Introduction
Osteoporosis is a skeletal disorder characterized by
reduced bone mass and micro-architectural deterioration of bone
tissue (1). It is estimated that,
in the United States, ~50% of women aged >50 years may sustain
an osteoporotic fracture during their lifetime (2). Despite the high incidence rate of
osteoporosis, the effect of therapeutic agents, including
bisphosphonates, calcitonin and estrogen in recovering bone mass is
limited. Osteoclasts are unique bone-resorbing cells derived from
cells of the monocyte-macrophage lineage (3). Numerous factors are involved in
osteoclast differentiation (4–6).
Receptor activator of nuclear factor-κB ligand (RANKL) is a
membrane protein of the tumor necrosis factor (TNF) family, and
increasing evidence indicates that RANKL serves a critical role in
osteoclast differentiation (6–8). It
has been reported that RANKL may activate various downstream
signaling pathways, including the nuclear factor-κB (NF-κB) and
mitogen-activated protein kinase (MAPK) signaling pathways.
Activation of these pathways may upregulate expression of
osteoclast-specific genes, including those encoding
tartrate-resistant acid phosphatase (TRAP) and enzymes involved in
cell fusion (9). Thus, inhibiting
RANKL signaling may suppress osteoclastogenesis.
Crocus sativus L. (saffron) belongs to the
Iridaceae family and has been utilized widely as a culinary spice,
an anodyne, or a tranquilizer in traditional Chinese medicine.
Crocin is a pharmacologically active component of Crocus
sativus L. Various studies have demonstrated that crocin
possesses a wide range of pharmacological effects, including
anti-arthritic, anti-inflammatory, anti-oxidant and anti-tumor
properties (10–13). Ding et al (14) demonstrated that crocin inhibited
interleukin (IL)-1β-induced activation of the NF-κB signaling
pathway by suppressing the degradation of inhibitor of κBα (IκBα)
in rabbit chondrocytes. In addition, crocin has been demonstrated
to restore cartilage and reduce bone deterioration, inflammation
and oxidative damage (10). A
recent study reported that administration of crocin may prevent
ovariectomy-induced osteoporosis in rats (15). However, the molecular mechanism
underlying the effect of crocin on osteoclast formation remains to
be investigated. The present study aimed to evaluate the effect of
crocin on RANKL-induced osteoclastogenesis and determine its
underlying molecular mechanism.
Materials and methods
Animals, reagents and antibodies
A total of 6 female C57BL/6 mice (age, 8–12 weeks
old) were obtained from the Animal Breeding Center of Shaanxi
Provincial People's Hospital (Xi'an, China) and maintained at a
constant temperature (21±2°C) and humidity in a holding facility
under a 12 h light/dark cycle, with free access to food and water.
Animal experiments were approved by the Institutional Animal Care
and Use Committee of Shaanxi Provincial People's Hospital (Shaanxi,
China). All reagent-grade chemicals, including crocin (purity
>95 %), fetal bovine serum (FBS), L-glutamine, penicillin,
streptomycin, recombinant macrophage-colony stimulating factor
(M-CSF), RANKL, α-minimum essential medium (α-MEM), TRAP Staining
kit, Cell Counting kit-8 (CCK8) and Bicinchoninic Acid (BCA)
Protein assay kit were purchased from Sigma-Aldrich; Merck
Millipore (Darmstadt, Germany). Radioimmunoprecipitation assay
(RIPA) lysis buffer was purchased from Beyotime Institute of
Biotechnology (Haimen, China). Polyvinylidene difluoride (PVDF)
membranes were purchased from EMD Millipore (Billerica, MA, USA).
Rabbit anti-mouse antibodies against nuclear factor of activated T
cells cytoplasmic 1 (NFATc1; 1:2,500; cat. no. sc-13033), c-Fos
(1:3,000; cat. no. sc-253), cathepsin K (1:1,500; cat. no.
sc-30056), IκBα (1:3,000; cat. no. sc-371), phosphorylated (p)-p65
(1:3,000; cat. no. sc-33020), p-c-Jun N-terminal kinase (JNK;
1:2,500; cat. no. sc-135642), JNK (1:3,000; cat. no. sc-572) and
GAPDH (1:3,000; cat. no. sc-25778) were purchased from Santa Cruz
Biotechnology, Inc., (Dallas, TX, USA). A goat anti-rabbit
horseradish peroxidase-conjugated IgG secondary antibody (1:3,000;
cat. no. A16104) and the Enhanced Chemiluminescence (ECL) reagent
were purchased from Invitrogen; Thermo Fisher Scientific, Inc.
(Waltham, MA, USA).
Cell culture
Primary bone marrow-derived macrophages (BMMs) were
prepared by removing bone marrow from the femora and tibiae of
C57BL/6 mice, and flushing the bone marrow cavity with α-MEM
supplemented with 10% FBS, 2 mM L-glutamine and 100 U/ml
penicillin/streptomycin. The following day, all non-adherent cells
were collected and considered to be osteoclast precursors. Mice
were sacrificed intraperitoneally with sodium pentobarbital (50
mg/kg; Sigma-Aldrich; Merck Millipore).
Osteoclast differentiation assay
During differentiation, BMMs were treated with 0,
10, 20 or 40 µM crocin at room temperature, and then cultured in
alpha-MEM supplemented with 10% FBS, 2 mM L-glutamine and 1%
penicillin/streptomycin in the presence of 50 ng/ml M-CSF and 50
ng/ml RANKL for 4 days. The control group was BMMs exposed to
medium alone at room temperature. Subsequently, osteoclast
formation was measured using a TRAP Staining kit according to the
manufacturer's protocol. Briefly, cells were fixed with 10%
formaldehyde in PBS for 3 min and stained with Naphthol AS-BI
phosphate and a tartrate solution for 1 h at 37°C, prior to
counterstaining with hematoxylin. Osteoclasts were considered to be
TRAP-positive if multinuclear staining (>3 nuclei) was observed
via light microscopy. The total number of TRAP-positive cells and
the number of nuclei per TRAP-positive cell in each well were
counted (data not shown).
Cell viability assay
Cell viability was measured with CCK8 according to
the manufacturer's protocol. Briefly, BMMs at a density of
1×104 cells/well were treated with 10, 20 or 40 µM
crocin for 3 days in 96-well plates. Subsequently, 10 µl CCK8
solution was added to each well for 2 h. The absorbance in each
well was measured at a wavelength of 490 nm using a microplate
reader (Omega Bio-Tek, Inc., Norcross, GA, USA).
Western blotting
Total protein was extracted from BMMs by using RIPA
lysis buffer according to the manufacturer's protocol. Lysates were
sonicated for 5 sec on ice and centrifuged at 6,000 × g for 5 min
at 4°C. Supernatants were collected and the protein concentration
was determined using a BCA assay kit. Equal quantities of protein
(40 µg per lane) were loaded onto 10% gels and subjected to sodium
dodecyl sulfate-polyacrylamide gel electrophoresis, prior to
transfer onto PVDF membranes. Membranes were blocked with 10%
fat-free milk in TBS (20 mmol/l Tris, 0.15 mol/l NaCl; pH 7.0)
containing 0.1% Tween-20 (TBST) for 1 h at room temperature prior
to probing with specific antibodies against NFATc1, c-Fos,
cathepsin K, p-JNK, JNK, IκBα, p-p65 and GAPDH at 4°C overnight.
Following washing with TBST, membranes were incubated with a
horseradish peroxidase-conjugated secondary antibody for 1 h at
room temperature. Proteins were visualized using an ECL reagent and
images were captured using the ImageQuant™ LAS 4000
imager (Fujifilm, Tokyo, Japan). Densitometry was performed using
Gel-Pro Analyzer software version 4.0 (Media Cybernetics, Inc.,
Rockville, MD, USA).
Statistical analysis
Data was analyzed using SPSS software version 13.0
(SPSS Inc., Chicago, IL, USA), and are presented as the mean ±
standard error of triplicate. Significant differences were analysed
using a Student's t-test or one-way analysis of variance followed
by Tukey's Honest Significant Difference post-hoc test. P<0.05
was considered to indicate a statistically significant
difference.
Results
Effect of crocin on cell
viability
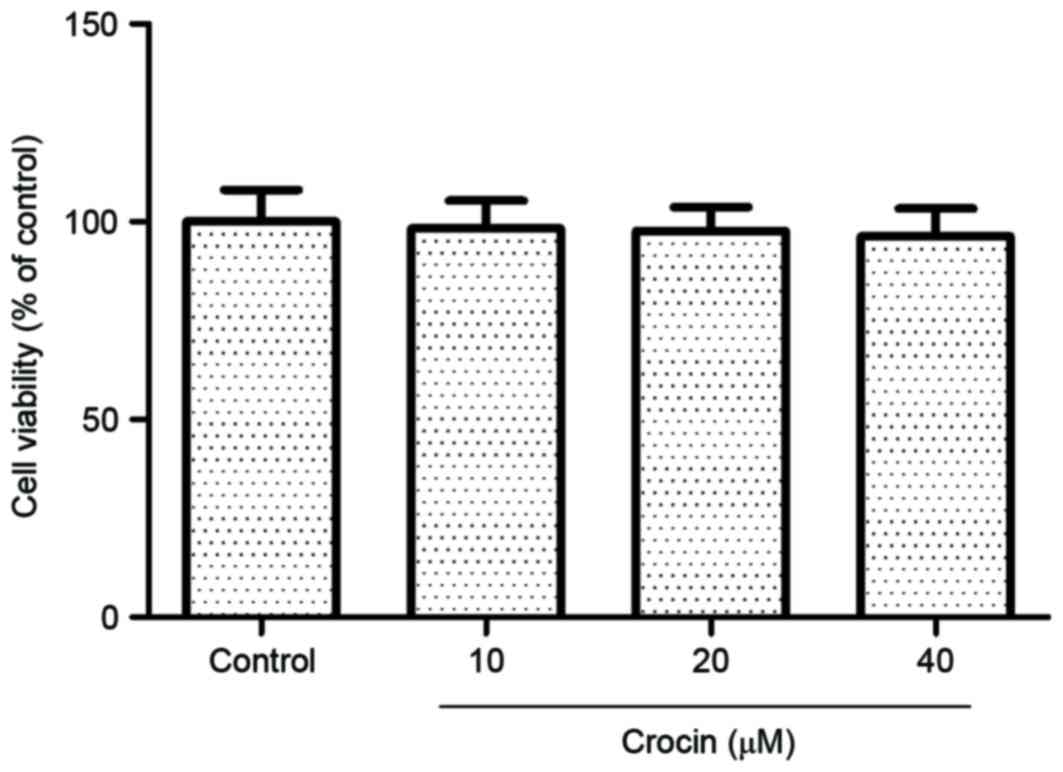
BMMs were treated with 10, 20 or 40 µM crocin for 72
h, and cell viability was assessed using a CCK8 assay. As presented
in Fig. 1, crocin treatment had no
cytotoxic effect on cells compared with the control group.
Crocin inhibits osteoclast formation
in BMMs
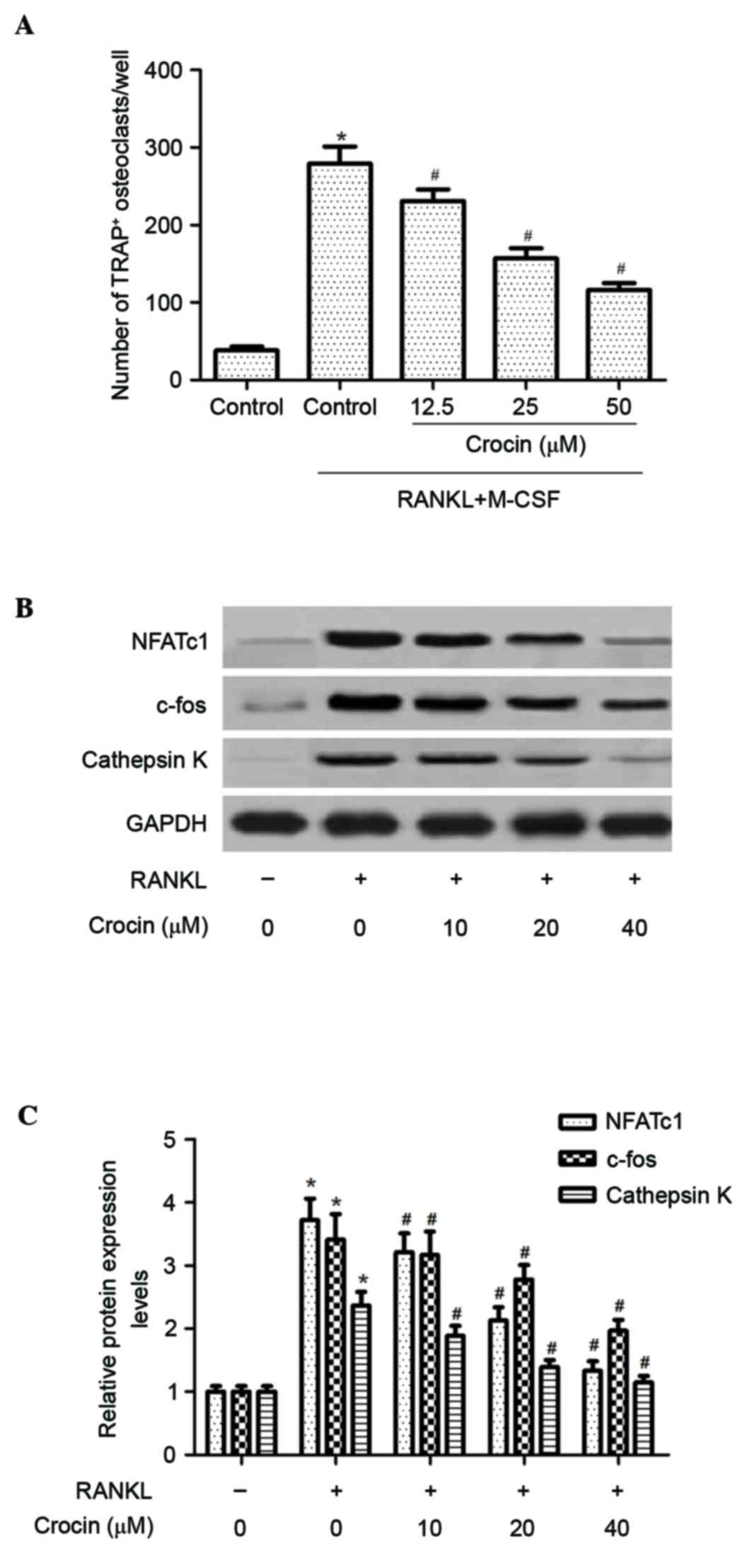
Subsequently, the effect of crocin on RANKL-induced
osteoclast differentiation from BMMs was investigated. As presented
in Fig. 2A, the number of
TRAP-positive multinuclear osteoclasts was significantly increased
following RANKL stimulation (P<0.05). However, crocin
significantly inhibited the formation of TRAP-positive multinuclear
osteoclasts in a dose-dependent manner (P<0.05).
Furthermore, the effect of crocin on the expression
of osteoclast-associated proteins, including NFATc1, c-Fos and
cathepsin K, was determined. Western blotting demonstrated that the
protein expression levels of NFATc1, c-Fos and cathepsin K were
increased following RANKL stimulation; crocin treatment
significantly inhibited the expression levels of these proteins in
BMMs compared with RANKL treatment alone (P<0.05), in a
dose-dependent manner (Fig. 2B and
C).
Crocin inhibits RANKL-induced
activation of JNK in BMMs
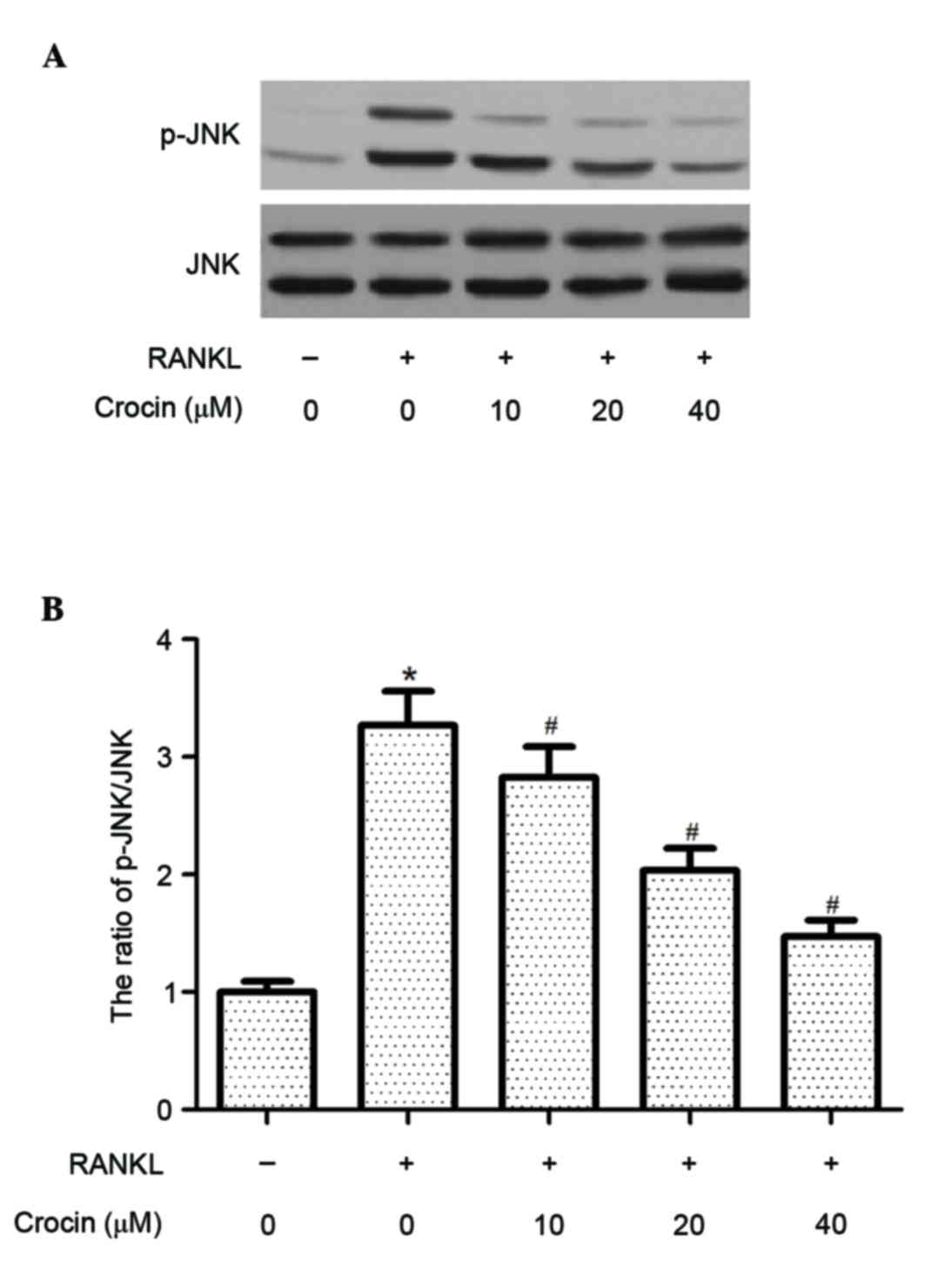
To investigate the molecular mechanisms underlying
the inhibitory effect of crocin on RANKL-induced osteoclast
formation, the effect of crocin on JNK activation in RANKL-induced
BMMs was examined. Western blotting demonstrated that the protein
expression levels of p-JNK were significantly increased following
RANKL stimulation (P<0.05), whereas crocin significantly
inhibited the RANKL-induced phosphorylation of JNK in BMMs
(P<0.05; Fig. 3).
Crocin inhibits RANKL-induced NF-κB
activation in BMMs
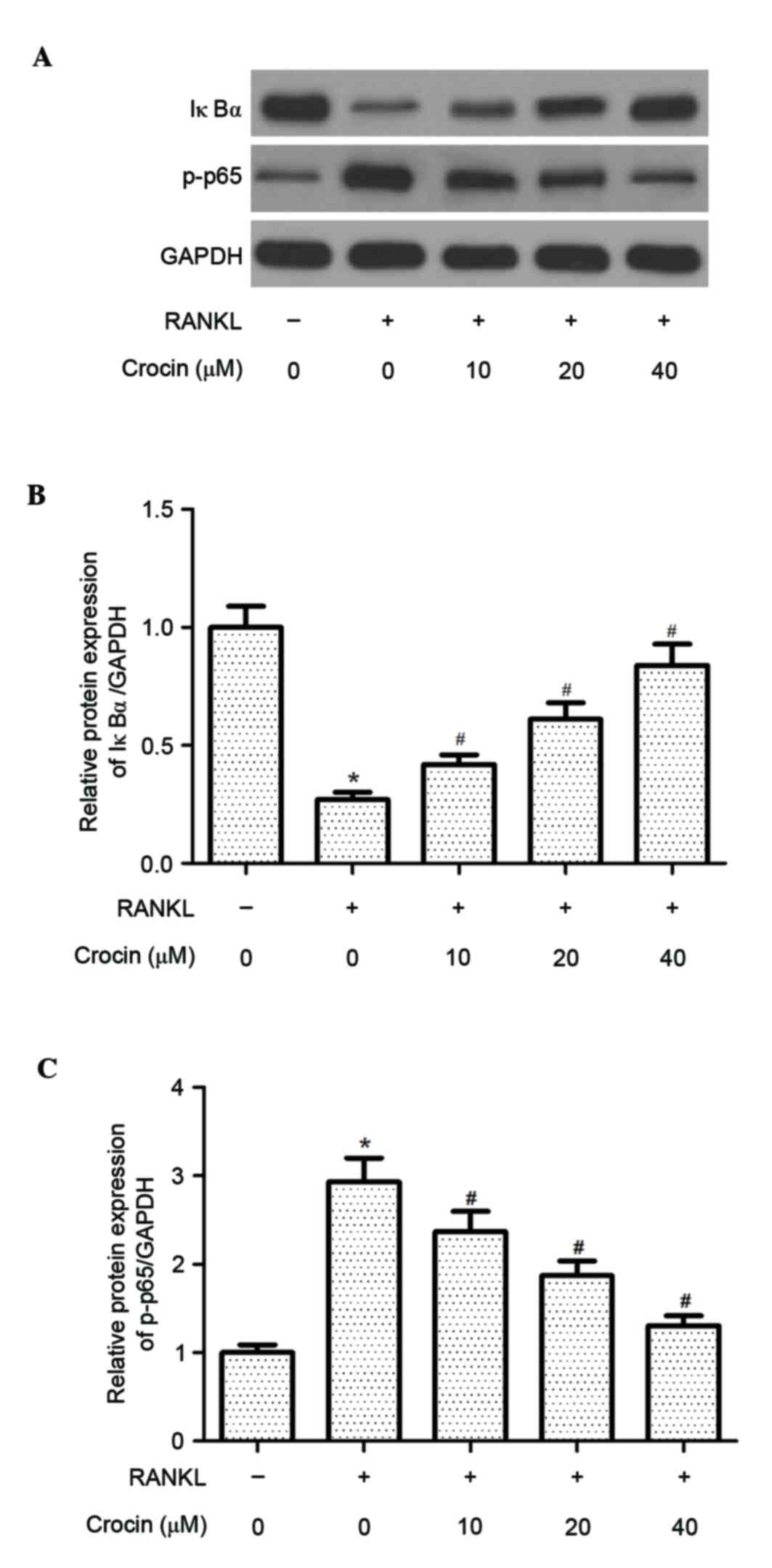
RANKL-induced NF-κB activation is essential for
osteoclast differentiation and function. Thus, the effect of crocin
on RANKL-induced NF-κB activation in BMMs was assessed. IκBα
protein expression levels were decreased following 1 h of RANKL
stimulation (P<0.05), whereas crocin significantly suppressed
the RANKL-induced degradation of IκBα in BMMs (P<0.05; Fig. 4). In addition, crocin inhibited the
RANKL-stimulated phosphorylation of NF-κB p65 in BMMs (P<0.05;
Fig. 4).
Discussion
The present study revealed that crocin inhibited the
osteoclastogenesis of BMMs, as the expression of osteoclast marker
proteins was downregulated by crocin. Mechanistically, crocin
inhibited RANKL-induced activation of NF-κB by suppressing IκBα
degradation and preventing NF-κB p65 nuclear translocation, and
activating JNK in BMMs.
To evaluate the effects of crocin on the formation
of osteoclasts, a bone marrow culture system was utilised to induce
the formation of osteoclasts in the presence of RANKL/M-CSF in
vitro. A previous study has reported that M-CSF induces the
proliferation of osteoclast precursor cells and their
differentiation into the osteoclast lineage, whereas RANKL induces
subsequent differentiation of osteoclast precursors into
osteoclasts (16). Consistent with
these results, the present study revealed that RANKL stimulation
induced the formation of osteoclasts; however, crocin treatment
inhibited the RANKL-induced formation of osteoclasts from
osteoclast precursors, as the number of TRAP-positive multinuclear
cells from BMMs was reduced following treatment with crocin. These
results suggested that crocin suppresses osteoclast differentiation
by acting directly on osteoclast precursors.
NFATc1, a key transcriptional factor downstream of
c-Fos, regulates the expression of TRAP, calcitonin receptor, and
cathepsin K during osteoclast differentiation (17). Takayanagi et al (18) reported that NFATc1-deficient
embryonic stem cells fail to differentiate into osteoclasts
following RANKL stimulation, and that ectopic expression of NFATc1
causes precursor cells to undergo differentiation. c-Fos serves an
essential role in the osteoclastic differentiation of precursor
cells generated by M-CSF and RANKL. c-Fos-deficient mice exhibit a
severe osteoporotic phenotype due to the absence of osteoclast
differentiation (19). The present
study demonstrated that crocin suppressed the expression of
osteoclast marker proteins in BMMs. These results suggested that
suppression of RANKL-induced activation of NFATc1 and c-Fos by
crocin is associated with the inhibition of osteoclastogenesis.
Previous studies have revealed that RANKL stimulates
JNK, leading to the activation of the transcription factor c-Jun
(20,21). Blockade of this signalling pathway
by treatment with a JNK inhibitor, SP600125, suppressed
RANKL-induced osteoclast differentiation (22). Furthermore, suppression of c-Jun
using small interfering RNA significantly inhibited RANKL-induced
osteoclast differentiation (23).
The present study demonstrated that crocin greatly inhibited
RANKL-induced phosphorylation of JNK in BMMs, which suggested that
the inhibitory effect of crocin on the differentiation of
osteoclast precursors into mature osteoclasts may be mediated by
the regulation of JNK phosphorylation.
The NF-κB signaling pathway serves an important role
in osteoclast formation (24–26).
It has been reported that NF-κB knockout mice exhibit severe
osteopetrosis and failed to stimulate osteoclastogenesis (27). Genetic inactivation of IκB kinase
(IKK) α or IKKβ is sufficient to inhibit osteoclastogenesis
(28). Furthermore, osteoclast
formation has been demonstrated to be mediated by RANKL-induced
NF-κB activation. For example, one study demonstrated that RANKL
induces TNF-α production primarily via the activation of NF-κB in
RAW264.7 cells (29). Therefore,
inhibition of NF-κB activity may be an effective strategy for the
treatment of osteoporosis. The present study revealed that crocin
greatly suppressed the RANKL-induced degradation of IκBα and
phosphorylation of NF-κB p65 in BMMs. This data supported the
hypothesis that crocin inhibits NF-κB activity in BMMs, resulting
in inhibition of osteoclast formation.
In conclusion, the results of the present study
suggested that crocin suppresses osteoclast differentiation via the
JNK and NF-κB signaling pathways. Thus, crocin may be a potential
therapeutic agent for the treatment of osteoclast-associated
diseases, including osteoporosis.
References
|
1
|
Hennemann A: Osteoporosis: Prevention,
diagnosis and therapy. Med Monatsschr Pharm. 25:164–167. 2002.(In
German). PubMed/NCBI
|
|
2
|
Burge R, Dawson-Hughes B, Solomon DH, Wong
JB, King A and Tosteson A: Incidence and economic burden of
osteoporosis-related fractures in the United States, 2005–2025. J
Bone Miner Res. 22:465–475. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Boyle WJ, Simonet WS and Lacey DL:
Osteoclast differentiation and activation. Nature. 423:337–342.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lacey DL, Timms E, Tan HL, Kelley MJ,
Dunstan CR, Burgess T, Elliott R, Colombero A, Elliott G, Scully S,
et al: Osteoprotegerin ligand is a cytokine that regulates
osteoclast differentiation and activation. Cell. 93:165–176. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Karst M, Gorny G, Galvin RJ and Oursler
MJ: Roles of stromal cell RANKL, OPG, and M-CSF expression in
biphasic TGF-beta regulation of osteoclast differentiation. J Cell
Physiol. 200:99–106. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Roodman GD: Regulation of osteoclast
differentiation. Ann N Y Acad Sci. 1068:100–109. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yasuda H, Shima N, Nakagawa N, Yamaguchi
K, Kinosaki M, Mochizuki S, Tomoyasu A, Yano K, Goto M, Murakami A,
et al: Osteoclast differentiation factor is a ligand for
osteoprotegerin/osteoclastogenesis-inhibitory factor and is
identical to TRANCE/RANKL. Proc Natl Acad Sci USA. 95:3597–3602.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Takayanagi H, Iizuka H, Juji T, Nakagawa
T, Yamamoto A, Miyazaki T, Koshihara Y, Oda H, Nakamura K and
Tanaka S: Involvement of receptor activator of nuclear factor
kappaB ligand/osteoclast differentiation factor in
osteoclastogenesis from synoviocytes in rheumatoid arthritis.
Arthritis Rheum. 43:259–269. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Asagiri M and Takayanagi H: The molecular
understanding of osteoclast differentiation. Bone. 40:251–264.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hemshekhar M, Santhosh Sebastin M, Sunitha
K, Thushara RM, Kemparaju K, Rangappa KS and Girish KS: A dietary
colorant crocin mitigates arthritis and associated secondary
complications by modulating cartilage deteriorating enzymes,
inflammatory mediators and antioxidant status. Biochimie.
94:2723–2733. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li K, Li Y, Ma Z and Zhao J: Crocin exerts
anti-inflammatory and anti-catabolic effects on rat intervertebral
discs by suppressing the activation of JNK. Int J Mol Med.
36:1291–1299. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Thushara RM, Hemshekhar M, Santhosh MS,
Jnaneshwari S, Nayaka SC, Naveen S, Kemparaju K and Girish KS:
Crocin, a dietary additive protects platelets from oxidative
stress-induced apoptosis and inhibits platelet aggregation. Mol
Cell Biochem. 373:73–83. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Aung HH, Wang CZ, Ni M, Fishbein A,
Mehendale SR, Xie JT, Shoyama CY and Yuan CS: Crocin from Crocus
sativus possesses significant anti-proliferation effects on
human colorectal cancer cells. Exp Oncol. 29:175–180.
2007.PubMed/NCBI
|
|
14
|
Ding Q, Zhong H, Qi Y, Cheng Y, Li W, Yan
S and Wang X: Anti-arthritic effects of crocin in
interleukin-1β-treated articular chondrocytes and cartilage in a
rabbit osteoarthritic model. Inflamm Res. 62:17–25. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cao PC, Xiao WX, Yan YB, Zhao X, Liu S,
Feng J, Zhang W, Wang J, Feng YF and Lei W: Preventive effect of
crocin on osteoporosis in an ovariectomized rat model. Evid Based
Complement Alternat Med. 2014:8251812014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Suda T, Takahashi N, Udagawa N, Jimi E,
Gillespie MT and Martin TJ: Modulation of osteoclast
differentiation and function by the new members of the tumor
necrosis factor receptor and ligand families. Endocr Rev.
20:345–357. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tanaka S, Nakamura K, Takahasi N and Suda
T: Role of RANKL in physiological and pathological bone resorption
and therapeutics targeting the RANKL-RANK signaling system. Immunol
Rev. 208:30–49. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Takayanagi H, Kim S, Koga T, Nishina H,
Isshiki M, Yoshida H, Saiura A, Isobe M, Yokochi T, Inoue JI, et
al: Induction and activation of the transcription factor NFATc1
(NFAT2) integrate RANKL signaling in terminal differentiation of
osteoclasts. Deve Cell. 3:889–901. 2002. View Article : Google Scholar
|
|
19
|
Arai A, Mizoguchi T, Harada S, Kobayashi
Y, Nakamichi Y, Yasuda H, Penninger JM, Yamada K, Udagawa N and
Takahashi N: Fos plays an essential role in the upregulation of
RANK expression in osteoclast precursors within the bone
microenvironment. J Cell Sci. 125:2910–2917. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kobayashi N, Kadono Y, Naito A, Matsumoto
K, Yamamoto T, Tanaka S and Inoue J: Segregation of TRAF6-mediated
signaling pathways clarifies its role in osteoclastogenesis. EMBO
J. 20:1271–1280. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Grigoriadis AE, Wang ZQ, Cecchini MG,
Hofstetter W, Felix R, Fleisch HA and Wagner EF: c-Fos: A key
regulator of osteoclast-macrophage lineage determination and bone
remodeling. Science. 266:443–448. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Huh JE, Lee WI, Kang JW, Nam D, Choi DY,
Park DS, Lee SH and Lee JD: Formononetin attunuates
osteoclastogenesis via suppressing the RANKL-induced activation of
NF-κB, c-Fos, and nuclear factor of activated T-cells cytoplasmic 1
signaling pathway. J Nat Prod. 77:2423–2431. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ikeda F, Nishimura R, Matsubara T, Tanaka
S, Inoue J, Reddy SV, Hata K, Yamashita K, Hiraga T, Watanabe T, et
al: Critical roles of c-Jun signaling in regulation of NFAT family
and RANKL-regulated osteoclast differentiation. J Clin Invest.
114:475–484. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Miyazaki T, Katagiri H, Kanegae Y,
Takayanagi H, Sawada Y, Yamamoto A, Pando MP, Asano T, Verma IM,
Oda H, et al: Reciprocal role of ERK and NF-kappaB pathways in
survival and activation of osteoclasts. J Cell Biol. 148:333–342.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lee ZH and Kim HH: Signal transduction by
receptor activator of nuclear factor kappa B in osteoclasts.
Biochem Biophys Res Commun. 305:211–214. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yamashita T, Yao Z, Li F, Zhang Q, Badell
IR, Schwarz EM, Takeshita S, Wagner EF, Noda M, Matsuo K, et al:
NF-kappaB p50 and p52 regulate receptor activator of NF-kappaB
ligand (RANKL) and tumor necrosis factor-induced osteoclast
precursor differentiation by activating c-Fos and NFATc1. J Biol
Chem. 282:18245–18253. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Novack DV, Yin L, Hagen-Stapleton A,
Schreiber RD, Goeddel DV, Ross FP and Teitelbaum SL: The IkappaB
function of NF-kappaB2 p100 controls stimulated osteoclastogenesis.
J Exp Med. 198:771–781. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chaisson ML, Branstetter DG, Derry JM,
Armstrong AP, Tometsko ME, Takeda K, Akira S and Dougall WC:
Osteoclast differentiation is impaired in the absence of inhibitor
of kappaB kinase alpha. J Biol Chem. 279:54841–54848. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zou W, Amcheslavsky A, Takeshita S, Drissi
H and Bar-Shavit Z: TNF-alpha expression is transcriptionally
regulated by RANK ligand. J Cell Physiol. 202:371–378. 2005.
View Article : Google Scholar : PubMed/NCBI
|