Introduction
Psoriasis is a chronic inflammatory skin disease
characterized by erythema, pimples, patches and silver scales
(1). It has been demonstrated that
psoriasis is a disorder resulting from a combination of mutations
in multiple genes and interaction between numerous other factors,
including trauma, infection and drugs (2). The primary pathological features of
psoriasis include overgrowth and abnormal differentiation of
epidermal keratinocytes, expansion of dermal papillary capillaries
and infiltration of inflammatory cells, among which keratinocyte
dysfunction serves a role in the pathogenesis of psoriasis
(2,3).
Keratinocytes are able to secrete a number of
cytokines, resulting in a local inflammatory response (4,5).
Upregulation of cytokine expression further stimulates the
keratinocytes and aggravates the inflammatory response of the skin
(4,6). A complex immune network formed by
keratinocytes and cytokines serves a role in psoriasis. Previous
studies demonstrated that psoriasis is initiated when keratinocytes
are subjected to a variety of stressors, including infection,
pregnancy and trauma (4,7,8).
These stressors result inactivation of the immune system, secretion
of a number of immune-associated cytokines and chemokines, and
induction of inflammatory responses (4). Cytokines are additionally produced by
activated T cells and dendritic cells (9). Therefore, the formation of psoriatic
lesions results from an interaction between the activation of skin
T cells and stimulation of keratinocytes (10).
Keratinocyte growth factor (KGF) is a member of the
fibroblast growth factor family, and is additionally termed
fibroblast growth factor-7 (11).
KGF is a paracrine growth factor secreted by various interstitial
cells, including fibroblasts, endothelial cells, smooth muscle
cells and dendritic epidermal T cells (11–13).
KGF interacts with a specific receptor, fibroblast growth factor
receptor 2 IIIb on epidermal cells (13). Although KGF is secreted by
interstitial cells, it targets epidermal cells. A previous study
demonstrated that the expression of KGF was markedly increased
following trauma in mice and humans (14). In addition, cytological studies
have demonstrated that KGF is able to specifically promote the
proliferation and migration of keratinocytes (14,15).
These studies suggested that KGF may serve a role in wound healing
and in promoting the proliferation of keratinocytes (15).
White peony is the dry root of plants in the
Ranunculaceae (crowfoot family), and its primary therapeutic
ingredient is a group of substances collectively termed total
glucosides of peony (TGP), which include peony bitter, hydroxyl
paeoniflorin, benzoyl and paeoniflorin, which accounts for >90%
of all the ingredients (16,17).
Clinical application and pharmacological studies have demonstrated
that TGP exhibit anti-inflammatory, analgesic and immune regulatory
effects, protect the liver and vascular endothelium against
inflammation (17,18). TGP is used for the treatment of a
variety of autoimmune diseases, including psoriasis (19), systemic lupus erythematosus
(19,20) and ankylosing spondylitis (21). A number of studies have suggested
that TGP is involved in numerous aspects of autoimmune processes
(1). The expression levels of
interleukin (IL)-2 and −4 are reduced and elevated, respectively,
in dinitrochlorobenzene-induced chronic dermatitis-eczema models
(22). TGP are involved in the G
protein-adenylate cyclase-cyclic adenosine monophosphate (cAMP)
signal transduction pathway (23).
It has additionally been demonstrated that TGP may downregulate the
expression of inflammatory cytokines, including tumor necrosis
factor-α (TNF-α), IL-6, IL-8 and interferon (IFN)-α (24).
A previous study reported that KGF may regulate the
expression of the proinflammatory mediators TNF-α and IL-1 to
stimulate inflammatory responses via activation of p38
mitogen-activated protein kinase (p38 MAPK) (25). The therapeutic effects of TGP may
be associated with the regulation of p38 MAPK. Hyperproliferative
HaCaT cells exhibit characteristics of keratinocytes in psoriatic
lesions, and are therefore used as in vitro models of
psoriasis in drug research (26).
Therefore, in order elucidate the underlying mechanism of action of
TGP in the treatment of psoriasis, the effects of TGP on the
proliferation of HaCaT cells induced by KGF were determined. The
p38 MAPK signaling pathway was selected to elucidate the molecular
mechanism of TGP.
Materials and methods
Cell culture
The immortal human keratinocyte line HaCaT was
purchased from the American Type Culture Collection (ATCC;
Manassas, VA, USA). The HaCaT cells were cultured in Dulbecco's
modified Eagle medium (DMEM; Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany) supplemented with 5% fetal bovine serum, 100 U/ml
penicillin and 100 µg/ml streptomycin, and were incubated in a cell
culture chamber at 37°C in a humidified atmosphere containing 5%
CO2. Cells were grown to 80% confluence, the culture
medium was removed and washed with PBS, and the cells were digested
using 0.25% trypsin. Cells in the logarithmic growth phase were
used for the subsequent experiments.
Cell experiments
Confluent cultures of HaCaT cells were incubated
with DMEM supplemented with 0, 1.25, 2.5, 5, 10, 20 and 40 ng/ml
KGF (Prospec-Tany TechnoGene, Ltd., East Brunswick, NJ, USA) for
24, 48 and 72 h at 37°C. The cell hyperproliferation model of
psoriasis was established in HaCaT cells via treatment with KGF (10
ng/ml) for 24 h at 37°C and confirmed using a Cell Counting Kit-8
(CCK-8) assay. To investigate the effect of TGP on
hyperproliferative HaCaT cells induced by KGF, cell proliferation
and the cell cycle were detected for these cells following
treatment with different concentrations of TGP: i) TGP-low (L; 10
mg/l); ii) TGP-medium (M; 50 mg/l); and iii) TGP-high (H; 120 mg/l)
or 48 h at 37°C. Subsequently, to determine the role of TGP, cells
were divided into five groups: i) Model, cells treated with 10
ng/ml KGF; ii) TGP, cells treated with 120 mg/lTGP; iii) control,
cells with no treatment; iv) KGF-IN [cells treated with 10 ng/ml
KGF+10 µm SB203580 (Calbiochem; Merck KGaA)]; and v) KGF-TGP (cells
treated with 10 ng/ml KGF+120 mg/lTGP). Following incubation for 48
h at 37°C, cells were harvested and used for subsequent
experimentation as described below.
Detection of cell proliferation
activity by CCK-8 assay
To determine alterations in cell proliferation,
HaCaT cells from all groups were seeded in 96-well plates at a
density of 1×105 cells/well. Cell proliferation was
detected using a CCK-8 kit (Dojindo Molecular Technologies, Inc.,
Kumamoto, Japan), according to the manufacturer's instructions. A
total of 20 µl CCK-8 was added and cells were incubated at 37°C for
4 h. The optical density values were measured at a wavelength of
450 nm using an ELISA reader (BioTek Instruments, Inc., Winooski,
VT, USA), according to the manufacturer's protocol.
Cell cycle detection by flow cytometry
assay
HaCaT cells collected from all groups were washed
with PBS and centrifuged at 500 × g for 5 min at 4°C. Cells were
adjusted to a density of 1×106 and re-suspended.
Propidium iodide (400 µl) and RNA-seA (100 µl; both from
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) were subsequently
added to the cells. Following staining for 30 min at room
temperature, the DNA content was determined using a FACSCalibur
system and analyzed using CellQuest software (version 3.3; BD
Biosciences, Franklin Lakes, NJ, USA).
Western blot analysis
HaCaT cell lysates from all groups were extracted
using lysis Triton X-100 buffer (250 mM HEPES, 50 mM
MgCl2, 10 mM EGTA and 5% Triton X-100). Protein
concentration was determined using a bicinchoninic acid protein
assay kit (Pierce; Thermo Fisher Scientific, Inc., Waltham, MA,
USA). Proteins (10 µg) were resolved on 10% SDS-PAGE gels and
transferred onto polyvinylidene fluoride membranes. Membranes were
blocked with 5% bovine serum albumin (BSA; Sigma-Aldrich; Merck
KGaA) in TBS with 0.1% Tween-20 for 1 h at room temperature, and
incubated with anti-β-actin (ab8226; 1:2,000), anti-IL-22,
anti-VEGF (ab9570; 1:1,000), anti-p38 MAPK (ab31828; 1:1,000),
anti-phosphorylated (p)-p38 MAPK (ab4822; 1:1,000) and anti-NF-κB
p65 (ab16502; 1:1,000) (all from Abcam) antibodies at 4°C
overnight. β-actin was used as a loading control. Following washing
with PBS, blots were incubated with horseradish
peroxidase-conjugated secondary antibodies (Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) at a concentration of
1:2,000–1:5,000 at room temperature for 1 h. Blots were visualized
using chemiluminescence (GE Healthcare Life Sciences, Little
Chalfont, UK) and developed on film (Kodak, Rochester, NY, USA),
according to the manufacturer's instructions. ImageJ software
version 1.42 (National Institutes of Health, Bethesda, MD, USA) was
used for densitometry analysis.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) assay
Total RNA was isolated using an RNeasy mini kit
(Qiagen, Inc., Valencia, CA, USA), according to the manufacturer's
instructions. A total of 1 µg total RNA was reverse transcribed to
cDNA using a PrimeScriptH RT reagent kit (Takara Biotechnology Co.,
Ltd., Dalian, China). Reverse transcription reaction conditions
were: 30°C for 10 min, 42°C for 30 min, 99°C 5 min and 4°C for 5
min. qPCR was performed using SYBR Green PCR master mix (Applied
Biosystems; Thermo Fisher Scientific, Inc.) with a SYBR Green
detection system [FS Universal SYBR Green Master (Rox); Roche
Applied Science, Penzberg, Germany]. Thermocycling procedures
consisted of 50°C for 2 min, 95°C for 10 min and 40 cycles of
amplification reactions at 95°C for 15 sec, and at 60°C for 1 min.
All samples were analyzed in triplicate and assessed by melting
curve analysis, and the PCR products obtained were analyzed by 1%
(w/v) agarose gel electrophoresis. The expression levels of IL-22
and VEGF were normalized to β-actin and calculated using the
2−ΔΔCq method (27).
The following primer sequences were used: IL-22,
5′-TGAGTGAGCGCTGCTATCTG-3′ (forward), 5′-TGTGCTTAGCCTGTTGCTGA-3′
(reverse); VEGF, 5′-CTGTCTAATGCCCTGGAGCC-3′ (forward),
5′-ACGCGAGTCTGTGTTTTTGC-3′ (reverse); β-actin,
5′-GCAGGAGTATGACGAGTCCG-3′ (forward), 5′-AACAACGCATCTCATATTTGGAA-3′
(reverse).
Statistical analysis
Data are presented as the mean ± standard deviation.
All samples were analyzed in triplicate. Statistical comparisons
between different groups were performed using SPSS software
(version 20; IBM Corp., Armonk, NY, USA). One-way analysis of
variance followed by Bonferroni post hoc pairwise comparison was
used for the evaluation of differences between groups. P<0.05
was considered to indicate a statistically significant
difference.
Results
HaCaT cell proliferation under
different concentrations of KGF
To investigate the response of HaCaT cells to KGF,
proliferation activity was determined in the presence of different
concentrations of KGF. Fig. 1
presents the alterations in cell morphology following treatment
with 0–40 ng/ml KGF for 24 h. Cell densities markedly increased in
cells treated with 0–10 ng/ml KGF, and marginally increased in the
presence of KGF at 10–40 ng/ml. Cells treated with 5–20 ng/ml KGF
appeared more elongated compared with cells treated with 0–2.5
ng/ml KGF. The viability of HaCaT cells in the presence of KGF was
determined at different time-points (24, 48 and 72 h) and
concentrations (0, 1.25, 2.5, 5, 10, 20 and 40 ng/ml). Cell
viability was notably elevated following treatment with KGF in a
time- and dose- dependent manner. Cell viability significantly
increased in cells treated with a dose of >10 ng/ml of KGF,
compared with the control.
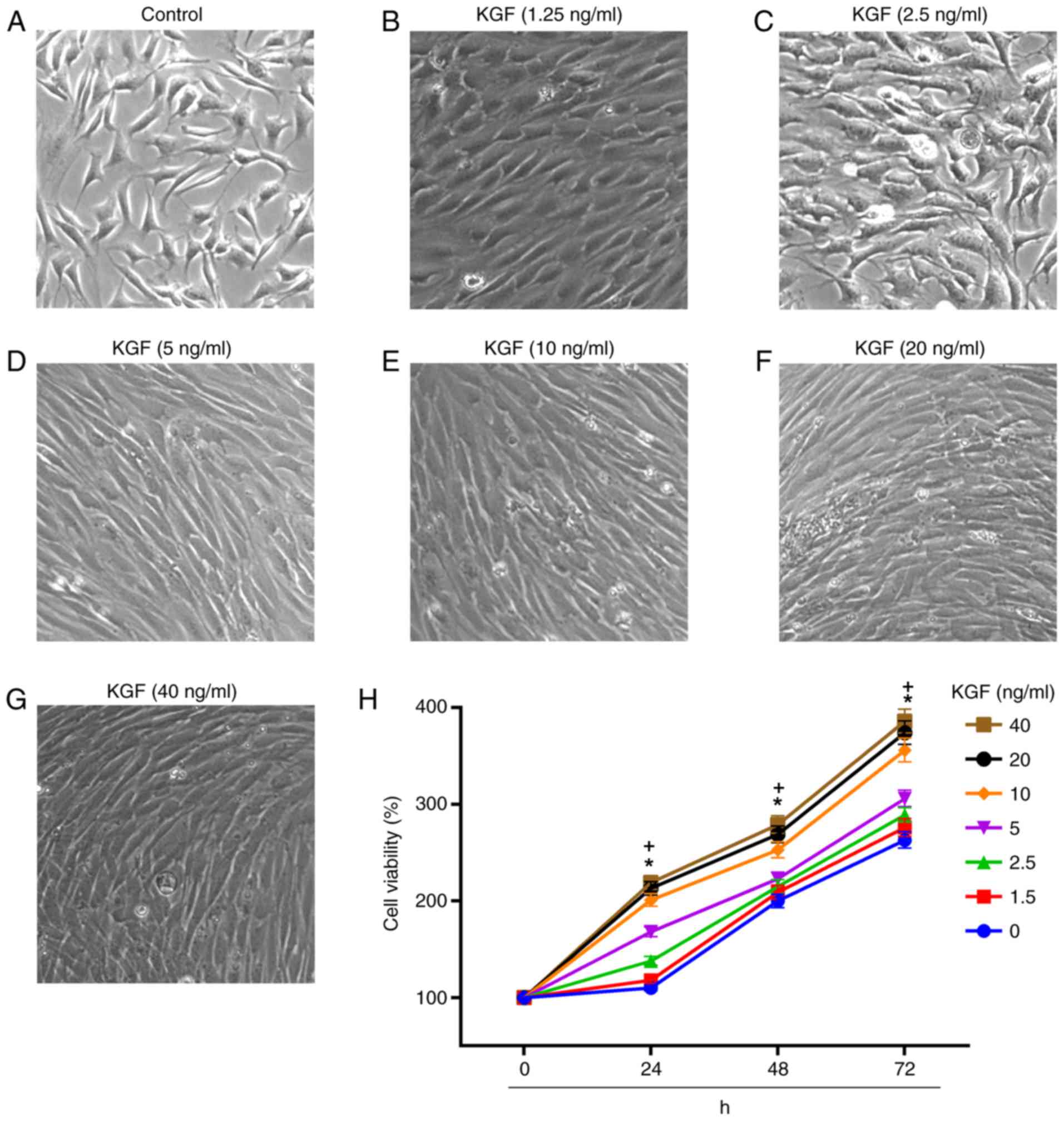 | Figure 1.Cell morphology and viability of
HaCaT cells following treatment with different concentrations of
KGF for 24, 48 and 72 h. Morphology of (A) control cells and cells
treated with (B) 1.25, (C) 2.5, (D) 5, (E) 10, (F) 20, and (G) 40
ng/ml KGF. Cell morphology alterations included cell elongation
following treatment with KGF (magnification, ×400). (H) The Cell
Counting Kit-8 assay demonstrated that the cell viability was
markedly elevated in cells following exposure to KGF. KGF,
keratinocyte growth factor. *P<0.05, >5 ng/ml KGF dosage
group vs. control (0 ng/ml KGF); +P<0.05, 24, 48 and
72 h vs. 0 h. |
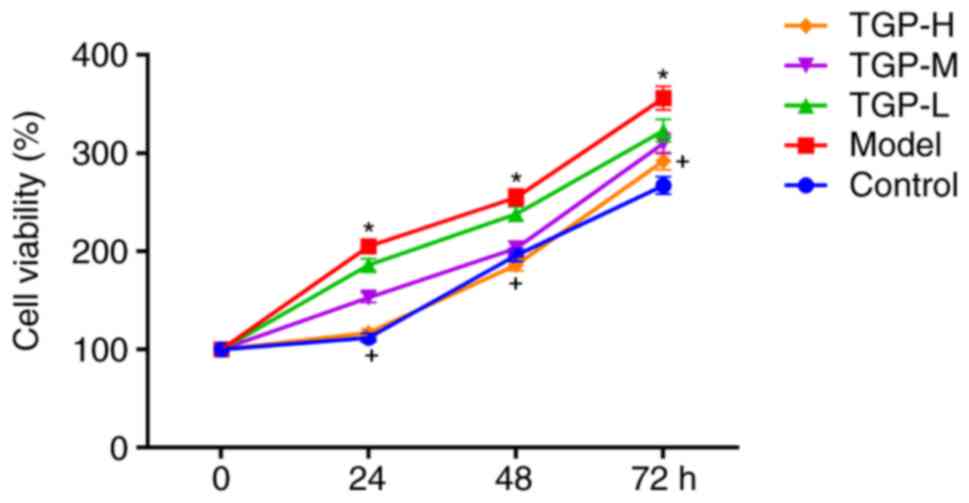
Effect of TGP treatment on cell
hyperproliferation triggered by KGF
One of the primary pathological characteristics of
psoriasis is overgrowth and abnormal differentiation of epidermal
keratinocytes (4,6). The present study investigated the
inhibitory effect of treatment with TGP on the hyperproliferation
of HaCaT cells. The cell viability of the model group treated with
10 ng/ml KGF (detected using a CCK-8 assay) significantly increased
compared with the control group. However, additional treatment with
TGP can reduce the excessive cell proliferation caused by KGF
(Fig. 2). Cell hyperproliferation
triggered by KGF was markedly inhibited in the TGP-H group,
compared with the model group. The effect of TGP treatment on cell
hyperproliferation induced by KGF was additionally validated in
HaCaT cells obtained from CLS Cell Lines Service GmbH (Eppelheim,
Germany; data not shown).
Treatment with TGP suppresses the
hyperproliferation of HaCaT cells stimulated by KGF by triggering
cell cycle arrest at the G1 phase
To determine the mechanism underlying TGP-mediated
inhibition of KGF-induced hyperproliferation, the cell cycle was
investigated by flow cytometry. The effect of different
concentrations of TGP on cells exposed to KGF was investigated
(Fig. 3). The results demonstrated
that among cells treated with KGF, fewer cells were in the
G0/G1-phase and an increased percentage was in the S-phase,
compared with the control group, indicating that KGF treatment
stimulated the proliferation of HaCaT cells (Fig. 3A, B and F). However, the
KGF-induced proliferation of HaCaT cells was inhibited by
increasing concentrations of TGP. Among cells treated with TGP, an
increased number of G0/G1-phase cells and a lower number of S-phase
cells was determined, compared with the model group, suggesting
that treatment with TGP suppressed the hyperproliferation of HaCaT
cells stimulated by KGF by inducing G1 arrest (Fig. 3B-F).
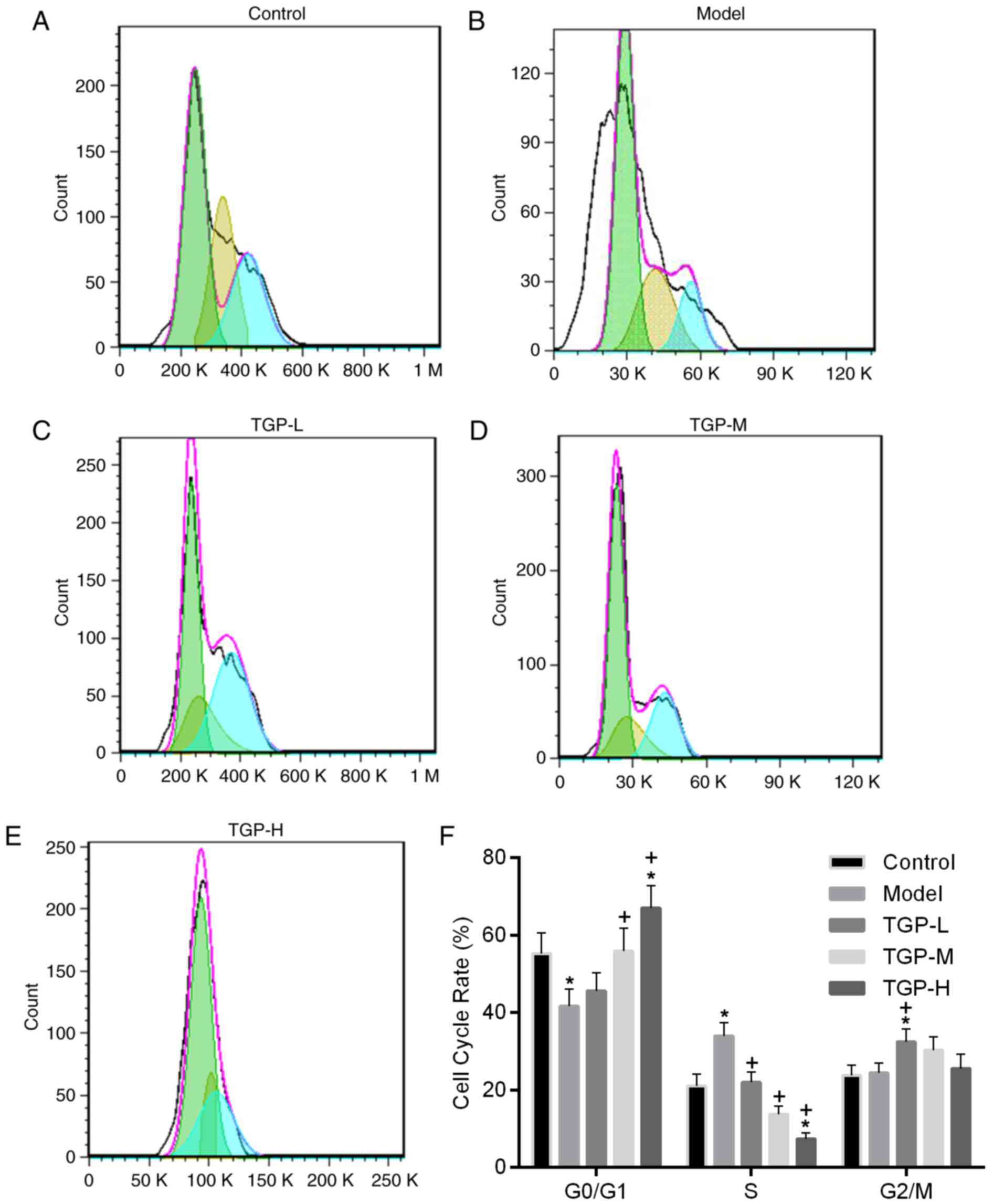 | Figure 3.Treatment with TGP demonstrated the
apparent suppression of KGF-induced hyperproliferation in HaCaT
cells. Flow cytometry assays were performed to assess the effect of
TGP on the hyperproliferation of HaCaT cells induced by KGF. Flow
cytometry results for (A) the control, (B) model, (C) TGP-L, (D)
TGP-M and (E) TGP-H groups (green, G0/G1, yellow, S; blue, G2/M).
(F) Groups treated with TGP demonstrated an increased percentage of
G0/G1-phase cells and a decreased number of cells in the S-phase
compared with the model group. TGP, total glucosides of peony; L,
low; M, medium; H, high; KGF, keratinocyte growth factor.
*P<0.05 vs. the control group; +P<0.05 vs. the
model group. |
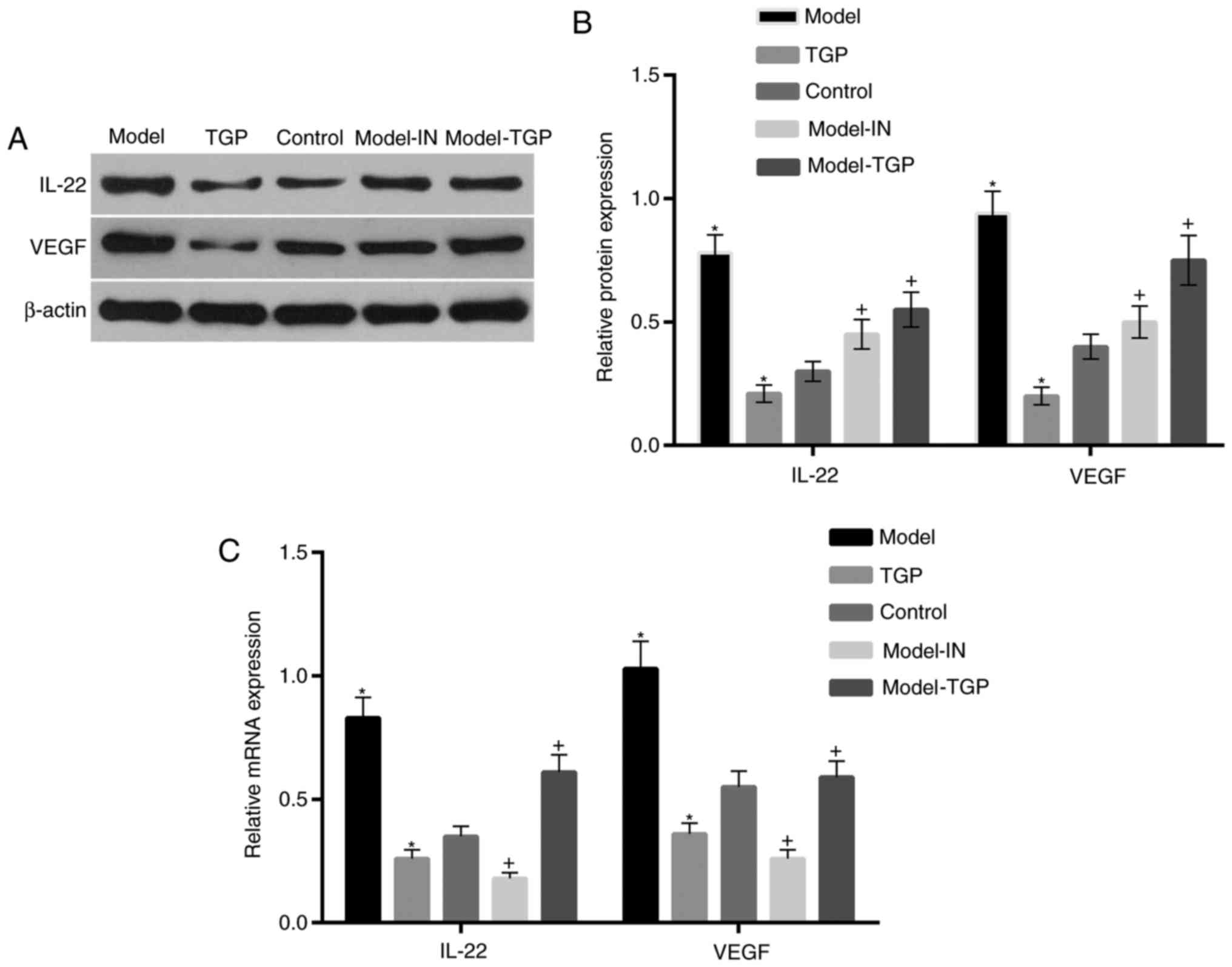
Expression of inflammatory cytokines
induced by KGF is suppressed by treatment with TGP
A previous study indicated that KGF can simulate the
expression of cytokines to promote cell proliferation via
activation of p38 MAPK (25). To
confirm this function of KGF, the p38 MAPK inhibitor SB203580 was
used to treat HaCaT cells. In addition, the effect of treatment
with TGP on HaCaT cells was assessed. The results revealed that the
expression levels of the inflammatory cytokines IL-22 and VEGF in
HaCaT cells were markedly elevated in the model group, compared
with the control (Fig. 4). The
expression levels of these cytokines decreased following treatment
with KGF and SB203580, and following treatment with TGP, compared
with the control. Furthermore, the expression levels of IL-22 and
VEGF were suppressed in cells treated with KGF and TGP, compared
with the model group.
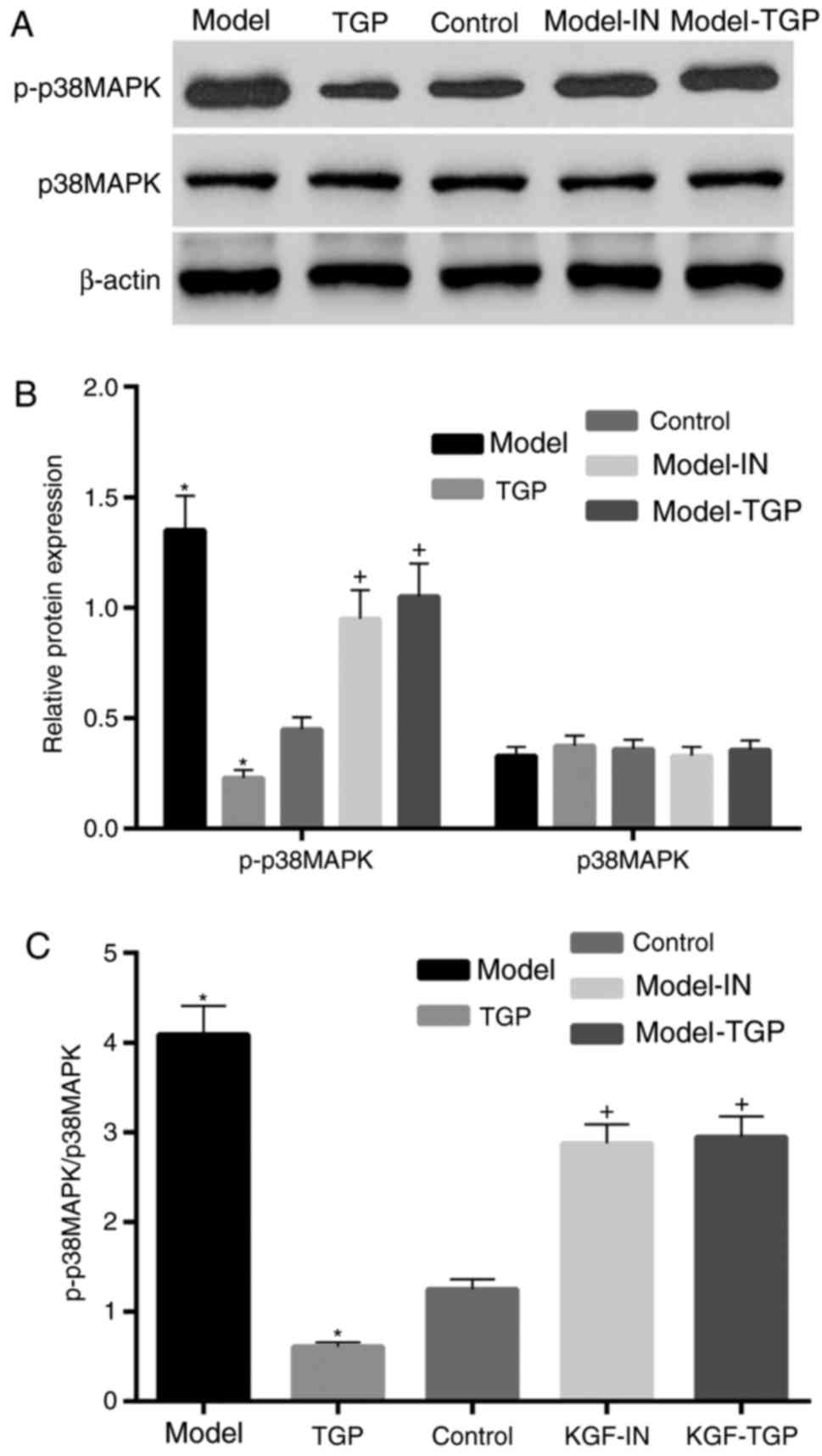
Activation of p38 MAPK is inhibited in
cells treated with TGP
Based on the results of a previous study (25), the authors of the present study
hypothesized that TGP may induce a similar inhibitory effect to
SB203580 on p38 MAPK. Therefore, the expression of p38 MAPK and
p-p38 MAPK was determined by western blot analysis. The expression
level of p-p38 MAPK was significantly elevated following treatment
with KGF compared with the control. By contrast, following
treatment with TGP, the expression level of p-p38 MAPK was markedly
inhibited compared with the control (Fig. 5). Furthermore, compared with the
model group, the expression level of p-p38 MAPK was suppressed by
combination treatment with KGF and SB203580, or KGF and TGP
(Fig. 5B). However, the expression
level of p38 MAPK was approximately equal in all groups. The ratio
of p-p38 MAPK to p38 MAPK exhibited a similar pattern to the levels
of p-p38 MAPK, since the expression of p38 MAPK was similar in all
groups. The inhibitory effect of TGP on p38 MAPK activation was
additionally validated in HaCaT cells obtained from CLS Cell Lines
Service GmbH (data not shown).
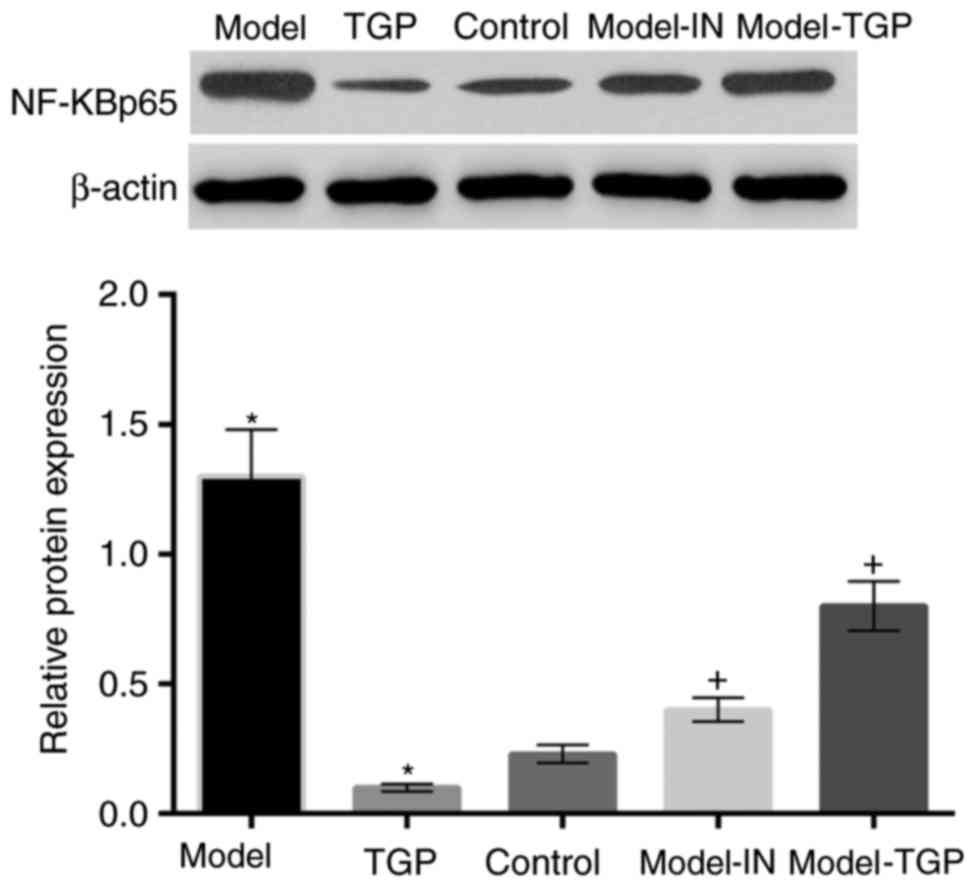
Activation of NF-κB p65 in cells is
suppressed by treatment with TGP
Expression of inflammatory cytokinesis regulated by
p38 MAPK/NF-κB p65 pathway. Therefore, the present study further
determined the expression levels of NF-κB p65 (Fig. 6). The NF-κB p65 protein expression
level in cells treated with KGF was upregulated compared with the
control. By contrast, treatment with TGP resulted in a
downregulation of the expression levels of NF-κB p65. NF-κB p65
protein expression levels were downregulated in cells with
combination treatment using KGF and SB203580 or KGF and TGP
compared with the model group. The above results were additionally
validated in HaCaT cells obtained from CLS Cell Lines Service GmbH
(data not shown).
Discussion
The present study aimed to determine whether p38
MAPK may be involved in the therapeutic effect mediated by TGP in
an in vitro model of psoriasis. Consistent with previous
studies (24–26), the results of the present study
demonstrated that KGF is associated with the activation of p38
MAPK, which was confirmed by the use of the p38 MAPK inhibitor
SB203580. Similar to SB203580, TGP exhibited an apparent inhibitory
effect on the activation of p38 MAPK. Furthermore, the expression
of IL-22 and VEGF protein and mRNA was suppressed by treatment with
TGP, which further supported the hypothesis that TGP may induce a
similar inhibitory effect to SB203580 on p38 MAPK.
TGP have been used to efficiently treat psoriasis
(19). Even though the mechanism
underlying the therapeutic effect of TGP on psoriasis has been
previously investigated, and it has been demonstrated to involve
the G protein-adenylate cyclase-cAMP signal transduction pathway,
it remains to be completely elucidated (22,23).
In addition, in a previous study, the proliferation of epidermal
cells was affected by KGF, epidermal growth factor, nerve growth
factor, TGF, TNF-α and other cytokines (28). However, KGF is the most potent and
specific cytokine that promotes keratinocyte proliferation
(14,15,29).
Therefore, the present study aimed to determine the effect of TGP
on the KGF-induced hyperproliferation of HaCaT cells. Consistent
with other studies, in the present study, KGF enhanced the
proliferation of HaCaT cells. However, this effect of KGF was
suppressed following treatment with TGP. Cell cycle analysis
demonstrated that among cells treated with KGF and TGP, an elevated
number of cells was in the G0/G1-phase and a decreased number of
cells was in the S-phase, compared with cells treated with KGF
alone. The above results indicated that TGP may suppress the
hyperproliferation of HaCaT cells stimulated by KGF, by triggering
G1-phase arrest.
It has been hypothesized that the abnormal
infiltration of T lymphocytes (primarily CD4+ T
lymphocytes) serves a role in the pathogenesis of psoriasis
(4,10). Elevated levels of T helper
(Th)1-type lymphocytes is the predominant response observed in
patients with pancreatic diseases, and Th1 type cytokines,
including TNF-α, interferon-γ and IL-2, contribute to the
aggravation of psoriasis (30).
Th2 type cytokines, including IL-4 and −10, serve a protective role
instead (31). Previous studies
suggested that psoriasis is a disease based on Th17/Th1 cell
responses and hypothesized that these induced
Th-17+/IFN-γ+ cells may be the cause of
psoriasis (32,33). However, psoriasis is a multi-gene
genetic disease associated with a number of factors, including
trauma, infection, and the underlying mechanisms of psoriasis
remain to be completely elucidated (2,34). A
number of previous studies demonstrated that the interaction
between IL-23, −17 and −22, and other cytokines, serves a role in
the development of psoriasis (35–37).
Elevated mRNA and protein expression levels of IL-21, −22, −17 and
−23 were detected in skin tissue with psoriatic lesions, and IL-23
may induce lesions similar to human psoriasis in mouse skin,
including epidermal hyperplasia erythema, leukocyte infiltration
and keratosis (35,36). Furthermore, inflammation and
acanthosis of the dermal layer have been hypothesized to be
associated with cytokine IL-22 and C-C chemokine receptor type 6
(38). The present study
demonstrated that the mRNA and protein expression levels of IL-22
were upregulated in cells treated with KGF, whereas they were
inhibited in model cells following treatment with TGP or the p38
MAPK inhibitor SB203580.
During the development of psoriasis, numerous
vascular-derived cytokines secreted by epidermal cells are involved
in angiogenesis, including VEGF (39). In psoriatic lesions, keratinocytes
are the primary source of angiogenic cytokines (4,6).
Previous studies demonstrated that the mRNA and protein expression
levels of VEGF in patients with psoriatic lesions were markedly
increased (40,41). Furthermore, it was demonstrated
that serum levels of VEGF in patients with severe psoriasis were
elevated and served a role in the progression of psoriasis
(42). The results of the present
study demonstrated that the protein and mRNA expression levels of
VEGF were increased in cells treated with KGF. Similar to the
levels of IL-22, the expression of VEGF protein and mRNA was
suppressed following treatment with TGP. Consistent with these
results, treatment of model cells with the p38 MAPK inhibitor
SB203580 or TGP additionally inhibited the expression of VEGF.
A previous study demonstrated that KGF simulates p38
MAPK to trigger stress fiber formation in human prostate DU145
cells (43). In addition, a study
revealed that the production of VEGF was upregulated following
treatment with KGF, and fibroblast growth factor-1 and −2 via the
MAPK and p38 MAPK pathways (44).
Furthermore, the results of the present study revealed that the
expression levels of VEGF protein and mRNA were elevated in cells
treated with KGF, while these levels were suppressed by combined
treatment with KGF and p38 MAPK inhibitor SB203580, indicating that
the expression of VEGF may be associated with the p38 MAPK
signaling pathway. Therefore, it may be hypothesized that KGF
induces p38 MAPK activation to promote the upregulation of VEGF,
and the results of the present study are consistent with this
hypothesis. The results of the present study additionally
demonstrated that the levels of p-p38 MAPK were associated with the
expression levels of IL-22 and VEGF. In addition, similar to the
p38 MAPK inhibitor SB20358, TGP inhibited p-p38 MAPK expression
levels. The above results suggested that TGP were able to
downregulate the expression of proinflammatory factors, including
IL-22 and VEGF, by inhibiting the activation of p38 MAPK.
Subsequently, alterations in the p38 MAPK/NF-κB p65 signaling
pathway were determined. The results indicated that the expression
level of NF-κB p65 was markedly elevated and reduced following
treatment with KGF and TGP, respectively. Furthermore, subsequent
experiments revealed that the expression levels of NF-κB p65 were
decreased following treatment of model cells with the p38 MAPK
inhibitor SB203580 or TGP, compared with the model group. The above
results suggested that the therapeutic effect of TGP on psoriasis
may be mediated by the regulation of the p38 MAPK/NF-κB p65
signaling pathway.
In conclusion, TGP may suppress the KGF-induced
hyperproliferation of HaCaT cells, which may be involved in the
downregulation of the expression of proinflammatory factors,
including IL-22 and VEGF. The results of the present study
indicated that the p38 MAPK/NF-κB p65 pathway may be associated
with psoriasis, and may be effectively suppressed by treatment with
TGP.
Acknowledgements
Not applicable.
Funding
Not applicable.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
WP and CC contributed equally to this study and
analyzed and interpreted data relating to the study. XQ and SZ
designed the study, implemented experiments and wrote the paper.
All authors read and approved the final study.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
All authors declare that they have no conflict of
interest.
References
|
1
|
Bagel J: Topical therapies for the
treatment of plaque psoriasis. Cutis. 84 4 Suppl:S3–S13. 2009.
|
|
2
|
Dedhar S, Hannigan G, Hunt DWC, Tao JS and
Fazli L: Treatment of inflammatory diseases including psoriasis. US
Patent 37863429 B2. Filed November 30, 2001; issued January 4.
2011.
|
|
3
|
Xia JX, Mei XL, Zhu WJ, Li X, Jin XH, Mou
Y, Yu K, Wang YY and Li QF: Effect of FGF10 monoclonal antibody on
psoriasis-like model in guinea pigs. Int J Clin Exp Pathol.
7:2219–2228. 2014.PubMed/NCBI
|
|
4
|
Feliciani C, Gupta AK and Sauder DN:
Keratinocytes and cytokine/growth factors. Crit Rev Oral Biol Med.
7:300–318. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zeng X, Zhao J, Wu X, Shi H, Liu W, Cui B,
Yang L, Ding X and Song P: PageRank analysis reveals topologically
expressed genes correspond to psoriasis and their functions are
associated with apoptosis resistance. Mol Med Rep. 13:3969–3976.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Carter EL, O'Herrin S, Woolery C and
Longley Jack B: Epidermal stem cell factor augments the
inflammatory response in irritant and allergic contact dermatitis.
J Inv Dermatol. 128:1861–1863. 2008. View Article : Google Scholar
|
|
7
|
Cather JC, Latremouille-Viau D, Horn EJ
and Bao Y: Psoriasis is significantly associated with lower rates
of pregnancy and live births. J Am Acad Dermatol. 66:AB2002012.
|
|
8
|
Thomas J, Jacobson GA, Narkowicz CK,
Peterson GM, Burnet H and Sharpe C: Toenail onychomycosis: An
important global disease burden. J Clin Pharm Ther. 35:497–519.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kubo T, Hatton RD, Oliver J, Liu X, Elson
CO and Weaver CT: Regulatory T cell suppression and anergy are
differentially regulated by proinflammatory cytokines produced by
TLR-activated dendritic cells. J Immunol. 173:7249–7258. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
de Boer OJ, van der Loos CM, Hamerlinck F,
Bos JD and Das PK: Reappraisal of in situ immunophenotypic analysis
of psoriasis skin: Interaction of activated HLA-DR+ immunocompetent
cells and endothelial cells is a major feature of psoriatic
lesions. Arch Dermatol Res. 286:87–96. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Strain AJ, Mcguinness G, Rubin JS and
Aaronson SA: Keratinocyte growth factor and fibroblast growth
factor action on DNA synthesis in rat and human hepatocytes:
Modulation by heparin. Exp Cell Res. 210:253–259. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yamamoto T, Matsuda Y, Kawahara K, Naito Z
and Ishiwata T: Keratinocyte growth factor stimulates growth of MIA
PaCa-2 cells through extracellular signal-regulated kinase
phosphorylation. Oncol Lett. 3:307–310. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Higashiyama S, Abraham JA, Miller J,
Fiddes JC and Klagsbrun M: A heparin-binding growth factor secreted
by macrophage-like cells that is related to EGF. Science.
251:936–939. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li M: KGF-1 and KGF receptor expression in
human periodontal disease and in vitro
microwounding-associated-ligand-independent KGFR activation.
2007.http://hdl.handle.net/2429/418
|
|
15
|
Putnins EE, Firth JD, Lohachitranont A,
Uitto VJ and Larjava H: Keratinocyte growth factor (KGF) promotes
keratinocyte cell attachment and migration on collagen and
fibronectin. Cell Adhes Commun. 7:211–221. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang K, Wu Y-G, Su J, Zhang JJ, Zhang P
and Qi XM: Total glucosides of paeony regulates JAK2/STAT3
activation and macrophage proliferation in diabetic rat kidneys. Am
J Chin Med. 40:521–536. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Harada M: Pharmacological studies on herb
paeony root. IV. Analysis of therapeutic effects of paeony- and
licorice-containing frequent prescriptions in Chinese medicine and
comparison with effects of experimental pharmacological tests.
Yakugaku Zasshi. 89:899–908. 1969.(In Japanese). View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang ZG: Antianxiety and sleep disorder
improving use of albiflorin or metabolite thereof in. Patent EP
2644198 A1. Filed November 9, 2011; issued October 2. 2013.
|
|
19
|
Li Y, Cheng H, Zuo XB, Sheng YJ, Zhou FS,
Tang XF, Tang HY, Gao JP, Zhang Z, He SM, et al: Association
analyses identifying two common susceptibility loci shared by
psoriasis and systemic lupus erythematosus in the Chinese Han
population. J Med Genet. 50:812–818. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang HF, Xiao WG and Hou P: Clinical
study of total glucosides of paeony in patients with systemic lupus
erythematosus. Zhongguo Zhong Xi Yi Jie He Za Zhi. 31:476–479.
2011.(In Chinese). PubMed/NCBI
|
|
21
|
Wang SL, Wang JP and Bian H: Clinical
observation on total glucosides of paeony combined with
sulfasalazine in treatment of ankylosing spondylitis. Zhongguo
Zhong XI Yi Jie He Za Zhi. 27:217–219. 2007.(In Chinese).
PubMed/NCBI
|
|
22
|
Chang Y, Wei W, Zhang L and Xu HM: Effects
and mechanisms of total glucosides of paeony on synoviocytes
activities in rat collagen-induced arthritis. J Ethnopharmacol.
121:43–48. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jia XY, Chang Y, Sun XJ, Wu HX, Wang C, Xu
HM, Zhang L, Zhang LL, Zheng YQ, Song LH and Wei W: Total
glucosides of paeony inhibit the proliferation of fibroblast-like
synoviocytes through the regulation of G proteins in rats with
collagen-induced arthritis. Int Immunopharmacol. 18:1–6. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhou Y, Li J, Kong F, Zhang H, Fang X,
Chen Z, Wang G and Li X and Li X: Clinical and immunological
consequences of total glucosides of paeony treatment in Sjögren's
syndrome: A randomized controlled pilot trial. Int Immunopharmacol.
39:314–319. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Westra J, Harmsen MC, van Rijswijk MH and
Limburg PC: Effect of P38 mapkinase inhibitor RWJ-67657 on
proinflammatory mediators produced by IL-1β-and/or TNFα-stimulated
rheumatoid synovial fibroblasts. Arthritis Res. 4 Suppl 1:S452002.
View Article : Google Scholar
|
|
26
|
Micallef L, Belaubre F, Pinon A,
Jayat-Vignoles C, Delage C, Charveron M and Simon A: Effects of
extracellular calcium on the growth-differentiation switch in
immortalized keratinocyte HaCaT cells compared with normal human
keratinocytes. Exp Dermatol. 18:143–151. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang B, Amerio P and Sauder DN: Role of
cytokines in epidermal Langerhans cell migration. J Leukoc Biol.
66:33–39. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yang K, Yin J, Sheng B, Wang Q, Han B, Pu
A, Yu M, Sun L, Xiao W and Yang H: AhR-E2F1-KGFR signaling is
involved in KGF-induced intestinal epithelial cell proliferation.
Mol Med Rep. 15:3019–3026. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nestle FO, Turka LA and Nickoloff BJ:
Characterization of dermal dendritic cells in psoriasis.
Autostimulation of T lymphocytes and induction of Th1 type
cytokines. J Clin Invest. 94:202–209. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
de Jong R, Bezemer AC, Zomerdijk TP, Van
dp KT, Ottenhoff TH and Nibbering PH: Selective stimulation of T
helper 2 cytokine responses by the anti-psoriasis agent
monomethylfumarate. Eur J Immunol. 26:2067–2074. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zaba LC, Fuentesduculan J, Eungdamrong NJ,
Abello MV, Novitskaya I, Pierson KC, Gonzalez J, Krueger JG and
Lowes MA: Psoriasis is characterized by accumulation of
immunostimulatory and Th1/Th17 cell-polarizing myeloid dendritic
cells. J Invest Dermatol. 129:79–88. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kanai Y, Satoh T, Igawa K and Yokozeki H:
Impaired expression of Tim-3 on Th17 and Th1 cells in psoriasis.
Acta Derm Venereol. 92:367–371. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Harden JL, Krueger JG and Bowcock AM: The
immunogenetics of psoriasis: A comprehensive review. J Autoimmun.
64:66–73. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Works MG, Yin F, Yin CC, Yiu Y, Shew K,
Tran TT, Dunlap N, Lam J, Mitchell T, Reader J, et al: Inhibition
of TYK2 and JAK1 ameliorates imiquimod-induced psoriasis-like
dermatitis by inhibiting IL-22 and the IL-23/IL-17 axis. J Immunol.
193:3278–3287. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Raychaudhuri SP: Role of IL-17 in
psoriasis and psoriatic arthritis. Clin Rev Allergy Immunol.
44:183–193. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Qin S, Wen J, Bai XC, Chen TY, Zheng RC,
Zhou GB, Ma J, Feng JY, Zhong BL and Li YM: Endogenous n-3
polyunsaturated fatty acids protect against imiquimod-induced
psoriasis-like inflammation via the IL-17/IL-23 axis. Mol Med Rep.
9:2097–2104. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zheng Y, Danilenko DM, Valdez P, Kasman I,
Eastham-Anderson J, Wu J and Ouyang W: Interleukin-22, a T(H)17
cytokine, mediates IL-23-induced dermal inflammation and
acanthosis. Nature. 445:648–651. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Shibuya M: Involvement of the
VEGF-Flt-receptor family in angiogenesis. Nihon Yakurigaku Zasshi.
107:119–131. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kwon YW, Kim KW, Jang HS, Kim MB, Oh CK
and Kwon KS: Insulin-like growth factor-II (IGF-II) induces
vascular endothelial growth factor (VEGF) expression through IGFR-1
and ERK1/2 activation in psoriasis. Korean J Dermatol. 42:728–734.
2004.
|
|
41
|
Verdes D, Popescu R, Gotia SR, Gotia L,
Filimon MN and Karabitsakos T: VEGF and VEGFRs expression in
psoriasis lesions. Curr Opinion Biotechnol. 22:S1142011. View Article : Google Scholar
|
|
42
|
Meki AR and Al-Shobaili H: Serum vascular
endothelial growth factor, transforming growth factor β1 and nitric
oxide levels in patients with psoriasis vulgaris: Their correlation
to disease severity. J Clin Lab Anal. 28:496–501. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Mehta PB, Robson CN, Neal DE and Leung HY:
Keratinocyte growth factor activates p38 MAPK to induce stress
fibre formation in human prostate DU145 cells. Oncogene.
20:5359–5365. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Aimond F, Rauzier JM, Bony C and Vassort
G: Simultaneous activation of p38 MAPK and p42/44 MAPK by ATP
stimulates the K+ current ITREK in cardiomyocytes. J Biol Chem.
275:39110–39116. 2000. View Article : Google Scholar : PubMed/NCBI
|