Introduction
Postoperative cognitive dysfunction (POCD) often
occurs in the elderly after undergoing anesthesia and surgery, and
is associated with increased perioperative complications and
mortality rate. According to recent studies, neuroinflammation
(1), blood-brain barrier
disruption (2,3), activation of the neurotrophin
receptor (4) and depression of
synaptic function (5) are likely
involved in the pathogenesis of POCD, but the etiology remains
unknown despite enormous research efforts. Clinical observational
studies revealing that POCD arises more frequently after surgery in
the elderly are consistent with an inflammatory component (6). Furthermore, animal studies have shown
that proinflammatory cytokines can cause neurological disorders
(1,7). Thus, it is possible that inflammation
plays a substantial role in the pathogenesis of POCD.
The commonly used inhalation anesthetic isoflurane
has been shown to induce activation of nuclear factor (NF)-κB, a
master regulator of inflammation (8), increase TNF-α, IL-6 and IL-1β in the
aged brain (9), and is associated
with increased risk of cognitive dysfunction in the elderly.
Hypoxia-inducible factor 1α (HIF-1α) is upregulated by factors such
as angiotensinII (10) and
carbachol (11), even though
HIF-1α plays a major role in the maintenance of oxygen homeostasis
and cellular adaptation (12). In
addition, isoflurane transiently increases HIF-1α protein in Hep3B
cells (13), myocardial tissue
(14) and renal organs (15).
Recent studies have indicated extensive interaction
between NF-κB and HIF-1α, the two main molecular players involved
in hypoxia and inflammation. It has been suggested that HIF-1α
promotes NF-κB activity (16), and
that activation of HIF-1α may simultaneously suppress the activity
of NF-κB (17). However, it has
also been reported that NF-κB regulates HIF-1α transcription in
pulmonary artery smooth muscle cells during hypoxia (18). At present, this interaction between
NF-κB and HIF-1α in POCD of the aging brain caused by isoflurane
anesthesia has proven elusive. Based on the above findings, we
preliminarily investigated whether HIF-1α plays a role in
isoflurane-induced cognitive dysfunction and whether the HIF-1α
inhibitor, 5-[1-(phenylmethyl)-1H-indazol-3-yl]-2-furanmethanol
(YC-1), can improve isoflurane-induced cognitive dysfunction by
preventing inflammation in aged rats. In addition, we observed a
marker of the NF-κB signaling pathway in the hippocampus of aged
rats, and the expression levels of IL-1β and TNF-1α for 7 days
following 4-h 1.5% isoflurane exposure.
Materials and methods
Animal model
Male Sprague-Dawley rats (n=144, 20 months of age
and weighing 500–600 g) were purchased from the Dongchuang
Laboratory Animal Center (Changsha, Hunan, China) and housed in
standard barrier facilities. They were maintained under a 12-h
light/dark cycle (lights on at 07:00) with food and water ad
libitum. The animals had a recovery period of at least 7 days
to adapt to their new environment before experiments began.
Isoflurane exposure was performed as previously
described (2). In a transparent
anesthetic chamber, rats were exposed to 1.5% isoflurane (Baxter
Healthcare, Deerfield, IL, USA) with 2 l/min of 100% oxygen
(Beijing Millennium City Gas Sales Center, China) as the carrying
gas, or to vehicle (2 l/min of 100% oxygen) for 4 h. At the outlet
of the chamber, gas composition (the concentrations of isoflurane,
oxygen, and carbon dioxide) within the chamber was continuously
analyzed by a gas monitor (Datex-Ohmeda, Inc., Louisville, CO,
USA). After anesthesia, the rats received 100% oxygen until they
recovered complete consciousness. In a previous study, it has been
shown that this anesthesia protocol does not cause significant
changes in glucose or blood gas (1).
Experiment protocols
Rats were randomly assigned to isoflurane (n=6) or
control (n=6) groups, and were exposed to isoflurane or vehicle
gas, respectively. Expression levels of hippocampal HIF-1α proteins
were examined by western blotting and immunofluorescence to
investigate changes in HIF-1α protein after 4-h isoflurane
exposure. Based on preliminary results, the HIF-1α inhibitor YC-1
(cat. no. 170632-47-0; Santa Cruz Biotechnology, Inc., Dallas, TX,
USA) was used for intervention studies. Subsequently, rats were
randomly assigned to control, ISO, YC-1 + ISO, and YC-1 groups (n=6
per group). Rats in the YC-1 + ISO and YC-1 groups received 2 mg/kg
of intraperitoneal YC-1 at 24 h 30 min before gas exposure. The
solution of YC-1 in 1% dimethyl sulfoxide (DMSO) was freshly
prepared before use, and 0.5 ml was injected. The rats in the other
two groups received 0.5 ml 1% DMSO. This YC-1 dosing protocol
effectively inhibits HIF-1α activation in rats (19).
The next day after 4-h isoflurane anesthesia or
vehicle gas exposure, hippocampal-dependent spatial memory ability
was evaluated using the Morris water maze (MWM) test (n=12 per
group). As the most well-characterized inhibitor of NF-κB (20,21),
the expression of IκBa and p-IκBa protein in the hippocampus of
aged rats was assessed immediately after isoflurane exposure, and a
1 day (1 day), 3 day, and 7 day after isoflurane exposure using
western blotting (n=4 per time point). The expression levels of
TNF-1α and IL-1β were also examined at the same time points using
enzyme-linked immunosorbent assay (ELISA; n=4 per time point).
Morphology
Immunofluorescence staining of brain sections were
performed as previously described (17) using an anti-HIF-1α primary antibody
(dilution, 1:500; cat. no. 13515; Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA) and a fluorescein isothiocyanate-labeled secondary
antibody (dilution, 1:200; cat. no. ab7010; Abcam, Cambridge, UK).
The nuclei were counterstained with mounting medium with 4,
6-diamidino-2-phenylindole (1:5,000; Roche Applied Science,
Mannheim, Germany).
Western blot analysis
The protein quantity of samples was determined using
western blot analysis. We used the following antibodies:
anti-HIF-1α (dilution, 1:500; cat. no. 13515; Santa Cruz
Biotechnology, Inc.); anti-Iκ-Bα and anti-p-IκBa (dilution,
1:1,000; cat. no. 4812 and 9246; Cell Signaling Technology, Inc.,
Danvers, MA, USA) and anti-β-actin (dilution, 1:10,000; cat. no.
4970; Santa Cruz Biotechnology, Inc.) primary antibodies overnight
at 4°C, followed by incubation with the IRDye®
800CW-conjugated goat anti-rabbit secondary antibody (dilution,
1:10,000; cat. no. 926-32211; LI-COR Biosciences, Inc., Lincoln,
NE, USA) for 2 h at room temperature.
ELISA
Homogenates from the hippocampus were centrifuged at
12,000 rpm for 10 min at 4°C as described previously (14), and the supernatants were collected.
The concentrations of IL-1β and TNF-1α were measured with an ELISA
kit (cat. no. ab100768 and ab46070; Abcam), using 100 µl of protein
for detection according to the manufacturer's instructions. Each
experimental condition was tested in three different wells and
measured in duplicate.
MWM test
As previously described (2), four groups with 12 rats per group
were tested for spatial learning and memory using the MWM test by
investigators blinded to the group conditions. Briefly, the rats
received four training trials daily for 5 consecutive days. During
each trial, the rats were gently placed in the water facing the
wall of the maze at one of the four equally spaced start positions
(north, south, east, or west). The time spent to locate the
submerged platform (defined by the latency cut-off time of 120 sec)
and the swimming velocity was recorded. On day 6, a series of probe
trials were conducted without the platform. The platform site
crossovers and the percentage of time spent in the previous
platform quadrant in a 90-sec period were determined.
Statistical analysis
For statistical analyses, SPSS 14.0 for Windows
(SPSS, Chicago, IL, USA) was used. The values for physiological
parameters and data obtained by western blotting analysis and ELISA
were expressed as the mean ± SD, and analyzed by one-way analysis
of variance (ANOVA), followed by a l Fisher's least significant
difference multiple comparison test. Data collected from the
behavioral studies were expressed as the mean ± SEM, and analyzed
by two-way repeated-measures ANOVA, with Bonferroni post hoc
analysis. P<0.05 was considered to indicate a statistically
significant difference.
Results
HIF-1α protein expression is induced
by isoflurane exposure, and degraded by YC-1
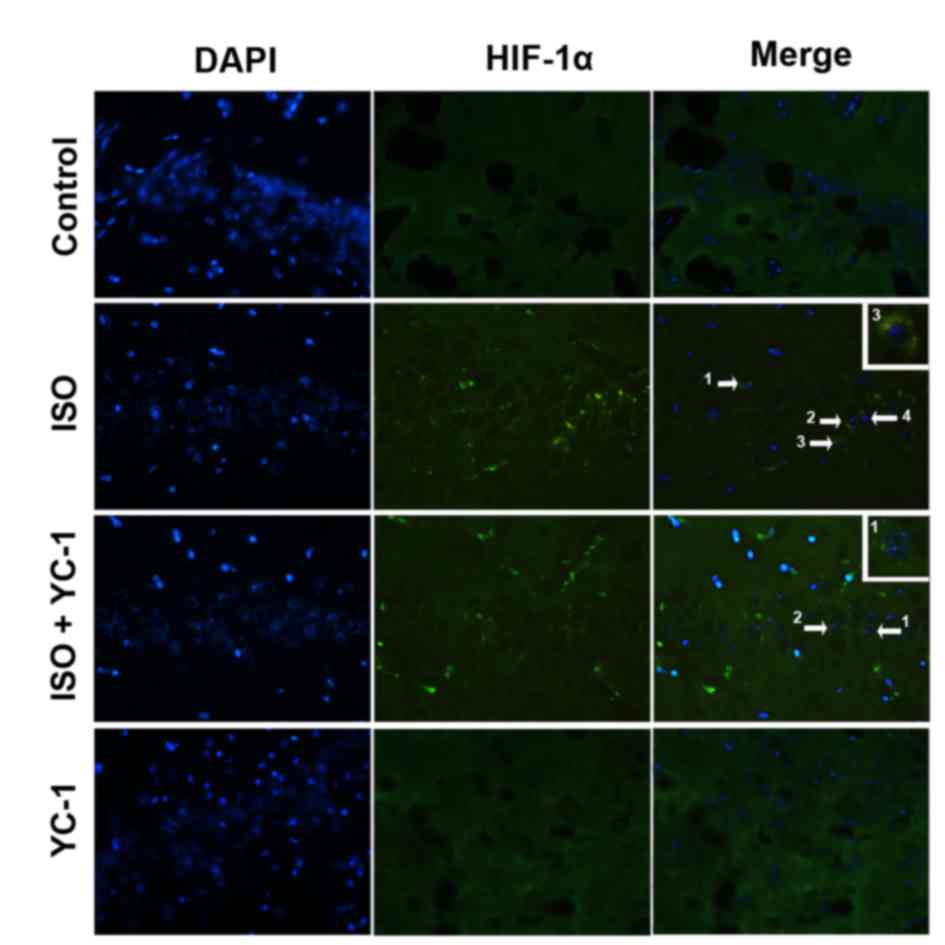
We used immunofluorescence staining to investigate
the nuclear translocation of HIF-1α. The increased HIF-1α was
mainly localized in neuronal nuclei in the hippocampus after a 4-h
isoflurane exposure compared with vehicle gas, but then it was
attenuated by YC-1 pretreatment (Fig.
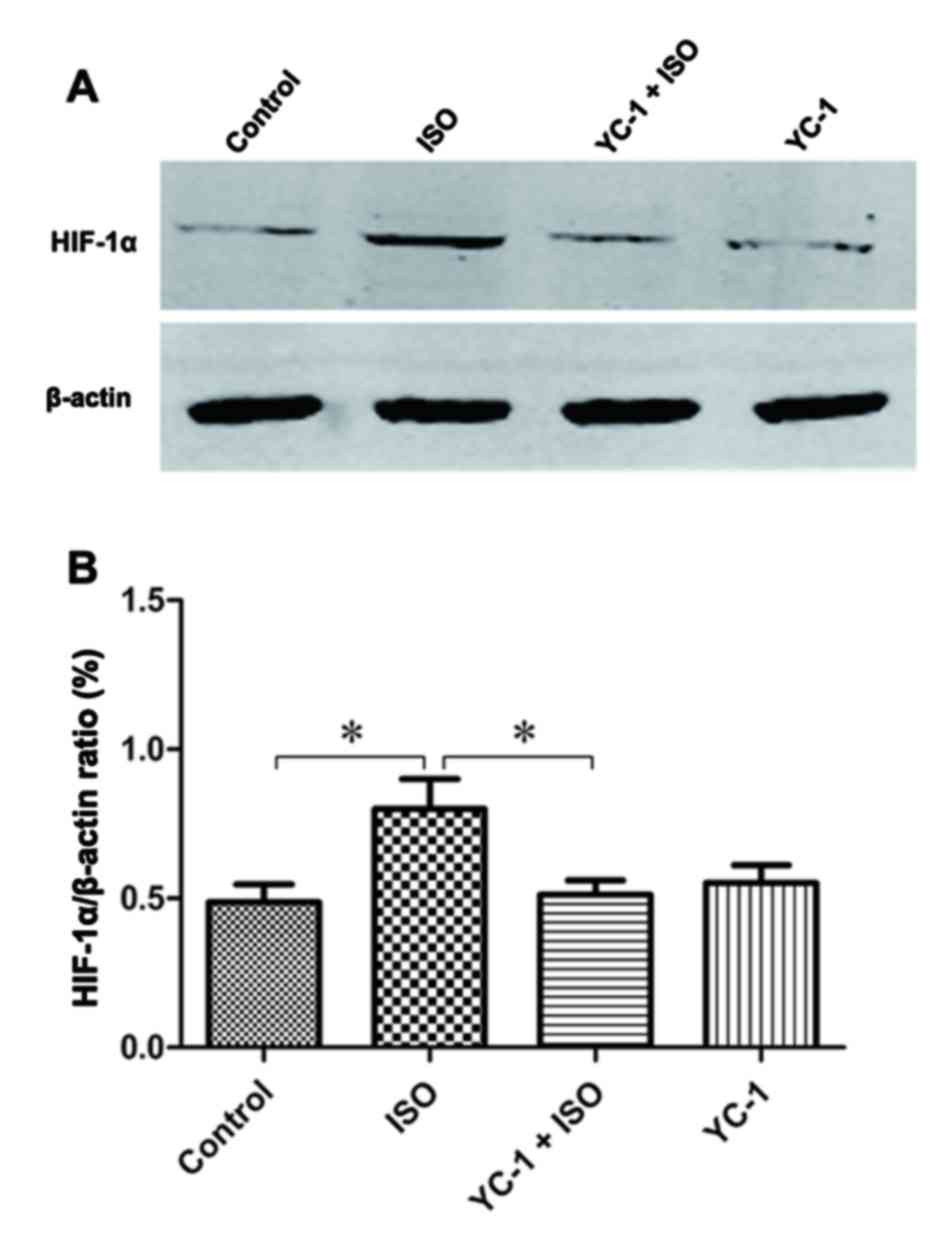
1). Similarly, compared with control rats, the expression of
HIF-1α protein increased after 4-h isoflurane exposure, and YC-1
pretreatment prevented the isoflurane-induced increase (Fig. 2). These changes indicate a
transient increase in HIF-1α after the 4-h isoflurane challenge,
and suggest YC-1 induces HIF-1α degradation.
YC-1 disturbs isoflurane-induced
activation of the NF-κB signaling pathway
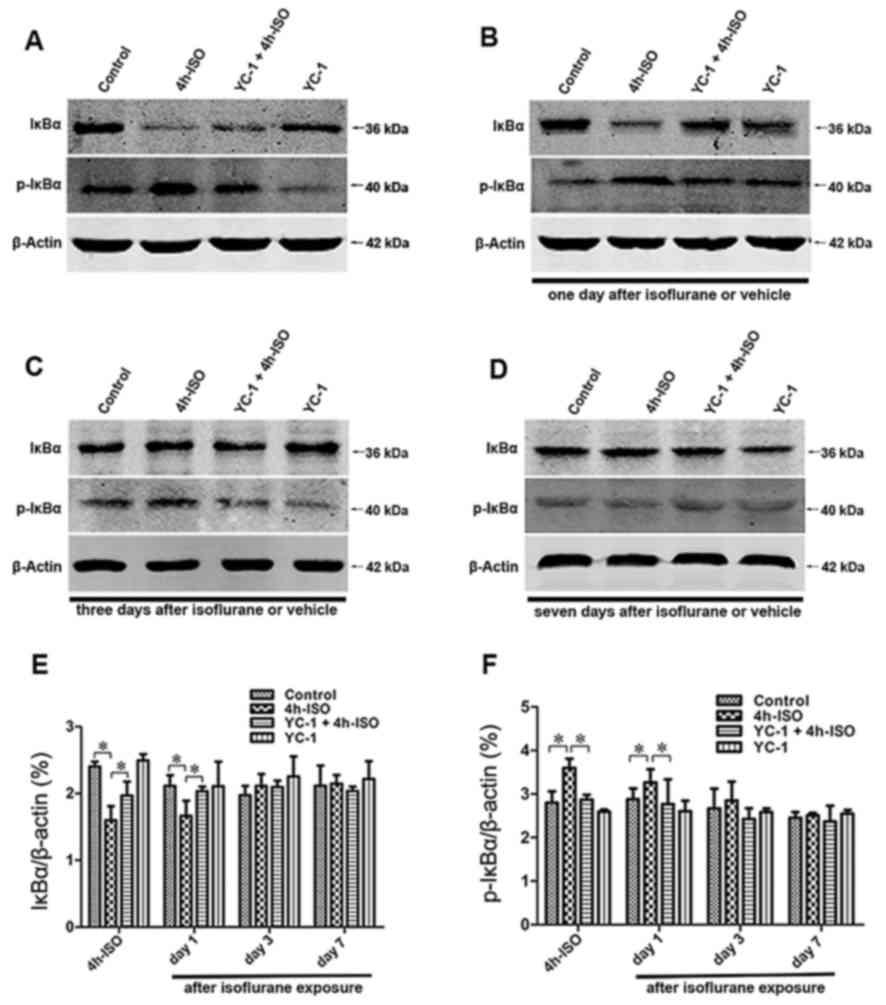
The expression of IκBα inhibited the activation of
NF-κB signaling pathways (20),
thus, hippocampal IκBα and p-IκBα protein expression following
isoflurane exposure was determined (Fig. 3). The IκBa phosphorylation levels
were significantly increased, and the protein levels of IκBa
significantly decreased after 4-h isoflurane exposure.
Specifically, these changes were detected immediately after
isoflruane exposure, and then recovery to the baseline levels
within 3 days. Western blotting demonstrated that YC-1 pretreatment
significantly prevented the isoflurane-induced the decrease in IκBa
and increase in p-IκBa protein levels in the hippocampus. The
results suggest that the NF-κB pathway was activated in the
hippocampus of aged rats, and was altered by YC-1 pretreatment
after a 4-h isoflurane challenge.
Isoflurane exposure-induced TNF-1α
accumulation is suppressed by YC-1
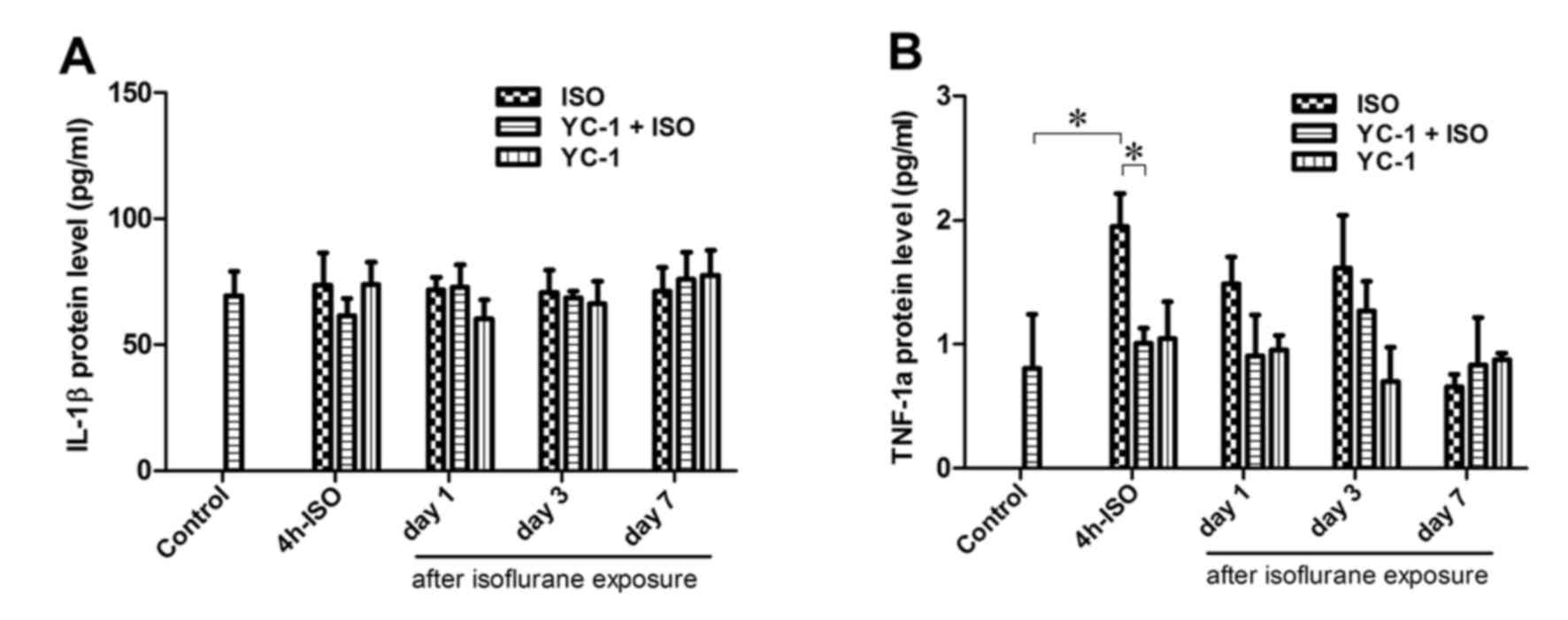
TNF-1α expression was significantly increased
immediately after 4-h isoflurane exposure, and then decreased to
the baseline level within 24 h after anesthesia. The
isoflurane-induced increase in TNF-1α was inhibited by YC-1
(Fig. 4). IL-1β was also examined
(Fig. 4), but showed no
significant alteration after isoflurane exposure. When administered
alone, YC-1 had no effect on the expression of TNF-1α or IL-1β at 4
h after anesthesia.
Isoflurane exposure-induced cognitive
impairment is attenuated by YC-1
Cognitive performance was assessed using the MWM.
There was no significant difference in swim speed among the four
groups (data not shown). During the 6-day training period, aged
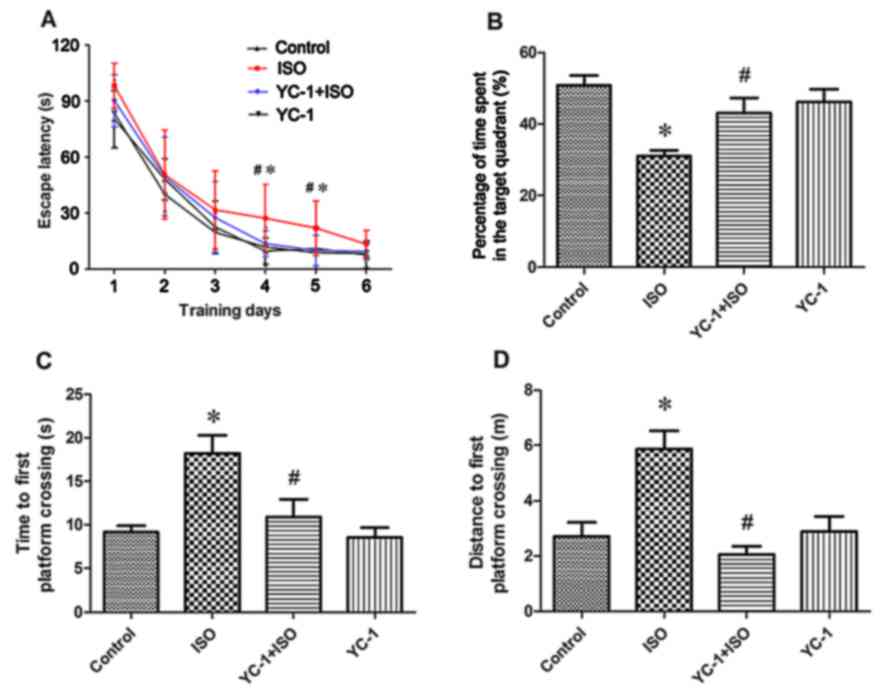
rats in the ISO group showed longer escape latencies compared with
the control group at days 4 and 5 (Fig. 5). Rats in the YC-1 + ISO group
required less time to find the platform than those in the ISO group
on days 4 and 5 of training (Fig.
5A). There were no significant differences in latencies between
the control and YC-1 groups (Fig.
5A). During the probe test, which evaluated memory in the rats,
the latency to the first entrance in the targeted area in the YC-1
+ ISO group was much shorter (Fig.
5C), and the percentages of time and distance in the targeted
area were much higher than those in the ISO group (Fig. 5B and D). These findings indicate
that there were memory impairments after isoflurane exposure, but
that YC-1 pretreatment could significantly prevent these
impairments in spatial learning and memory.
Discussion
In the present study, we demonstrated that YC-1
pretreatment can protect against cognitive impairment in aged rats
after 4-h isoflurane exposure. Specifically, pretreatment with YC-1
significantly attenuated the increase in HIF-1α protein caused by
longer isoflurane exposure. Furthermore, YC-1-induced attenuation
of HIF-1α expression appeared to significantly disturb the
isoflurane-induced NF-κB activation and decrease the expression of
inflammatory cytokines in vivo.
The expression of HIF-1α protein maintain its
balance via protein synthesis and degradation in physiological
conditions (22). In this study,
we showed that exposure to clinically relevant concentrations of
isoflurane enhanced expression of HIF-1α in the hippocampus of aged
rats, and demonstrated that isoflurane disrupted the cognition of
aged rats, which might be partly mediated by the HIF-1α pathway.
Whereas, the defined molecular mechanisms by which isoflurane
up-regulate HIF-1α is remains unknown, previous studies have
reported that isoflurane activation of the Akt-mTOR pathway induced
HIF-1α expression in vivo (14), and the levels of phospho-Akt and
GSK 3β also played roles in isoflurane-induced HIF-1α expression
in vitro (23). Our data
confirmed that 4 h-isoflurane exposure resulted in up-regulation of
HIF-1α without hypoxia interventions, moreover, this is the first
experiment to demonstrate that the interaction between NF-κB and
HIF-1α pathway in isoflurane-induced cognitive decline in aging
brain.
Recent evidence has shown that isoflurane may
increase the levels of proinflammatory cytokines in the brain of
aged rats, and that this process plays a critical role in the
progression of cognitive dysfunction (1,9).
Other studies have reported that HIF-1α may be considered
proinflammatory in that it promotes inflammatory cell survival
(24). In the present study,
isoflurane-induced HIF-1α protein expression in aged rat brain was
inhibited by YC-1, and the canonical NF-κB signaling pathway was
transiently activated, as evidenced by the increased expression of
p-IκBa protein and decreased expression of IκBa protein in the
hippocampus immediately after isoflurane exposure, which persisted
until 24 h. Moreover, isoflurane-induced IκBa was increased with
YC-1 pretreatment, as the product of the NFKBIA gene, IκBa is
thought to be an inhibitor of NF-κB due to its ability to bind to
the p65/p50 dimers, preventing them from translocating into the
nucleus, and thus counteracting NF-κB signaling (20,21).
In addition, studies have reported a positive correlation between
the NF-κB signaling pathway and TNF-1α (1,25).
TNF-1α protein expression was dramatically decreased after
depleting HIF-1α in the present study, therefore, we speculate that
NF-κB signaling pathway activity may be suppressed through
inactivation of the HIF-1α protein. Conversely, IL-1β was not
changed, which may be because IL-1β contents were measured in the
hippocampus of rats exposed to isoflurane at different time points,
which was increased from 1 to 6 h after isoflurane exposure
(1). The findings suggest that
inhibition of HIF-1α by YC-1 disrupted NF-κB signaling pathway
activity, suppressed its downstream TNF-1α expression and mitigated
the isoflurane-induced cognitive impairment.
Previous studies have shown that hippocampal
apoptosis increased when HIF-1α was inhibited by YC-1, suggesting
that HIF-1α has a neuroprotective effect against the cognitive
function deteriorated induced by subarachnoid hemorrhage of rats
(26). Contrarily, it has been
reported that YC-1 protects neurons from the toxicity associated
with Aβ25–35 peptides is mainly mediated by the induction of Hsp70
(27), and YC-1 also against
ketamine-induced long-term cognitive defects in neonatal rats via
the ROS/HIF-1α pathway (28).
Thus, because of the complexity of such regulations, HIF-1α may
play different roles under different conditions, and its activation
may even prove to be a promising intervention against
isoflurane-induced POCD. Our results showed improvement of
isoflurane-induced spatial learning and memory deficits with HIF-1α
inhibitor pretreatment. However, the pathophysiology of the
cognitive deficits is complicated; other signaling pathways or
mechanisms activated by YC-1 might also be responsible for
improving cognitive function during the perioperative period. This
will require further investigation. The current findings lead to a
better understanding of the underlying mechanisms of POCD in the
aged brain.
In conclusion, we report that isoflurane-induced
spatial cognitive impairment in aged rats was associated with
expression of the HIF-1α protein. Suppressing HIF-1α protein
expression attenuated the neuroinflammation in the hippocampus by
disturbing NF-κB signaling pathway activity and decreasing TNF-1α
after 4-h isoflurane exposure. These data suggest that HIF-1α may
be an upstream regulator of the NF-κB signaling pathway in POCD,
and inhibition of this pathway may serve as a potential treatment
target.
Acknowledgements
Not applicable.
Funding
This work was supported by grants from the National
Natural Science Foundation of China (grant no. 81371205 and
81571036) (awarded to Dr Xiangyang Guo, Peking University Third
Hospital, Beijing, China), and the Shanghai University of Medicine
and Health Sciences Basic Research Program of China (grant no.
HMSF-17-21-019) (awarded to Dr Yiyun Cao, the Sixth People's
Hospital East Campus, Shanghai University, of Medicine and Health
Sciences, Shanghai, China).
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
YC, XG and ZL designed and performed experiments;
YC, LM, CN, LL and NY performed the experiments; YC and CS analyzed
the data; YC and XG wrote the paper. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
The present study was performed following the
approval of Peking University Biomedical Ethics Committee
Experimental Animal Ethics Branch (approval no. 20150041).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Li ZQ, Rong XY, Liu YJ, Ni C, Tian XS, Mo
N, Chui DH and Guo XY: Activation of the canonical nuclear
factor-κB pathway is involved in isoflurane-induced hippocampal
interleukin-1β elevation and the resultant cognitive deficits in
aged rats. Biochem Biophys Res Commun. 438:628–634. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cao Y, Ni C, Li Z, Li L, Liu Y, Wang C,
Zhong Y, Cui D and Guo X: Isoflurane anesthesia results in
reversible ultrastructure and occludin tight junction protein
expression changes in hippocampal blood-brain barrier in aged rats.
Neurosci Lett. 587:51–56. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li Z, Cao Y, Li L, Liang Y, Tian X, Mo N,
Liu Y, Li M, Chui D and Guo X: Prophylactic angiotensin type 1
receptor antagonism confers neuroprotection in an aged rat model of
postoperative cognitive dysfunction. Biochem Biophys Res Commun.
449:74–80. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Schallner N, Ulbrich F, Engelstaedter H,
Biermann J, Auwaerter V, Loop T and Goebel U: Isoflurane but not
sevoflurane or desflurane aggravates injury to neurons in vitro and
in vivo via p75NTR-NF-kB activation. Anesth Analg. 119:1429–1441.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li L, Li Z, Cao Y, Fan D, Chui D and Guo
X: Increased extrasynaptic GluN2B expression is involved in
cognitive impairment after isoflurane anesthesia. Exp Ther Med.
12:161–168. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lemstra AW, Kalisvaart KJ, Vreeswijk R,
van Gool WA and Eikelenboom P: Pre-operative inflammatory markers
and the risk of postoperative delirium in elderly patients. Int J
Geriatr Psychiatry. 23:943–948. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhu J, Jiang Y, Wu L, Lu T, Xu G and Liu
X: Suppression of local inflammation contributes to the
neuroprotective effect of ginsenoside Rb1 in rats with cerebral
ischemia. Neuroscience. 202:342–351. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li Z, Ni C, Xia C, Jaw J, Wang Y, Cao Y,
Xu M and Guo X: Calcineurin/nuclear factor-κB signaling mediates
isoflurane-induced hippocampal neuroinflammation and subsequent
cognitive impairment in aged rats. Mol Med Rep. 15:201–209. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wu X, Lu Y, Dong Y, Zhang G, Zhang Y, Xu
Z, Culley DJ, Crosby G, Marcantonio ER, Tanzi RE and Xie Z: The
inhalation anesthetic isoflurane increases levels of
proinflammatory TNF-α, IL-6, and IL-1β. Neurobiol Aging.
33:1364–1378. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dai GH, Ma PZ, Song XB, Liu N, Zhang T and
Wu B: MicroRNA-223-3p inhibits the angiogenesis of ischemic cardiac
microvascular endothelial cells via affecting RPS6KB1/hif-1a signal
pathway. PLoS One. 9:e1084682014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hui Y, Zhao Y, Ma N, Peng Y, Pan Z, Zou C,
Zhang P and Du Z: M3-mAChR stimulation exerts anti-apoptotic effect
via activating the HIF-1α/HO-1/VEGF signaling pathway in H9c2 rat
ventricular cells. J Cardiovasc Pharmacol. 60:474–482. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Semenza GL: HIF-1 and mechanisms of
hypoxia sensing. Curr Opin Cell Biol. 13:167–171. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li QF, Wang XR, Yang YW and Su DS:
Up-regulation of hypoxia inducible factor 1alpha by isoflurane in
Hep3B cells. Anesthesiology. 105:1211–1219. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Raphael J, Zuo Z, Abedat S, Beeri R and
Gozal Y: Isoflurane preconditioning decreases myocardial infarction
in rabbits via up-regulation of hypoxia inducible factor 1 that is
mediated by mammalian target of rapamycin. Anesthesiology.
108:415–425. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Benzonana LL, Perry NJ, Watts HR, Yang B,
Perry IA, Coombes C, Takata M and Ma D: Isoflurane, a commonly used
volatile anesthetic, enhances renal cancer growth and malignant
potential via the hypoxia-inducible factor cellular signaling
pathway in vitro. Anesthesiology. 119:593–605. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Walmsley SR, Print C, Farahi N,
Peyssonnaux C, Johnson RS, Cramer T, Sobolewski A, Condliffe AM,
Cowburn AS, Johnson N and Chilvers ER: Hypoxia-induced neutrophil
survival is mediated by HIF-1alpha-dependent NF-kappaB activity. J
Exp Med. 201:105–115. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Carbia-Nagashima A, Gerez J, Perez-Castro
C, Paez-Pereda M, Silberstein S, Stalla GK, Holsboer F and Arzt E:
RSUME, a small RWD-containing protein, enhances SUMO conjugation
and stabilizes HIF-1alpha during hypoxia. Cell. 131:309–323. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Belaiba RS, Bonello S, Zähringer C,
Schmidt S, Hess J, Kietzmann T and Görlach A: Hypoxia up-regulates
hypoxia-inducible factor-1alpha transcription by involving
phosphatidylinositol 3-kinase and nuclear factor kappaB in
pulmonary artery smooth muscle cells. Mol Biol Cell. 18:4691–4697.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hieber S, Huhn R, Hollmann MW, Weber NC
and Preckel B: Hypoxia-inducible factor 1 and related gene products
in anaesthetic-induced preconditioning. Eur J Anaesthesiol.
26:201–206. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Karin M: Nuclear factor-kappaB in cancer
development and progression. Nature. 441:431–436. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xia Y, Shen S and Verma IM: NF-κB, an
active player in human cancers. Cancer Immunol Res. 2:823–830.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cummins EP, Keogh CE, Crean D and Taylor
CT: The role of HIF in immunity and inflammation. Mol Aspects Med.
47–48:24–34. 2016. View Article : Google Scholar
|
|
23
|
Sun Y, Li QF, Zhang Y, Hu R and Jiang H:
Isoflurane preconditioning increases survival of rat skin
random-pattern flaps by induction of HIF-1α expression. Cell
Physiol Biochem. 31:579–591. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cramer T, Yamanishi Y, Clausen BE, Förster
I, Pawlinski R, Mackman N, Haase VH, Jaenisch R, Corr M, Nizet V,
et al: HIF-1alpha is essential for myeloid cell-mediated
inflammation. Cell. 112:645–657. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Janyou A, Wicha P, Jittiwat J, Suksamrarn
A, Tocharus C and Tocharus J: Dihydrocapsaicin attenuates blood
brain barrier and cerebral damage in focal cerebral
ischemia/reperfusion via oxidative stress and inflammatory. Sci
Rep. 7:105562017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Dong Y, Li Y, Feng D, Wang J, Wen H, Liu
D, Zhao D, Liu H, Gao G, Yin Z and Qin H: Protective effect of
HIF-1α against hippocampal apoptosis and cognitive dysfunction in
an experimental rat model of subarachnoid hemorrhage. Brain Res.
1517:114–121. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tsai YC, Lee YM, Lam KK, Lin JF, Wang JJ,
Yen MH and Cheng PY: The role of heat shock protein 70 in the
protective effect of YC-1 on β-amyloid-induced toxicity in
differentiated PC12 cells. PLoS One. 8:e693202013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yan J, Huang Y, Lu Y, Chen J and Jiang H:
Repeated administration of ketamine can induce hippocampal
neurodegeneration and long-term cognitive impairment via the
ROS/HIF-1α pathway in developing rats. Cell Physiol Biochem.
33:1715–1732. 2014. View Article : Google Scholar : PubMed/NCBI
|