intellectual
Property Office) (20–22). During EA treatment, stainless steel
needles were percutaneously inserted into the right hind leg at two
points: Zusanli acupoint (ST-36, 5 mm below the head of the fibula
under the knee joint, and 2 mm lateral to the anterior tubercle of
the tibia) and the Yanglingquan acupoint (GB-34, ~5 mm
superior-lateral to ST-36; Fig. 1)
(23,24). It has been previously reported that
2 Hz induced analgesia mediated by the release of met-enkephalin
and β-endorphin, while 100 Hz via release of dynorphin-A in the CNS
(25). Combining the low (2 Hz)
and high (100 Hz) frequencies alternately (named ‘2/100 Hz’)
elicits a synergistic analgesic effect (26). The intensity of the stimulation was
set at 1.5 mA, which elicited a slight twitch. A HANS-200E (Jisheng
Medical Instruments Co., Ltd., Nanjing, China) was used to deliver
the acupuncture stimulus and enabled the performance of EA.
Acupuncture needles were inserted 2–3 mm deep and stimulated
electrically at a frequency of 2/100 Hz to permit the muscle to
shrink slightly. The total stimulation period was 30 min.
Immunohistochemistry
Rats (n=5 per group) were anaesthetized as described
above and perfused via the aorta with 0.9% normal saline, followed
by addition of 4% paraformaldehyde in phosphate-buffered saline on
day 14 following surgery. Spinal cord segments (L4-L6) were
collected and fixed in 4% paraformaldehyde for 24 h. Following
fixation, they were dehydrated, cleared and embedded in paraffin
for transverse paraffin sections. The paraffin sections (5-µm
thick) were made and mounted on poly-L-lysine-coated slides for
immunohistochemistry. To block the activity of endogenous
peroxidase, the slides were incubated in 3%
H2O2 for 10 min at room temperature and
blocked in 10% normal goat serum with 0.3% Triton X-100 in PBS for
1 h. Sections were probed with mouse anti-KCC2 (1:200; cat. no.
ab134300; Abcam, Cambridge, MA, USA) and rabbit anti-GABAA receptor
γ2 subunit (1:100; cat. no. orb5291; Biorbyt Ltd., Cambridge, UK)
antibodies overnight at 4°C. The next day, slides were incubated
for 30 min at 37°C with the following secondary antibodies:
Two-step plus Poly-horseradish peroxidase (HRP) anti-mouse
immunoglobulin (Ig)G detection system (1:1; cat. no. PV-6002;
Beijing Zhongshan Golden Bridge Biotechnology, Co., Ltd., Beijing,
China) and two-step plus Poly-HRP anti-rabbit IgG detection system
(1:1; cat. no. PV-6001; Beijing Zhongshan GoldenBridge
Biotechnology, Co., Ltd.). Slides were then stained with DAB for 20
sec and counterstained with haematoxylin for 8 min at room
temperature. Finally, images were captured via a light microscope
(Olympus Corporation, Tokyo, Japan). Image-Pro Plus 5.1 software
(Media Cybernetics, Inc., Rockville, MD, USA) was used to define
and determine positive regions of interest.
Western blotting
Rats (n=5 per group) were deeply anesthetized as
described above and perfused with 0.9% normal saline. The L4-L6
spinal cord segments (1 cm) were dissected on day 14 post-CCI
surgery. The segments were lysed in fresh radioimmunoprecipitation
assay protein lysis buffer containing phenylmethylsulfonyl fluoride
(PMSF) (RIPA: PMSF=100:1) for 30 min and centrifuged at 15,294 × g
for 5 min at 4°C, and the supernatant was subsequently collected.
Protein concentration of each sample was determined using a
bicinchoninic acid assay protein assay kit (Beyotime Institute of
Biotechnology, Haimen, China). The proteins were mixed with loading
buffer, heated to 100°C for 10 min and separated on an 8 or 10%
Tris-HCl SDS-PAGE gel (Bio-Rad Laboratories, Inc., Hercules, CA).
Following electrophoresis, the proteins were transferred to a
polyvinylidene fluoride membrane. The membranes were blocked with
5% non-fat milk for 2 h and incubated overnight at 4°C with the
following antibodies: Mouse anti-KCC2 (1:400; cat. no. ab134300;
Abcam); rabbit anti-GABAA receptor γ2 subunit (1:400; cat. no.
orb5291; Biorbyt Ltd.). Next, the membranes were incubated with
peroxidase-conjugated goat anti-mouse IgG (H+L) (1:5,000; cat. no.
BL001A; Biosharp, Hefei, China) or peroxidase-conjugated goat
anti-rabbit IgG (H+L) secondary antibody (1:5,000; cat. no. BL003A;
Biosharp). Following being washed, the labeled proteins were
visualized using the enhanced chemiluminescence (ECL) kit (Beyotime
Institute of Biotechnology). The quantity of band intensity was
detected by aDNR microchemiluminescence gel imaging system (DNR
Bio-Imaging Systems, Jerusalem, Israel) and using AlphaEase FC
4.0.0 software from ProteinSimple, (San Jose, CA, USA). The band
densities were normalized to GAPDH.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the L4-L6 segment with
TRIzol (Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) according to the manufacturer's instructions (n=5 per group).
Total RNA (1 µg) was reverse transcribed using High Capacity
RNA-to-cDNA Master Mix (cat. no. A3500; Promega Corporation,
Madison, WI, USA). Real-time amplification was conducted using SYBR
Green Supermix (Toyobo Co., Ltd., Osaka, Japan) and a Light Cycler
480 system (Roche Diagnostics GmbH, Mannheim, Germany). Primer
sequences were the following: KCC2 forward,
5′-AGCAAAGAGCACGAAGAAGC-3′ and reverse, 5′-GCCGCAGAAAGAGGATAACA-3′;
GABAA receptor γ2 subunit forward, 5′-TTCTGTCCTGGGTGTCCTTC-3′ and
reverse, 5′-AGAGACTTCCGGGCTATGGT-3′; RPS16 forward,
5′-AAGTCTTCGGACGCAAGAAA-3′ and reverse, 5′-TTGCCCAGAAGCAGAACAG-3′.
The PCR reactions were prepared in a total volume of 10 µl,
containing 1 µl cDNA, 0.6 µl forward and reverse primer, 5 µl SYBR
Green Supermix, 1 µl plus solution (this solution is a component
that increases the specificity and reliability of the PCR reaction)
and 1.8 µl RNase-free water. To amplify genomic DNA, PCR was
performed under the following conditions: 95°C for 5 min, followed
by 40 cycles of 95°C for 10 sec, 60°C for 10 sec, and 72°C for 10
sec. The quantification values were obtained from the
quantification cycle (Cq) number at which the increase in the
signal was associated with an exponential growth of the PCR
products. All samples were run in triplicate and repeated three
times. RPS16 quantification was used as an internal control for
normalization. Fold differences of mRNA levels over vehicle control
were calculated by 2-ΔΔCq method (27).
Statistical analysis
Statistical analyses were carried out using the SPSS
statistical software (version, 16.0; SPSS, Inc., Chicago, IL, USA).
Numerical data of protein and mRNA assays were analyzed and
compared by one-way analysis of variance (ANOVA) followed by a post
hoc comparison test using the LSD (equal variances assumed) or
Kruskal-Wallis (equal variances not assumed) method. Behavioral
results with multiple comparisons were statistically analyzed by
ANOVA for repeated measures. All values were presented as the means
± standard deviation. P<0.05 were considered to indicate a
statistically significant difference.
Results
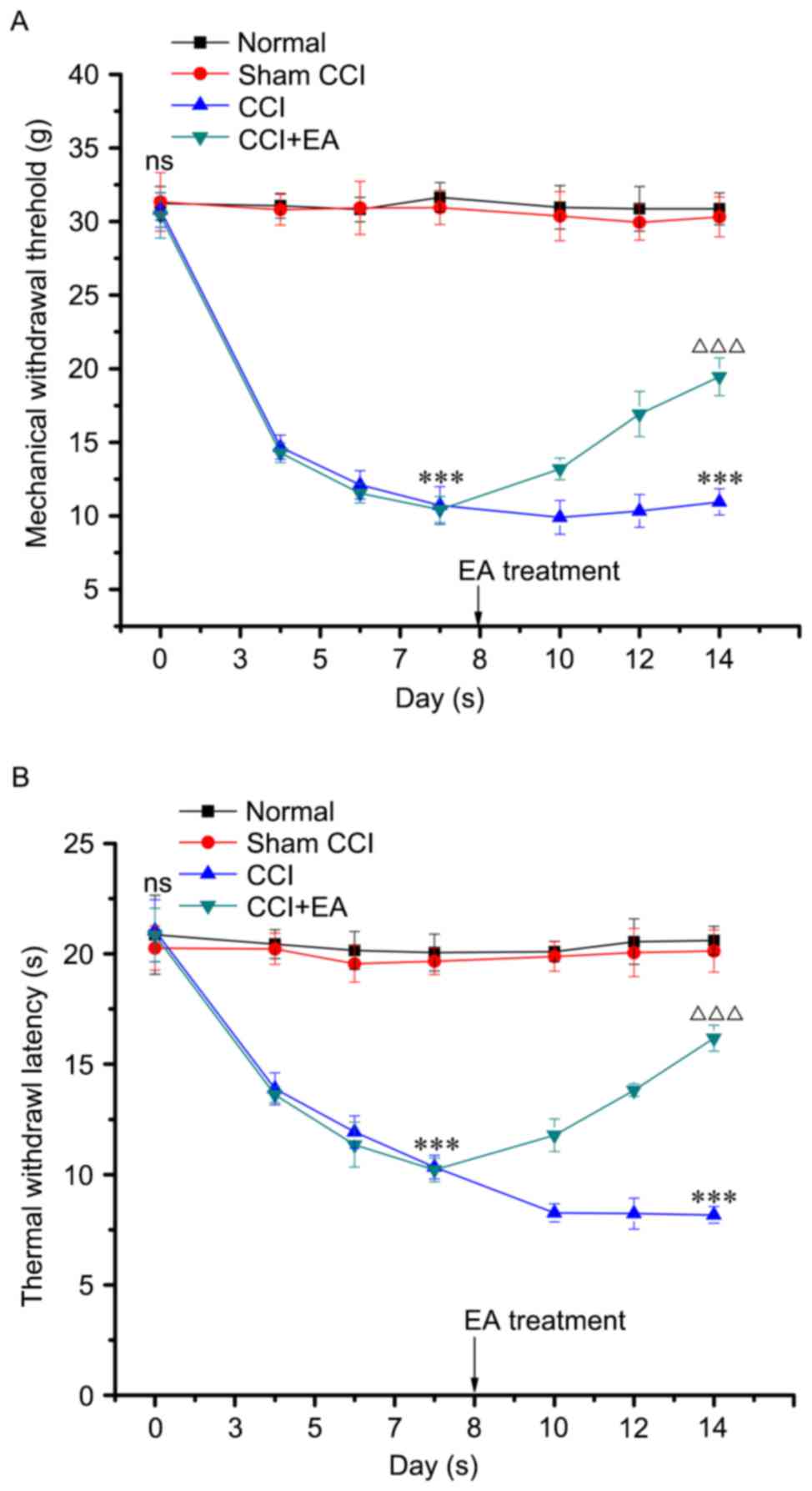
EA treatment reversed CCI-induced mechanical
allodynia and thermal hyperalgesia. The MWT and TWL values were
normalized to baseline (day 0) to determine the time course of
CCI-induced mechanical allodynia and thermal hyperalgesia. There
was no significant difference among the groups. In behavioral tests
on day 3 post-CCI operation, MWT and TWL were significantly lower
in the CCI group compared with the normal and sham-CCI groups
(P<0.001). On day 7, the MWT was markedly reduced from 30.8±1.16
to 10.7±1.28 g, and TWL was reduced from 21.0±1.40 to 10.3±0.54 sec
compared with the normal and sham-CCI groups (P<0.001). EA
treatment markedly reduced the symptoms of neuropathic pain. As
presented in Fig. 2A and B, EA
significantly reversed the established mechanical allodynia and
thermal hyperalgesia in the CCI model occurring on day 10. On day
14, the MWT and TWL of the CCI+EA group were markedly higher than
those in the CCI group (P<0.001). Taken together, these results
suggest that EA therapy effectively reduces the severity of pain
induced by sciatic nerve injury.
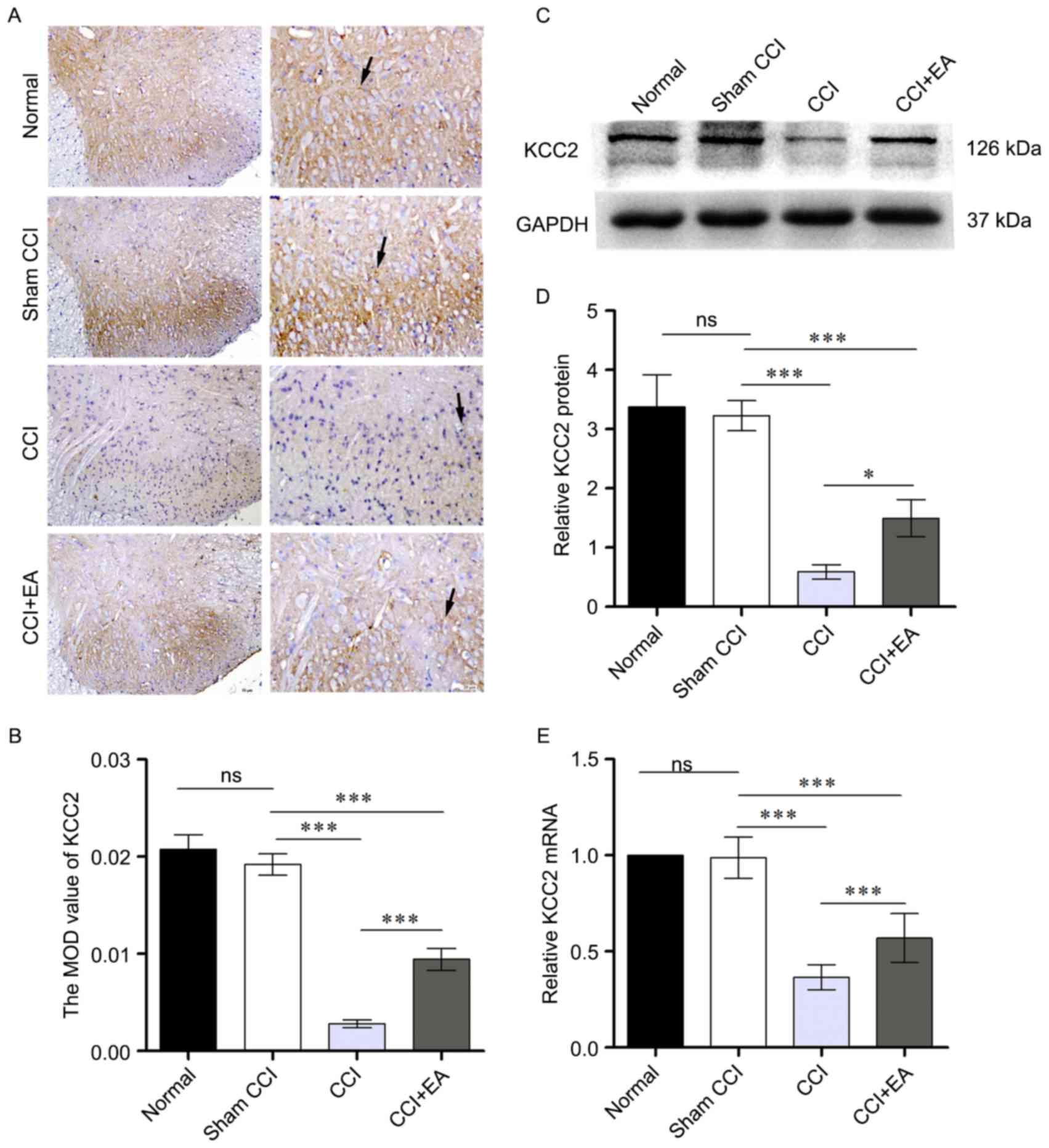
EA upregulated KCC2 protein and mRNA levels in
spinal cord. To explore the analgesic effect of EA, and prove the
critical role of KCC2 for maintaining neuropathic pain, the authors
next examined the protein and mRNA levels of KCC2 in the spinal
cord following repeated EA treatments. Immunohistochemistry was
performed in each group (Fig. 3A and
B). Comparisons between the CCI and CCI+EA group revealed
significant effect, KCC2 was evident in CCI+EA group relative to
the CCI group. Furthermore, using western blotting, the authors
examined the protein level of KCC2 in the L4-L6 spinal cord. CCI
resulted in a significant reduction in the KCC2 protein levels
compared to the sham-CCI group. The relative optical density value
for KCC2 in the EA group was significantly higher than that in the
CCI group (P<0.05; Fig. 3C and
D). RT-qPCR analysis confirmed the downregulation of KCC2 under
chronic constriction at the sciatic nerve. However, following 7
days of EA treatment, KCC2 expression was significantly higher than
it in the CCI group (Fig. 3E).
Together, these results suggested that CCI may lead to a loss of
KCC2 mRNA and protein, 2/100 Hz EA may increase KCC2 expression in
the spinal cord of CCI rats, which, in turn may potentiate
EA-induced analgesia in the spinal cord.
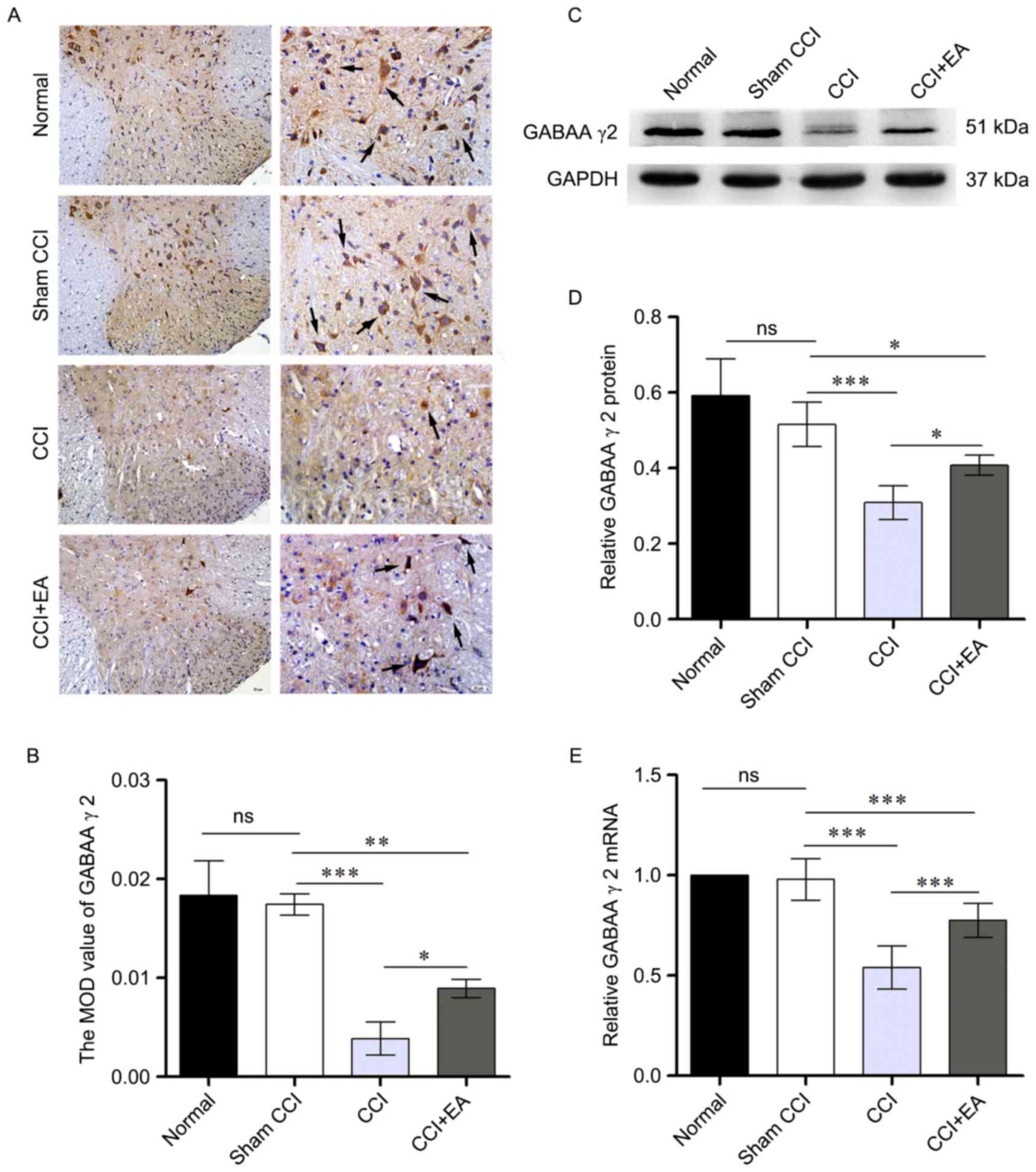
EA treatment increased spinal GABAA receptor γ2
subunit expression in CCI rats. To evaluate the effect of EA on the
GABAA receptor γ2 subunit, immunohistochemistry, western blotting
and RTq-PCR analyzed protein and mRNA levels. Direct analysis of
GABAA receptor γ2 subunit in the spinal cord was investigated using
immunohistochemistry. As is demonstrated in Fig. 4A and B, EA significantly increased
the expression of GABAA receptor γ2 subunit (P<0.05). Western
blotting produced similar results at the protein level (P<0.05;
Fig. 4C and D). Data from RT-qPCR
revealed a prominent reduction in GABAA receptor γ2 subunit mRNA in
the spinal cord of CCI rats. However, EA significantly attenuated
the CCI-induced downregulation of GABAA receptor γ2 subunit mRNA
(P<0.001; Fig. 4E). These data
suggested that CCI-induced NP inhibits the expression of GABAA
receptor γ2 subunit, thus indicating a potential role for GABAA
receptor γ2 subunit in mediating pain inhibition in response to
CCI.
Discussion
The current study demonstrated that EA treatment has
a beneficial effect on the pain threshold of rats undergoing CCI.
The data indicated that the mechanical and thermal nociceptive
thresholds maybe increased by EA on ST-36 and GB-34 acupoints in
the CCI plus EA group. Furthermore, the authors examined the mRNA
and protein levels of KCC2 and GABAA receptor γ2 subunit in CCI
rats and indicated that EA treatment significantly increased KCC2
and GABAA receptor γ2 subunit expression.
A great deal of evidence from diverse animal models
of neuropathic pain has suggested that glia, particularly activated
microglia, serve a critical role in the defense of the neural
parenchyma (28). Following
peripheral nerve injury, activated microglia will upregulate their
surface expression of immunomodulatory proteins and release
neurotransmitters and immunomodulators such as opioids, serotonin,
noradrenaline and adenosine (29).
Brain-derived neurotrophic factor (BDNF), which can be synthesized
and released of microglia, has been reported to be crucial in the
occurrence and development of NP (30). Most of the cellular behaviors of
BDNF in the spinal cord are mediated by its high-affinity receptor
tropomyosin-related kinase B (TrkB), and BDNF-TrkB has previously
been shown to downregulate expression of the KCC2 (6,30).
It is reported that the BDNF-TrkB-KCC2 signaling pathway may
contribute to NP not only during its occurrence but also during its
maintenance (6). Based on these
data, it was hypothesized that chronic pain may be correlated to
KCC2 and its signal transduction pathway, the analgesic effect of
EA may be associated with regulation of this pathway.
Accumulated evidence suggests that KCC2, the main
Cl-transporter in spinal lamina I neurons, leads to decrease
intracellular Cl-concentration (31). It serves a pivotal role in the
inhibitory mechanisms that control neuronal excitation in the CNS
(30). The downregulation of KCC2
has been implicated in various excitatory disorders, such as in
epilepsy or in nerve injury resulting from various pathological
conditions (32). As a result,
GABAA receptors can be reduced which potentially shift the
receptor-mediated responses from hyperpolarizing inhibition to
depolarizing excitation (32,33).
The increased neuronal excitability may then sustain the enhanced
(and exaggerated) communication between primary afferents and
dorsal horn neurons to contribute to the early behavioral signs of
pain (34).
The current study demonstrated that NP decreased the
expression of KCC2 and GABAA receptor γ2 subunit mRNA and protein
in CCI rats and this occurred concomitantly with a marked increase
in allodynia. However, EA significantly relieved mechanical
allodynia and thermal hyperalgesia and increased the expression of
KCC2 and GABAA receptor γ2 subunit.
EA has been established as a viable therapeutic
intervention to alleviate chronic pain and has shown positive
effects in the clinic, but the biological basis of its action
remains unknown (35). Several
studies have demonstrated that EA has an immunomodulatory effect
(36). Zhang Wang and McAlonan
(37) suggested that acupuncture
stimulation can release pain killers and CNS mediators, including
endorphin, morphine and GABA. GABA has been implicated not only in
peripheral responses but also in interfering with pain impulse
input in the CNS. In the present study, EA prevented decreased
levels of KCC2 and GABAA in the central sensory pathways and
reduced NP following peripheral nerve injury. Based on these
findings, the authors suspect that the mechanical allodynia and
thermal hyperalgesia mediated by KCC2-GABAA receptor signaling
pathway is attenuated by EA via its immunomodulatory effects in the
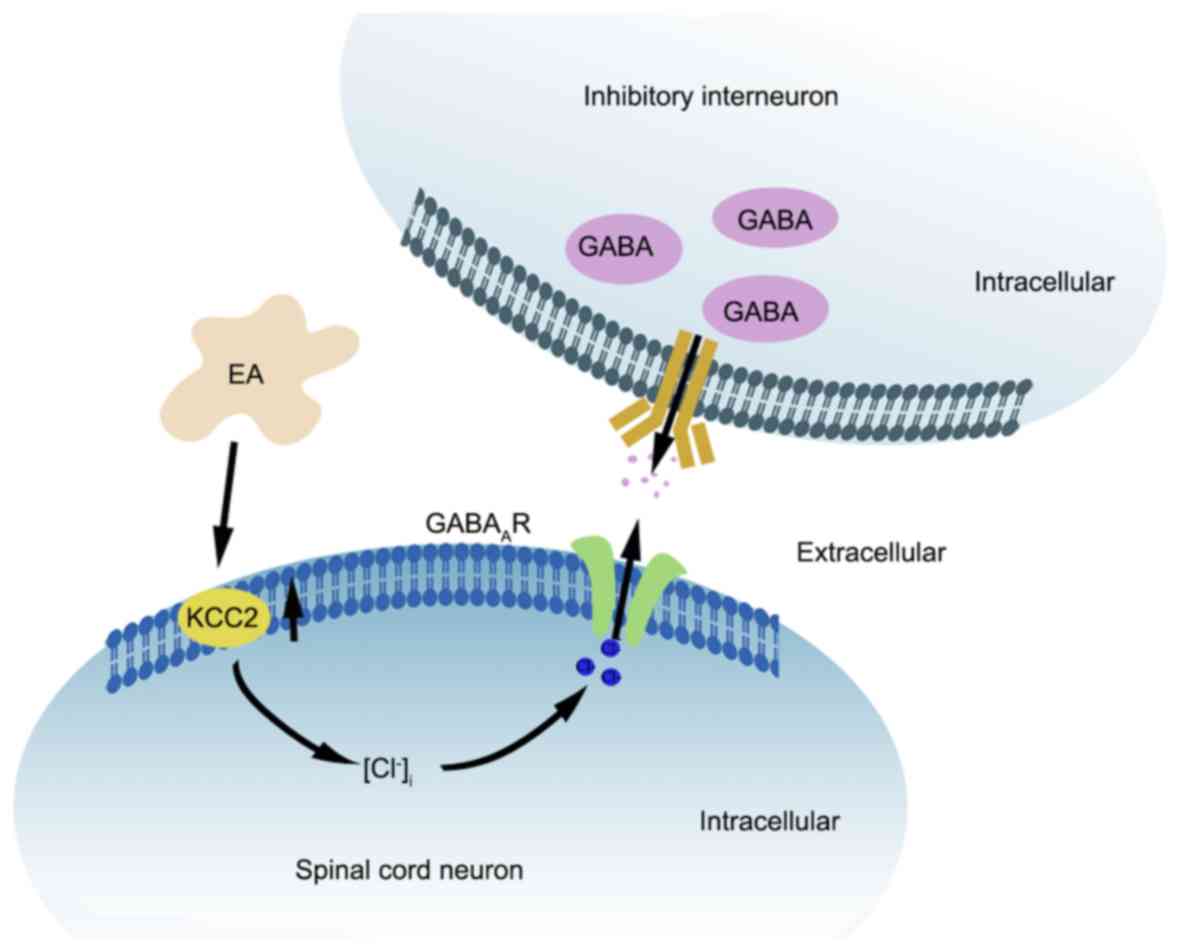
spinal cord (Fig. 5). To further
characterize the analgesic effect of EA, pharmacology,
electrophysiological techniques and other methods will be conducted
in the future.
In conclusion, these data suggested that peripheral
nerve injury caused a reduction in KCC2 and GABAA receptor γ2
subunit expression in the spinal cord and that the upregulation of
KCC2 and GABAA receptor γ2 subunit may account for the analgesic
effect of EA. Overall, the results of the present initial study
provide a promising foundation for future research on the impact of
EA on NP.
Acknowledgements
Not applicable.
Funding
The current study was supported by the Natural
Science Foundation of Zhejiang Province (grant no. LY16H270016) and
the Foundation of Wenzhou Scientific and Technological Bureau
Project (grant no. Y20140221).
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding author on reasonable
request.
Authors' contributions
SL, WT, SJ and BL conceived and designed the
experiments. CJ, WC and XJ performed the experiments. XQ, GY and QH
analyzed the data and prepared the figures. WT and SL wrote the
paper. BL and SJ contributed to the modification of the manuscript.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All experiments were maintained and cared for
according to the guidelines of the National Institutes of Health
Guide for the Care and Use of Laboratory Animals (NIH Publications
No. 80–23, revised 1978). The experimental protocol was approved by
the Institutional Animal Care and Use Committee of Wenzhou Medical
University (Wenzhou, China).
Consent for publication
Not applicable.
Competing interests
The authors declare they have no competing
interests.
References
|
1
|
Amit Z and Galina ZH: Stress-induced
analgesia: Adaptive pain suppression. Physiol Rev. 66:1091–1120.
1986. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fox A, Kesingland A, Gentry C, McNair K,
Patel S, Urban L and James I: The role of central and peripheral
Cannabinoid1 receptors in the antihyperalgesic activity of
cannabinoids in a model of neuropathic pain. Pain. 92:91–100. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Woolf CJ and Mannion RJ: Neuropathic pain:
Aetiology, symptoms, mechanisms, and management. Lancet.
353:1959–1964. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Miletic G and Miletic V: Loose ligation of
the sciatic nerve is associated with TrkB receptor-dependent
decreases in KCC2 protein levels in the ipsilateral spinal dorsal
horn. Pain. 137:532–539. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhang W, Liu LY and Xu TL: Reduced
potassium-chloride co-transporter expression in spinal cord dorsal
horn neurons contributes to inflammatory pain hypersensitivity in
rats. Neuroscience. 152:502–510. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Coull JA, Boudreau D, Bachand K, Prescott
SA, Nault F, Sík A, De Koninck P and De Koninck Y: Trans-synaptic
shift in anion gradient in spinal lamina I neurons as a mechanism
of neuropathic pain. Nature. 424:938–942. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Delpire E: Cation-chloride cotransporters
in neuronal communication. News Physiol Sci. 15:309–312.
2000.PubMed/NCBI
|
|
8
|
Medina I, Friedel P, Rivera C, Kahle KT,
Kourdougli N, Uvarov P and Pellegrino C: Current view on the
functional regulation of the neuronal K(+)-Cl(−) cotransporter
KCC2. Front Cell Neurosci. 8:272014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Acton BA, Mahadevan V, Mercado A, Uvarov
P, Ding Y, Pressey J, Airaksinen MS, Mount DB and Woodin MA:
Hyperpolarizing GABAergic transmission requires the KCC2 C-terminal
ISO domain. J Neurosci. 32:8746–8751. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Price TJ, Cervero F and de Koninck Y: Role
of cation-chloride-cotransporters (CCC) in pain and hyperalgesia.
Curr Top Med Chem. 5:547–555. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Price TJ, Cervero F, Gold MS, Hammond DL
and Prescott SA: Chloride regulation in the pain pathway. Brain Res
Rev. 60:149–170. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bowery NG, Hudson AL and Price GW: GABAA
and GABAB receptor site distribution in the rat central nervous
system. Neuroscience. 20:365–383. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Malherbe P, Sigel E, Baur R, Persohn E,
Richards JG and Mohler H: Functional characteristics and sites of
gene expression of the alpha 1, beta 1, gamma 2-isoform of the rat
GABAA receptor. J Neurosci. 10:2330–2337. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Barnard EA, Skolnick P, Olsen RW, Mohler
H, Sieghart W, Biggio G, Braestrup C, Bateson AN and Langer SZ:
International union of pharmacology. XV. Subtypes of
gamma-aminobutyric acidA receptors: Classification on the basis of
subunit structure and receptor function. Pharmacol Rev. 50:291–313.
1998.PubMed/NCBI
|
|
15
|
Bonnert TP, McKernan RM, Farrar S, le
Bourdellès B, Heavens RP, Smith DW, Hewson L, Rigby MR,
Sirinathsinghji DJ, Brown N, et al: theta, a novel
gamma-aminobutyric acid type A receptor subunit. Proc Natl Acad Sci
USA. 96:pp. 9891–9896. 1999; View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sieghart W and Sperk G: Subunit
composition, distribution and function of GABA(A) receptor
subtypes. Curr Top Med Chem. 2:795–816. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Araki T, Sato M, Kiyama H, Manabe Y and
Tohyama M: Localization of GABAA-receptor gamma 2-subunit
mRNA-containing neurons in the rat central nervous system.
Neuroscience. 47:45–61. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kelly RB: Acupuncture for pain. Am Fam
Physician. 80:481–484. 2009.PubMed/NCBI
|
|
19
|
Ulett GA, Han S and Han JS:
Electroacupuncture: Mechanisms and clinical application. Biol
Psychiatry. 44:129–138. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tu W, Wang W, Xi H, He R, Gao L and Jiang
S: Regulation of neurotrophin-3 and interleukin-1β and inhibition
of spinal glial activation contribute to the analgesic effect of
electroacupuncture in chronic neuropathic pain states of rats. Evid
Based Complement Alternat Med. 2015:6420812015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang WS, Tu WZ, Cheng RD, He R, Ruan LH,
Zhang L, Gong YS, Fan XF, Hu J, Cheng B, et al: Electroacupuncture
and A-317491 depress the transmission of pain on primary afferent
mediated by the P2X3 receptor in rats with chronic neuropathic pain
states. J Neurosci Res. 92:1703–1713. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tu WZ, Cheng RD, Cheng B, Lu J, Cao F, Lin
HY, Jiang YX, Wang JZ, Chen H and Jiang SH: Analgesic effect of
electroacupuncture on chronic neuropathic pain mediated by P2X3
receptors in rat dorsal root ganglion neurons. Neurochem Int.
60:379–386. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lee CH, Kim DK, Yook TH, Sasaki M and
Kitamura N: Effectiveness of electroacupuncture at Zusanli (ST36)
on the immunohistochemical density of enteroendocrine cells related
to gastrointestinal function. J Acupunct Meridian Stud. 5:63–71.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen SP, Kan Y, Zhang JL, Wang JY, Gao YH,
Qiao LN, Feng XM, Yan YX and Liu JL: Involvement of hippocampal
acetylcholinergic receptors in electroacupuncture analgesia in
neuropathic pain rats. Behav Brain Funct. 12:132016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang Y, Zhang Y, Wang W, Cao Y and Han JS:
Effects of synchronous or asynchronous electroacupuncture
stimulation with low versus high frequency on spinal opioid release
and tail flick nociception. Exp Neurol. 192:156–162. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Silva JR, Silva ML and Prado WA: Analgesia
induced by 2- or 100-Hz electroacupuncture in the rat tail-flick
test depends on the activation of different descending pain
inhibitory mechanisms. J Pain. 12:51–60. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cao H and Zhang YQ: Spinal glial
activation contributes to pathological pain states. Neurosci
Biobehav Rev. 32:972–983. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Suzuki T, Hide I, Ido K, Kohsaka S, Inoue
K and Nakata Y: Production and release of neuroprotective tumor
necrosis factor by P2X7 receptor-activated microglia. J Neurosci.
24:1–7. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Coull JA, Beggs S, Boudreau D, Boivin D,
Tsuda M, Inoue K, Gravel C, Salter MW and De Koninck Y: BDNF from
microglia causes the shift in neuronal anion gradient underlying
neuropathic pain. Nature. 438:1017–1021. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kahle KT, Staley KJ, Nahed BV, Gamba G,
Hebert SC, Lifton RP and Mount DB: Roles of the cation-chloride
cotransporters in neurological disease. Nat Clin Pract Neurol.
4:490–503. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang D, Gopalakrishnan SM, Freiberg G and
Surowy CS: A thallium transport FLIPR-based assay for the
identification of KCC2-positive modulators. J Biomol Screen.
15:177–184. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
De Koninck Y: Altered chloride homeostasis
in neurological disorders: A new target. Curr Opin Pharmacol.
7:93–99. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Nomura H, Sakai A, Nagano M, Umino M and
Suzuki H: Expression changes of cation chloride cotransporters in
the rat spinal cord following intraplantar formalin. Neurosci Res.
56:435–440. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Berman BM, Langevin HM, Witt CM and Dubner
R: Acupuncture for chronic low back pain. N Engl J Med.
363:454–461. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ouyang BS, Gao J, Che JL, Zhang Y, Li J,
Yang HZ, Hu TY, Yang M, Wu YJ and Ji LL: Effect of
electro-acupuncture on tumor necrosis factor-α and vascular
endothelial growth factor in peripheral blood and joint synovia of
patients with rheumatoid arthritis. Chin J Integr Med. 17:505–509.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang ZJ, Wang XM and McAlonan GM: Neural
acupuncture unit: A new concept for interpreting effects and
mechanisms of acupuncture. Evid Based Complement Alternat Med.
2012:4294122012. View Article : Google Scholar : PubMed/NCBI
|